Back to Journals » Clinical Ophthalmology » Volume 9
Current perspective of neuroprotection and glaucoma
Authors Tian K, Shibata-Germanos S, Pahlitzsch M, Cordeiro MF
Received 3 August 2015
Accepted for publication 26 September 2015
Published 11 November 2015 Volume 2015:9 Pages 2109—2118
DOI https://doi.org/10.2147/OPTH.S80445
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Scott Fraser
Kailin Tian,1,2 Shannon Shibata-Germanos,1 Milena Pahlitzsch,1 M Francesca Cordeiro1,3
1Glaucoma and Retinal Neurodegeneration Research Group, UCL Institute of Ophthalmology, London, UK; 2Eye Centre, Renmin Hospital of Wuhan University, Wuhan, People’s Republic of China; 3Western Eye Hospital, ICORG, Imperial College NHS Trust, London, UK
Abstract: Glaucoma is the second leading cause of blindness worldwide and is most notably characterized by progressive optic nerve atrophy and advancing loss of retinal ganglion cells (RGCs). The main concomitant factor is the elevated intraocular pressure (IOP). Existing treatments are focused generally on lowering IOP. However, both RGC loss and optic nerve atrophy can independently occur with IOP at normal levels. In recent years, there has been substantial progress in the development of neuroprotective therapies for glaucoma in order to restore vital visual function. The present review intends to offer a brief insight into conventional glaucoma treatments and discuss exciting current developments of mostly preclinical data in novel neuroprotective strategies for glaucoma that include recent advances in noninvasive diagnostics going beyond IOP maintenance for an enhanced global view. Such strategies now target RGC loss and optic nerve damage, opening a critical therapeutic window for preventative monitoring and treatment.
Keywords: glaucoma, retinal ganglion cell, neuroprotection
Introduction
Glaucoma is the second leading cause of blindness worldwide,1 with an estimated 8.4 million people suffering from blindness in 2010 alone, accounting for 21% of total blindness in the world today, and it is estimated that the number of those with glaucoma worldwide will rise to 79.6 million by the year 2020.2 Glaucoma is considered a progressive neurodegenerative eye disorder with varied symptoms most notably characterized in earlier stages by the degeneration and loss of retinal ganglion cells (RGCs) and their axons, leading to optic neuropathy and visual field loss. This occurs through the loss of function and death of RGC through initially the process of programmed cell death (apoptosis). In later stages of the disease, progression includes neuronal loss in the lateral geniculate nucleus and visual cortex.3 Elevated intraocular pressure (IOP) is considered to be one of the key factors in glaucoma; however, elevated IOP is typically followed by the appearance of structural damage at the optic nerve head, including neuroretinal rim thinning, cupping of the optic disc, and sectoral retinal nerve fiber layer (RNFL) thinning, indicating the pathological progression of the disease within the retina as a whole.4
The best known classification of glaucoma relies on the anatomy of the anterior chamber and the drainage pathway (open and narrow angles). It is usually categorized as primary or secondary, based on several combined pathologies including comorbid conditions such as infection, mechanical injury, or neovascularization that often affect a single eye alone. In addition, primary open-angle glaucoma (POAG) can present with high and low IOPs, with the latter termed “normal tension glaucoma” representing 40% of patients with POAG.5 Furthermore, there is increasing recognition that even significant lowering of IOP may be insufficient to prevent visual loss in some patients. Therefore, alternative treatments besides lowering IOP, especially via prevention of neuronal cell loss, are now being investigated with an emphasis on neuroprotection to slow down the progression of glaucoma. Most data cited are obtained from preclinical studies.
The present review will 1) give a brief overview of the current treatments aiming to maintain IOP, 2) discuss neuroprotective and neuroregenerative strategies targeting RGC loss and the prevention of optic nerve damage, and 3) introduce new advances in diagnostic technologies.
Current and existing medication for glaucoma
The therapeutic aim for glaucoma treatment remains the prevention of vision loss by predominantly decreasing the risk factors of the disease. At present, the only effective treatment is the lowering of IOP.6,7 Currently available treatment modalities for glaucoma focus on the lowering of IOP through various strategies often in tandem, including decreasing the production of aqueous humor and/or increasing the drainage through the trabecular meshwork (TM) and/or enhancing uveoscleral outflow. A significant reduction of IOP (20%–40%) can limit progressive loss in the visual field.8 To date, there are numerous drugs that control IOP, which are most commonly in the form of topically applied eyedrops – a convenient noninvasive method of administration. These topical drugs decrease the production of aqueous humor and facilitate drainage through the TM, increasing uveoscleral outlflow.9 There are mainly five categories: β-blockers, carbonic anhydrase inhibitors, prostaglandin analogs, sympathomimetic drugs, and parasympathomimetic drugs.7,9 In addition, some fixed combination therapies are also given to some patients for effective IOP control when they do not respond to one pure form of medication. To date, fixed combination therapies for glaucoma mainly include prostaglandin analogs/β-blockers, carbonic anhydrase inhibitors/β-blockers, and α2-adrenergic agonists/β-blockers and carbonic anhydrase inhibitors/α2-adrenergic agonists. All the therapies were approved by the US Food and Drug Administration (FDA) in 2013.9 It is also worth mentioning that as an exception, triple fixed combination of prostaglandin analogs/α2-adrenergic agonists/β-blockers is available in Mexico.9 However, it was found elucidated that the currently available fixed combination of dorzolamide/dimolol and brimonidine/timolol could reduce IOP more effectively with reduced ocular allergy than their component used separately as monotherapy.10
Neuroprotection therapies for glaucoma
Although there are clinically effective IOP reduction treatments, there has been a growing realization that alternative treatments are needed to also target the prevention of RGC loss as patients with glaucoma have been seen to continue to lose visual function even after successful IOP lowering.
Neuroprotection, by definition, is “the relative preservation of neuronal structure and/or function”.11 In addition to reduction of IOP, neuroprotective therapies have the potential to halt RGC loss through promoting cell survival12 and preventing further structural and functional damage of the optic nerve to preserve vision in those showing glaucomatous signs.4,13
Though the primary mechanisms of glaucoma have not yet been fully elucidated, several key processes have been identified, such as mechanical compression,14 ischemia,15 oxidative stress,16 neurotropic growth factor deprivation,17 intracellular calcium toxicity,18 activation of autoimmunity,19 and glutamate neurotoxicity.20 In recent years, researchers have investigated a number of compounds that can protect RGCs via targeting the aforementioned pathways.
