Back to Journals » Journal of Pain Research » Volume 12
Could epigenetics help explain racial disparities in chronic pain?
Authors Aroke EN ![]() , Joseph PV
, Joseph PV ![]() , Roy A
, Roy A ![]() , Overstreet DS, Tollefsbol TO, Vance DE
, Overstreet DS, Tollefsbol TO, Vance DE ![]() , Goodin BR
, Goodin BR ![]()
Received 23 October 2018
Accepted for publication 4 January 2019
Published 18 February 2019 Volume 2019:12 Pages 701—710
DOI https://doi.org/10.2147/JPR.S191848
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Katherine Hanlon
Edwin N Aroke,1 Paule V Joseph,2 Abhrarup Roy,2 Demario S Overstreet,3 Trygve O Tollefsbol,4 David E Vance,1 Burel R Goodin3
1School of Nursing, University of Alabama at Birmingham, Birmingham, AL, USA; 2Sensory Science and Metabolism Unit (SenSMet), Division of Intramural Research, National Institute of Nursing Research, National Institute of Health, DHHS, Bethesda, MD, USA; 3Department of Psychology, College of Arts and Sciences, University of Alabama at Birmingham, Birmingham, AL, USA; 4Department of Biology, College of Arts and Sciences, University of Alabama at Birmingham, Birmingham, AL, USA
Abstract: African Americans disproportionately suffer more severe and debilitating morbidity from chronic pain than do non-Hispanic Whites. These differences may arise from differential exposure to psychosocial and environmental factors such as adverse childhood experiences, racial discrimination, low socioeconomic status, and depression, all of which have been associated with chronic stress and chronic pain. Race, as a social construct, makes it such that African Americans are more likely to experience different early life conditions, which may induce epigenetic changes that sustain racial differences in chronic pain. Epigenetics is one mechanism by which environmental factors such as childhood stress, racial discrimination, economic hardship, and depression can affect gene expression without altering the underlying genetic sequence. This article provides a narrative review of the literature on epigenetics as a mechanism by which differential environmental exposure could explain racial differences in chronic pain. Most studies of epigenetic changes in chronic pain examine DNA methylation. DNA methylation is altered in the glucocorticoid (stress response) receptor gene, NR3C1, which has been associated with depression, childhood stress, low socioeconomic status, and chronic pain. Similarly, DNA methylation patterns of immune cytokine genes have been associated with chronic stress states. Thus, DNA methylation changes may play an essential role in the epigenetic modulation of chronic pain in different races with a higher incidence of epigenetic alterations contributing to more severe and disabling chronic pain in African Americans.
Keywords: chronic pain, epigenetics, racial health disparities, epigenomics, DNA methylation, stress
Introduction
In the United States, chronic pain is a significant public health problem that affects over 100 million people and produces an estimated annual cost of about $635 billion.1,2 Despite significant improvements in health care access and chronic pain management, African Americans continue to experience more frequent, severe, and disabling chronic pain than do non-Hispanic Whites.3,4 For instance, a National Health Interview Survey of over 85,000 participants found that African Americans reported more severe joint pain and activity limitation due to arthritis than did non-Hispanic Whites.5 Also, African Americans are more likely to report more severe back pain, and more disability outcomes than White Americans.4 Even though African Americans use more pain-related coping strategies (such as, praying, hoping, diverting attention, and catastrophizing) than Whites, racial differences persist.6 Frequently cited explanations for disparities in chronic health conditions include differences in socioeconomic status (SES),3,7,8 education,4 childhood trauma,3,9 accumulated stress,10 racial discrimination,11,12 and neighborhood crime3,8 as well as mental disorders such as anxiety and depression.13,14 Thus, it can be argued that the experience of chronic pain extends to an individual’s interaction with his or her psychosocial environment.
Recent reviews have been published on the topic of epigenetics and chronic pain.15,16 However, we are not aware of any articles that addressed the inter-relationship between race, lived experience, epigenetic changes, and chronic pain disparities. The purpose of this narrative review is to explore epigenetics as a mechanism by which differential environmental exposure could explain racial differences in chronic pain. To achieve this premise, first, an overview of the fundamental concept of epigenetic mechanisms is presented. Second, the relationship between environmental exposure and epigenetic changes that cause and sustain chronic pain is presented within the context of chronic stress. A conceptual model is provided to depict the relationship between chronic stress due to early life adversities, racial discrimination, SES, depression, and DNA methylation patterns that cause and sustain chronic pain (Figure 1). Third, hypothetical case comparisons are provided to highlight the relationship between racial differences in environmental exposure and patterns of epigenetic changes (Figure 2). Finally, implications for clinical practice and research are provided. Undoubtedly, there are sex differences in chronic pain sensitivity and perception, and it is possible that racial disparities in chronic pain are actually driven by sexual dimorphic differences rather than differences due to the social construct of race or a combination. Several authors have published excellent review articles on sex differences and clinical pain,17–20 and detailed discussion of sex difference and chronic pain is beyond the scope of this paper.
