Back to Journals » OncoTargets and Therapy » Volume 13
Curcumin Negatively Regulates Cigarette Smoke-Induced Renal Cell Carcinoma Epithelial–Mesenchymal Transition Through the ERK5/AP-1 Pathway
Authors Zhang T, Zhao L, Zhang T, Wu W, Liu J, Wang X, Wan Y, Geng H, Sun X, Qian W, Yu D ![]()
Received 3 June 2020
Accepted for publication 20 August 2020
Published 29 September 2020 Volume 2020:13 Pages 9689—9700
DOI https://doi.org/10.2147/OTT.S265847
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjeev K. Srivastava
Tao Zhang,1,* Li Zhao,2,* Taotao Zhang,1,* Wangyu Wu,1 Jie Liu,3 Xian Wang,1 Yang Wan,4 Hao Geng,1 Xin Sun,1 Weiwei Qian,1 Dexin Yu1
1Department of Urologic Surgery, The Second Affiliated Hospital of Anhui Medical University, Hefei 230032, People’s Republic of China; 2Department of Urologic Surgery, The Fourth Affiliated Hospital of Nanjing Medical University, Nanjing, 210048, People’s Republic of China; 3Department of Urologic Surgery, The First People’s Hospital of Wuhu, Wuhu 241000, People’s Republic of China; 4Department of Hematology/Hematological Lab, The Second Affiliated Hospital of Anhui Medical University, Hefei 230032, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Dexin Yu
Department of Urologic Surgery, The Second Affiliated Hospital of Anhui Medical University, 80 Feicui Road, Hefei 230032, People’s Republic of China
Email [email protected]
Background: So far, little research has been conducted regarding the underlying mechanism of renal carcinogenesis at molecular level. Epithelial–mesenchymal transition (EMT) exerts an important part during tumor genesis as well as the development through mitogen-activated protein kinase (MAPK) pathways. Therefore, we hypothesized that EMT could promote renal cell carcinoma (RCC) progression via the ERK5/AP-1pathway.
Materials and Methods: The RCC cell lines were utilized to be the models with in vitro exposure to cigarette smoke extract (CSE). We used the curcumin for the EMT intervention study. In the present study, immunohistochemistry (IHC), Western blotting, and real-time quantitative reverse transcription PCR had been used to determine the experimental results. EMT phenotypic alterations were assessed by changes in cell morphology, invasion and transfer ability, as well as expression of epithelial and mesenchymal markers.
Results: In human renal cell carcinoma tissue, E-cadherin expression within the smoking renal cancer patients was down-regulated compared with that among the non-smokers. However, Vimentin, N-cadherin, and TWIST levels increased (P< 0.05). Significantly, we clarified that ERK5/AP-1 exerted positive regulation on the renal cell carcinoma EMT mediated by CS, which was suggested based on the results of CS activating the ERK5/AP-1 pathway, as well as ERK5 inhibition via XMD8-92 reversed AP-1 protein levels and the EMT process. Furthermore, curcumin showed the same inhibitory effect as XMD8-92 and significantly reversed CS-induced EMT through inhibiting the ERK5/AP-1 signaling pathway.
Conclusion: The above results indicated that ERK5/AP-1 signaling pathway exerts a vital part for CS-associated RCC development and cancer intervention.
Keywords: renal cell carcinoma, epithelial–mesenchymal transition, cigarette smoke, ERK5/AP-1 pathway, curcumin
Introduction
Epidemiological investigations found that over 90% of renal cancers are renal cell carcinoma, and 50% of renal cancer in patients is associated with long-term smoking.1,2 Although cigarette smoke (CS) shows tight correlation with renal tumor occurrence and progression, few studies have examined the mechanisms by which CS is involved in renal carcinoma progression at molecular level. Consequently, the current research aimed to investigate CS pathogenesis in renal cell carcinoma at molecular level.
Epithelial–mesenchymal transition (EMT) has been recognized as an essential physiopathological process for tumor genesis and progression.3 In the process of EMT, cells have lost the epithelial characteristics, obtaining the e mesenchymal characteristics. Moreover, abundant evidence demonstrates that EMT could enhance the invasive capabilities and metastatic growth attributes of tumor cells.4,5 Additionally, evidence also suggests that EMT plays a critical role in initiating tumor genesis.6,7 However, no molecular mechanism has been proposed for CS-induced EMT in renal cell carcinoma.
Extracellular signal-regulated kinase 5 (ERK5), as one of the MAPKfamily members, has been rarely investigated. The ERK5/AP-1 (the transcriptional factor TF subunit and the Jun proto-oncogene) pathway participates in numerous vital cell processes, such as angiogenesis, apoptosis, proliferation, and viability of cells.8 The AP-1, which is constituted by FOS and JUN proteins, participates in mediating tumorigenesis and progression. Based on compelling evidence from our preliminary research, the activity of MAPK/AP-1 plays a vital part in the CS effect.9–11 Besides, activating the ERK5/AP-1 pathway can up-regulate EMT induced by CS within non-carcinoma urothelial cells.12 Arias-Gonzalez et al found that high levels of ERK5 are associated with a poor prognosis of RCC.13 Nonetheless, little research concentrates on the role of ERK5/AP-1 in regulating the EMT in renal carcinoma induced by CS.
Curcumin is currently the most widely studied phytochemical, which has a distinct safety profile. As one of the primary turmeric bioactive components, curcumin is the traditional Chinese herbal medicine that has been used for a long time across Asia.14 According to the previous studies both in vivo and in vitro, curcumin exerts anti-tumor effects on various cancer types, such as bladder, lung, thyroid, and colorectal cancer.4,5,12 Nonetheless, little research is carried out to examine its interventional effects.
So far, no studies have examined whether curcumin negatively regulates cigarette smoke-induced renal cell carcinoma EMT. Consequently, the present research aimed to examine the role of ERK5/AP-1 in regulating EMT induced by CS, aiming to further clarify the mechanism of CS-induced renal cell carcinoma progression, and provide new ideas for cancer intervention.
Materials and Methods
Chemicals and Reagents
ACHN and 786-O cells were provided by the American Type Culture Collection (ATCC, Manassas, VA, USA). Dimethyl sulfoxide (DMSO) and the remaining reagents were provided by Sigma-Aldrich (St. Louis, MO, USA) and had the greatest purity. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) had been provided by Sigma-Aldrich. Antibodies recognizing the total-ERK5, phosphorylated-ERK5 (p-ERK5), c-jun, phosphorylated-c-jun (p-c-jun), E-cadherin, c-fos, phosphorylated-c-fos (p-c-fos), Vimentin, and N-cadherin were provided by the Cell Signaling Technology (Monoclonal, Rabbit, Danvers, MA, USA). Moreover, those antibodies recognizing Twist family BHLH transcription factor (TWIST) and zonula occludens protein 1 (ZO-1, which is also referred to as the tight junction protein 1 (TJP1)), had been offered by Santa Cruz Biotechnology (Monoclonal, Rabbit, Santa Cruz, CA, USA). Biogot Technology (Nanjing, China) supplied the antibodies against glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Nanjing, China). Curcumin had been provided by Sigma-Aldrich (at the purity of 99.0%). XMD8-92 had been provided by the Selleck Chemicals (Huston, TX, USA). In terms of the IHC staining tests, mouse antibodies against E-cadherin and N-cadherin, together with rabbit antibody against Vimentin, and the DAB chromogenic kit, were provided by the Beijing Zhongshan Jinqiao Company. Moreover, other material sources are marked within the manuscript.