Glutamate toxicity-blocking agents
Excitotoxicity in neurodegenerative diseases can occur by stimulation of N-methyl-d-aspartic acid receptors (NMDARs), the receptor for the amino acid transmitter glutamate.21 Glutamate is an essential excitatory neurotransmitter released by bipolar cells onto RGCs in the vertebrate retina22 and has been shown to induce excitotoxicity in glaucoma.23 By the stimulation of NMDARs, extensive levels of Ca2+ enter the cells, activating phospholipases, endonucleases, proteases, etc, thereby leading to the apoptotic and/or necrotic cell death in neurons.24 It was demonstrated that the glutamate concentration was elevated chronically in the inner eye both in patients with glaucoma and in glaucoma animal models.25,26 This then caused the overactivation of the glutamate receptor, leading to neuron damage.9 Thus, antiexcitotoxic agents such as NMDAR antagonists, glutamate release inhibitors, and calcium channel blockers could all potentially be employed to inhibit excitotoxicity with the intent of halting related neuronal cells loss.
“Memantine” is the best known antagonist of NMDARs and is approved by the FDA for the treatment of Alzheimer’s disease following a multinational clinical trial in 2009.27 However, it was not found to be effective in a clinical trial in Alzheimer’s disease (NCT00235716).28 In addition, despite some evidence of its efficacy preclinically,29 a clinical trial of Memantine against chronic glaucoma (NCT00168350) finished in 2010 with results yet unpublished.
Ca2+ channel blockers have been suggested to be neuroprotective in glaucoma.30 Topical 2% flunarizine was confirmed to significantly ameliorate ischemic retinal damage in rabbit models.31 Iganidipnie, nimodipine, and lomerizine were applied to cultured rat RGCs under hypoxic conditions, evidencing a significant increase in RGCs’ viability.30 Clinically, orally administered nilvadipine slightly slowed down visual field degeneration and maintained the optic disc rim of patients with normal low-tension IOP open-angle glaucoma in a 3-year study.32
Neurotrophic factors
Among other roles, neurotrophic factors (NTFs) are involved in the maintenance and enhancement of neuronal cell survival and contribute to their differentiation, growth, and regeneration. They are essential for neuronal development within the nervous system. In theory, a therapeutic approach could involve the direct supply of exogenous neurotrophins or through the upregulation of endogenous expression of neurotrophins.
Brain-derived neurotrophic factor (BDNF) is secreted into the synaptic cleft, binding to surface tropomyosin receptor kinases B, which are then transported retrogradely to neuronal cells to regulate their activities.33 BDNF was reported to rescue RGCs after optic nerve axotomy,34 with exogenously applied BDNF proving neuroprotective on RGCs in moderately chronic hypertensive rat models.35 Leucine rich repeat and lg domain containing 1 (LINGO-1) is specific to the central nervous system and functions as a negative regulator of axonal regeneration and neuronal survival. Fu et al demonstrated that combing LINGO-1 antagonist with BDNF could provide long-term protection for RGCs in a chronic ocular hypertension (OHT) models, solving the problem of the neuroprotective limitations of BDNF downregulating the tropomyosin receptor kinases B receptor.36
Ciliary neurotropic factor (CNTF),37,38 nerve growth factor (NGF),39 and glial cell-line-derived neurotropic factor40 have all been identified as neuroprotective factors for RGCs. Among them, CNTF, delivered by an encapsulated cell technology implant, has been used in a Phase II clinical trial against geographic atrophy and has been proved to slow the progression of vision loss.41 A Phase I clinical trial for CNTF implant in patients with POAG (NCT01408472) completed in October 2014 is yet to be published. A significant reduction in RCG loss has been demonstrated with topical NGF application in rat glaucoma models at 7 weeks. Its efficacy is directly associated with the inhibition of cell death by apoptosis.42 Lentiviral vector-mediated retinal gene transfer of pigment epithelium-derived factor was achieved through subretinal injection in both transient OHT and N-methyl-d-aspartic acid-induced models, indicating pigment epithelium-derived factor had led to neuroprotective effects rescuing RGCs.43
Brimonidine (BMD) is a selective α2-adrenergic receptor agonist that is used clinically to lower IOP by decreasing the production of aqueous humor and facilitating its outflow via the TM.44 Neuroprotective efficacy of BMD has been found in several animal experimental disease models, such as optic nerve injury45 and OHT models,46 with both convincing histological evidence and functional protection. BMD has been found to prevent progressive RGC loss and thinning of the inner retina layer and visual impairment in a normal tension glaucoma murine model. This was achieved through excitotoxic inhibition of upregulated phosphorylated NR2B, stimulation of production of NGF, BDNF, and basic fibroblast growth factor in multiple pathways.47 In recent years, there have been several ongoing clinical trials for BMD. In a 4-year double-masked, randomized, and multicenter clinical trial testing the efficacy of monotherapy with BMD tartrate 0.2% versus timolol maleate 0.5% eyedrops (NCT00317577), the BMD group had a significantly lower rate of visual field progression, attributed to an IOP–independent protective effect of BMD or conversely, a potentially harmful effect of timolol.48,49
To date, some identified limitations regarding neurotrophins include potential instability of eyedrops, the necessity of repeated intravitreal administration, the long-term or short lasting effects, and the mode of delivery (invasive vs noninvasive).50,51 Therefore, further novel approaches, such as gene therapy or stem cell therapy, need to be comprehensively investigated and developed to make neurotrophin application a more realistic therapy for glaucoma.
Oxidative stress suppression
Reactive oxygen species are chemically reactive molecules, including oxygen ions and peroxides, generated in the metabolism of oxygen and cleared by antioxidants. Their accumulation induces oxidative stress imbalance, which overburdens the cell’s ability to readily clear the intermediates and repair the subsequent damage. It could also lead to damage of cellular components, such as DNA, fatty acids in lipids, and amino acids in protein. During the past 2 decades, oxidative stress was shown to be involved in the pathogenesis of glaucoma,16,52 especially in POAG.53
Aminoguanidine, an inhibitor of NOS-2, is believed to reduce reactive oxygen species production and showed significant prevention of RGC loss in rat glaucoma models, offering a hint that this well-tolerated pharmacological agent could be a reasonable candidate for neuroprotective therapy in glaucoma.54 Vitamin E, a scavenger of peroxyl radicals,9 has been indicated as protective of RGCs. Its daily supplementation in patients was found to be associated in decreasing the rate of primary glaucomatous progression.55–57 Conversely, a higher rate of RGC death in vitamin E deficiency was linked to elevated lipid peroxidation.58 Recent studies showed that vitamin E-modified silicone-hydrogel contact lenses can also help to increase the sustained release of such drugs, making the effect of the therapy last longer than the wear duration.59,60
Ginkgo biloba extract is another compound of interest, mainly containing antioxidants such as flavonoids and cyanidine,16,61 which exert significant protective effects against free radical and lipid peroxidation.62 They can be used as a neuroprotective therapy for RGC survival against glaucoma63 though the underlying mechanism is not established.