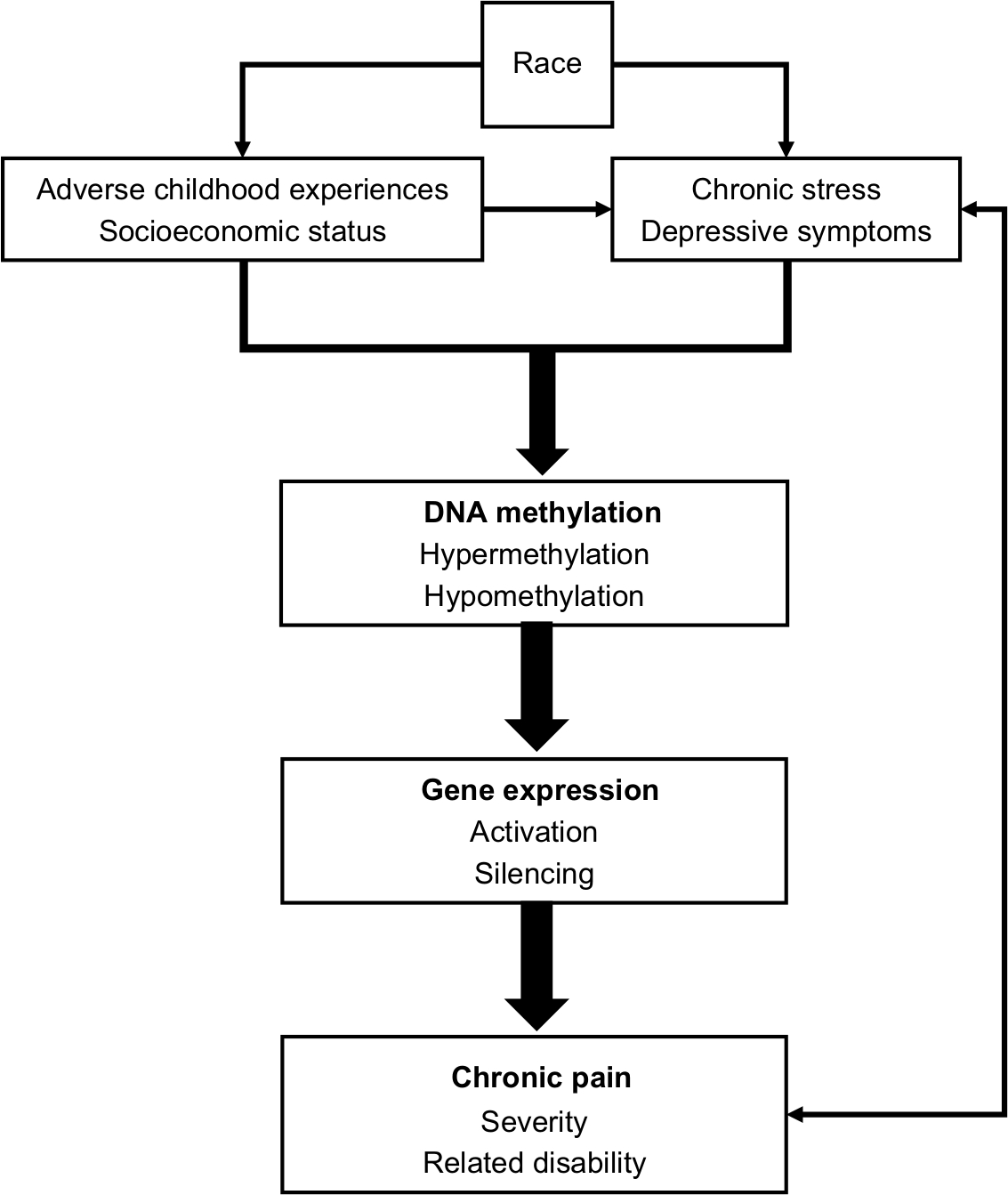 | Figure 1 The conceptual model of the inter-relationship between life experiences, chronic stress, DNA methylation patterns, and chronic pain. |
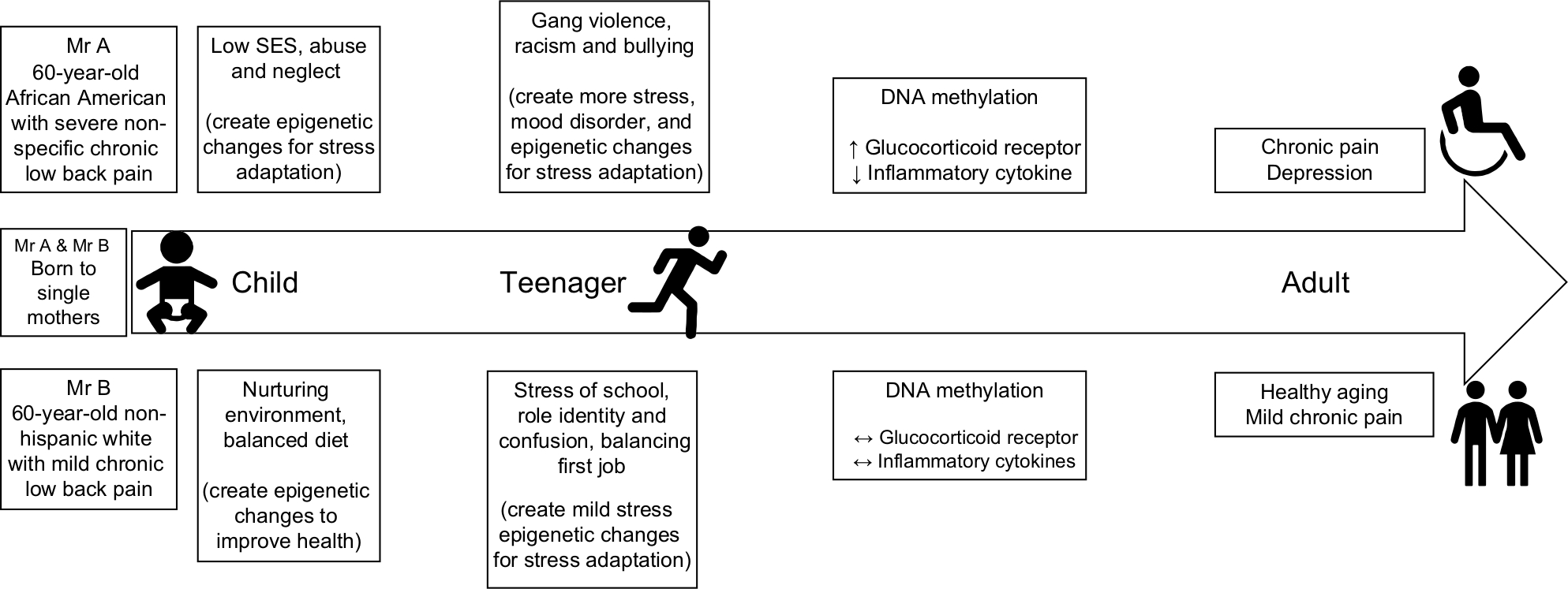 | Figure 2 Hypothetical cases of epigenetic changes in chronic pain. Abbreviation: SES, socioeconomic status. |
An overview of epigenetic mechanisms
Epigenetics refers to heritable changes in chromatin and gene expression without alterations in the underlying DNA sequence.21 In other words, the genotype of an organism remains intact while the outward expression (phenotype) is altered. In this regard, nutrition, stress, and lifestyle choices are some factors that contribute to epigenetic changes22 with non-Mendelian inheritance patterns.23 At the molecular level, three major types of mechanisms drive epigenetics: non-coding RNAs (ncRNAs), histone modifications such as acetylation, and DNA methylation.23
The majority of human genetic material does not code for proteins and are referred to as ncRNAs. Initially thought to be junk RNA, ncRNAs mediate intracellular processes and can influence gene expression and silencing.24 Many types of ncRNAs have been identified based on several attributes including regions of the human genome involved and size of the sequence such as microRNAs (miRNAs) and long non-coding RNAs (lncRNAs).24,25 For example, miRNAs are approximately 22-nucleotide double-stranded RNAs that interfere with transcribed messenger RNA (mRNA) in the cytoplasm, while lncRNAs are over 200 nucleotides long and may encode short peptides.24 The mechanism by which ncRNAs silence gene expression is known as RNA interference, which is commonly associated with post-transcriptional modification of mRNA.25
Histone acetylation does not directly affect mRNA. Instead, it remodels chromatin. This process of chromatin remodeling is catalyzed by histone acetyltransferase, an enzyme that typically adds acetyl groups to lysine groups of the histone protein. This causes the chromatin to unravel, which stimulates gene expression.23 By neutralizing the positive charge on the histone tail, histone acetylation opens chromatin, giving the transcriptional machinery easier access to the DNA. Acetyl groups are commonly added to lysine residues such as H3K9.26 Generally, acetylation of histones is associated with gene expression, and histone deacetylation is associated with gene repression. Also, histone modification can influence DNA methylation and vice versa.
DNA methylation is an epigenetic mechanism that involves direct chemical modification of DNA.27 Methylation of DNA is catalyzed by a family of DNA methyltransferases, which add a methyl group to cytosine. In humans, the majority of the methyl groups are added to a cytosine that precedes guanine in a cytosine-guanine dinucleotide rich region known as CpG islands. Interestingly, many CpG sites are found in the 5’ regulatory (promoter) region of most genes, where they provide a transcription binding site and enhance the accessibility of the DNA for transcription. This is a dynamic (reversible) process whereby methylation of the CpG island interferes with the binding of transcription factors and leads to stable gene silencing.27 The process of gene regulation through DNA methylation is sensitive to environmental input and life experiences. Landmark work by Weaver et al showed that exposure to a nurturing environment resulted in hypomethylation of the glucocorticoid gene and better adaptation to stress, while an uncaring environment resulted in higher levels of DNA methylation.28 Given the dynamic nature of DNA methylation, reversal of the environmental conditions resulted in a reversal of the epigenetic changes.29 Given that most studies of epigenetic changes in chronic pain have focused on DNA methylation, the rest of this article will focus on DNA methylation changes.
Lived experience and epigenetic regulation
