Back to Journals » Infection and Drug Resistance » Volume 14
Correlation Between Biofilm-Formation and the Antibiotic Resistant Phenotype in Staphylococcus aureus Isolates: A Laboratory-Based Study in Hungary and a Review of the Literature
Authors Senobar Tahaei SA, Stájer A ![]() , Barrak I
, Barrak I ![]() , Ostorházi E, Szabó D, Gajdács M
, Ostorházi E, Szabó D, Gajdács M ![]()
Received 27 January 2021
Accepted for publication 11 March 2021
Published 23 March 2021 Volume 2021:14 Pages 1155—1168
DOI https://doi.org/10.2147/IDR.S303992
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Seyyed Askhan Senobar Tahaei,1 Anette Stájer,2 Ibrahim Barrak,2 Eszter Ostorházi,3 Dóra Szabó,3 Márió Gajdács1,3
1Department of Pharmacodynamics and Biopharmacy, Faculty of Pharmacy, University of Szeged, Szeged, 6720, Hungary; 2Department of Periodontology, Faculty of Dentistry, University of Szeged, Szeged, 6720, Hungary; 3Institute of Medical Microbiology, Faculty of Medicine, Semmelweis University, Budapest, 1089, Hungary
Correspondence: Márió Gajdács Tel +36 62-341-330
Email [email protected]
Introduction: Staphylococcus aureus (S. aureus) is an important causative pathogen in human infections. The production of biofilms by bacteria is an important factor, leading to treatment failures. There has been significant interest in assessing the possible relationship between the multidrug-resistant (MDR) status and the biofilm-producer phenotype in bacteria. The aim of our present study was to assess the biofilm-production rates in clinical methicillin-susceptible S. aureus [MSSA] and methicillin-resistant S. aureus [MRSA] isolates from Hungarian hospitals and the correlation between resistance characteristics and their biofilm-forming capacity.
Methods: A total of three hundred (n=300) S. aureus isolates (corresponding to MSSA and MRSA isolates in equal measure) were included in this study. Identification of the isolates was carried out using the VITEK 2 ID/AST system and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Antimicrobial susceptibility testing was performed using the Kirby–Bauer disk diffusion method and E-tests, confirmation of MRSA status was carried out using PBP2a agglutination assay. Biofilm-production was assessed using the crystal violet (CV) tube-adherence method and the Congo red agar (CRA) plate method.
Results: There were significant differences among MSSA and MRSA isolates regarding susceptibility-levels to commonly used antibiotics (in case of erythromycin, clindamycin and ciprofloxacin: p< 0.001, gentamicin: p=0.023, sulfamethoxazole/trimethoprim: p=0.027, rifampin: p=0.037). In the CV tube adherence-assay, 37% (n=56) of MSSA and 39% (n=58) of MRSA isolates were positive for biofilm-production, while during the use of CRA plates, 41% (n=61) of MSSA and 44% (n=66) of MRSA were positive; no associations were found between methicillin-resistance and biofilm-production. On the other hand, erythromycin, clindamycin and rifampin resistance was associated with biofilm-positivity (p=0.004, p< 0.001 and p< 0.001, respectively). Biofilm-positive isolates were most common from catheter-associated infections.
Discussion: Our study emphasizes the need for additional experiments to assess the role biofilms have in the pathogenesis of implant-associated and chronic S. aureus infections.
Keywords: Staphylococcus aureus, MSSA, MRSA, biofilm, antibiotic resistance, crystal violet, Congo red agar, phenotypic assay
Introduction
Staphylococcus aureus (S. aureus) is a Gram-positive, catalase-positive and bacitracin-resistant coccus, which is a common colonizer of the human body. These bacteria are frequently found on mucosal surfaces (eg, the nares, the throat and the rectum) and moist regions of the skin (eg, axilla, groin and perineum).1,2 According to recent data, 60% of the population is transiently colonized, while in 30%, this colonization is persistent.3 S. aureus (both MSSA [methicillin-sensitive S. aureus] and MRSA [methicillin-resistant S. aureus]) is an exceptionally successful and adaptable pathogen, relevant in both community-associated and nosocomial infections.4 They are an important cause of skin and soft tissue infections (SSTIs), osteoarticular infections, medical device-related infections, pneumonia, infective endocarditis and bacteremia (in addition, through hematogenous spread, this microorganism may cause a wide range of secondary pathologies).5,6 MRSA was first identified in 1961 and has emerged as the first multidrug-resistant (MDR) bacterium in human medicine.7 MRSA is resistant to all β-lactam antibiotics (with the exception of fifth-generation cephalosporins), severely narrowing safe and effective treatment options; additionally, these strains often possess a battery of resistance-determinants against other antibiotic groups (eg, fluoroquinolones, macrolides, tetracyclines, aminoglycosides), thus earning the name “superbug” for the first time.8 Methicillin-resistance is mediated by modifications in penicillin-bindings proteins (namely PBP2a/2c/2ʹ), owing to the presence of mecA or mecC genes. Initially, MRSA was mainly associated with nosocomial infections (hospital-associated MRSA; HA-MRSA), however, some 20–30 years after its initial description, community-associated MRSA (CA-MRSA) infections have also emerged.4,9 Since the 2000s, extensive research regarding livestock-associated MRSA (LA-MRSA) has been published, both due to its impact in veterinary medicine (for animal husbandry and for companion animals) and due to the possible relevance of animals as vectors for MRSA transmission.10 The prevalence of MRSA infections shows large geographical differences: it is around 1–10% in Northern Europe, 15–30% in the United States, 40–50% in Southern and Eastern Europe, while it may exceed 80% in some parts of Asia.11 Risk factors associated with acquiring an MRSA infection include advanced age (≥60 years), prolonged hospital stay, prior antimicrobial treatment and the use of nasogastric tubes or endovascular catheters.12 MRSA infections are associated with decreased quality of life (QoL), excess mortality and substantial economic costs, compared to MSSA infections.4,12
The emergence of MDR isolates in human infections considerably limits clinicians in administering adequate antimicrobial therapy.13,14 A variety of resistance-determinants have been described in the literature (both intrinsic resistance and genes acquired on mobile genetic elements), allowing bacteria to withstand otherwise lethal doses of antibiotics.15 In addition to these resistance-determinants, the production of biofilms by bacteria is another important factor leading to treatment failures.16 The first record on the existence of bacterial biofilms was published by Henrici (1933), while a recent publication by the National Institute of Health (NIH) suggested that in in vivo conditions, 60% of all infections are caused by bacteria embedded in biofilms.17,18 Biofilms are aggregates of mono-species or multispecies bacterial communities, enveloped in a protective extracellular matrix.19,20 This matrix is typically made up of secreted exopolysaccharides (EPS), environmental DNA (eDNA), proteins, surfactants, lipids and water.21 Biofilms allow bacterial communities to attach to and persist on inanimate surfaces and inside the body. The initial step of biofilm-production is the attachment of bacteria to relevant surfaces (most commonly coarse or hydrophobic surfaces, such as catheters, implanted medical devices and other biomaterials), with the aid of EPS, surface proteins, fimbriae and pili.22 After the development of the mature biofilm, bacteria residing inside this protective structure will be in different metabolic states: bacteria in the surface layer of the biofilm will be aerobic and metabolically active; while in the deeper layers, due to nutrient deficiency and lower oxygen concentrations, bacteria are fermentative and dormant.23 In essence, biofilms provide double protection against antibiotics: as most antibiotics are only effective against actively-replicating (ie, planktonic) cells, the eradication of these persisters is an important challenge; additionally, the thick biofilm also acts as a pharmacokinetic barrier, limiting the diffusion of antimicrobials and other noxious agents in the vicinity of the pathogens.24,25 This may result in minimal inhibitory concentrations (MICs) 10–10,000-times higher against bacteria embedded in biofilms.26 Owing to this resistance against antibiotics and the protective effects of biofilms against harsh environmental stressors (eg, sheer forces, drying) and the immune system (eg, phagocytosis), it is unsurprising that biofilms are an important virulence factor in the development of skin and soft tissue infections, catheter- (intravascular or urinary) and medical device-associated infections, oral infections, dental caries and chronic infections.27–30
At present, the group of “ESKAPE” bacteria – which is a list consisting of MDR pathogens, including MRSA – is considered as the most concerning in respect to their resistance rates, clinical impact and mortality.31 As most of the ESKAPE-members are biofilm-producers, there has been significant interest in assessing the possible relationship between their MDR status and the biofilm-producer phenotype.32 Although several studies have provided experimental data on the subject, corresponding to both MDR and wild-type Gram-negative (eg, Escherichia coli,33 Klebsiella spp.,34 Pseudomonas aeruginosa,35 Acinetobacter spp.36) and Gram-positive bacteria (eg, MSSA/MRSA,37 Enterococcus spp.38), the findings of these studies are often controversial. With this in mind, the aim of our present study was to assess the rates of biofilm-production in various clinical MSSA and MRSA isolates from Hungarian hospitals with phenotypic methods, and the potential correlation between the resistance characteristics and their biofilm-forming capacity.