CSE Preparation
CSE was prepared freshly for all experiments according to a previously reported method.9 The commercially available cigarettes (the Hongtashan filter-tipped cigarettes manufactured in Yunnan Province of China; each containing 1.1 mg nicotine and 12 mg tar) were smoked. Subsequently, one vacuum was used to draw the mainstream smoke via the pre-warmed (37°C, 10 mL) fetal bovine serum (FBS)-free 1640 medium containing streptomycin and penicillin at the speed of 5 min/cigarette. To acquire the 100% solution, a 0.22-µm filter was used to filter and further dilute the resultant solution. Different concentrations of CSE were used to treat the 786-O and ACHN cells for 6 days.
Culture and Treatment of Cells
The 786-O and ACHN cell lines had been grown within the 10 cm2 flasks filled with the Roswell Park Memorial Institute (RPMI) 1640 medium containing antibiotics (100 µg/mL streptomycin and 100 U/mL penicillin) as well as the 10% FBS, and cultured under the 37°C conditions within an incubator under 5% CO2. Afterwards, the medium would be replaced daily till reaching 80–90% cell confluence. Later, cells were subjected to CSE curcumin or XMD8-92 treatments at different contents.
Preparation of Case Data
The current work gained approval from the Ethics Committee at the Second Affiliated Hospital of Anhui Medical University. Sixty cases with renal cell carcinoma diagnosed at the Second Affiliated Hospital of Anhui Medical University from February 2017 to February 2019 and treated with surgery in urology were selected. Both smoking groups (SG) and non-smoking groups (NSG) comprised 30 cases. No patients underwent neoadjuvant targeted therapy or interventional therapy before surgery. The patients were asked about their smoking status before surgery. Immediately after the surgical specimen was isolated, the cancer tissue and some normal kidney tissues were taken and frozen at −80°C.
MTT Assay
The 786-O and ACHN cell lines were subjected to CSE treatments at various doses (0%, 0.1%, 0.25%, 0.5%, 1%, and 2%) for 6 days. Then, the MTT assay would be performed to examine the viability of cells under different CSE concentration treatments. XMD8-92 (0–3 µM) or curcumin (0–6 µM) was added into 786-O and ACHN cells with CSE (0%, 0.25%, 0.5%) to select the appropriate concentrations for further experiments.
Wound-Healing Assay
Medium without CSE or with CSE at 0.25% or 0.5% was used to pretreat 786-O and ACHN cells for 4 days. Then, the cells lines would be grown at the concentration of 4 × 105/well within the 6-well plates and incubated for 24h in serum-free medium before wounding. Using a 20-µL pipette tip, a wound was made on the surface of the cell layer on the plates. The movement of the cells at the sides of the scratch was evaluated at 0, 12, and 24 h. Photographs were taken and evaluated under a light microscope.
Transwell Assay
For cell pretreatment, refer to the wound-healing assay section. The assay was carried out within the 24-well Boyden chamber coated with the polycarbonate membrane (with the pore size of 8-µm, Millipore, Billerica, MA, USA) as well as Matrigel for the formation of the matrix barrier. RPMI 1640 medium would be put into bottom chamber with 10% FBS to be the chemoattractant. Afterwards, the cells were cultured for 24 h in the incubator. In addition, those that were still staying on the internal channel membrane would be eliminated by the cotton swab, whereas those adhering to the lower membranes would be subjected to methanol fixation and crystal violet staining. Afterwards, the migrating cell number in each microscopic field of view (FOV) was recorded and examined.
Western Blotting
Proteins were extracted from 786-O and ACHN cells, and surgical specimen tissues (including cancer and normal kidney tissues). Western blotting was carried out to determine the levels of protein. To conduct densitometry, each protein band on the blot was determined by the Eagle Eye II software (Stratagene Inc., La Jolla, CA, USA)
Quantitative Real-Time Reverse Transcription PCR (qRT-PCR)
Relative Vimentin and E-cadherin mRNA expression levels in 786-O and ACHN cells were assessed with qRT-PCR, and GAPDH was used as the reference for loading. All primers involved in the current study were shown below: E-cadherin, 5′-TCGACACCCGATTCAAAGTGG-3′(forward), 5′-TTCCAGAAACGGAGGCCTGAT-3′ (reverse); Vimentin, 5′-CCTTGACATTGAGATTGCCA-3′ (forward), 5′-GTATCAACCAGAGGGAGTGA-3′(reverse); and GAPDH, 5′-GCTGCCCAACGCACCGAATA-3′ (forward), 5′- GAGTCAACGGATTTGGTCGT-3′(reverse).
Immunohistochemistry
Wax blocks from 60 patients (SG: 30 cases and NSG: 30 cases) were chosen and sliced into sections. Conventional methods of dewaxing, hydration, antigen retrieval, dropwise addition of hydrogen peroxide, and the addition of appropriate amounts of EMT marker protein antibodies were used. The sections were subjected to 60 min of incubation under 37°C, after which, antibody would be rinsed off, an appropriate amount of secondary antibody would be jointed, and all sections were subsequently placed under ambient temperature for a period of 20 min. Thereafter, secondary antibody was rinsed off, DAB staining solution was added, and staining degree would be monitored. The sections were rinsed with distilled water to a suitable depth and dyeing was completed. Finally, the sections were counterstained with hematoxylin and sealed. The results of section staining were interpreted by two experienced pathologists. When the interpretation results were inconsistent, a third pathologist was asked to make the final decision. Stain evaluation using semi-quantitative counting method: determine tissue staining intensity under light microscope 100 times (0: no staining; 1: low intensity dyeing; 2: medium intensity dyeing; 3: strong dyeing), then after 400 times to determine the dyeing range (0: <5%; 1: 5–25%; 2: 26–75%; 3:> 75%). Meanwhile, the intensity score and the range score are added to obtain the final IHC score (0–6 points).
Statistical Analysis
Data were analyzed with the SPSS 19.0 (IBM Corp., Armonk, NY, USA) software. Results were expresses as mean ±S.D. from at least three independent experiments. Statistical analysis was performed according to Student`s t-test by one-way analysis of variance. The Student`s t-test was used to compare the labeling index and relative density for different treatments and χ2 test was used for qualitative data. At the same time, a difference of *P< 0.05 would be deemed to be of significant difference.
Results
Patient Demographic Data
Totally 60 patients were included, including 30 in the smoking and 30 in the non-smoking groups. The tumors were staged as stages I–IV in accordance with the American Joint Committee on Cancer (AJCC) 2010 renal cancer classification criteria. The tumors were graded according to the pathological results of the patients, and the levels 1 and 2 of the Fuhrman and WHO/ISUP grades were unified as high differentiation. Those assessed as grade 3 by the two methods were unified as moderate differentiation, and those assessed as grade 4 by the two methods were unified into low differentiation. The maximum diameter of the tumors for smoking group would be increased compared with that for non-smoking group, and the stage IV patient number and patients with poor tumor differentiation for smoking group were highly compared with those for non-smoking counterpart. There were fewer stage I tumors, and fewer patients had highly differentiated tumors in the smoking group relative to those for non-smoking group. However, differences in tumor stage and tumor differentiation between them were not statistically significant (Table 1).