The United States is a racially and ethnically diverse country. However, individuals of different racial backgrounds have different life experiences, which influence their health and health outcomes.4 In the postgenomic era, the debate over race as biological vs social entity continues.30,31 For this review, the term race refers to the social construct that influences health and lived experiences. This view of race as a lived experience is primarily associated with skin color.32 Informed by the social determinants of health, we assert that the lived experience of African Americans tends to be more stressful than non-Hispanic Whites,33,34 which can be reflected epigenetically and increase the risk that African Americans will suffer from more debilitating chronic pain. In the following sections, we discuss racial differences in life experiences such as adverse childhood experiences (ACEs), SES, chronic stress, and depression, which are contributing factors to changes in DNA methylation patterns that may explain racial differences in chronic pain. Figure 1 is a conceptual depiction of this inter-relationship.
ACEs
In the United States, African American children experience more ACEs than children of other races/ethnicities.35,36 Following the landmark study by Felitti et al,37 many studies have documented relationships between ACEs and neurobehavioral, cognitive, and psycho-emotional chronic illnesses such as chronic pain later in life.38–41 Using data from the ACEs study, Anda et al reported a significant positive relationship between the number of ACEs and risk of frequent headaches in a dose-response fashion. An adult with five or more ACEs had more than a 2-fold risk of frequent headaches than adults with no ACEs.39 More recently, Nelson et al examined the incidences and impact of ACEs on the psychological and pain-related distress among youth (between 9 and 19 years) with chronic pain. Over 80% of the youth with chronic pain had experienced at least one ACE. There was a significant relationship between ACE exposure, depression, anxiety, and fear of pain.38 The fact that associations are found between ACEs and psychosocial factors among youth as young as 9 years old suggest that the factors mediating the body’s response to ACEs (stress response) could be happening much earlier than the ACE itself. There is considerable evidence showing that prenatal exposure to maternal psychosocial stress influences the neurocognitive development of children.42–44
Emerging evidence suggests that from the first possible exposure to stress – in utero – organisms may be susceptible to epigenetic programming that persists into adult life and can be transmitted from one generation to the next.45,46 The Dutch Hunger Winter birth epidemiological study compared the DNA methylation patterns of siblings exposed to famine against non-exposed siblings. They found that siblings exposed to famine showed differential methylation of several genes including IGF2 and IL-10.47,48 Contrary to other studies, the Dutch study did not find any correlation between DNA methylation status of the genes in the hypothalamic-pituitary-adrenal (HPA) axis (for example, NR3C1) and exposure to stress.
In a meta-analysis, Palma-Gudiel et al examined primary evidence linking prenatal stress to DNA methylation of the NR3C1 gene. They found a significant correlation between prenatal exposure to stress and DNA methylation at specific sites in the promoter region of the human glucocorticoid receptor gene (NR3C1).49 The same groups performed a critical review of the link between early life stress and DNA methylation of the NR3C1 gene and concluded that early life adversity is associated with hypermethylation at several CpG sites in the promoter region of the NR3C1 gene.50 DNA methylation of other genes that affect the functioning of the HPA and are associated with early life adversity include 11b-HSD2 and FKBP5 genes, which modulate the glucocorticoid receptor activity in response to a stressor.51,52 Collectively, these studies suggest that DNA methylation may modulate the relationship between the stress of early life adversity, the functioning of the HPA, and adulthood conditions. Also, the social construct of race makes it such that Americans of some racial backgrounds are more likely to experience different early life conditions, which may induce epigenetic changes that sustain racial differences in chronic pain.