Coenzyme Q10 (CoQ10), an important potent antioxidant, was previously proved to be an effective neuroprotectant in Parkinson’s and Huntington’s diseases.64 Furthermore, it was suggested that the intraocular administration of CoQ10 minimizes glutamate release and protects RGCs against ischemia-induced injury.65 Additionally, CoQ10 was also shown to protect RGCs both in vitro and in vivo against oxidative stress.66 Recently, topical CoQ10 has been shown to positively affect retinal function in patients with POAG.67
Immune modulatory treatment
Recent research has confirmed that cellular interactions play an important role in the immune system’s regulation of glaucoma.68 These include the stress-induced immune response, innate immune cells, autoreactive T-cells, dysfunctional cross-talk between neurons and glia, and overproduction of proinflammatory cytokines.57–73 One of the key proinflammatory cytokines, tumor necrosis factor-alpha (TNF-α), is secreted by damaged glial cells and through the binding of TNF-receptor-1 (TNF-R1) contributes to apoptotic RGC death.74 Dysfunction of immunoregulation of RGC-related T-cells is implicated in glaucoma.68,75
Agmatine, an anti-inflammatory agent, was shown in vitro to inhibit TNF-α production in hypoxic RGC conditions.76 Agmatine was also found to effectively lower IOP via topical administration and rescue RGCs in chronic ocular hypertensive rat models.77 Etanercept, a TNF-α blocker, is commonly used clinically for other indications. It was evidenced to inhibit microglial response, preventing axonal degeneration and subsequent RGC loss in a rat glaucoma models.78
The primary task of microglia cells, astrocytes, and Müller cells in the retina is to provide support, create an interface between RGCs and surrounding blood vessels, and help maintain the ion homeostasis. They are also able to remove excess glutamate and prevent excitotoxicity.9,79 In addition, they participate in several inflammatory processes, such as the production of inflammatory mediators, cytokines, and chemokines, which trigger their activity in glaucoma.66,80,81 It is believed that further understanding of glial changes at the molecular level may provide innovative potential treatments that seek to selectively inhibit neuronal damage due to unwanted glial activation response in glaucoma.69
RGC apoptosis may also be mediated by Fas or TNF-receptors, initiated by the efflux of cytochrome c from the mitochondria.24 It was demonstrated that glaucoma was associated with increased RGC apoptosis.82,83 TNF-α inhibitors could also be used as neuroprotective agents in line with immunomodulatory treatments as discussed previously in this paper: for example, TNF-α inhibitors could be utilized along with other agents such as Baculoviral IAP repeat-containing protein-4, a potent caspase inhibitor, or a modified viral (adeno-associated Virus) vector, which was seen to significantly protect against optic nerve axon deterioration in chronic OHT rat models.84
Stem cell therapy
Stem cells are not only able to self-renew but are also characterized by their ability to differentiate into various cell types.85 The use of stem cells in neuroprotection and neuroregeneration within glaucoma has been a burgeoning area of research and interest.
Due to the unique properties of stem cells, there are two potential avenues where stem cells could be applied in the treatment of glaucomatous damage. The most promising and powerful therapeutic potential of stem cells lies in their ability to differentiate into new cell types and to effect tissue regeneration. It is conceivable that stem cells may offer a replacement for lost RGCs and optic nerves in glaucoma.86,87 Stem cell therapies could target TM cells and RGCs, as TM cells are one of the prime culprits in the increased resistance to the aqueous outflow leading to elevated IOP. The main aqueous outflow pathway of the eye includes a series of channels, such as the TM, Schlemm’s canal, collector channels, and the episcleral venous system. In glaucoma, a reduction of TM cells is frequently observed.88 Therefore, using stem cells to replace or repair TM cells becomes an attractive and plausible approach for IOP management. Isolated and characterized stem cells from human TM proved that they were able to differentiate into TM cells and exhibit phagocytic function, which was expected to have the potential to functionally maintain the aqueous outlow.89,90 Mesenchymal stem cells (MSCs) were demonstrated to be capable of migrating predominantly to the area of tissue damage, decreasing IOP, and restoring aqueous humor drainage, as well as stimulating proliferation of nestin-expressing ocular progenitor cells in the ciliary body.91 Emre et al intravitreally transplanted bone marrow-derived and adipose tissue-derived MSCs to an OHT model and demonstrated that RGC numbers significantly improved in stem cell-treated OHT groups, with the number of cells expressing proinflammatory cytokins (interferon-gamma and TNF-α) decreasing in the MSC-transferred group.12
Embryonic stem cells have the potential to self-renew and retain pluripotency to generate all specialized cell types, including cells damaged in retinal neuroregeneration.92,93 RGC bodies are located in the retina, while the terminal axons extend into the brain, making stem cell transplantation into the inner membrane for retinal integration difficult. Cell lineage restricted mouse pluripotent stem cells converted into retinal ganglion-like cells, which exhibited long synapses, and were unable to integrate into the normal retina after transplantation.95 A safe and efficient therapeutic application of retinal progenitor cells is still in process due to the complexity of their cellular components and their transient state, along with the challenge of the retina’s multilayered architecture, but it is given full attention in experimental research nowadays.
Schwann cells (SCs) are important glial cells within the peripheral nervous system and are able to de-differentiate into progenitor cells under the condition of axonal injury, which could potentially serve as a way to replace damaged axons and provide an environment suitable for neuronal survival and axonal regrowth.96,97 Guo et al compared SC delivery to an injured optic nerve sheath with intravitreal SC delivery. SC delivery to an injured optic nerve sheath resulted in delayed but long-lasting effects on TGC protection, significantly increasing RGC survival through targeting secondary rather than primary degeneration.98
Though stem cells transplantation methods and strategies for the retina still have a long journey of development ahead, they promise a fundamental prospective neuroprotective approach for structural and functional recovery in the treatment of glaucoma. It is clear that stem cells possess multiple protective characteristics that may be capable of alleviating disease progression and promoting cell survival through various mechanisms, and we have also pointed out previously that neurotrophic factor deprivation has been implicated as one of the underlying pathophysiology of RGC death in glaucoma.33 Thus, it is hypothesized that transplantation of certain types of stem cells may activate multiple endogenous neuroprotective pathways via secretion of various factors.96 Stem cells could be applied as an intraocular slow-release device for multiple bioactive factors to achieve RGC neuroprotection, which could potentially solve the problem of the short-lived effect of neurotrophins via a single administration. To this aim, the Sadan group investigated culturing MSCs in a defined environment, which upregulated the secretion of a variety of NTFs.97 This approach revealed a neuroprotective impact on several neurodegenerative models, such as Parkinson’s disease99 and optic nerve transection.100 Flachsbarth et al genetically modified neural stem (NS) cells by intravitreally grafting them into intravitreal optic nerve crush mice models. The results were encouraging. NS cells were shown to stably express CNTF and significantly attenuate the loss of axotomized RGCs over a 4-month period.101 A Phase I clinical trial for genetically modified CNTF-secreting retinal pigment epithelium cell implant was finished last year (NCT01408472), though results are still awaited.