Materials and Methods
Collection of Isolates
A total of three hundred (n=300) S. aureus isolates (corresponding to MSSA and MRSA isolates in equal measure; n=150 isolates, respectively) were included in this study, which were kindly provided by a tertiary-care teaching hospital and two smaller regional hospitals in Hungary. The study uses a cross-sectional study design; the microorganisms were isolated between 2019.01.01 and 2020.01.01., including n=100 isolates from catheter-associated infections (CAI-SA), skin and soft tissue infections (SSTI-SA) and urinary tract infections (UTI-SA). During our experiments, S. aureus ATCC 29213 (MSSA; positive for biofilm-production, icaAB gene negative), S. aureus ATCC 43300 (MRSA; positive for biofilm-production, icaAB gene positive), S. aureus ATCC 12600 (MSSA; non-biofilm producing, icaAB gene negative), S. epidermidis ATCC 35984 (positive for biofilm-production, icaAB gene positive) and S. epidermidis ATCC 12224 (non-biofilm producing, icaAB gene negative) were used as control strains, obtained from the American Type Culture Collection (ATCC; Manassas, VI, USA).39 Stock cultures were stored at −80 °C in a cryopreservation medium (700 µL trypticase soy broth + 300 µL 50% glycerol).
Bacterial Identification
Identification of S. aureus isolates was carried out using the VITEK 2 ID/AST automated system (bioMérieux, Marcy-l’Étoile, France) and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS; Bruker Daltonics, Bremen, Germany). During the MALDI-TOF assay, bacterial cells from fresh overnight cultures were transferred to a stainless-steel target. An on-target extraction was performed by adding 1 µL of 70% formic acid prior to the matrix. After drying at room temperature, the cells were covered with 1 µL matrix (α-cyano-4-hydroxy cinnamic acid in 50% acetonitrile/2.5% trifluoro-acetic acid; Bruker Daltonics, Bremen, Germany). Mass spectrometry analyses were performed by the Microflex MALDI Biotyper (Bruker Daltonics, Bremen, Germany) in positive linear mode across the m/z range of 2 to 20 kDa; for each spectrum, 240 laser shots at 60 Hz in groups of 40 shots per sampling area were collected. The MALDI Biotyper RTC 3.1 software (Bruker Daltonics, Bremen, Germany) and the MALDI Biotyper Library 3.1 were used for spectrum analysis. As a result of the MALDI-TOF spectrum analysis, a log(score) value was provided, indicating the reliability of MALDI-TOF MS identification. The log(score) values were evaluated as follows: a log(score) <1.69 showed unreliable identification, 1.70–1.99 cor-responded to probable genus-level identification, 2.00–2.29 corresponded to reliable genus-level identification, while a score ≥2.30 corresponded to reliable species-level identification.40 All isolates included in the study were re-identified as S. aureus before further experiments.
Antimicrobial Susceptibility Testing, Resistance Detection
Antimicrobial susceptibility testing (AST) was performed either using the Kirby–Bauer disk diffusion method or E-tests (Liofilchem, Teramo, Italy) on Mueller–Hinton agar (MHA) plates. During testing, the susceptibilities to erythromycin (ERY; 15 µg), clindamycin (CLI; 2 µg), ciprofloxacin (CIP; 5 µg), gentamicin (GEN; 10 µg), sulfamethoxazole/trimethoprim (SXT; 23.75/1.25 µg), vancomycin (VAN; E-test), tigecycline (TIG; 15 µg), linezolid (LZD; 10 µg), fusidic acid (FUS; 10 µg), quinupristin/dalfopristin (QDP; 15 µg), rifampicin (RIF; 5 µg) and ceftaroline (CFT; 5 µg) were determined. Interpretation of testing results and classification of isolates as MDR (being non-susceptible to at least one antimicrobial agent in three or more antimicrobial classes) was based on EUCAST standards and breakpoints v. 9.0. (http://www.eucast.org). VITEK 2 ID/AST (bioMérieux, Marcy-l’Étoile, France) was used for the verification of discrepant results. During data analysis, intermediate results were grouped with and reported as resistant. Inducible CLI resistance was detected using the ERY-CLI D test; these strains were also reported as resistant.41
Methicillin-resistance was verified using mannitol salt agar (MSA) plates using cefoxitin (FOX) disks (zone diameters under 22 mm were considered positive for methicillin-resistance) and PBP2ʹ Latex Agglutination Test (Thermo Fisher Scientific Hungary GmbH, Budapest, Hungary). A MRSA strain was automatically considered to be resistant to all β-lactam antibiotics other than CFT.41 MSSA S. aureus ATCC 29213 and MRSA S. aureus ATCC 43300 were used as quality control strains.
Detection of Biofilm-Production by the Tube-Adherence Method
Assessment of biofilm-formation was carried out in the tube-adherence method described previously.42 In short, glass tubes containing 1 mL of sterile trypticase soy broth (bioMérieux, Marcy-l’Étoile, France) were inoculated with 1 µL of the overnight culture of a respective bacterial strains. Respective tubes were then incubated statically for 24 h at 37 °C. Verification of planktonic growth was observed visually. After the incubation period, the supernatant was then discarded, the adhered cells were rinsed three times with phosphate buffer saline (PBS; Sigma-Aldrich; Budapest, Hungary) and the tubes were patted dry on a paper towel. The contents of the tubes were treated with a 1 mL solution of 0.1% crystal violet (CV; Sigma-Aldrich; Budapest, Hungary) to stain the adhered biomass; the tubes were incubated for 3 h at room temperature with the staining solution. The CV solution was then discarded and the tubes were again rinsed three times with PBS and the tubes were patted dry on a paper towel. Biofilm-formation was observed visually; in case of the appearance of visible biofilm lining at the bottom and on wall of the glass tubes, the strain was considered a biofilm-producer in this assay.33,42 All experiments were performed in triplicate and were evaluated by two independent researchers.