SES
SES and chronic stress are firmly interweaved. The construct of SES is also categorized by education level, financial security, and social status. SES is known to be heavily correlated with a plethora of psychological and environmental conditions that contribute to the disproportionate burden of chronic stress.53 Recent studies suggest that childhood SES (not adulthood SES) may be a stronger determinant of adult health status. Lower SES or socioeconomic deprivation in early life is associated with chronic physiologic stress that affects health outcomes later in life.54 Children growing up in lower SES neighborhoods are at increased risk of exposure to crime, economic hardship, and pollution, which negatively influences their adult health.55 African Americans are more likely to experience childhood crime, discrimination, racism, and live in a lower SES neighborhood, which may contribute to chronic biopsychosocial stress.3 Using proton magnetic resonance spectroscopy, McLean et al examined the relationship between early life SES, chronic stress, and N-acetyl aspartate (a myelin lipid that plays a critical role in oligodendrocytes’ synthesis) in the hippocampus of randomly selected healthy men. They reported that lower SES in early life was associated with greater chronic stress and reduced N-acetyl aspartate level.56 These findings suggest that early life lower SES and associated chronic stress could have a cumulative physiological effect on the functioning of the white matter that persists into adulthood.
Epigenetic modification (such as DNA methylation) is another mechanism by which early life SES may affect adulthood chronic disease. Using a life course approach, Needham et al examined the relationship between life course of SES and DNA methylation in 18 genes related to stress and inflammatory response. They found a significant relationship between childhood SES and DNA methylation of both stress-related (AVP, BDNF, FKBP5, and OXTR) and inflammatory (CCL1, CD1D, and F8) genes.57 Although the pattern of methylation (hypomethylation vs hypermethylation) varied by genes and site (promoter vs non-promoter CpG site), their findings suggest that differences in childhood SES may correlate with different epigenetic modifications, which in turn have been associated with chronic diseases.
Chronic stress and DNA methylation
Chronic stress and associated maladaptive processes play a crucial role in modulating life experiences (for example, ACEs and low SES) and many chronic diseases such as depression and chronic pain.53,58 During the evolutionary process, humans developed a complex adaptive process to ensure survival when faced with an actual or perceived life-threatening acute stressor.54 This adaptive process typically involves the activation of multiple systems including the autonomic, endocrine, and immune systems. One pathway by which these multiple systems are activated to mount a concerted response to a stressor is the HPA axis pathway. Upon activation, the HPA axis releases glucocorticoid hormones (and other mediators such as catecholamines and inflammatory cytokines), which modulate the stress response through negative feedback.59,60 Overactivation of this system by the continuous and sustained release of the stress response mediators can result in a maladaptive response, which often results in feelings of being overwhelmed that characterizes chronic stress.58–61 For instance, chronic stress such as the stress of ACEs, racism/racial discrimination, low SES, and depression have been associated with alterations in the neuroendocrine and immune system response to stimuli.45,59 Even more alarming is the fact that the adverse effects of chronic stress can induce changes in global DNA methylation and gene expression, which increases susceptibility to adverse health outcomes and diseases such as depression.46,62
Racial differences in depression
Chronic pain, often comorbid with depression and alterations in the HPA axis, plays an essential role in the pathophysiology of both depression and chronic pain.63 As a result, some researchers have suggested the pain cannot be fully understood without considerations for depression.64 Theoretical and empirical evidence suggests that dysfunction in the HPA and immune systems may play a significant role in the depression-pain dyad.64,65 Among patients with chronic pain, African Americans report higher levels of depressive symptoms compared to Whites.66–68 Thus, we suggest that disparities in pain cannot be fully understood without considerations of the emotional aspects of pain (depression). Although no studies have explicitly examined the inter-relationship among chronic stress, depressive symptoms, chronic pain, and epigenetic modifications, emerging evidence suggests that these chronic disorders may share common DNA methylation patterns.50,69
Chronic pain, DNA methylation, and gene expression
Growing evidence suggests that epigenetic modifications are mechanisms through which environmental exposure interacts with an individual’s genetic makeup to determine a lifetime risk for chronic pain.70 In this section, we summarize current evidence of the impact of epigenetic changes on chronic low back pain and fibromyalgia.