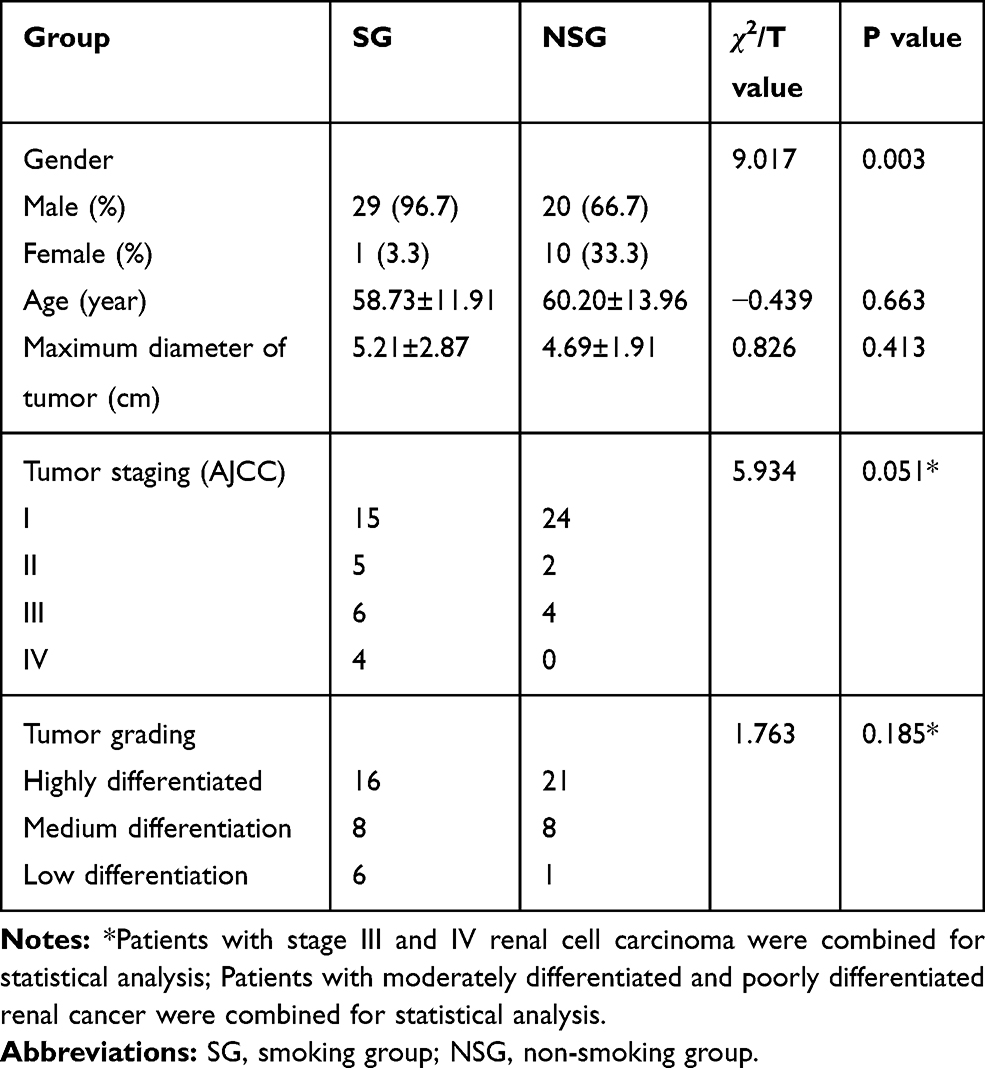 |
Table 1 Demographic Data of Two Groups of Patients with RCC (n=60, χ±s) |
CSE Induces EMT in Renal Cell Carcinoma Cells
To examine the CSE influence on inducing EMT, 786-O and ACHN cell lines were subjected to CSE exposure at different contents for a period of 6 days. CSE treatment at 0.5% dose would not significantly affect the viability of cells (Figure 1A). As a result, 0.25% and 0.5% CSE were chosen as the doses for subsequent experiments. Besides, 786-O and ACHN cells were subjected to CSE treatment for a period of 6 days, leading to obvious morphologic changes to the spindle-like mesenchymal one from the epithelial round shape (Figure 1B). Afterwards, all cells had become thinner and longer, and slender tails were generated in some of them. Transwell and Wound-healing assays would be carried out to further examine the CSE role in EMT. According to our results, CSE increased the migratory capacities of renal cell carcinoma cells significantly (Figure 1C). Additionally, the EMT process is also demonstrated by the changes in mesenchymal and epithelial marker expression. In addition, Western blotting was carried out to analyze the EMT protein markers, including N-cadherin, Vimentin, E-cadherin and ZO-1. The ZO-1 and E-cadherin levels within cells subjected to 0.25% and 0.5% CSE treatment were significantly decreased versus the control (P<0.01). The N-cadherin and Vimentin levels were up-regulated relative to those in control cells in vitro (Figure 2A). Similar results were detected for the E-cadherin and Vimentin mRNA expression in CSE-treated 786-O and ACHN cells (Figure 2B).
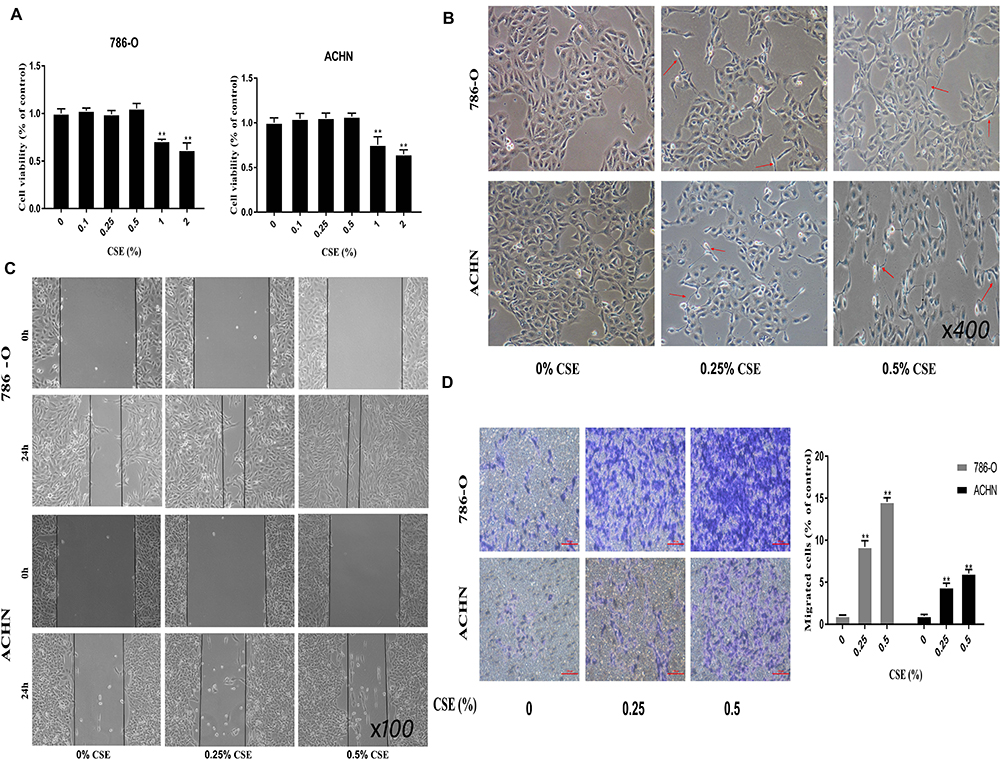 |
Figure 1 CSE induces EMT in RCC cells. (A) 786-O and ACHN cell lines are subjected to CSE exposure at different doses for a 6-day period. The viability of cells is not significantly influenced in CSE - treated cells at the dose of 0.5%. Moreover, cells are treated by CSE at the dose of ≥1%, which is found to show RCC cell toxicity. (B) CSE can induce the morphologic changes from the epithelial round shape to the spindle-resembling mesenchymal form. The cells become thinner and longer, and slender tails are generated in some cells, as indicated by the red arrow (×400 magnification). (C and D) Transwell migration and Wound-healing assays (×100 magnification) demonstrate that CSE increased migratory capacities of renal cell carcinoma cells significantly. Data are expressed in the form of mean ± SD. **P< 0.01 relative to control group. |