Diagnostic technologies of detection in glaucoma
Due to the irreversible blindness of glaucomatous damage, treatment strategies are insufficient, requiring an early and thorough diagnostic approach in tandem to lower the risk of visual impairment progression, opening a critical neuroprotective window before the damage becomes irreversible. Adequate and efficient monitoring and detection for the progression of the disease is essential.
Conventional examination of glaucoma
Other than the commonly used detection of elevated IOP, traditional tests for diagnosing glaucoma largely depend on pathological structural change observed in the retina along with functional tests. Assessment of structural changes of the optic nerve head and detection of visual field defects are fundamental in establishing the diagnosis and evaluating the progression of glaucoma.102 However, in plenty of cases, visual field loss develops unnoticed, accompanied by a substantial and unrecoverable loss of RGCs.103,104 Visual field testing is performed via standard automated perimetry (SAP) that has been used as the golden standard for managing glaucoma.101 However, previous studies have demonstrated that 25%–35% of RGCs are lost, before any visual field defect can be detected through SAP.105,106
Therefore, developing tools for early identification and diagnosis of structural damage of the optic nerve head and RNFL along with RGC loss is a pressing need within glaucoma. Along such lines, Alencar and Medeiros developed an approach to estimate RGC counts from structural and functional measurements, providing a single combined metric of structure and function.101 The method combined estimates of RGC counts obtained from OCT RNFL thickness with those obtained from SAP. Advances in spectral domain optical coherence tomography also appear to be very promising, although none of these technologies have been validated in their own right for glaucoma diagnosis.107,108
Detection of apoptosing retinal cell technology
As irreversible glaucomatous vision loss can be slowed or halted, the search for an early diagnostic tool for glaucoma is now at a peak. Apoptosis is a form of programmed cell death that is implicated in both pathological and physiological processes, which plays essential roles in several neurodegenerative diseases.109 RGC apoptosis has been demonstrated to be one of the most critical in ophthalmological study as it occurs through the whole etiological process of glaucoma.72,82,110 In early apoptosis, externalization and exposure of phosphatidylserine (PS) occur at the cell membrane surface, causing asymmetry within the phospholipid membrane of the cells.110,111 The annexin family is known to be able to bind to negatively charged PS with high affinity in a Ca2+-dependent manner.112,113 Fluorescently conjugated annexin V has been used for detection of apoptosis in a variety of studies. Technetium-99m (99m Tc)-labeled annexin V first enabled apoptosis to be detected in vivo. In recent years, this in vivo technique has been performed in various clinical trials, including post-heart transplantation,114 acute myocardial infarction,115 breast cancer,116 and rectal cancer.117
Detection of apoptosing retinal cell (DARC) provides a real-time in vivo imaging technology, using the transparent nature of the eye to view apoptotic RGCs via a nonradioactive method, allowing direct microscopic observation of cellular processes in the retina. DARC is based on the visualization of fluorescently labeled annexin V-positive cells using an ophthalmoscope such as the confocal laser scanning ophthalmoscope, with originally an argon laser of 488 nm necessary to excite the administered annexin V-bound fluorophore. Fluorescence was then detected by a solid-state photodetector with a wide band-pass filter and a short-wavelength cutoff of a 521 nm filter; finally, the signal was digitized by a frame grabber and stored on the computer.118,119
During the last decade, DARC has been refined and successfully applied in various animal studies to assess RGC apoptosis in glaucomatous rat models and Alzheimer’s disease models. Furthermore, DARC has been performed at various time points (hours, days, and months) with histological validation of RGCs labeling. The results have indicated and confirmed that RGC apoptosis happens in early stages of glaucoma.83,118,120,121 DARC has also been used in the evaluation of neuroprotective agents in glaucoma, such as beta amyloid targeting therapy,122 topical-administrated CoQ10,120 glutamate receptor agonists,121 and retinal photocoagulative treatments.123 As previously noted, the limitation of existing glaucoma diagnostic devices are that patients could only be identified when significant structural changes have already occurred and damage had progressed past the point of effective intervention. The most immediate advantageous benefit of DARC is to monitor progression and evaluate neuroprotective efficacy in real-time (Figure 1). DARC is currently undergoing a Phase I clinical trial in glaucoma (NCT02394613).
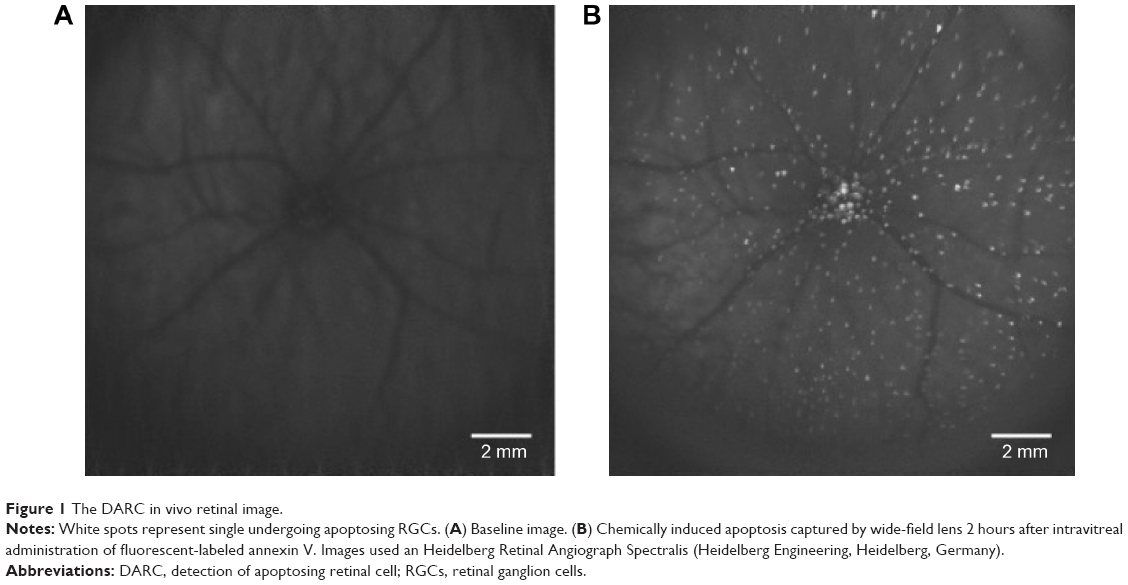 | Figure 1 The DARC in vivo retinal image. |
Conclusion
Existing available glaucoma diagnostics, medications, and therapies need further advancement, especially if visual loss of any kind is to be eliminated.124 Diagnostics are now taking into consideration a variety of structural and pathological developments within the retina to identify the early onset and progression of glaucoma with a critical focus on RGC loss and degeneration moving beyond conventional IOP measurement diagnostic alone. With this in mind, the early and accurate identification of glaucoma, providing a longer, critical therapeutic window is fundamental to reduce and halt progression. This can be achieved through neuroprotection and neuroenhancement as the next exciting step into the future of glaucoma treatment with an emphasis on early prevention.