Detection of Biofilm-Production by the Congo Red Agar Method
Biofilm-formation of the isolates was also evaluated on Congo Red Agar (CRA) plates, based on the previously described protocol.43 Briefly, CRA plates were prepared using trypticase soy agar supplemented with 5% sucrose and 40 μg/mL Congo red dye (Sigma-Aldrich; Budapest, Hungary). Congo red is a secondary diazo dye, which can be used as a pH indicator (with a detectable color change at pH 3.0–5.2). Strains were cultured on trypticase soy agar plates at 37 °C for 16–18 h; cells were resuspended in trypticase soy broth at a density of OD600=2; 10 µl of the suspension was spotted on CRA plates. The inoculated CRA plates were incubated at 37 °C in aerobic conditions for 24 h, followed by incubation at room temperature before the reading of the plates for an additional 24 h. The isolates were assessed for their colony-morphologies: black colonies with a dry consistency and rough surface edges were considered as biofilm-producers in this assay, while red colonies with smooth, round and shiny surface were read as negative for biofilm-production.43 All experiments were performed in triplicate and were evaluated by two independent researchers.
Statistical Analysis
Descriptive statistical analysis (including means and percentages to characterize data) was performed using Microsoft Excel 2013 (Microsoft Corp.; Redmond, WA, USA). Additional statistical analyses were performed with IBM SPSS Statistics for Windows 24.0 (IBM Corp., Armonk, NY, USA), using the χ2-test.44 p values <0.05 were considered statistically significant. Additionally, consistency-assessment was also performed between the results of the two different biofilm-production studies (CV assay vs CRA agar).30
Ethical Considerations
Clinical, personal and epidemiological data pertaining to the affected patients were not collected or provided during the study, bacterial isolates were only identifiable based on their serial number; therefore, our present study was not subject to ethics review.
Results
Antibiotic Susceptibility of MSSA and MRSA Isolates Included in the Study
Out of the tested S. aureus isolates, the following susceptibilities were detected overall: complete susceptibility (100%; n=300) was seen for VAN, CFT, QDP, FUS, LZD and TIG 100%; on the other hand, varying levels of resistance were observed for other antibiotics, such as GEN 90% (n=271), SXT 89% (n=268), RIF 86% (n=257), ERY 54% (n=162), CIP 51% (n=153) and CLI 48% (n=144). The detailed susceptibilities for every group of isolates (namely CAI-SA, SSTI-SA and UTI-SA) are presented in Table 1. There were significant differences among MSSA and MRSA isolates regarding susceptibility-levels to commonly used antibiotics (ERY: p < 0.001, χ2 = 156.52, degrees of freedom [DOF]: 1; CLI: p < 0.001, χ2 = 155.63, DOF: 1; CIP: p < 0.001, χ2 = 192.03, DOF: 1; GEN: p < 0.023, χ2 = 12.03, DOF: 1; SXT: p = 0.0027, χ2 = 8.95, DOF: 1; RIF: p = 0.0037, χ2 = 8.95, DOF: 1). Similarly, the subset of MRSA isolates could be classified as MDR more commonly (85% [n=128] vs 8% [n=12]), compared to isolates from the MSSA group.
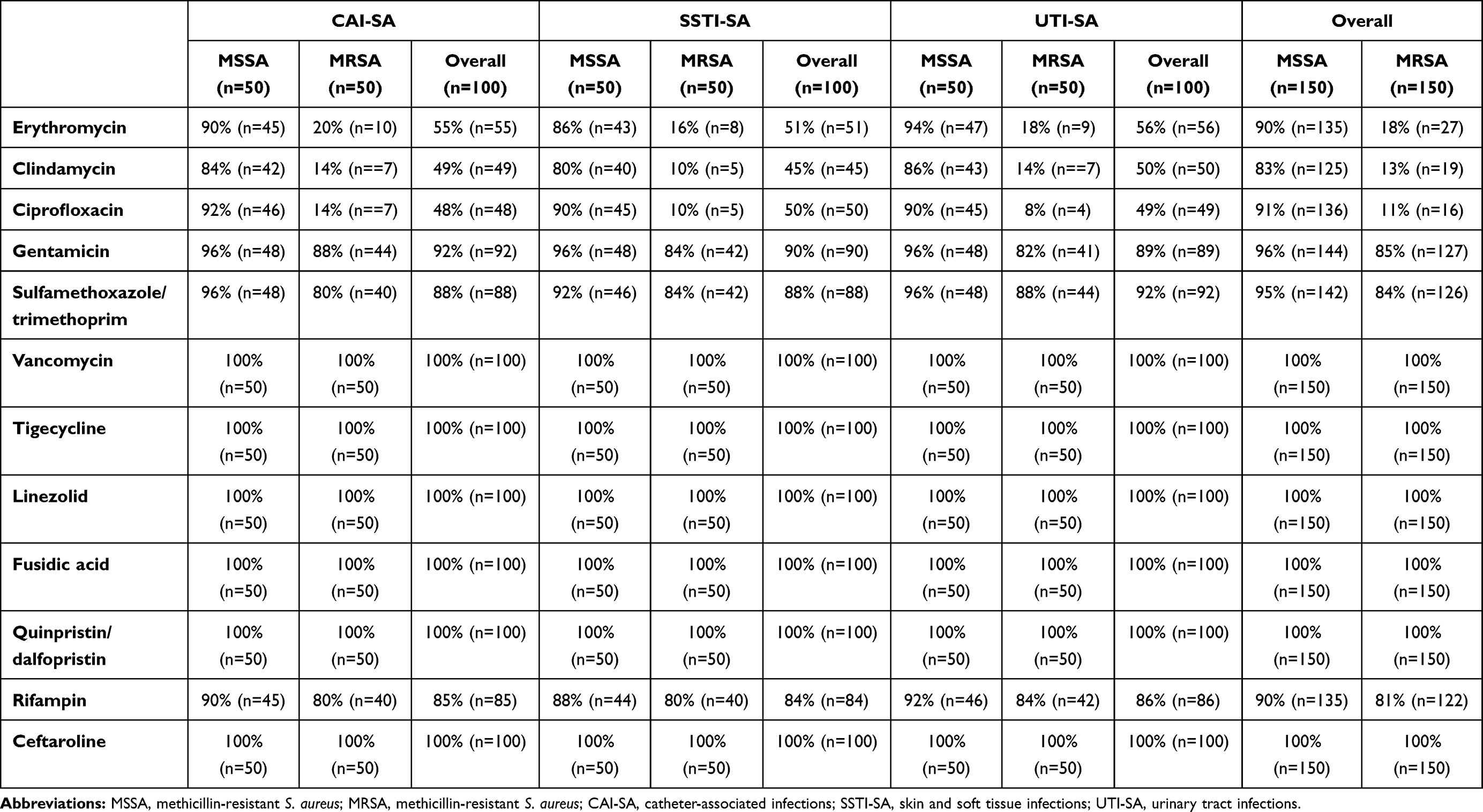 |
Table 1 Antibiotic Susceptibility Rates Among S. aureus Isolates Included in This Study |
Association of MSSA/MRSA Status and Resistance to Other Antibiotics with Biofilm-Production
In the CV tube adherence-assay, 37% (n=56) of MSSA and 39% (n=58) of MRSA isolates were positive for biofilm-production, while during the use of CRA plates, 41% (n=61) of MSSA and 44% (n=66) of MRSA were positive; no associations were found between methicillin-resistance and biofilm-production (p>0.05 in both cases). The agreement between the results of the two phenotypic testing methods was 0.897 (89.7%) overall (91.8% in case of MSSA and 87.8% in case of MRSA isolates). Interestingly, biofilm-production was more commonly detected from both MSSA and MRSA CAI-SA isolates (p<0.0001 and p=0.0018 for MSSA, p=0.02 and p=0.015 for MRSA, respectively), compared to the isolates from other origins. The detailed distribution of biofilm-positive S. aureus among the isolates of different origin, the results of the statistical analyses and the agreements among the CV adherence assay and CRA plates are presented in Figure 1. Among the control strains, S. aureus ATCC 29213, S. aureus ATCC 43300, S. epidermidis ATCC 35984 were positive for biofilm-production in both phenotypic assays, while S. aureus ATCC 12600 and S. epidermidis ATCC 12224 were both negative.