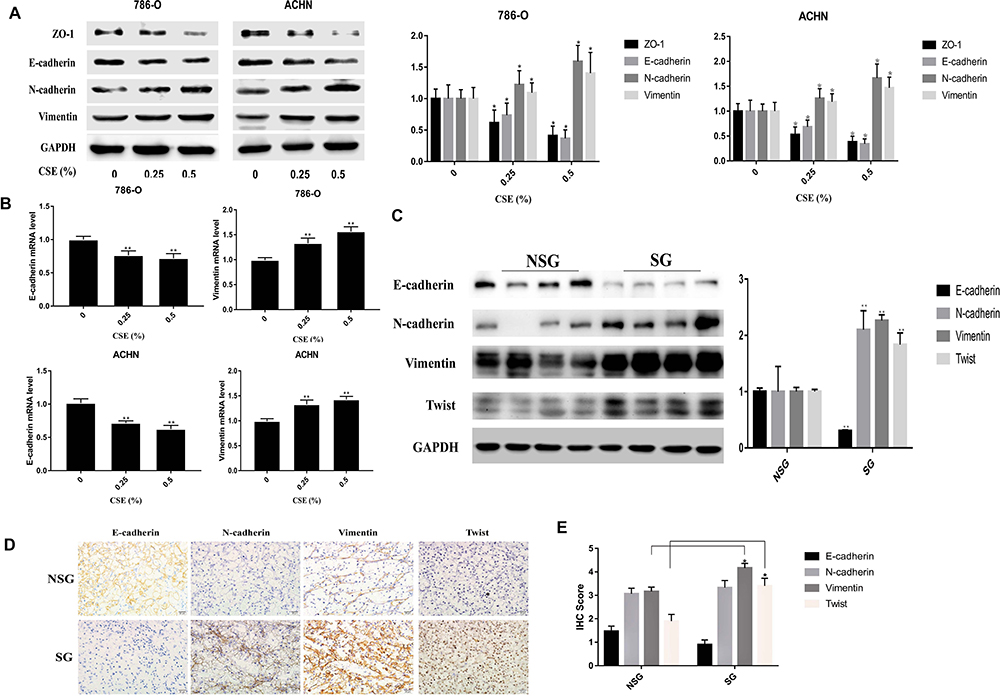 |
Figure 2 CSE alters EMT marker expression within RCC tissues and cells. (A) Epithelial marker levels, ZO-1 and E-cadherin, at 0.25% CSE and 0.5% CSE were significantly decreased versus the control. However, N-cadherin and Vimentin levels are elevated. (B) Similar results were detected for E-cadherin and Vimentin mRNA expression in CSE-treated RCC cells. (C) In tumor tissues, E-cadherin level is down-regulated of smoking patients relative to those of non-smoking patients, while Vimentin, N-cadherin, and Twist levels increased, as suggested by Western blotting. (D) IHC presented that N-cadherin and E-cadherin staining is more intense within cell membrane, while Vimentin and TWIST showed deeper staining in the cytoplasm. E-cadherin was highly expressed in non-smoking group RCC and poorly expressed in the RCC tissues of the smoking group. N-cadherin, TWIST and Vimentin were poorly expressed in the non-smoking group but were highly expressed in the smoking group. (E) The IHC scores of E-cadherin: SG (0.90 ± 1.09); NSG (1.47 ± 1.20) (P = 0.0604), the smoking group was down-regulated, but the difference was not statistically significant. The IHC scores of N-cadherin: SG (3.33±1.63); NSG (3.07±1.28), the difference was not statistically significant (P = 0.4837). The IHC scores of Vimentin: SG (4.17±1.05); NSG (3.17±1.02), the smoking group was higher than the non-smoking group (P<0.01). The IHC scores of TWIST: SG (3.4±0.33); NSG (1.9±0.29), the smoking group was higher than the non-smoking group (**P<0.01). Data are expressed in the form of mean ± SD. *P< 0.05, relative to respective control group. **P<0.01 relative to respective control group. |
CSE Activates the ERK5/AP-1 Pathway
To examine whether the ERK5/AP-1 signaling pathway was triggered following exposure of 786-O and ACHN cells to CSE for a 6-day period, the ERK5 and AP-1 protein levels were detected. Findings suggested that the t-ERK5, -fos and -jun levels were notably decreased depending on the CSE dose. However, the phosphorylated-ERK5, -fos and -jun levels were notably increased (Figure 3A). Correspondingly, inhibition of ERK5 (XMD8-92) decreased p-c-fos and p-c-Jun activation, suggesting the positive modulation of ERK5 on the AP-1 pathway (Figure 4A).