Disclosure
MF Cordeiro is a named inventor on a patent on DARC technology owned by UCL. The authors report no other conflicts of interest in this work.
References
Kingman S. Glaucoma is second leading cause of blindness globally. Bull World Health Organ. 2004;82(11):887–888. | ||
Quigley HA, Broman AT. The number of people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol. 2006;90(3):262–267. | ||
Kaushik S, Pandav SS, Ram J. Neuroprotection in glaucoma. J Postgrad Med. 2003;49(1):90. | ||
Chang EE, Goldberg JL. Glaucoma 2.0: neuroprotection, neuroregeneration, neuroenhancement. Ophthalmology. 2012;119(5):979–986. | ||
Dielemans I, Vingerling JR, Wolfs RCW, Hofman A, Grobbee DE, de Jong PTVM. The prevalence of primary open-angle glaucoma in a population-based study in the Netherlands: the Rotterdam study. Ophthalmology. 1994;101(11):1851–1855. | ||
Leske MC, Wu SY, Hennis A, et al; BESs Study Group. Risk factors for incident open-angle glaucoma: the Barbados eye studies. Ophthalmology. 2008;115(1):85–93. | ||
Bagnis A, Papadia M, Scotto R, Traverso CE. Current and emerging medical therapies in the treatment of glaucoma. Expert Opin Emerg Drugs. 2011;16(2):293–307. | ||
Heijl A, Bengtsson B, Hyman L, Leske MC; Early Manifest Glaucoma Trial Group. Natural history of open-angle glaucoma. Ophthalmology. 2009;116(12):2271–2276. | ||
Kolko M. Present and new treatment strategies in the management of glaucoma. Open Ophthalmol J. 2015;9:89–100. | ||
Higginbotham EJ. Considerations in glaucoma therapy: fixed combinations versus their component medications. Clin Ophthalmol. 2010;4(1):1–9. | ||
Casson RJ, Chidlow G, Ebneter A, Wood JP, Crowston J, Goldberg I. Translational neuroprotection research in glaucoma: a review of definitions and principles. Clin Exp Ophthalmol. 2012;40(4): 350–357. | ||
Emre E, Yüksel N, Duruksu G, et al. Neuroprotective effects of intravitreally transplanted adipose tissue and bone marrow–derived mesenchymal stem cells in an experimental ocular hypertension model. Cytotherapy. 2015;17(5):543–559. | ||
Sena DF, Ramchand K, Lindsley K. Neuroprotection for treatment of glaucoma in adults. Cochrane Database Syst Rev. 2010;2:CD006539. | ||
O’Neill EC, Mackey DA, Connell PP, Hewitt AW, Danesh-Meyer HV, Crowston JG. The optic nerve head in hereditary optic neuropathies. Nat Rev Neurol. 2009;5(5):277–287. | ||
Osborne NN, Ugarte M, Chao M, et al. Neuroprotection in relation to retinal ischemia and relevance to glaucoma. Surv Ophthalmol. 1999;43:S102–S128. | ||
Izzotti A, Bagnis A, Saccà SC. The role of oxidative stress in glaucoma. Mutat Res. 2006;612(2):105–114. | ||
Quigley HA, McKinnon SJ, Zack DJ, et al. Retrograde axonal transport of BDNF in retinal ganglion cells is blocked by acute IOP elevation in rats. Investig Ophthalmol Vis Sci. 2000;41(11):3460–3466. | ||
D’hondt C, Srinivas SP, Vereecke J, Himpens B. Adenosine opposes thrombin-induced inhibition of intercellular calcium wave in corneal endothelial cells. Invest Ophthalmol Vis Sci. 2007;48(4):1518–1527. | ||
Kuehn MH, Kim CY, Ostojic J, et al. Retinal synthesis and deposition of complement components induced by ocular hypertension. Exp Eye Res. 2006;83(3):620–628. | ||
Dreyer EB. A proposed role for excitotoxicity in glaucoma. J Glaucoma. 1998;7(1):62–67. | ||
Michaelis EK. Molecular biology of glutamate receptors in the central nervous system and their role in excitotoxicity, oxidative stress and aging. Prog Neurobiol. 1998;54(4):369–415. | ||
Massey SC, Redburn DA. Transmitter circuits in the vertebrate retina. Prog Neurobiol. 1987;28(1):55–96. | ||
Dkhissi O, Chanut E, Wasowicz M, et al. Retinal TUNEL-positive cells and high glutamate levels in vitreous humor of mutant quail with a glaucoma-like disorder. Investig Ophthalmol Vis Sci. 1999;40:990–994. | ||
Song W, Huang P, Zhang C. Neuroprotective therapies for glaucoma. Drug Des Devel Ther. 2015;9:1469–1479. | ||
Dreyer EB, Zurakowski D, Schumer RA, Podos SM, Lipton SA. Elevated glutamate levels in the vitreous body of humans and monkeys with glaucoma. Arch Ophthalmol. 1996;114(3):299–305. | ||
Brooks DE, Garcia GA, Dreyer EB, Zurakowski D, Franco-Bourland RE. Vitreous body glutamate concentration in dogs with glaucoma. Am J Vet Res. 1997;58(8):864–867. | ||
Osborne NN. Recent clinical findings with memantine should not mean that the idea of neuroprotection in glaucoma is abandoned. Acta Ophthalmol. 2009;87(4):450–454. | ||
Dysken MW, Sano M, Asthana S, et al. Effect of vitamin E and memantine on functional decline in Alzheimer disease: the TEAM-AD VA cooperative randomized trial. JAMA. 2014;311(1):33–44. | ||
Gabelt BT, Rasmussen CA, Tektas OY, et al. Structure/function studies and the effects of memantine in monkeys with experimental glaucoma. Investig Ophthalmol Vis Sci. 2012;53(4):2368–2376. | ||
Mayama C. Calcium channels and their blockers in intraocular pressure and glaucoma. Eur J Pharmacol. 2013;739:96–105. | ||
Takahashi K, Lam TT, Edward DP, Buchi ER, Tso MOM. Protective effects of flunarizine on ischemic injury in the rat retina. Arch Ophthalmol. 1992;110(6):862–870. | ||
Koseki N, Araie M, Tomidokoro A, et al. A placebo-controlled 3-year study of a calcium blocker on visual field and ocular circulation in glaucoma with low-normal pressure. Ophthalmology. 2008;115(11): 2049–2057. | ||
Poo M. Neurotrophins as synaptic modulators. Nat Rev Neurosci. 2001;2(1):24–32. | ||