Over 70%–80% of adults experience at least one episode of back pain in their lifetime. Chronic low back pain refers to any persistent pain in the lower spine, lasting more than 3 months. Common causes of chronic low back pain can be attributed to injury, or disease or non-specific factors, and the pain is usually described as axial or radicular pain or both axial and radicular. Radicular back pain usually radiates to the lower extremity through nerves; axial pain is localized to the spine and evoked spontaneously or with movement. Tajerian et al investigated whether DNA methylation down-regulated gene expression and precipitated chronic low back pain in mice and humans.71 They treated 1-year-old mice with a DNA demethylating agent (5-azacytidine) and observed that demethylation of the SPARC gene promoter region resulted in a >4-fold increase in SPARC mRNA. Also, SPARC methylation correlated significantly with cold pain sensitivity. They also investigated the role of SPARC methylation in chronic low back pain. Compared with pain-free controls, patients with chronic low back pain had higher levels of methylation in the promoter region of the SPARC gene.71 Using a two-stage epigenome-wide association design, Bell et al (2014) compared the methylation patterns of 50 monozygotic twins who were discordant for pain sensitivity and 50 unrelated individuals. They reported that increased DNA methylation levels at the promoter region of the TRPA1 gene were associated with decreased gene expression and pain sensitivity.72 More recently, Sukenaga et al examined DNA methylation levels of the TRPA1 gene (cg12668482, chr8: 72987762) in 12 patients with chronic neuropathic and low back pain. They found that higher pain scores correlated with higher DNA methylation levels at the CpG Island of the TRPA1 gene.73 We agree with their conclusion that DNA methylation of the TRPA1 gene might be a mediator between chronic pain, mood disorders such as depression, and anxiety.73
Chronic widespread pain and mood disturbances characterize fibromyalgia pain. It has been associated with disturbances in the HPA axis. The glucocorticoid receptor gene (NR3C1) is crucial to the maintenance of homeostasis and response to stress. Macedo et al compared the sensitivity and expression of glucocorticoid receptor and mineralocorticoid receptor genes in 27 women with fibromyalgia and 27 sex-matched controls. Compared with the healthy controls, patients with fibromyalgia had significantly lower mRNA expression of both glucocorticoid receptor and mineralocorticoid receptor.74 A more recent study compared the DNA methylation patterns and chromosomal instability of ten women with fibromyalgia pain and 42 age-matched healthy controls using an epigenome-wide approach. Overall, compared to controls, women with fibromyalgia pain had significantly different DNA methylation patterns in many genes involved in neural development and maturation such as brain neurotrophic factors, N-acetyltransferase 15, and protein kinases.75
Case comparison
Initially, it was thought that once in the committed stem cells, DNA methylation remains mostly unchanged through the life course of an individual. However, there is growing evidence that environmental factors influence DNA methylation (epigenome) and can lead to adulthood disorders.45,48,50,62 Similarly, the relationship between SES and racial disparities in health has been well documented. However, after controlling for SES other lived experiences such as early life adversity, the burden of racism and discrimination, and exposure to psychosocial stressors influence racial differences in adulthood diseases and health outcomes.34 Figure 2 depicts the life course of two hypothetical individuals Mr A and Mr B.
Mr A is a 60-year-old African American male, who is wheelchair-bound due to severe non-specific chronic low back pain. He was born and raised by a single mother living in a metropolitan city, while his dad was serving a 20-year sentence in jail for possession of illegal drugs. Poverty, abuse, and neglect characterized his early childhood. At age 5 years, Mr A was physically abused by his stepfather, while his mother worked night shift as a housekeeper at the local hospital. The abuse continued for several years and ended at the age of 10 when Mr A returned from school to find his lifeless mother who had been killed by a member of the neighborhood gang. Young Mr A moved to live with his grandmother in an affluent neighborhood. In middle and high school, he experienced racial discrimination and bullying from classmates, and was diagnosed with attention deficit hyperactivity disorder and depression. He eventually graduated from college and began working at the local superstore as branch manager. As a manager, Mr A trained many employees, many of whom went on to become his supervisor and credit him for their success. Two days after his 50th birthday, Mr A started complaining of non-specific back pain. Five years later, the company was sold, and he was laid-off as part of the restructuring by the new owners. At the age of 60, Mr A lives on disability and is wheelchair-bound. His medical diagnosis includes chronic back pain, major depression, anxiety, hypertension, and mild diabetes. Mr A recently enrolled in a chronic low back pain study and his sample showed hypermethylation of the NR3C1 gene and hypomethylation of pro-inflammatory cytokines.
On the other hand, Mr B is a 60-year-old non-Hispanic White male with chronic low back pain. He was born to a single mother who worked as a nurse at the local hospital. Even though he came from a modest background, his childhood was relatively uneventful. Some of the most significant stressors in his early childhood were the death of his grandmother, balancing school work and volunteer activities through the Boys Scouts, and peer pressure. While in high school, he had minor infractions with law enforcement officers including traffic tickets and drinking alcohol as a minor. After graduating from high school, he gained employment as a railroad technician, eventually growing in the company to his current position as Assistant Manager. At the age of 52 years, he was diagnosed with chronic low back pain, a condition he attributes to many years of hard labor as a technician. He underwent a lumbar microdiscectomy and fusion, with minimal relief. He currently undergoes epidural steroid injections multiple times a year for pain control and his employer has adjusted his work schedule to accommodate his back pain. At the age of 60, he remains gainfully employed and looks forward to retirement at the age of 67. Mr B enrolled in the chronic low back study, and his DNA methylation profile showed “normal” methylation of NR3C1 and pro-inflammatory cytokine genes.
These two hypothetical exemplars highlight the fact that differential environmental exposures can result in racial differences in DNA methylation patterns of genes that contribute to chronic pain. The pathophysiology of chronic pain and the gene–environment interaction are much more complicated. Also, racial discrimination is not the only source of stress, and the environmental exposure of individuals of different racial/ethnic backgrounds is not very distinct. Other psychosocial factors, such as coping mechanisms, resilience, and sleep hygiene, are all critical considerations.
Implications for research and practice
Clinical implications
Most chronic pain conditions are challenging to treat because the exact mechanism that causes and sustains them remains mostly unknown. For nurses and other health care providers caring for patients with chronic pain, suggestions are provided in the areas of assessment, and symptom management (intervention). For patients with chronic pain, pain assessment should go beyond the frequently assessed severity, location, duration, and precipitating factors. Although these factors provide valuable information, they do not provide a full assessment of the ongoing pathophysiologic disorder. Given the strong association between mood disorders and chronic pain, a thorough pain assessment should include a brief assessment of depression (for example, Beck’s depression inventory) and anxiety (for example, Hamilton anxiety rating scale), family history, coping, and perceived stressors. If concerns arise, a detailed assessment of childhood trauma (for example, Childhood Trauma Questionnaire76) and personal impact of childhood adversity can be administered during a clinic visit. Patients with inconclusive or positive results for psychosocial dysfunction should be referred to a psychologist for holistic management.
Management of chronic pain syndromes should include a comprehensive approach to address associated psychosocial factors. First, every effort should be made to relieve pain and prevent further distress. This may include the use of pharmacological interventions such as non-steroidal anti-inflammatory drugs, acetaminophen, opioids, and mood stabilizers. Second, psychological interventions (for example, counseling) can address mood disorders and childhood adversity. Addressing childhood adversity and enacting policies to prevent chronic stress (for example, racial discrimination, crime, and violence) in some segments of the population may control the chronification of pain. Given that epigenetic changes are reversible, future intervention should be geared toward reversing the epigenetic changes (for example, using DNA methyltransferase inhibitors and HDAC inhibitors) associated with chronic pain syndromes.77 Because the focus on epigenetic changes (DNA methylation) is relatively new to chronic pain and chronic pain differences, this also represents an emerging research topic.