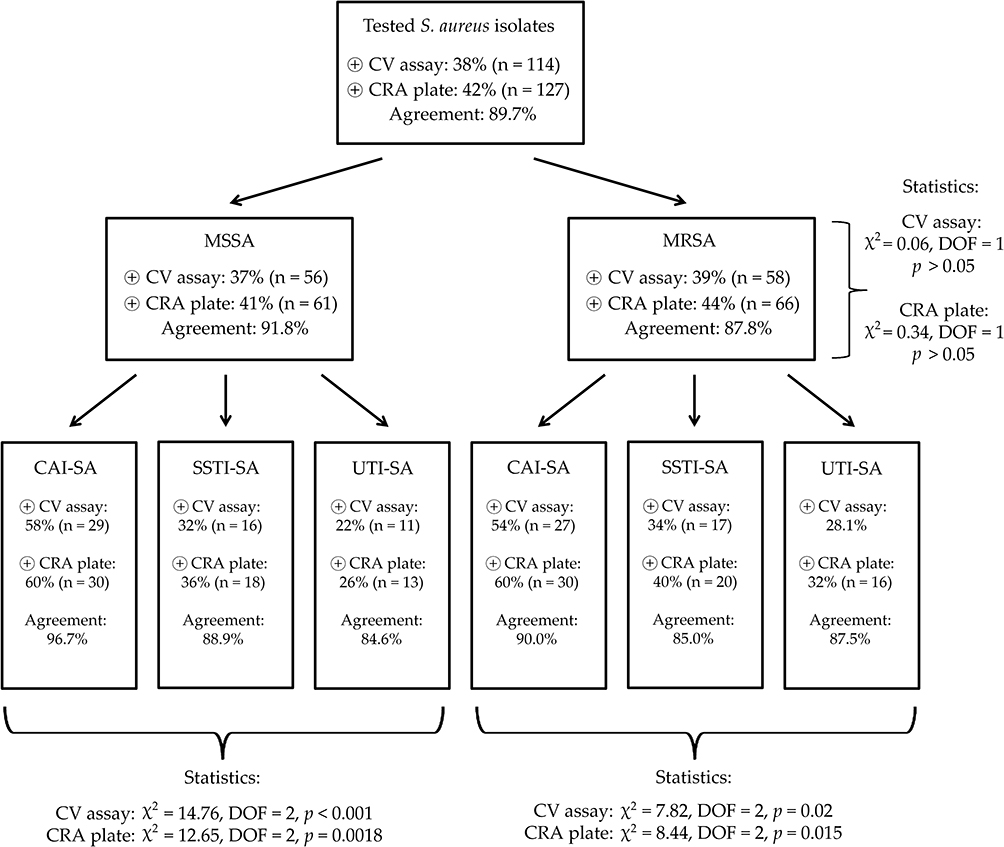 |
Figure 1 Antibiotic susceptibility rates among S. aureus isolates included in this study. Abbreviations: MSSA, methicillin-resistant S. aureus; MRSA, methicillin-resistant S. aureus; CAI-SA, catheter-associated infections; SSTI-SA, skin and soft tissue infections; UTI-SA, urinary tract infections. |
The relationship between biofilm-production and resistance to other antibiotics was also assessed; during these analyses, only the results from the CRA plates were considered. It was found that resistance to ERY (p = 0.004, χ2 = 8.12, DOF: 1), CLI (p < 0.001, χ2 = 44.57, DOF: 1) and RIF (p < 0.001, χ2 = 96.95, DOF: 1) was associated with biofilm-positivity; in fact, 37 out of 43 (86%) of RIF-resistant isolates were biofilm-producers. On the other hand, this association was not shown for other antibiotics (ie, CIP, GEN, SXT; p>0.05).
Discussion and Review of the Literature
S. aureus infections are associated with considerable morbidity, mortality and economic costs for the healthcare institutions worldwide.45 Owing to the adaptability, the plethora of virulence factors and the increasing levels of antimicrobial resistance in S. aureus, treatment of these infections is a considerable challenge for clinicians.46 Biofilm-formation has been classified as an important defense mechanism and pathogenic hallmark for both MSSA and MRSA isolates, both as a means to persist in the environment (eg, on a hospital ward) and in the host during infections.47 It has been described that the staphylococcal colonization of the skin is dependent of the biofilm-formation of these bacteria; in addition, S. aureus strains adhere to damaged skin and mucosal surfaces more easily, leading to the development of SSTIs.48 At the same time, all inserted and implanted medical devices (contact lenses, cardiac pacemakers, prosthetic valves, cerebrospinal fluid shunts, implanted catheters and synthetic joints) are at risk to be associated with S. aureus infections (eg, endocarditis, osteomyelitis, bacteremia).49 In the present study, we have investigated n=150 MSSA and n=150 MRSA isolates – originating from clinical materials – regarding their antibiotic susceptibilities and their biofilm-forming capacities using two phenotypic tests, namely the CV tube-adherence assay and the plate-based CRA medium. There are a plethora of methods available for the characterization of the biofilm-forming capacity of bacteria – self-developed and chromogenic media (both in liquid and in plate form), staining methods, assessment via measurements with a spectrophotometer or electron microscopy and most recently, flow chamber systems – however, these are pronounced differences among these methods in the price, reproducibility, high-throughput nature and the in vivo adaptability of the results.50–52 Our two utilized methods have been described for a number of years (the CV tube-based assay was described by Christensen et al in 1982,53 while the CRA method was developed by Freeman et al in 1989);54 although subsequent studies have demonstrated that these methods needed to be modified to improve accuracy and sensitivity, these methods are cheap, easy to perform, the criteria for their evaluation are straightforward and their results are comparable to other, more expensive assays.39 In our study, no significant association was noted between MSSA/MRSA-status and biofilm-production in either phenotypic tests; however, ERY, CLI and RIF resistance was more common in biofilm-producing S. aureus isolates. It is also interesting to note that 86% of isolates resistant to RIF were biofilm-producers, especially as RIF is considered an effective antimicrobial agent with good penetration into bacterial biofilms in vivo.55 Although the exact reason behind this phenomenon is unknown, it has been described that the sub-inhibitory concentrations of several antibiotics may induce biofilm-formation in S. aureus isolates.56 Interestingly, the interplay between sub-inhibitory doses of antibiotics has been most frequently published in relation to the MLKS (macrolide-lincosamide-ketolide-streptogramin; which are all protein synthesis inhibitors affecting the 50S ribosome) group of drugs and rifampin.57 For example, Lima-e-Silva et al reported that in sub-lethal doses (MIC/2 and MIC/4), rifampin strongly stimulated biofilm-formation (when measured by the CRA plate and CV microtiter plate assay), in contrast to minocycline, which did not have such inducing effects.58 The effects of low antibiotic doses on the biofilm-forming capacity are thought to occur by differential expression of genes of interest due to the noxious agents.59 The literature has shown that sub-MIC concentrations of tetracycline, quinupristin and dalfopristin were presented as strong inducers, while erythromycin was noted as a weak inducer of ica-gene expression. In contrast, gentamicin, chloramphenicol (which are also protein synthesis inhibitors), penicillin, oxacillin, ofloxacin and vancomycin did not present with similar effects.60 The clinical origin of the isolates also had an effect on their biofilm-producing capacity: they were the most prevalent in isolates from catheter-associated infections and the least common in strains isolated from the urinary tract. Finally, the agreement among the results of the two in vitro methods was 89.7%, which is a very good result, based on other studies from the literature.39
Many studies have aimed to assess the correlation of biofilm-production with antibiotic-resistance in S. aureus strains; nevertheless, the literature has shown conflicting data on the topic, thus, at present, we are unable to draw far-reaching conclusions. To make the interpretation of the currently available results even more troublesome, many different methodologies have been used to assess biofilm-formation, with or without the molecular characterization of the isolates. Similarly to our results, Arslan et al found no association between slime production and methicillin-resistance or resistance to other antibiotics in a sample of n=187 S. aureus isolates; in their report, CRA plates and the CV tube adherence assay were utilized, while molecular testing (for clonality or the presence of agr genes) was not performed.61 Ghasemian et al assessed biofilm-production in n=209 S. aureus isolates, among which no relevant differences were shown for biofilm-positive and negative isolates, based on methicillin-resistance (36.1% and 28.9% for MRSA and MSSA, respectively). The extent of biofilm-production was assessed by a microtiter plate biofilm assay, and the genetic testing of the isolates was also performed: 58.3% and 22.0% of isolates belonged to the agr groups I and II, respectively, while 84.0% of MRSA isolates possessed the SCCmec III cassette.62 Rodríguez-Lopez et al came to similar conclusions, when studying S. aureus isolates originating from animal and environmental samples at heavy swine farms in Italy. In this report, the same methodology was utilized (plate-based quantitative measurements), and spa-typing was also performed: overall, isolates belonging to the ST398/t899 and ST398/t011 were the most common among biofilm-producers.63 El-Nagdy et al detected biofilm-forming S. aureus (using the CRA plate method, complemented with scanning and transmission electron microscopy) from febrile neutropenic patients in Egypt; interestingly, they have found that 72.7% of isolates were biofilm-positive. Among the tested strains, 37.5% were positive for icaA, and 22.9% were positive for icaD; however, only 50% of biofilm-producers carried either the icaA or icaD genes.64 Similarly, no correlation among methicillin-resistance and biofilm-production was seen in the reports of Knobloch et al (including n=128 ica-positive isolates, with utilizing CRA plates and the microtiter plate method)65 and Mathur et al (where n=152 Staphylococcus spp. were tested using the microtiter plate method).66