Yan Q, Wang J, Matheson CR, Urich JL. Glial cell line – derived neurotrophic factor (GDNF) promotes the survival of axotomized retinal ganglion cells in adult rats: comparison to and combination with brain-derived neurotrophic factor (BDNF). J Neurobiol. 1999;38(3):382–390. | ||
Ko M-L, Hu D-N, Ritch R, Sharma SC, Chen C-F. Patterns of retinal ganglion cell survival after brain-derived neurotrophic factor administration in hypertensive eyes of rats. Neurosci Lett. 2001;305(2): 139–142. | ||
Fu QL, Li X, Yip HK, et al. Combined effect of brain-derived neurotrophic factor and LINGO-1 fusion protein on long-term survival of retinal ganglion cells in chronic glaucoma. Neuroscience. 2009; 162(2):375–382. | ||
Parrilla-Reverter G, Agudo M, Sobrado-Calvo P, Salinas-Navarro M, Villegas-Pérez MP, Vidal-Sanz M. Effects of different neurotrophic factors on the survival of retinal ganglion cells after a complete intraorbital nerve crush injury: a quantitative in vivo study. Exp Eye Res. 2009;89(1):32–41. | ||
Ji JZ, Elyaman W, Yip HK, et al. CNTF promotes survival of retinal ganglion cells after induction of ocular hypertension in rats: the possible involvement of STAT3 pathway. Eur J Neurosci. 2004;19(2):265–272. | ||
Cheng L, Sapieha P, Kittlerová P, Hauswirth WW, Di Polo A. TrkB gene transfer protects retinal ganglion cells from axotomy-induced death in vivo. J Neurosci. 2002;22(10):3977–3986. | ||
Checa-Casalengua P, Jiang C, Bravo-Osuna I, et al. Retinal ganglion cells survival in a glaucoma model by GDNF/Vit E PLGA microspheres prepared according to a novel microencapsulation procedure. J Control Release. 2011;156(1):92–100. | ||
Zhang K, Hopkins JJ, Heier JS, et al. Ciliary neurotrophic factor delivered by encapsulated cell intraocular implants for treatment of geographic atrophy in age-related macular degeneration. Proc Natl Acad Sci U S A. 2011;108(15):6241–6245. | ||
Lambiase A, Aloe L, Centofanti M, et al. Experimental and clinical evidence of neuroprotection by nerve growth factor eye drops: implications for glaucoma. Proc Natl Acad Sci U S A. 2009;106(32):13469–13474. | ||
Miyazaki M, Ikeda Y, Yonemitsu Y, et al. Pigment epithelium-derived factor gene therapy targeting retinal ganglion cell injuries: neuroprotection against loss of function in two animal models. Hum Gene Ther. 2010;22(5):559–565. | ||
Akman A, Cetinkaya A, Akova YA, Ertan A. Comparison of additional intraocular pressure-lowering effects of latanoprost vs brimonidine in primary open-angle glaucoma patients with intraocular pressure uncontrolled by timolol – dorzolamide combination. Eye. 2005;19(2):145–151. | ||
Levkovitch-Verbin H, Harris-Cerruti C, Groner Y, Wheeler LA, Schwartz M, Yoles E. RGC death in mice after optic nerve crush injury: oxidative stress and neuroprotection. Investig Ophthalmol Vis Sci. 2000;41(13):4169–4174. | ||
Wheeler LA, Gil DW, WoldeMussie E. Role of alpha-2 adrenergic receptors in neuroprotection and glaucoma. Surv Ophthalmol. 2001;45:S290–S294. | ||
Semba K, Namekata K, Kimura A, Harada C, Mitamura Y, Harada T. Brimonidine prevents neurodegeneration in a mouse model of normal tension glaucoma. Cell Death Dis. 2014;5(7):e1341. | ||
Krupin T, Liebmann JM, Greenfield DS, Ritch R, Gardiner S. A randomized trial of brimonidine versus timolol in preserving visual function: results from the low-pressure glaucoma treatment study. Am J Ophthalmol. 2011;151(4):671–681. | ||
De Moraes CG, Liebmann JM, Greenfield DS, Gardiner SK, Ritch R, Krupin T. Risk factors for visual field progression in the low-pressure glaucoma treatment study. Am J Ophthalmol. 2012;154(4): 702–711. | ||
Kolomeyer AM, Zarbin MA. Trophic factors in the pathogenesis and therapy for retinal degenerative diseases. Surv Ophthalmol. 2014;59(2):134–165. | ||
Cuenca N, Fernández-Sánchez L, Campello L, et al. Cellular responses following retinal injuries and therapeutic approaches for neurodegenerative diseases. Prog Retin Eye Res. 2014;43:1–59. | ||
Izzotti A, Saccà SC, Cartiglia C, De Flora S. Oxidative deoxyribonucleic acid damage in the eyes of glaucoma patients. Am J Med. 2003; 114(8):638–646. | ||
Saccà SC, Pascotto A, Camicione P, Capris P, Izzotti A. Oxidative DNA damage in human trabecular meshwork and its correlation with intraocular pressure and visual field in primary open angle glaucoma. Arch Ophthalmol. 2005;123:458–463. | ||
Neufeld AH. Pharmacologic neuroprotection with an inhibitor of nitric oxide synthase for the treatment of glaucoma. Brain Res Bull. 2004;62(6):455–459. | ||
Birich TV, Birich TA, Marchenko LN, Remizonova DN, Fedylov AS. [Vitamin E in the complex treatment of patients with primary glaucoma]. Vestn Oftalmol. 1985;102(2):10–13. | ||
Cellini M, Caramazza N, Mangiafico P, Possati GL, Caramazza R. Fatty acid use in glaucomatous optic neuropathy treatment. Acta Ophthalmol Scand. 1998;76(S227):41–42. | ||
Buschini E, Fea AM, Lavia CA, et al. Recent developments in the management of dry age-related macular degeneration. Clin Ophthalmol. 2015;9:563–574. | ||
Ko M-L, Peng P-H, Hsu S-Y, Chen C-F. Dietary deficiency of vitamin E aggravates retinal ganglion cell death in experimental glaucoma of rats. Curr Eye Res. 2010;35(9):842–849. | ||
Hsu KH, de la Jara PL, Ariyavidana A, et al. Release of betaine and dexpanthenol from vitamin E modified silicone-hydrogel contact lenses. Curr Eye Res. 2014;40(3):267–273. | ||
Hsu K-H, Carbia BE, Plummer C, Chauhan A. Dual drug delivery from vitamin E loaded contact lenses for glaucoma therapy. Eur J Pharm Biopharm. 2015;94:312–321. | ||
DeFeudis FV. Ginkgo Biloba Extract (EGb 761): Pharmacological Activities and Clinical Applications. Paris: Elsevier; 1991. | ||