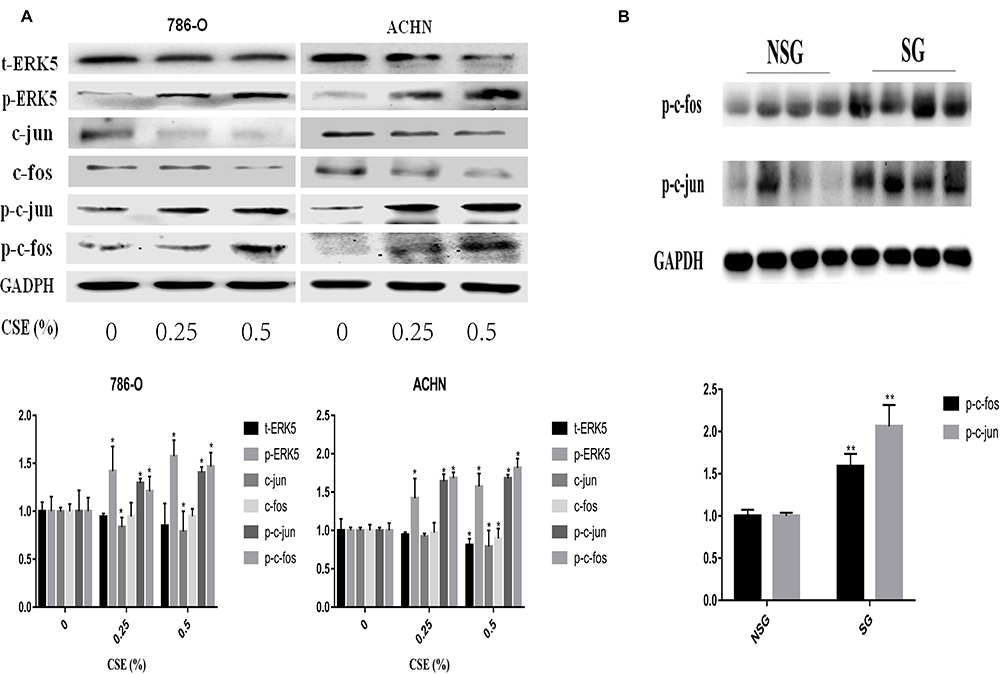 |
Figure 3 CSE can activate the ERK5/AP-1 signaling pathway. (A) t-ERK5, -fos and -jun levels were notably decreased depending on the CSE dose. However, the phosphorylated-ERK5, -fos and -jun levels were notably increased via Western blotting. (B) Western blotting analysis in RCC tissue also detected an increase in AP-1 protein (p-c-fos and p-c-jun) levels. Data are expressed in the form of mean ± SD. *P< 0.05, relative to respective control group. **P<0.01, relative to respective NSG group. |
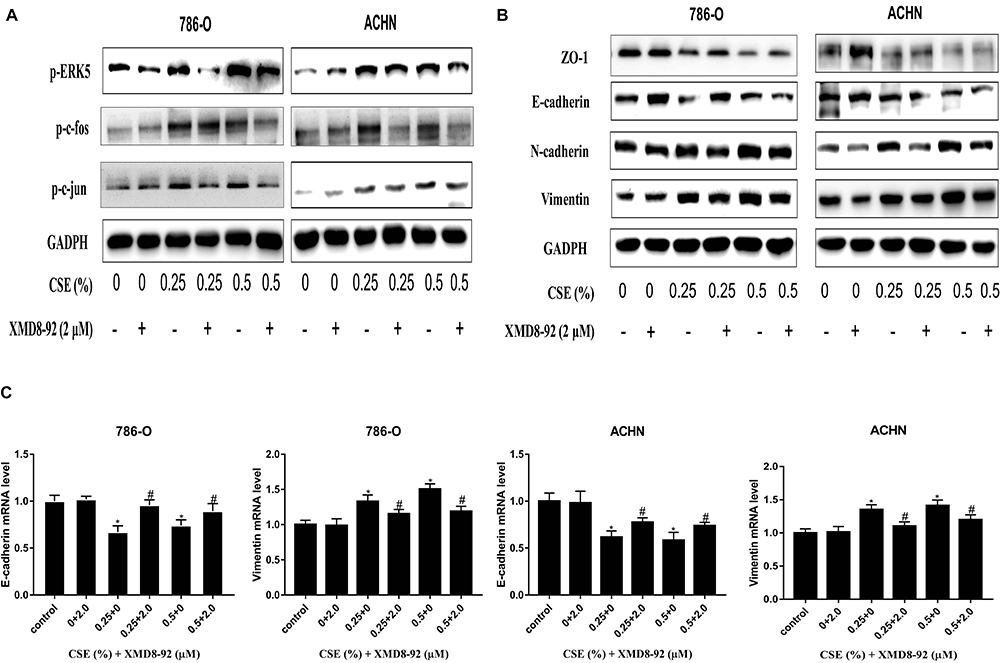 |
Figure 4 Molecular changes were further tested in XMD8-92-treated RCC cells. (A) XMD8-92 reduced p-c-jun and p-c-fos levels according to ERK5 suppression. (B and C) Meanwhile, inhibition of ERK5 results in ZO-1 and E-cadherin down-regulation, as well as Vimentin and N-cadherin up-regulation at both mRNA and protein levels. Data are expressed in the form of mean ± SD. *P<0.05, relative to control group; #P <0.05, XMD8-92 group relative to respective CSE group. |
CSE Altered Expression of EMT Markers and Activated AP-1 in RCC Tissue
In tumor tissues, according to Western blotting, E-cadherin expression was down-regulated of smoking group compared with that for non-smoking one (P<0.01), while the N-cadherin, Vimentin, and TWIST levels were up-regulated (P<0.05) (Figure 2C). Western blotting analysis in renal cell carcinoma tissue also detected an increase in AP-1 proteins (p-c-fos, p-c-jun) levels (Figure 3B). To further examine Vimentin, N-cadherin, TWIST and E-cadherin expression within renal clear cell carcinoma, IHC was used to detect the four proteins. As shown in Figure 2E, the IHC scores of E-cadherin: SG (0.90 ± 1.09); NSG (1.47 ± 1.20), (P=0.0604). The IHC scores of N-cadherin: SG (3.33±1.63); NSG (3.07±1.28), (P=0.4837). The IHC scores of Vimentin: SG (4.17±1.05); NSG (3.17±1.02), (P<0.01). The IHC scores of TWIST: SG (3.4±0.33); NSG (1.9±0.29), (P<0.01). As presented in Figure 2D, we find that E-cadherin showed high expression within RCC tissue from the non-smoking group, showing low expression in RCC tissues of the smoking group. N-cadherin, TWIST and Vimentin were poorly expressed in non-smoking group and highly expressed in smoking group.
ERK5/AP-1 Inhibition Diminished EMT Induced by CSE Within the RCC Cell Line
The specific inhibitor of ERK5, XMD8-92, is frequently used for in vitro trials. Besides, 786-O and ACHN cells were exposed to XMD8-92 (1–3 µM with or without 0.25% and 0.5% CSE) for 6 days. Finally, 2 µM XMD8-92 was selected as a suitable concentration for subsequent experiments (Figure 5A). Then, it could be observed that XMD8-92 attenuated the invasion and migration capacities of cells enhanced by CSE (Figure 5B and C). At the molecular level, XMD8-92 suppression of ERK5 reduced p-c-jun and p-c-fos levels (Figure 4A). Meanwhile, inhibition of ERK5 resulted in the down-regulated ZO-1 and E-cadherin, whereas up-regulated Vimentin and N-cadherin at both mRNA and protein levels (Figure 4B and C). The above findings are in consistent with CSE inducing EMT in ACHN and 786-O cells via the ERK5 signaling pathway.