However, there have been studies that identified differences in the rates of biofilm-production based on the phenotypic resistance of S. aureus isolates. In the study by Bose et al (in which the authors have utilized similar methodologies to our study, in addition to the tissue culture plate method to quantify their results) found higher levels of antibiotic resistance in biofilm-producing S. aureus and S. epidermidis isolates.67 Piechota et al compared the biofilm-forming capacity of MSSA and MRSA isolates from Poland: in this report, MRSA isolates were stronger biofilm-producers overall in the microtiter plate-based assay (39.7% vs 36.8%) and the occurrence of icaABCD genes (51.5% overall) was also more common in methicillin-resistant strains.68 Cha et al characterized n=126 MRSA isolates from a Korean teaching hospital, during which they have found higher levels of MDR isolates (defined as MRSA+resistance to at least three non-β-lactam agents) among the biofilm-forming isolates. The study group has used the microtiter plate assay to quantify biofilm-formation and has also performed molecular testing: they have shown that the majority of the isolates were ST5 (69.8%) and 64.0% of isolates are from the agr group II.69 Souli et al have clearly demonstrated that strong biofilm-producers (tested with the CV tube-based assay) among S. epidermidis isolates had also possessed higher levels of resistance in vitro.70 Agarwal and Jain tested commensal, colonizing and invasive S. aureus isolates for their biofilm-forming capacity using the microtiter plate assay; in their study, biofilm-producers were more frequently MDR in all groups, and 94.0% of biofilm-producers carried ica-genes.71 De Araujo et al reported that methicillin-resistant and MDR S. epidermidis isolates were more frequent among biofilm-producers (tested by the microtiter plate assay). They have found that 96.0% of biofilm-positive isolates carried icaA or icaD genes, while 86.0% were positive for atlE and aap genes.72 In contrast, in a report concerning S. aureus isolates originating from pork, Zhang et al found 83.8% of the bacteria to be agr-positive (agr I: 39.2%, agr IV: 32.3%) and noted the high prevalence of MDR isolates in moderate and weak biofilm-producers, when tested with the microplate method.73 Our study demonstrated an association of CLI-resistance and biofilm-production corresponding to our isolates, which has been identified in other reports as well, although these studies usually also showed a positive correlation with methicillin-resistance. In the study by Belbase et al, S. aureus isolates from pus/wound swab samples were assessed for the susceptibility and microtiter-based biofilm-production in a tertiary-care hospital in Nepal. Overall, methicillin-resistance and inducible CLI-resistance were more common in biofilm-producers.38 In another study from Nepal, Manandhar et al also showed that the in vitro biofilm-production (assessed by CRA plates) of S. aureus isolates was associated with methicillin-resistance and inducible CLI-resistance.74 Bhattacharya et al tested n=100 S. aureus isolates, including 47% MRSA isolates: biofilm-positivity was shown in 55.0% of isolates, with MRSA isolates in higher numbers among biofilm-producers; in addition, resistance-levels against CIP, RIF, ERY, CLI and SXT were also significantly higher in biofilm-positive isolates.75 Finally, Neopane et al tested n=150 S. aureus isolates, originating from pus samples: in their study – although no clear association was seen with methicillin-resistance and biofilm-production, strong biofilm-production and the MDR status has shown good agreement.76 In the latter three studies, only phenotypic characterization of resistance and biofilm-production was utilized. Overall, the literature findings suggest a possible relationship between the expression of antibiotic-resistance-determinants and biofilm-production, however, the clarification of nature of this association will require further studies.
Pulse-field gel electrophoresis (PFGE) and multi-locus sequence typing (MLST) are not routinely performed in Hungary by diagnostic laboratories; thus, there is a scarcity of local data regarding the molecular epidemiology of S. aureus isolates, such data may be sourced from major national public health surveillance studies: the Hungarian clone (ST239-III, PFGE type E) – which was predominant before 2000 – was almost completely replaced by the Southern German clone (ST228-I, PFGE type B) and the New York/Japan epidemic clone (ST5-II, PFGE type A/C), and since 2006, the breakthrough of the EMRSA-15 (ST22-IV, PFGE type D) was described.77 Based on the most recent data (2017–2018) available from the National Institute of Public Health in Hungary, ~45% of HA-MRSA isolates were ST22-IV clones, while ~24% of isolates were ST5-II; in case of CA-MRSA isolates, ST8-IV and ST80-IV clones were the most common, while among LA-MRSA, CC398 isolates were the most prevalent.78,79 Literature data also suggest that the clonal background of S. aureus isolates may play a major role in the biofilm-forming capacity, however (as demonstrated by the publications discussed in the previous section), the clonality of the isolates is seldom reported in these studies.80 Croes et al reported that S. aureus isolates from MLST clonal complex CC8 were the most potent biofilm-producers – irrespective of the glucose-concentration (0–0.5%) in the media; at physiological glucose (0.1%) concentration, >60% of CC8 isolates were strong biofilm-producers, compared to 0–7% in other tested CCs.81 These findings were also supported by the results of Luther et al: from n= 182 clinical MRSA strains, isolates belonging to the CC8 group and spa type t008 group were significantly more common among strong biofilm-producers (p=0.01), while spa type t895 and β-toxin-producing isolates showed a negative correlation with biofilm-production.82 The study of Recker et al performed combined study, including laboratory assays and a data analysis regarding bacterial genotype and phenotype with available clinical metadata in a machine-learning framework, corresponding to n=300 S. aureus, including CC22 and CC30 isolates from bacteremia. Their results showed no relevant differences in biofilm-formation among MSSA and MRSA isolates or among CC22 and CC30 isolates. Although this report showed no association with biofilm-production and SCCmec-type, other studies highlighted that the SCCmec II element is associated with decreased capability to form biofilm.83 On the other hand, Lim et al (who assessed n=465 clinical S. aureus isolates) and da Fonseca Batistao (during the study of fifteen isolates) both concluded that the presence of the SCCmec III cassette is a good predictor of strong biofilm-forming ability.84,85 The importance of the SCCmec cassette and mecA genes in biofilm formation were highlighted by Pozzi et al, where ΔSCCmec (deletion mutants) presented with decreased expression of virulence determinants, including biofilms; the authors of the study have concluded that these genes may have the potential to affect phenotypic characteristics mediated by other operons (ie, agr or icaADBC) to facilitate the adaptation of hospital-associated MRSA to the harsh environment in hospitals.86 This may be the reason for the larger number of biofilm-producing isolates from catheter-associated (CAI-SA) infections.