Detry-Morel M. Prospects in the medical treatment of glaucomatous neuropathy. Basics of neuroprotection. J Fr Ophtalmol. 1999;22(1): 122. | ||
Cybulska-Heinrich AK, Mozaffarieh M, Flammer J. Ginkgo biloba: an adjuvant therapy for progressive normal and high tension glaucoma. Mol Vis. 2012;18:390. | ||
Beal MF. Coenzyme Q10 administration and its potential for treatment of neurodegenerative diseases. Biofactors. 1999;9(2–4):261–266. | ||
Nucci C, Tartaglione R, Cerulli A, et al. Retinal damage caused by high intraocular pressure – induced transient ischemia is prevented by coenzyme Q10 in rat. Int Rev Neurobiol. 2007;82:397–406. | ||
Nakajima Y, Inokuchi Y, Nishi M, Shimazawa M, Otsubo K, Hara H. Coenzyme Q 10 protects retinal cells against oxidative stress in vitro and in vivo. Brain Res. 2008;1226:226–233. | ||
Parisi V, Centofanti M, Gandolfi S, et al. Effects of coenzyme Q10 in conjunction with vitamin E on retinal-evoked and cortical-evoked responses in patients with open-angle glaucoma. J Glaucoma. 2014; 23(6):391–404. | ||
Tezel G. Immune regulation toward immunomodulation for neuroprotection in glaucoma. Curr Opin Pharmacol. 2013;13(1):23–31. | ||
Tezel G. The immune response in glaucoma: a perspective on the roles of oxidative stress. Exp Eye Res. 2011;93(2):178–186. | ||
Wax MB, Tezel G. Immunoregulation of retinal ganglion cell fate in glaucoma. Exp Eye Res. 2009;88(4):825–830. | ||
Tezel G. The role of glia, mitochondria, and the immune system in glaucoma. Invest Ophthalmol Vis Sci. 2009;50(3):1001–1012. | ||
Naskar R, Wissing M, Thanos S. Detection of early neuron degeneration and accompanying microglial responses in the retina of a rat model of glaucoma. Investig Ophthalmol Vis Sci. 2002;43(9):2962–2968. | ||
Tezel G, Li LY, Patil RV, Wax MB. TNF- and TNF-receptor-1 in the retina of normal and glaucomatous eyes. Investig Ophthalmol Vis Sci. 2001;42(8):1787–1794. | ||
McKinnon SJ. The cell and molecular biology of glaucoma: common neurodegenerative pathways and relevance to glaucoma. Invest Ophthalmol Vis Sci. 2012;53(5):2485–2487. | ||
Wax MB, Tezel G, Yang J, et al. Induced autoimmunity to heat shock proteins elicits glaucomatous loss of retinal ganglion cell neurons via activated T-cell-derived fas-ligand. J Neurosci. 2008;28(46): 12085–12096. | ||
Hong S, Park K, Kim CY, Seong GJ. Agmatine inhibits hypoxia-induced TNF-alpha release from cultured retinal ganglion cells. Biocell. 2008;32(2):201–205. | ||
Hong S, Kim CY, Lee WS, Shim J, Yeom HY, Seong GJ. Ocular hypotensive effects of topically administered agmatine in a chronic ocular hypertensive rat model. Exp Eye Res. 2010;90(1):97–103. | ||
Roh M, Zhang Y, Murakami Y, et al. Etanercept, a widely used inhibitor of tumor necrosis factor-α (TNF-α), prevents retinal ganglion cell loss in a rat model of glaucoma. PLoS One. 2012;7(7):e40065. | ||
Bringmann A, Grosche A, Pannicke T, Reichenbach A. GABA and glutamate uptake and metabolism in retinal glial (Müller) cells. Front Endocrinol. 2013;4:48. | ||
Wang L, Cioffi GA, Cull G, Dong J, Fortune B. Immunohistologic evidence for retinal glial cell changes in human glaucoma. Invest Ophthalmol Vis Sci. 2002;43(4):1088–1094. | ||
Neufeld AH. Microglia in the optic nerve head and the region of parapapillary chorioretinal atrophy in glaucoma. Arch Ophthalmol. 1999;117(8):1050–1056. | ||
Quigley HA, Nickells RW, Kerrigan LA, Pease ME, Thibault DJ, Zack DJ. Retinal ganglion cell death in experimental glaucoma and after axotomy occurs by. Invest Ophthalmol. 1995;36(April):774–786. | ||
Cordeiro MF, Guo L, Coxon KM, et al. Imaging multiple phases of neurodegeneration: a novel approach to assessing cell death in vivo. Cell Death Dis. 2010;1(1):e3. | ||
McKinnon SJ, Lehman DM, Tahzib NG, et al. Baculoviral IAP repeat-containing-4 protects optic nerve axons in a rat glaucoma model. Mol Ther. 2002;5(6):780–787. | ||
Morrison SJ, Kimble J. Asymmetric and symmetric stem-cell divisions in development and cancer. Nature. 2006;441(7097):1068–1074. | ||
Johnson TV, Bull ND, Martin KR. Stem cell therapy for glaucoma: possibilities and practicalities. Expert Rev Ophthalmol. 2011;6(2): 165–174. | ||
Bull ND, Martin KR. Using stem cells to mend the retina in ocular disease. Regen Med. 2009;4(6):855–864. | ||
Liton PB, Challa P, Stinnett S, Luna C, Epstein DL, Gonzalez P. Cellular senescence in the glaucomatous outflow pathway. Exp Gerontol. 2005;40(8):745–748. | ||
Du Y, Roh DS, Mann MM, Funderburgh ML, Funderburgh JL, Schuman JS. Multipotent stem cells from trabecular meshwork become phagocytic TM cells. Invest Ophthalmol Vis Sci. 2012;53(3): 1566–1575. | ||
Du Y, Yun H, Yang E, Schuman JS. Stem cells from trabecular meshwork home to TM tissue in vivo. Investig Ophthalmol Vis Sci. 2013; 54(2):1450–1459. | ||
Manuguerra-Gagné R, Boulos PR, Ammar A, et al. Transplantation of mesenchymal stem cells promotes tissue regeneration in a glaucoma model through laser-induced paracrine factor secretion and progenitor cell recruitment. Stem Cells. 2013;31(6):1136–1148. | ||
Meyer JS, Katz ML, Maruniak JA, Kirk MD. Embryonic stem cell-derived neural progenitors incorporate into degenerating retina and enhance survival of host photoreceptors. Stem Cells. 2006;24(2): 274–283. | ||
Vugler A, Lawrence J, Walsh J, et al. Embryonic stem cells and retinal repair. Mech Dev. 2007;124(11):807–829. | ||
Chen M, Chen Q, Sun X, et al. Generation of retinal ganglion – like cells from reprogrammed mouse fibroblasts. Invest Ophthalmol Vis Sci. 2010;51(11):5970–5978. | ||