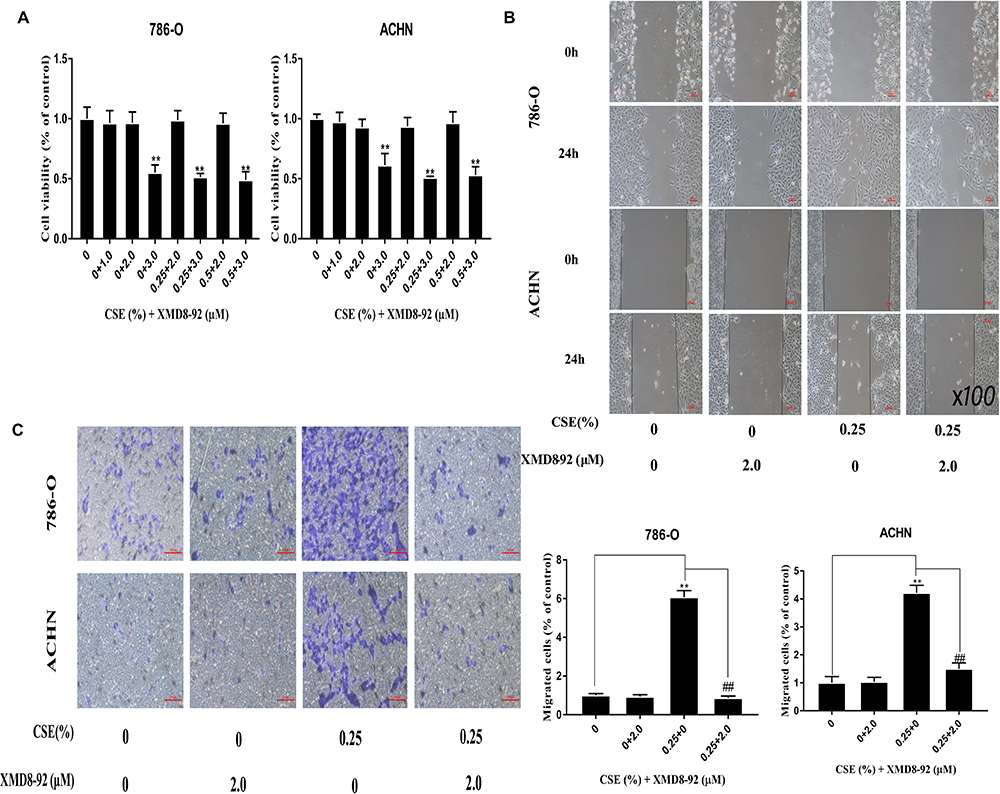 |
Figure 5 Inhibiting ERK5/AP-1 diminishes the EMT induced by CSE within RCC cell lines. (A) 786-O and ACHN cells were exposed to XMD8-92 (1–3 µM with or without 0.25–0.5% CSE) for 6 days. 2 µM of XMD8-92 is chosen to be the most suitable dose in subsequent assays. (B and C) Transwell and wound-healing assays are carried out to observe the XMD8-92 effect in the attenuation of CSE-enhanced cell migration. Data are expressed in the form of mean ± SD. **P<0.01 relative to control group, ##P<0.01, XMD8-92 group relative to respective CSE group. |
Curcumin Decreased the EMT Induced by CSE Through Suppressing ERK5/AP-1
Curcumin is extensively used in clinical applications because of its anti-tumor, anti-oxidation, and anti-inflammatory effects. To examine the interference of curcumin EMT induced by CSE, MTT assays were carried out to screen a suitable dose of curcumin (5 µM) for subsequent experiments (Figure 6A). A wound-healing assay showed that the cells’ CSE-enhanced migration ability was also reduced through curcumin (Figure 6B). Afterwards, it was found that treating cells with curcumin inhibited the ERK5/AP-1 activation induced by CSE (Figure 7A). Moreover, according to our results, curcumin treatment up-regulated the ZO-1 and E-cadherin mRNA and protein levels, and down-regulated the levels of and N-cadherin and Vimentin, demonstrating curcumin-mediated inhibition of EMT induced by CSE (Figure 7B and C). Therefore, curcumin also has an anti-tumor effect in RCC and regulates the EMT process by inhibiting the ERK5/AP-1 signaling pathway.
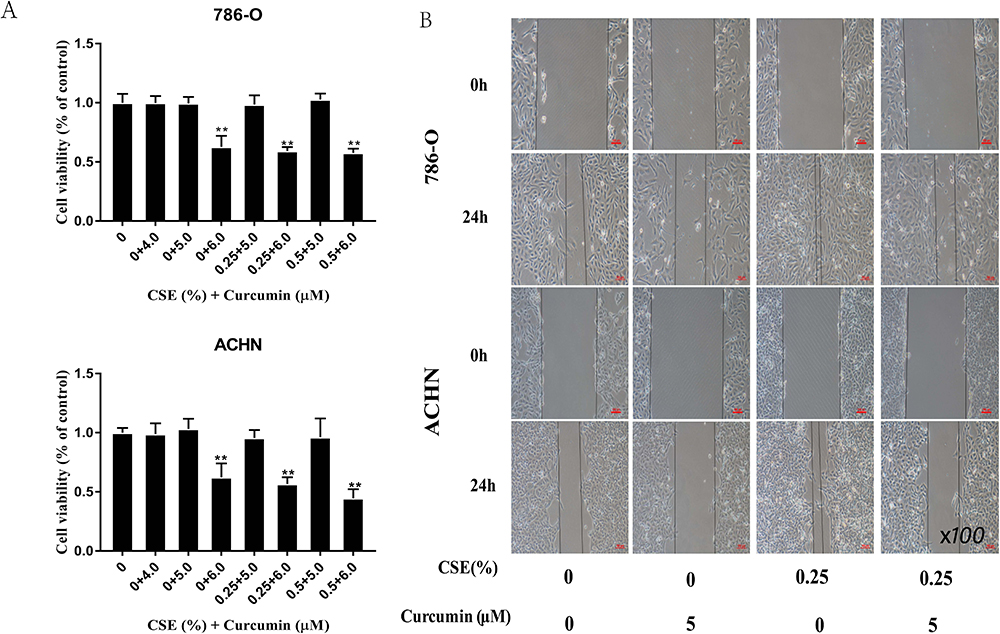 |
Figure 6 Curcumin reduces the EMT induced by CSE through suppressing ERK5/AP-1. (A) An MTT assay was conducted to screen a suitable concentration for subsequent experiments. Treatment with CSE at various doses under 6 µM curcumin revealed lower cell viability below 70%, which proved to be toxic to ACHN and A498 cells. (B) A wound-healing assay showing that CSE-enhanced migration was weakened by curcumin. Data are expressed in the manner of mean ± SD. **P<0.01 relative to control group. |
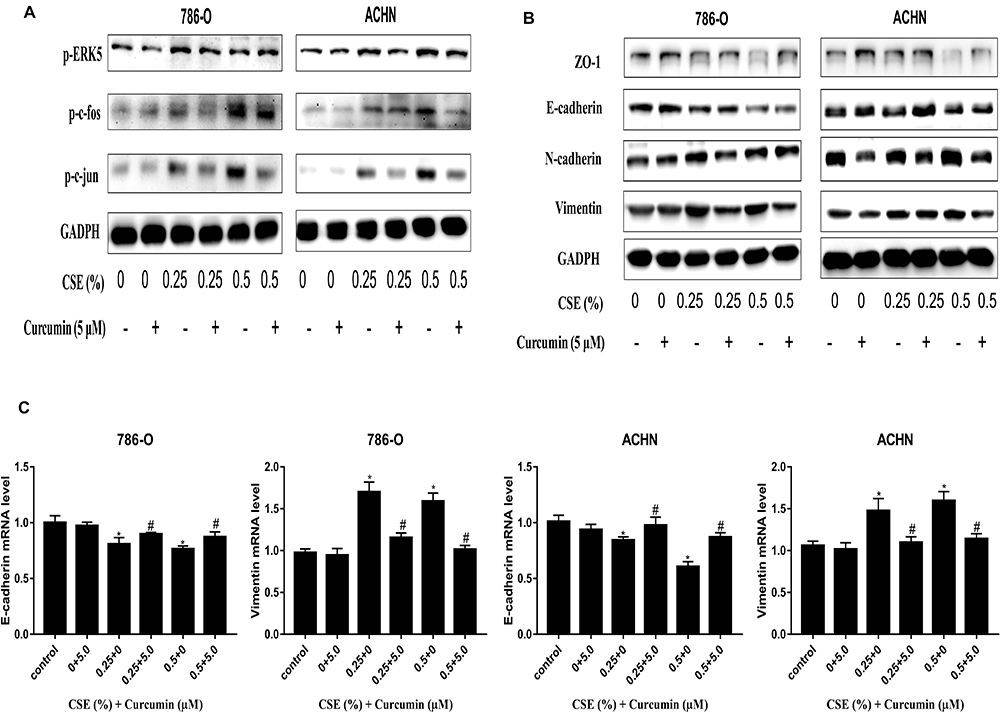 |
Figure 7 Molecular changes are tested in curcumin-treated RCC cells. (A) Curcumin inhibits ERK5/AP-1 activated by CSE, according to the activation of ERK5/AP-1 detection in curcumin-exposed cells. (B) Curcumin ameliorated the CSE-triggered reduction in ZO-1 and E-cadherin at protein levels, whereas the up-regulated N-cadherin and Vimentin protein expression, which is revealed using Western blotting. (C) Curcumin treatment increased the E-cadherin mRNA expression, but decreases Vimentin mRNA expression. All data are expressed in the manner of mean ± SD. *P<0.05, relative to control group; #P<0.05, curcumin group relative to the respective CSE group. |
Discussion
The incidence of RCC is increasing year by year and there are significantly more male patients than female patients.1,2 Cigarette smoke (CS) has been recognized as one of the most important risk factors of RCC development and numerous epidemiological studies have revealed that CS exposure can promote the initiation and progression of RCC.15,16 Tobacco contains a variety of carcinogens, and their combustion products can attack cells through various mechanisms, consequently activating signaling pathways related to each tumor in a network, leading to the rapid development of tumors.17 Although a lot of studies have found that smoking is associated with renal cancer progression, the specific molecular mechanisms involved remain unclear. The current study aimed to explore the mechanism by which smoking promotes the progression of RCC and the anti-tumor effect of curcumin.