It has been described that S. aureus strains usually form multilayered biofilms; this biofilm is useful in the evasion of the non-specific and adaptive immune responses of the host, including decreased rate of opsonization, phagocytosis, killing by neutrophil granulocytes, and Toll-like receptor activation.87,88 The latter is particularly relevant, because TLR-activation is important in facilitating a Th2-type immune response, which may act to prevent S. aureus biofilm-associated infections.89 Biofilms are also protective against reactive oxygen species (ROS) in in vivo environments.90 Biofilms also facilitate the metabolic transformation of S. aureus into the small colony variant (SCV; which may be seen as “dwarf colonies” on solid media) morphotype: in this sub-population, bacteria exist at a lower metabolic activity (leading to increased antimicrobial resistance), which also enables in vivo persistence and chronic infections.91 In addition to metabolic switching, SCVs are also characterized by adaptation for intracellular survival in mammalian cells, where the pathogen acts similarly to other microorganisms with a strictly intracellular life cycle.92,93 The relevance of eDNA in the stability and antimicrobial resistance of staphylococcal biofilms has been demonstrated, as DNase treatment (eg, in cystic fibrosis) clearly negatively impacts the biofilm structure.94 Biofilm-formation in S. aureus is genetically mediated by the regulatory genetic locus staphylococcal accessory regulator (sarA); this controls two pathways – namely the intracellular adhesin (ica) operon and accessory gene regulator (agr) regulated pathways – both of which have been suggested as determinants of the extent of biofilm-formation in these bacteria.95,96 The product of the genes of the ica operon (icaADBC) are the IcaA and IcaD transferase membrane proteins; these proteins have important roles in the biosynthesis of PIA (polysaccharide intercellular adhesion protein; or poly-β-1,6-N-acetylglucosamine [PNAG]), which is a major component of staphylococcal biofilms.97 Although our study did not demonstrate pronounced differences among the biofilm-producing capabilities of the locally collected MSSA and MRSA isolates, there have been studies offering possible biological explanations to this phenomenon.38 It has been suggested that biofilm-formation in MSSA is mediated by cell-cell adhesion via the production of PIA (encoded by icaADBC), while MRSA biofilm-production is PIA-independent, and rather, it is dependent on a protein adhesion, which is negatively regulated by the agr system.98,99 O’Neill et al showed that media supplementation with NaCl results in the induction of biofilm-production in MSSA only, as this activated the expression of the ica operon.100 In contrast, Croes et al showed that the presence of excess glucose in the media represses the agr system (through the generation of low pH), which resulted in the induction of biofilm-production in MRSA only;81,101 additionally, the deletion of the agr system also enhanced the biofilm-production of MRSA isolates, while it had no effect on its methicillin-susceptible counterparts.102
Conclusions
The production of biofilm by pathogenic bacteria in vivo provides important protection from external forces and antimicrobials, in addition to facilitating chronicity. S. aureus is an exceptionally adaptable pathogen both in natural environment and in clinical situations. Biofilm-formation in both MSSA and MRSA isolates is an important step in the pathogenesis of implant-associated infections and leads to a synergistic interaction between “classical” resistance-determinants and the inability of antibiotics and immune cells to reach S. aureus isolates. The relationship between the MDR phenotype and biofilm-positivity has been studied for many relevant pathogens, however, the culmination of these results is inconclusive. In our study, one hundred and fifty MSSA and one hundred and fifty MRSA isolates (from a variety of clinical situations) were tested for their antibiotic-susceptibility and their biofilm-forming capacity to ascertain a possible relationship between the two. Among our tested isolates, we have found complete susceptibility to the last-resort agents, while there were significant differences in the resistance rates between MSSA and MRSA isolates regarding almost all other, commonly used agents. Thirty-eight percent and 42% of isolates were biofilm-producers based on the CV tube adherence assay and the CRA plate methods, respectively. Overall, no association was found between methicillin-resistance and biofilm-positivity in our settings; on the other hand, resistant isolates to erythromycin, clindamycin and rifampin were significantly more common among biofilm-producers.
Our study possesses some limitations: i) the cross-sectional nature of the study: although isolates were collected from different clinical specimen groups, they may not represent Hungarian S. isolates, their biofilm-forming capacity or susceptibility overall; ii) selection bias: isolates usually originated from tertiary-care centers, corresponding to patients with more severe conditions or underlying illnesses; iii) interpretation: both phenotypic methods were evaluated by organoleptic methods, therefore the reading of the results was dependent on the expertise of the researchers; iv) lack of molecular methods: the molecular characterization of resistance determinants, clonal lineages (with PFGE or MLST) or genetic determinants of biofilm-production (eg, agr, ica genes) in the mentioned isolates was not performed. Nevertheless, our study provides additional data to the existing pool of literature on the association of drug resistance and biofilm-formation in S. aureus. Additional studies – with the inclusion of other isolates and utilization – are needed to provide clarity on this subject.
Data Sharing Statement
All data generated during the study are presented in this paper.
Funding
The article processing charge (APC) was funded by the University of Szeged Open Access fund (ID: 5175). M.G. was supported by the János Bolyai Research Scholarship (BO/00144/20/5) of the Hungarian Academy of Sciences and the New National Excellence Programme (ÚNKP-20-5-SZTE-330) of the Ministry of Human Resources. Support from Ministry of Human Capacities, Hungary grant 20391-3/2018/FEKUSTRAT is acknowledged. M.G. would also like to acknowledge the support of ESCMID’s “30 under 30” Award.
Disclosure
The authors declare no conflict of interest, monetary or otherwise. The authors alone are responsible for the content and writing of this article.
References
1. Shaw C, Stitt JM, Cowan ST. Staphylococci and their Classification. J Gen Microbiol. 1951;5:1010–1023. doi:10.1099/00221287-5-5-1010
2. Tong SYC, Davis JS, Eichenberger E, Holland TL, Fowler VG. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28:603–661. doi:10.1128/CMR.00134-14
3. Gould D, Chamberlaine A. Staphylococcus aureus: a review of the literature. J Clin Nurs. 1995;4:5–12. doi:10.1111/j.1365-2702.1995.tb00004.x
4. Gajdács M. The continuing threat of methicillin-resistant Staphylococcus aureus. Antibiotics. 2019;8:e52. doi:10.3390/antibiotics8020052
5. Kahl BC, Becker K, Löffler B. Clinical significance and pathogenesis of staphylococcal small colony variants in persistent infections. Clin Microbiol Rev. 2016;29:401–427. doi:10.1128/CMR.00069-15
6. Ericson JE, Popoola VO, Smith PB, et al. Burden of invasive Staphylococcus aureus infections in hospitalized infants. JAMA Pediatr. 2015;169:1105–1111. doi:10.1001/jamapediatrics.2015.2380
7. Chambers HF. The changing epidemiology of Staphylococcus aureus. Emerg Infect Dis. 2001;7:178–182. doi:10.3201/eid0702.010204
8. Enright MC, Robinson DA, Randle G, Feil EJ, Grundmann H, Spratt BG. The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA). Proc Natl Acad Sci USA. 2002;99:7687–7692. doi:10.1073/pnas.122108599