Research implications
The evidence supporting the role of epigenetic changes in chronic pain is increasing; however, the number of clinical studies investigating the role of epigenetic changes in various chronic pain syndromes remain sparse. For investigators, this has implications for pain research, disparity studies in pain, and interventional studies in pain management. First, given the strong association between childhood life experiences and adulthood disorders, studies of an adulthood disorder such as chronic pain should utilize an epigenetic/epigenomic approach to account for the gene–environment interaction. Such an approach can help increase our understanding of mechanisms that cause and sustain chronic pain.
Second, as alluded to earlier, compared with non-Hispanic Whites, African Americans experience more severe and disabling chronic pain, and the cause of the racial differences remain unknown. However, we know that African Americans are relatively more likely to experience early life adversity, chronic stress, and adverse environmental exposure, all of which are known to affect adulthood health adversely (for example, depression and hypertension). Moreover, all of the factors have been shown to induce epigenetic changes associated with adulthood disorders. Thus, studies of racial differences using an epigenetic approach are needed.
Lastly, emerging epigenetic literature points to potential mechanisms for novel therapeutics targeting epigenetic changes.77 Studies investigating these therapeutic interventions are needed. Also, intervention studies geared toward reversing the effects of or addressing early life adversity may help to reduce the racial/ethnic differences in chronic pain.
Conclusion
Racial differences in adulthood disorders such as chronic pain may result from a lifetime of adversity, chronic stress, poor coping, racial discrimination, this along with genetic makeup all interact to influence adaptation continually. Over time, these interactions result in epigenetic changes such as hypermethylation of the NR3C1 gene, which suppresses the expression of the glucocorticoid receptor. Because the HPA is crucial in adaptation to psychological and physical stress, different methylation patterns of this gene and others may explain differences in chronic pain. Racial disparities in chronic pain represent a complex interaction of racism, environmental exposure, and genetic makeup. Epigenetics represents one mechanism that could help explain how these factors interact to sustain such differences.
Acknowledgments
The authors would like to thank Dr Joan Austin for her thoughtful comments and edits. The American Association of Nurse Anesthetists (AANA) Foundation Post-Doctoral Fellowship (2018-FS-4) provided support to ENA. The University of Alabama at Birmingham Health Services Research Training Program; T32HS013852 (DSO) and NIH/NIMHD Grant R01MD010441 (BRG) provided financial support for this research. The National Institutes of Nursing Research, Division of Intramural Research provided support to PVJ under award 1ZIANR000035-01. PVJ is supported by the National Institute of Nursing Research (1ZIANR000035-01), the Office of Workforce Diversity and the National Institutes of Health Distinguished Scholars Award, and by the Rockefeller University Heilbrunn Nurse Scholar Award. NIH Intramural Research Training Award supports AR. The National Institutes of Nursing Research, is an institute in the National Institutes of Health (NIH), US Department of Health and Human Services (DHHS), Bethesda, MD.
Disclosure
The content expressed herein and the interpretation and reporting of these data are the responsibility of the authors and should not be seen as an official recommendation or interpretation of the National Institutes of Health or the University of Alabama Birmingham. The authors report no conflicts of interest in this work.
References
Institute of Medicine Committee on Advancing Pain Research C, and Education. Relieving Pain in America: A Blueprint for Transforming Prevention, Care, Education, and Research. Washington (DC): The National Academies Press; 2011. | ||
Gaskin DJ, Richard P. The Economic Costs of Pain in the United States. The Journal of Pain. 2012;13(8):715–724. | ||
Green CR, Hart-Johnson T. The association between race and neighborhood socioeconomic status in younger black and white adults with chronic pain. J Pain. 2012;13(2):176–186. | ||
Anderson KO, Green CR, Payne R. Racial and ethnic disparities in pain: causes and consequences of unequal care. J Pain. 2009;10(12):1187–1204. | ||
Bolen J, Schieb L, Hootman JM, et al. Differences in the prevalence and severity of arthritis among racial/ethnic groups in the United States, National Health Interview survey, 2002, 2003, and 2006. Prev Chronic Dis. 2010;7(3):A64. | ||
Meints SM, Miller MM, Hirsh AT, Sm M, Meints SM. Differences in pain coping between black and white Americans: a meta-analysis. J Pain. 2016;17(6):642–653. | ||
Janevic MR, McLaughlin SJ, Heapy AA, Thacker C, Piette JD. Racial and socioeconomic disparities in disabling chronic pain: findings from the health and retirement study. J Pain. 2017;18(12):1459–1467. | ||
Maly A, Vallerand AH. Neighborhood, socioeconomic, and racial influence on chronic pain. Pain Manag Nurs. 2018;19(1):14–22. | ||
Alvarez P, Green PG, Levine JD. Stress in the adult rat exacerbates muscle pain induced by early-life stress. Biol Psychiatry. 2013;74(9):688–695. | ||
Pervanidou P, Agorastosa, Kolaitis G, Chrousos G. Neuroendocrine responses to early life stress and trauma and susceptibility to disease. Eur J Psychotraumatol. 2017;8 (Suppl 4):1351218. | ||