Studies have found that EMT contributes to the metastasis of RCC through transforming the polarity of cancer cells as well as the adhesion between cells and boosting cancer aggression. Loss of epithelial cell characteristics makes it difficult for cells to adhere, and the acquisition of interstitial cell characteristics makes it easier for cells to metastasize.18 Numerous molecular alternations occur in the process of EMT, such as the loss or low E-cadherin expression as well as the up-regulated N-cadherin expression, referred to as “cadherin switch”, which is typically observed during EMT via the interruption of tight junctions and the concomitant gain of cell motility. In vitro IHC had been carried out to evaluate levels of N-cadherin, E-cadherin, Vimentin, as well as ZO-1 EMT markers in RCC specimens, proving that they correlated with the prognosis of patients with RCC.18,19 Yu et al demonstrated that EMT caused epithelial cells to significantly change their cell morphology, and increase their proliferation and invasion in human normal bladder epithelial cells and mouse models.9 Giordano et al20 found that EMT-inducing factors not only induced the occurrence of EMT in breast cancer cells but also enhance their stem cell characteristics. EMT plays a vital role during tumor metastasis and occurrence. Regulating EMT might become a new strategy to treat cancer recurrence and metastasis in the future. In consistence with previous results, our study also suggested that exposure to CSE induced EMT within 786-O and ACHN cells, manifested by changes in the cell phenotype and EMT marker levels.
In RCC tissue, we found that the tumor size, tumor stage, and tumor grade in smoking patients were slightly enhanced relative to than those for non-smoking ones. However, the differences were not statistically significant. At the same time, EMT-related markers expression would be different between smoking group and non-smoking group using both Western blotting and IHC detection. Combined with the results of the in vitro cell experiments, we believe that CS induces EMT of RCC cells, and that EMT exerts a vital part during smoking-induced RCC progression. Nevertheless, the specific regulatory mechanisms remain to be further studied. The current study was a single-center retrospective study with limited clinical cases, no long-term patient follow-up, and a lack of follow-up data, such as tumor-specific survival and overall survival. However, the relationship between smoking and EMT has been studied clinically. Cases have been presented, and preliminary findings have revealed the induction of EMT in RCC, which is in consistent with previous in vitro cell experiments.
As suggested in a prior research, ERK5 plays a vital part within CSE or benzidine-induced EMT in bladder tumor cells,7,12 yet it remains unclear about ERK5 effect on the development of RCC promoted by CSE. Numerous studies have reported the regulation of the EMT signaling pathway by, for instance, ERK5, MAPK, nuclear factor kappa B (NF-κB), and phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K).7,21,22 In primary RCC, TGFB1 mRNA (encoding transforming growth factor beta 1) was detected, indicating that the stimulation of EMT in RCC by TGFB1 induces and contributes to tumor growth.23 Laura et al found that high levels of ERK5 correlated with stages associated with worse prognosis in RCC.13 Different from other ERKs, ERK5 can activate the transcription activity. Specifically, its special structure allows ERK5 to regulate downstream molecule transcription by two manners, either via phosphorylation or transcriptional boosting.24 In this study, the 786-O and ACHN cell lines were subjected to CSE exposure for a 6-day period, which induced a marked increase in p-ERK5 in a CSE dose-dependent manner, indicating that CSE up-regulation exerted certain effect on the activity of ERK5. Additionally, CSE enhanced p-c-fos and p-c-jun levels. An ERK5 inhibitor (XMD8-92) could remarkably reverse EMT induced by CSE, which was shown through the reduced p-c-jun and p-c-fos levels after inhibiting ERK5. However, the present study did not reveal the exact mechanism of ERK5 in regulating the activation of AP-1, which requires further investigation.
Curcumin is a natural compound with great application prospects, which is used in the treatment of malignancy. Besides, it can modulate numerous signaling pathways, such as transcriptional, signal transduction, and adhesion.4,12 Evidence suggests that curcumin can regulate different signaling pathways or cytokines to achieve anti-tumor effects, and could be used as an effective adjuvant for cisplatin to regulate the erythroid 2 like 2 (NRF2), nuclear factor, as well as signal transducer and activator of transcription 3 (STAT3) signaling within humans to treat squamous cell carcinoma.25 According to Woo et al, curcumin inhibited the MAPK signal transduction pathways within astroglioma cells in humans.26 Moreover, Sun et al also demonstrated that curcumin repaired the EMT induced by benzidine within phenotype and genotype levels.21 Our results suggested that curcumin reversed the CSE-enhanced invasion and migration abilities of RCC cells. Additionally, activating ERK5/AP-1 and EMT markers might be inhibited via curcumin, as assessed by detecting the activation of ERK5/AP-1 in the case of cell exposure to curcumin. Therefore, we believe that curcumin attenuated EMT induced by CSE through suppressing the ERK5/AP-1 signaling.
Conclusion
To conclude, ERK5 is found to have a positive part within EMT progression of smoking-promoted renal cell carcinoma. In addition, curcumin is related to the smoking-associated renal malignancies. The above results further clarify the molecular mechanisms of smoking-promoted renal cell carcinoma progression, contributing to formulating the novel preventing and interventional strategies.
Data Sharing Statement
All data analyzed during this study are included in this manuscript.
Ethical Approval and Consent to Participate
This work gained approval from the Ethics Committee of the Second Affiliated Hospital of Anhui Medical University. Written informed consent was obtained from all participants included in the study.