9. David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev. 2010;23:616–687.
10. Algammal AM, Hetta HF, Elkelish A, et al. Methicillin-resistant Staphylococcus aureus (MRSA): one health perspective approach to the bacterium epidemiology, virulence factors, antibiotic-resistance, and zoonotic impact. Infect Drug Res. 2020;13:3255–3265. doi:10.2147/IDR.S272733
11. Dulon M, Haarnmann F, Peters C, Schablon A, Nienhaus A. MRSA prevalence in European healthcare settings: a review. BMC Infect Dis. 2011;11:e138. doi:10.1186/1471-2334-11-138
12. Kang C-I, Song J-H, Ko KS, Chung DR, Peck KR. Clinical features and outcome of Staphylococcus aureus infection in elderly versus younger adult patients. Int J Infect Dis. 2011;15:e58–e62. doi:10.1016/j.ijid.2010.09.012
13. Gajdács M. The concept of an ideal antibiotic: implications for drug design. Molecules. 2019;24:892. doi:10.3390/molecules24050892
14. Stefani S, Chung DR, Lindsay JA, et al. Meticillin-resistant Staphylococcus aureus (MRSA): global epidemiology and harmonisation of typing methods. Int J Antimicrob Agents. 2012;39:273–282. doi:10.1016/j.ijantimicag.2011.09.030
15. Arzanlou M, Chai WC, Venter H. Intrinsic, adaptive and acquired antimicrobial resistance in Gram-negative bacteria. Essays Biochem. 2017;61:49–59. doi:10.1042/EBC20160063
16. Lebeaux D, Ghigo J-M, Beloin C. Biofilm-related infections: bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol Mol Boil Rev. 2014;78:510–543.
17. Henrici AT. Studies of freshwater bacteria: I. A direct microscopic technique. J Bacteriol. 1933;25:277–287. doi:10.1128/JB.25.3.277-287.1933
18. Bryers JD. Medical biofilm. Biotechnol Bioeng. 2008;100:1–18. doi:10.1002/bit.21838
19. Stewart PS. Mechanisms of antibiotic resistance in bacterial biofilms. Int J Med Microbiol. 2002;292:107–113. doi:10.1078/1438-4221-00196
20. Artini M, Papa R, Scoarughi GL, et al. Comparison of the action of different proteases on virulence properties related to the staphylococcal surface. J Appl Microbiol. 2013;114:266–277. doi:10.1111/jam.12038
21. Tan X, Qin N, Wu C, et al. Transcriptome analysis of the biofilm formed by methicillin-susceptible Staphylococcus aureus. Sci Rep. 2015;5:e11997. doi:10.1038/srep11997
22. Chatterjee S, Maiti P, Dey R, Kundu A, Dey R. Biofilms on indwelling urologic devices: microbes and antimicrobial management prospect. Ann Med Health Sci Res. 2014;4:100–104. doi:10.4103/2141-9248.126612
23. Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science. 1995;284:1318–1322. doi:10.1126/science.284.5418.1318
24. Craft KM, Nyugen JM, Berg LJ, Townsend SD. Methicillin-resistant Staphylococcus aureus (MRSA): antibi-otic-resistance and the biofilm phenotype. Med Chem Comm. 2019;10:1231–1241. doi:10.1039/C9MD00044E
25. Singh R, Ray P, Das A, Sharma M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. J Antimicrob Chemother. 2010;65:1955–1958. doi:10.1093/jac/dkq257
26. Soto SM. Importance of biofilms in urinary tract infections: new therapeutic approaches. Adv Biol. 2014;2014:e543974. doi:10.1155/2014/543974
27. Wu H, Moser C, Wang H-Z, Høiby N, Song Z-J. Strategies for combating bacterial biofilm infections. Int J Oral Sci. 2015;7:1–7. doi:10.1038/ijos.2014.65
28. Holby N. A short history of microbial biofilms and biofilm infections. APMIS. 2017;125:272–275.
29. Stájer A, Barrak I, Gajdács M, Urbán E, Baráth Z. Diagnosis and management of cervicofacial actinomycosis: lessons from two distinct clinical cases. Antibiotics. 2020;9:e139. doi:10.3390/antibiotics9040139
30. Silva-Santana G, Cabral-Oliviera G, Oliveira DR, Nogueira BA, Pereira-Ribeiro PMA, Mattos-Guaraldi AL. Staphylococcus aureus biofilms: an opportunistic pathogen with multidrug resistance. Rev Med Microbiol. 2020. doi:10.1097/MRM.0000000000000223
31. Boucher HW, Talbot GH, Bradley JS, et al. Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America. Clin Infect Dis. 2009;48:1–12. doi:10.1086/595011
32. McCarthy H, Rudkin KJ, Black NS, Gallagher L, O’Neill E, O’Gara JP. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front Cell Infect Microbiol. 2015;5:e1. doi:10.3389/fcimb.2015.00001
33. Behzadi P, Urbán E, Gajdács M. Association between biofilm-production and antibiotic resistance in uropathogenic Escherichia coli (UPEC): an in vitro study. Diseases. 2020;8:17. doi:10.3390/diseases8020017
34. Nirwati H, Sinanjung K, Fahrunissa F, et al. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc. 2019;13:e20. doi:10.1186/s12919-019-0176-7
35. Rodulfo H, Acria A, Hernández A, et al. Virulence factors and integrons are associated with MDR and XDR phenotypes in nosocomial strains of Pseudomonas aeruginosa in a Venezuelan university hospital. Rev Inst Med Trop Sao Paolo. 2019;61:e20. doi:10.1590/s1678-9946201961020
36. de Campos PA, Royer S, Batistao DW, et al. Multi-drug resistance related to biofilm formation in Acinetobacter baumannii and Klebsiella pneumoniae clinical strains from different Pulsotypes. Curr Microbiol. 2016;72:617–627. doi:10.1007/s00284-016-0996-x
37. Hashem YA, Amin HM, Essam TM, Yassin AS, Aziz RK. Biofilm formation in enterococci: genotype-phenotype correlations and inhibition by vancomycin. Sci Rep. 2017;7:5733. doi:10.1038/s41598-017-05901-0
38. Belbase A, Pant ND, Nepal K, et al. Antibiotic resistance and biofilm production among the strains of Staphylococcus aureus isolated from pus/wound swab samples in a tertiary care hospital in Nepal. Ann Clin Microbiol Antimicrob. 2017;16:e15. doi:10.1186/s12941-017-0194-0
39. Lee JS, Bae YM, Han A, Lee SY. Development of Congo red broth method for the detection of biofilm-forming or slime-producing Staphylococcus sp. Food Sci Technol. 2016;73:707–714.
40. Gajdács M, Ábrók M, Lázár A, Burián K. Increasing relevance of Gram‑positive cocci in urinary tract infections: a 10‑year analysis of their prevalence and resistance trends. Sci Rep. 2020;10. doi:10.1038/s41598-020-7483.
41. Gajdács M, Urbán E. Epidemiology and resistance trends of Staphylococcus aureus isolated from vaginal samples: a 10-year retrospective study in Hungary. Acta Dermatovenerol Alpina Pannon Adriatica. 2019;28:143–147.