Hoffman KM, Trawalter S, Axt JR, Oliver MN. Racial bias in pain assessment and treatment recommendations, and false beliefs about biological differences between blacks and whites. Proc Natl Acad Sci USA. 2016;113(16):4296–4301. | ||
Burgess DJ, Grill J, Noorbaloochi S, et al. The effect of perceived racial discrimination on bodily pain among older African American men. Pain Med. 2009;10(8):1341–1352. | ||
Cruz-Almeida Y, King CD, Goodin BR, et al. Psychological profiles and pain characteristics of older adults with knee osteoarthritis. Arthritis Care Res. 2013;65(11):1786–1794. | ||
Moix J, Kovacs FM, Martín A, Plana MN, Royuela A, Spanish Back Pain Research Network. Catastrophizing, state anxiety, anger, and depressive symptoms do not correlate with disability when variations of trait anxiety are taken into account. a study of chronic low back pain patients treated in Spanish pain units [NCT00360802]. Pain Med. 2011;12(7):1008–1017. | ||
Liang L, Lutz BM, Bekker A, Tao YX. Epigenetic regulation of chronic pain. Epigenomics. 2015;7(2):235–245. | ||
Denk F, McMahon SB. Chronic pain: emerging evidence for the involvement of epigenetics. Neuron. 2012;73(3):435–444. | ||
Re S, Strath LJ. Sex differences in pain responses. Curr Opin Physiol. 2018;6:75–81. | ||
Re S, Totsch SK. Sex differences in pain. J Neurosci Res. 2017;95(6):1271–1281. | ||
Mogil JS. Sex-based divergence of mechanisms underlying pain and pain inhibition. Curr Opin Behav Sci. 2018;23:113–117. | ||
Fillingim RB, King CD, Ribeiro-Dasilva MC, Rahim-Williams B, Riley JL 3rd. Sex, gender, and pain: a review of recent clinical and experimental findings. J Pain. 2009;10(5):447–485. | ||
Dupont C, Armant DR, Brenner CA. Epigenetics: definition, mechanisms and clinical perspective. Semin Reprod Med. 2009;27(5):351–357. | ||
Matboli M, Shafei A, Ali M, et al. Emerging role of nutrition and the non-coding landscape in type 2 diabetes mellitus: a review of literature. Gene. 2018;675:54–61. | ||
Huang B, Jiang C, Zhang R. Epigenetics: the language of the cell? Epigenomics. 2014;6(1):73–88. | ||
Palazzo AF, Lee ES. Non-coding RNA: what is functional and what is junk? Front Genet. 2015;6:2. | ||
Booton R, Lindsay MA. Emerging role of microRNAs and long noncoding RNAs in respiratory disease. Chest. 2014;146(1):193–204. | ||
Anacker C, O’Donnell KJ, Meaney MJ. Early life adversity and the epigenetic programming of hypothalamic-pituitary-adrenal function. Dialogues Clin Neurosci. 2014;16(3):321–333. | ||
Moore LD, Le T, Fan G. DNA methylation and its basic function. Neuropsychopharmacology. 2013;38(1):23–38. | ||
Weaver IC, Cervoni N, Champagne FA, et al. Epigenetic programming by maternal behavior. Nat Neurosci. 2004;7(8):847–854. | ||
Weaver IC, Champagne FA, Brown SE, et al. Reversal of maternal programming of stress responses in adult offspring through methyl supplementation: altering epigenetic marking later in life. J Neurosci. 2005;25(47):11045–11054. | ||
Meloni M. Race in an epigenetic time: thinking biology in the plural. Br J Sociol. 2017;68(3):389–409. | ||
Yudell M, Roberts D, Desalle R, Tishkoff S. Science and society: taking race out of human genetics. Science. 2016;351(6273):564–565. | ||
Garcia JA, Sanchez GR, Sanchez-Youngman S, Vargas ED, Ybarra VD. Race as lived experience: the impact of multi-dimensional measures of Race/Ethnicity on the self-reported health status of Latinos. Du Bois Rev. 2015;12(2):349–373. | ||
Sternthal MJ, Slopen N, Williams DR. Racial disparities in health: how much does stress really matter? Du Bois Rev. 2011;8(1):95–113. | ||
Williams DR, Priest N, Anderson NB. Understanding associations among race, socioeconomic status, and health: patterns and prospects. Health Psychol. 2016;35(4):407–411. | ||
Sack V, Murphey D. The prevalence of adverse childhood experiences, nationally, by state, and by race or ethnicity. Child Trends. 2018 (03). | ||
Slopen N, Shonkoff JP, Albert MA, et al. Racial disparities in child adversity in the U.S.: interactions with family immigration history and income. Am J Prev Med. 2016;50(1):47–56. | ||
Felitti VJ, Anda RF, Nordenberg D, et al. Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults. the adverse childhood experiences (ACE) study. Am J Prev Med. 1998;14(4):245–258. | ||
Nelson S, Simons LE, Logan D. The incidence of adverse childhood experiences (aces) and their association with pain-related and psychosocial impairment in youth with chronic pain. Clin J Pain. 2018;34(5):402–408. | ||
Anda R, Tietjen G, Schulman E, Felitti V, Croft J. Adverse childhood experiences and frequent headaches in adults. Headache. 2010;50(9):1473–1481. | ||
Zhang Z, Hayward MD, Yu YL. Life course pathways to racial disparities in cognitive impairment among older Americans. J Health Soc Behav. 2016;57(2):184–199. | ||
Umberson D, Williams K, Thomas PA, Liu H, Thomeer MB. Race, gender, and chains of disadvantage: childhood adversity, social relationships, and health. J Health Soc Behav. 2014;55(1):20–38. | ||
Bale TL, Baram TZ, Brown AS, et al. Early life programming and neurodevelopmental disorders. Biol Psychiatry. 2010;68(4):314–319. | ||
Khashan AS, Abel KM, Mcnamee R, et al. Higher risk of offspring schizophrenia following antenatal maternal exposure to severe adverse life events. Arch Gen Psychiatry. 2008;65(2):146–152. | ||
Blair MM, Glynn LM, Sandman CA, Davis EP. Prenatal maternal anxiety and early childhood temperament. Stress. 2011;14(6):644–651. | ||
Monk C, Spicer J, Champagne FA. Linking prenatal maternal adversity to developmental outcomes in infants: the role of epigenetic pathways. Dev Psychopathol. 2012;24(4):1361–1376. | ||