Funding
The current study was funded by the Key Projects of Natural Science Research in Anhui Colleges and Universities (KJ2018A0209) and Natural Science Foundation of Anhui Province (2008085MH290). The funder did not play any role in the study design, data collection, data analysis, data interpretation, writing of the manuscript or decision to publish results.
Disclosure
The authors declare that they have no conflicts of interest for this work.
References
1. Sun M, De Velasco G, Brastianos PK, et al. The development of brain metastases in patients with renal cell carcinoma: epidemiologic trends, survival, and clinical risk factors using a population-based cohort. Eur Urol Focus. 2019;5(3):474–481. doi:10.1016/j.euf.2017.12.007
2. Sanchez A, Allard CB, Wilson KM. Epidemiology of Renal Cell Carcinoma. 2017.
3. Chen T, You Y, Jiang H, et al. Epithelial-mesenchymal transition (EMT): a biological process in the development, stem cell differentiation, and tumorigenesis. J Cell Physiol. 2017;232(12):3261–3272. doi:10.1002/jcp.25797
4. Liang Z, Wu R, Xie W, et al. Curcumin suppresses MAPK pathways to reverse tobacco smoke-induced gastric epithelial-mesenchymal transition in mice. Phytother Res. 2015;29(10):1665–1671. doi:10.1002/ptr.5398
5. Liang Z, Xie W, Wu R, et al. Inhibition of tobacco smoke-induced bladder MAPK activation and epithelial-mesenchymal transition in mice by curcumin. Int J Clin Exp Pathol. 2015;8(5):4503–4513.
6. Peng Z, Wang CX, Fang EH, et al. Role of epithelial-mesenchymal transition in gastric cancer initiation and progression. World J Gastroenterol. 2014;20(18):5403–5410. doi:10.3748/wjg.v20.i18.5403
7. Sun X, Zhang T, Deng Q, et al. Benzidine induces epithelial-mesenchymal transition of human bladder cancer cells through activation of ERK5 pathway. Mol Cells. 2018;41(3):188–197. doi:10.14348/molcells.2018.2113
8. Deng Y, Lei T, Li H, et al. ERK5/KLF2 activation is involved in the reducing effects of puerarin on monocyte adhesion to endothelial cells and atherosclerotic lesion in apolipoprotein E-deficient mice. Biochim Biophys Acta. 2018;1864(8):2590–2599. doi:10.1016/j.bbadis.2018.04.021
9. Yu D, Geng H, Liu Z, et al. Cigarette smoke induced urocystic epithelial mesenchymal transition via MAPK pathways. Oncotarget. 2017;8(5):8791–8800. doi:10.18632/oncotarget.14456
10. Zhao L, Zhang T, Geng H, et al. MAPK/AP-1 pathway regulates benzidine-induced cell proliferation through the control of cell cycle in human normal bladder epithelial cells. Oncol Lett. 2018;16(4):4628–4634. doi:10.3892/ol.2018.9155
11. Zhao L, Geng H, Liang ZF, et al. Benzidine induces epithelial-mesenchymal transition in human uroepithelial cells through ERK1/2 pathway. Biochem Biophys Res Commun. 2015;459(4):643–649. doi:10.1016/j.bbrc.2015.02.163
12. Liu Z, Liu J, Zhao L, et al. Curcumin reverses benzidine-induced epithelial-mesenchymal transition via suppression of ERK5/AP-1 in SV-40 immortalized human urothelial cells. Int J Oncol. 2017;50(4):1321–1329. doi:10.3892/ijo.2017.3887
13. Arias-Gonzalez L, Moreno-Gimeno I, Del Campo AR, et al. ERK5/BMK1 is a novel target of the tumor suppressor VHL: implication in clear cell renal carcinoma. Neoplasia. 2013;15(6):649–659. doi:10.1593/neo.121896
14. Geng S, Wang S, Zhu W, et al. Curcumin attenuates BPA-induced insulin resistance in HepG2 cells through suppression of JNK/p38 pathways. Toxicol Lett. 2017;272:75–83. doi:10.1016/j.toxlet.2017.03.011
15. Boudreau DM, Yu O, Johnson J. Statin use and cancer risk: a comprehensive review. Expert Opin Drug Saf. 2010;9(4):603–621. doi:10.1517/14740331003662620
16. Tsivian M, Moreira DM, Caso JR, et al. Cigarette smoking is associated with advanced renal cell carcinoma. J Clin Oncol. 2011;29(15):2027–2031. doi:10.1200/JCO.2010.30.9484
17. Cumberbatch MG, Rota M, Catto JW, et al. The role of tobacco smoke in bladder and kidney carcinogenesis: a comparison of exposures and meta-analysis of incidence and mortality risks. Eur Urol. 2016;70(3):458–466. doi:10.1016/j.eururo.2015.06.042
18. Sugimoto M, Kohashi K, Itsumi M, et al. Epithelial to mesenchymal transition in clear cell renal cell carcinoma with rhabdoid features. Pathobiology. 2016;83(6):277–286. doi:10.1159/000445752
19. Piva F, Giulietti M, Santoni M, et al. Epithelial to mesenchymal transition in renal cell carcinoma: implications for cancer therapy. Mol Diagn Ther. 2016;20(2):111–117. doi:10.1007/s40291-016-0192-5
20. Giordano A, Gao H, Anfossi S, et al. Epithelial-mesenchymal transition and stem cell markers in patients with HER2-positive metastatic breast cancer. Mol Cancer Ther. 2012;11(11):2526–2534. doi:10.1158/1535-7163.MCT-12-0460
21. Sun X, Deng QF, Liang ZF, et al. Curcumin reverses benzidine-induced cell proliferation by suppressing ERK1/2 pathway in human bladder cancer T24 cells. Exp Toxicol Pathol. 2016;68(4):215–222. doi:10.1016/j.etp.2015.12.003
22. Deng QF, Sun X, Liang ZF, et al. Cigarette smoke extract induces the proliferation of normal human urothelial cells through the NF-kappaB pathway. Oncol Rep. 2016;35(5):2665–2672. doi:10.3892/or.2016.4623
23. Liu Y, Hu H, Wang K, et al. Multidimensional analysis of gene expression reveals TGFB1I1-induced EMT contributes to malignant progression of astrocytomas. Oncotarget. 2014;5(24):12593–12606. doi:10.18632/oncotarget.2518
24. Wang X, Pesakhov S, Harrison JS, et al. ERK5 pathway regulates transcription factors important for monocytic differentiation of human myeloid leukemia cells. J Cell Physiol. 2014;229(7):856–867. doi:10.1002/jcp.24513
25. Fetoni AR, Paciello F, Mezzogori D, et al. Molecular targets for anticancer redox chemotherapy and cisplatin-induced ototoxicity: the role of curcumin on pSTAT3 and Nrf-2 signalling. Br J Cancer. 2015;113(10):1434–1444. doi:10.1038/bjc.2015.359
26. Woo MS, Jung SH, Kim SY, et al. Curcumin suppresses phorbol ester-induced matrix metalloproteinase-9 expression by inhibiting the PKC to MAPK signaling pathways in human astroglioma cells. Biochem Biophys Res Commun. 2005;335(4):1017–1025. doi:10.1016/j.bbrc.2005.07.174
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.