Jagatha B, Divya MS, Sanalkumar R, et al. In vitro differentiation of retinal ganglion-like cells from embryonic stem cell derived neural progenitors. Biochem Biophys Res Commun. 2009;380(2):230–235. | ||
Bull ND, Johnson TV, Martin KRÃ. Stem cells for neuroprotection in glaucoma. Prog Brain Res. 2008;173(08):511–519. | ||
Sadan O, Shemesh N, Cohen Y, Melamed E, Offen D. Adult neurotrophic factor-secreting stem cells: a potential novel therapy for neurodegenerative diseases. Isr Med Assoc J. 2009;11(4):201–204. | ||
Guo L, Davis B, Nizari S, et al. Direct optic nerve sheath (DONS) application of Schwann cells prolongs retinal ganglion cell survival in vivo. Cell Death Dis. 2014;5(10):e1460. | ||
Sadan O, Bahat-Stromza M, Barhum Y, et al. Protective effects of neurotrophic factor-secreting cells in a 6-OHDA rat model of Parkinson disease. Stem Cells Dev. 2009;18(8):1179–1190. | ||
Levkovitch-Verbin H, Sadan O, Vander S, et al. Intravitreal injections of neurotrophic factors secreting mesenchymal stem cells are neuroprotective in rat eyes following optic nerve transection. Invest Ophthalmol Vis Sci. 2010;51(12):6394–6400. | ||
Flachsbarth K, Kruszewski K, Jung G, et al. Neural stem cell-based intraocular administration of ciliary neurotrophic factor attenuates the loss of axotomized ganglion cells in adult mice. Invest Ophthalmol Vis Sci. 2014;55(11):7029–7039. | ||
Schwartz M, Yoles E. Neuroprotection: a new treatment modality for glaucoma? Curr Opin Ophthalmol. 2000;11(2):107–111. | ||
Medeiros FA, Alencar LM, Zangwill LM, Sample PA, Weinreb RN. The relationship between intraocular pressure and progressive retinal nerve fiber layer loss in glaucoma. Ophthalmology. 2009;116(6):1125–1133. | ||
Harwerth RS, Carter-Dawson L, Smith EL, Barnes G, Holt WF, Crawford MLJ. Neural losses correlated with visual losses in clinical perimetry. Invest Ophthalmol Vis Sci. 2004;45(9):3152–3160. | ||
Kerrigan-Baumrind LA, Quigley HA, Pease ME, Kerrigan DF, Mitchell RS. Number of ganglion cells in glaucoma eyes compared with threshold visual field tests in the same persons. Invest Ophthalmol Vis Sci. 2000;41(3):741–748. | ||
Medeiros FA, Lisboa R, Weinreb RN, Liebmann JM, Girkin C, Zangwill LM. Retinal ganglion cell count estimates associated with early development of visual field defects in glaucoma. Ophthalmology. 2013;120(4):736–744. | ||
Bussel II, Wollstein G, Schuman JS. OCT for glaucoma diagnosis, screening and detection of glaucoma progression. Br J Ophthalmol. 2013;98(suppl 2):ii15–ii19. | ||
El Chehab H, Delbarre M, Maréchal M, et al. [New neuroretinal rim analysis with spectral domain optical coherence tomography, spectralis (Heidelberg Engineering, Germany). Preliminary study]. J Fr Ophtalmol. 2015;38(1):46–52. | ||
Friedlander RM. Apoptosis and caspases in neurodegenerative diseases. N Engl J Med. 2003;348(14):1365–1375. | ||
Bizrah M, Dakin SC, Guo L, et al. A semi-automated technique for labeling and counting of apoptosing retinal cells. BMC Bioinformatics. 2014;15(1):169. | ||
van Engeland M, Nieland LJ, Ramaekers FC, Schutte B, Reutelingsperger CP. Annexin V-affinity assay: a review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry. 1998;31(1):1–9. | ||
Carroll AD, Moyen C, Van Kesteren P, Tooke F, Battey NH, Brownlee C.Ca2+, annexins, and GTP modulate exocytosis from maize root cap protoplasts. Plant Cell. 1998;10(8):1267–1276. | ||
Meers P, Mealy T. Calcium-dependent annexin V binding to phospholipids: stoichiometry, specificity, and the role of negative charge. Biochemistry. 1993;32(43):11711–11721. | ||
Blankenberg FG, Katsikis PD, Tait JF, et al. In vivo detection and imaging of phosphatidylserine expression during programmed cell death. Proc Natl Acad Sci U S A. 1998;95(11):6349–6354. | ||
Flotats A, Carrió I. Non-invasive in vivo imaging of myocardial apoptosis and necrosis. Eur J Nucl Med Mol Imaging. 2003;30(4): 615–630. | ||
Yang DJ, Azhdarinia A, Wu P, et al. In vivo and in vitro measurement of apoptosis in breast cancer cells using 99mTc-EC-annexin V. Cancer Biother Radiopharm. 2001;16(1):73–83. | ||
Horisberger K, Erben P, Ströbel P, et al. Annexin and survivin in locally advanced rectal cancer: indicators of resistance to preoperative chemoradiotherapy? Onkologie. 2010;33(8–9):439–444. | ||
Cordeiro MF, Guo L, Luong V, et al. Real-time imaging of single nerve cell apoptosis in retinal neurodegeneration. Proc Natl Acad Sci U S A. 2004;101(36):13352–13356. | ||
Cordeiro MF, Migdal C, Bloom P, Fitzke FW, Moss SE. Imaging apoptosis in the eye. Eye (Lond). 2011;25(5):545–553. | ||
Guo L, Moss SE, Alexander RA, Ali RR, Fitzke FW, Cordeiro MF. Retinal ganglion cell apoptosis in glaucoma is related to intraocular pressure and IOP-induced effects on extracellular matrix. Invest Ophthalmol Vis Sci. 2005;46(1):175. | ||
Guo L, Salt TE, Maass A, et al. Assessment of neuroprotective effects of glutamate modulation on glaucoma-related retinal ganglion cell apoptosis in vivo. Invest Ophthalmol Vis Sci. 2006;47(2):626. | ||
Guo L, Salt TE, Luong V, et al. Targeting amyloid-beta in glaucoma treatment. Proc Natl Acad Sci U S A. 2007;104(33):13444–13449. | ||
Schmitz-Valckenberg S, Guo L, Maass A, et al. Real-time in vivo imaging of retinal cell apoptosis after laser exposure. Invest Ophthalmol Vis Sci. 2008;49(6):2773. | ||
Prevention of blindness and visual impairment: Priority eye diseases. World Health Organization. Available from: http://www.who.int/blindness/causes/priority/en/index9.html. Accessed October 16, 2015. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.