Buss C, Entringer S, Moog NK, et al. Intergenerational transmission of maternal Childhood maltreatment exposure: implications for fetal brain development. J Am Acad Child Adolesc Psychiatry. 2017;56(5):373–382. | ||
Tobi EW, Lumey LH, Talens RP, et al. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum Mol Genet. 2009;18(21):4046–4053. | ||
Heijmans BT, Tobi EW, Stein AD, et al. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc Natl Acad Sci USA. 2008;105(44):17046–17049. | ||
Palma-Gudiel H, Córdova-Palomera A, Eixarch E, Deuschle M, Fañanás L. Maternal psychosocial stress during pregnancy alters the epigenetic signature of the glucocorticoid receptor gene promoter in their offspring: a meta-analysis. Epigenetics. 2015;10(10):893–902. | ||
Palma-Gudiel H, Córdova-Palomera A, Leza JC, Fañanás L. Glucocorticoid receptor gene (NR3C1) methylation processes as mediators of early adversity in stress-related disorders causality: a critical review. Neurosci Biobehav Rev. 2015;55:520–535. | ||
Conradt E, Hawes K, Guerin D, et al. The contributions of maternal sensitivity and maternal depressive symptoms to epigenetic processes and neuroendocrine functioning. Child Dev. 2016;87(1):73–85. | ||
Sosnowski DW, Booth C, York TP, Amstadter AB, Kliewer W. Maternal prenatal stress and infant DNA methylation: a systematic review. Dev Psychobiol. 2018;60(2):127–139. | ||
Baum A, Garofalo JP, Yali AM. Socioeconomic status and chronic stress. does stress account for Ses effects on health? Ann N Y Acad Sci. 1999;896(1):131–144. | ||
Doan SN, Dich N, Evans GW. Childhood cumulative risk and later allostatic load: mediating role of substance use. Health Psychol. 2014;33(11):1402–1409. | ||
Cohen S, Janicki-Deverts D, Chen E, Matthews KA. Childhood socioeconomic status and adult health. Ann N Y Acad Sci. 2010;1186:37–55. | ||
Mclean J, Krishnadas R, Batty GD, et al. Early life socioeconomic status, chronic physiological stress and hippocampal N-acetyl aspartate concentrations. Behav Brain Res. 2012;235(2):225–230. | ||
Needham BL, Smith JA, Zhao W, et al. Life course socioeconomic status and DNA methylation in genes related to stress reactivity and inflammation: the multi-ethnic study of atherosclerosis. Epigenetics. 2015;10(10):958–969. | ||
Radley J, Morilak D, Viau V, Campeau S. Chronic stress and brain plasticity: mechanisms underlying adaptive and maladaptive changes and implications for stress-related CNS disorders. Neurosci Biobehav Rev. 2015;58:79–91. | ||
Maniam J, Antoniadis C, Morris MJ. Early-life stress, HPA axis adaptation, and mechanisms contributing to later health outcomes. Front Endocrinol. 2014;5(Suppl 3):73. | ||
Chrousos GP. Stress and disorders of the stress system. Nat Rev Endocrinol. 2009;5(7):374–381. | ||
Jackson JS, Knight KM, Rafferty JA. Race and unhealthy behaviors: chronic stress, the HPA axis, and physical and mental health disparities over the life course. Am J Public Health. 2010;100(5):933–939. | ||
Provençal N, Binder EB. The effects of early life stress on the epigenome: from the womb to adulthood and even before. Exp Neurol. 2015;268:10–20. | ||
Maletic V, Raison CL. Neurobiology of depression, fibromyalgia and neuropathic pain. Front Biosci. 2009;14(1):5291–5338. | ||
Goldenberg DL. Pain/depression dyad: a key to a better understanding and treatment of functional somatic syndromes. Am J Med. 2010;123(8):675–682. | ||
Hruschak V, Cochran G. Psychosocial predictors in the transition from acute to chronic pain: a systematic review. Psychol Health Med. 2018:1–17. | ||
Ruehlman LS, Karoly P, Newton C. Comparing the experiential and psychosocial dimensions of chronic pain in African Americans and Caucasians: findings from a national community sample. Pain Med. 2005;6(1):49–60. | ||
Edwards CL, Fillingim RB, Keefe F. Race, ethnicity and pain. Pain. 2001;94(2):133–137. | ||
Green CR, Baker TA, Sato Y, Washington TL, Smith EM. Race and chronic pain: a comparative study of young black and white Americans presenting for management. J Pain. 2003;4(4):176–183. | ||
Chen D, Meng L, Pei F, Zheng Y, Leng J. A review of DNA methylation in depression. J Clin Neurosci. 2017;43:39–46. | ||
Penas C, Navarro X. Epigenetic modifications associated to neuroinflammation and neuropathic pain after neural trauma. Front Cell Neurosci. 2018;12:158. | ||
Tajerian M, Alvarado S, Millecamps M, et al. DNA methylation of SPARC and chronic low back pain. Mol Pain. 2011;7:65. | ||
Bell JT, Loomis AK, Butcher LM, et al. Differential methylation of the TRPA1 promoter in pain sensitivity. Nat Commun. 2014;5:2978. | ||
Sukenaga N, Ikeda-Miyagawa Y, Tanada D, et al. Correlation between DNA methylation of TRPA1 and chronic pain states in human whole blood cells. Pain Med. 2016;17(10):1906–1910. | ||
Macedo JA, Hesse J, Turner JD, Meyer J, Hellhammer DH, Muller CP. Glucocorticoid sensitivity in fibromyalgia patients: decreased expression of corticosteroid receptors and glucocorticoid-induced leucine zipper. Psychoneuroendocrinology. 2008;33(6):799–809. | ||
Menzies V, Lyon DE, Archer KJ, et al. Epigenetic alterations and an increased frequency of micronuclei in women with fibromyalgia. Nurs Res Pract. 2013;2013(3):1–12. | ||
Pennebaker JW, Susman JR. Childhood Trauma Questionnaire. In Measurement Instrument Database for the Social Science, 2013. Available from: http://www.midss.org/content/childhood-trauma-questionnaire. Accessed January 25, 2019. | ||
Ligon CO, Moloney RD, Greenwood-van Meerveld B. Targeting epigenetic mechanisms for chronic pain: a valid approach for the development of novel therapeutics. J Pharmacol Exp Ther. 2016;357(1):84–93. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.