42. Dumaru R, Baral R, Shrestha LB. Study of biofilm formation and antibiotic resistance pattern of gram-negative Bacilli among the clinical isolates at BPKIHS, Dharan. BMC Res. 2019;12:38. doi:10.1186/s13104-019-4084-8
43. Melo PC, Ferreira LM, Filho AN, Zafalon LF, Vicente HIG, de Souza V. Comparison of methods for the detection of biofilm formation by Staphylococcus aureus isolated from bovine subclinical mastitis. Braz J Microbiol. 2013;44:119–124. doi:10.1590/S1517-83822013005000031
44. Greenwood PE, Nikulin MS. A Guide to Chi-Squared Testing. New York: Wiley; 1996.
45. Zhen X, Lundborg CS, Zhang M, et al. Clinical and economic impact of methicillin-resistant Staphylococcus aureus: a multicentre study in China. Sci Rep. 2020;10:e3900. doi:10.1038/s41598-020-60825-6
46. Chen L, Tang ZY, Cui SY, et al. Biofilm production ability, virulence and antimicrobial resistance genes in Staphylococcus aureus from various veterinary hospitals. Pathogens. 2020;9:e264. doi:10.3390/pathogens9040264
47. Amorena B, Gracia E, Monzón M, et al. Antibiotic susceptibility assay for Staphylococcus aureus in biofilms developed in vitro. J Antimicrob Chemother. 1999;44:43–55. doi:10.1093/jac/44.1.43
48. Grice EA, Segre JA. The skin microbiome. Nat Rev Microbiol. 2011;9:244–253. doi:10.1038/nrmicro2537
49. Lister LJ, Horswill AR. Staphylococcus aureus biofilms: recent developments in biofilm dispersal. Front Cell Infect Microbiol. 2014;4:e178. doi:10.3389/fcimb.2014.00178
50. Verderosa AD, Totsika M, Fairfull-Smith KE. Bacterial biofilm eradication agents: a current review. Front Chem. 2019;7:824. doi:10.3389/fchem.2019.00824
51. Delcaru C, Alexandru I, Podgoreanu P, et al. Microbial biofilms in urinary tract infections and prostatitis: etiology, pathogenicity, and combating strategies. Pathogens. 2016;5:65. doi:10.3390/pathogens5040065
52. Dakheel KH, Rahim RA, Neela VK, Al-Obaidi JR, Hun TG, Yusoff K. Methicillin-resistant Staphylococcus aureus biofilms and their influence on bacterial adhesion and cohesion. BioMed Res Int. 2016;2016:e4708425. doi:10.1155/2016/4708425
53. Christensen GD, Simpson WA, Bisno AL, Beachey EH. Adherence of slime producing strains of Staphylococcus epidermidis to smooth surfaces. Infect Immun. 1982;37:318–326. doi:10.1128/IAI.37.1.318-326.1982
54. Freeman DJ, Falkiner FR, Keane CT. New method for detecting slime production by coagulase negative staphylococci. J Clin Pathol. 1989;42:872–874. doi:10.1136/jcp.42.8.872
55. Zimmerli W, Sendi P. Role of rifampin against Staphylococcal biofilm infections in vitro, in animal models, and in orthopedic-device-related infections. Antimicrob Agents Chemother. 2019;2:e01746–e01718.
56. Yu W, Hallinen KM, Wood KB. Interplay between antibiotic efficacy and drug-induced lysis underlies enhanced biofilm formation at subinhibitory drug concentrations. Antimicrob Agents Chemother. 2018;62:e01603–e01617. doi:10.1128/AAC.01603-17
57. Penesyan A, Paulsen IT, Gillings MR, Kjelleberg S, Manefieldm MJ. Secondary effects of antibiotics on microbial biofilms. Front Microbiol. 2020;11:e2109. doi:10.3389/fmicb.2020.02109
58. Lima-e-Silva AA, Silva-Filho RG, Fernandes HMZ, et al. Sub-inhibitory concentrations of rifampicin strongly stimulated biofilm production in S. aureus. Open Microbiol J. 2017;11:142–151. doi:10.2174/1874285801711010142
59. Seneviratne CJ, Suriyanarayanan T, Swarup S, Chia KHB, Nagarajan N, Zhang C. Transcriptomics analysis reveals putative genes involved in biofilm formation and biofilm-associated drug resistance of Enterococcus faecalis. J Endodont. 2017;43:949–955. doi:10.1016/j.joen.2017.01.020
60. Rachid S, Ohlsen K, Witte W, Hacker J, Ziebuhr W. Effect of subinhibitory antibiotic concentrations on polysaccharide intercellular adhesin expression in biofilm-forming Staphylococcus epidermidis. Antimicrob Agents Chemother. 2000;44:3357–3363. doi:10.1128/AAC.44.12.3357-3363.2000
61. Arslan S, Ozkardes F. Slime production and antibiotic susceptibility in staphylococci isolated from clinical samples. Mem Inst Oswaldo Cruz. 2007;102:29–33. doi:10.1590/S0074-02762007000100004
62. Ghasemian A, Najar Peerayeh S, Bakhshi B, Mirzaee M. Comparison of biofilm formation between Methicillin-resistant and methicillin-susceptible isolates of Staphylococcus aureus. Iran Biomed J. 2016;20:175–181. doi:10.7508/ibj.2016.03.007
63. Rodríguez-Lopez P, Filipello V, Di Ciccio PA, et al. Assessment of the antibiotic resistance profile, genetic heterogeneity and biofilm production of Methicillin-Resistant Staphylococcus aureus (MRSA) isolated from the Italian swine production chain. Foods. 2020;9:e1141. doi:10.3390/foods9091141
64. El-Nagdy AH, Abdel-Fattah GM, Emarah Z. Detection and control of biofilm formation by Staphylococcus aureus from febrile neutropenic patient. Infect Drug Res. 2020;13:3091–3101. doi:10.2147/IDR.S259914
65. Knobloch JKM, Horstkotte MA, Rodhe H, Mack D. Evaluation of different detection methods of biofilm formation in Staphylococcus aureus. Med Microbiol Immunol. 2002;191:101–106. doi:10.1007/s00430-002-0124-3
66. Mathur T, Singhal S, Khan S, Upadhyay DJ, Fatma T, Rattan A. Detection of biofilm formation among the clinical isolates of Staphylococci: an evaluation of three different screening methods. Indian J Med Microbiol. 2006;24:25–29. doi:10.4103/0255-0857.19890
67. Bose S, Khodke M, Basak S, Mallick SK. Detection of biofilm-producing staphylococci: need of the hour. J Clin Diagn Res. 2009;3:1915–1920.
68. Piechota M, Kot B, Frankowska-Maciejewska A, Gruzewska A, Wozniak-Kosek A. Biofilm formation by methicil-lin-resistant and methicillin-sensitive Staphylococcus aureus strains from hospitalized patients in Poland. BioMed Res Int. 2018;2018:e4657396. doi:10.1155/2018/4657396
69. Cha JO, Yoo JI, Yoo JS, et al. Investigation of biofilm formation and its association with the molecular and clinical characteristics of methicillin-resistant Staphylococcus aureus. Osong Public Health Res Perspect. 2013;4:225–232. doi:10.1016/j.phrp.2013.09.001
70. Souli M, Giamarellou H. Effects of slime produced by clinical isolates of coagulase negative Staphylococci on activities of various antimicrobial agents. Antimicrob Agents Chemother. 1998;42:939–941. doi:10.1128/AAC.42.4.939
71. Agarwal A, Jain A. Glucose and sodium chloride induced biofilm production and ica operon in clinical isolates of staphylococci. Indian J Med Res. 2013;138:262–266.
72. De Araujo GL, Coelho RL, de Carvalho CB, et al. Commensal isolates of methicillin-resistant Staphylococcus epidermidis are also well equipped to produce biofilm on polystyrene surfaces. J Antimicrob Chemother. 2006;57:855–864. doi:10.1093/jac/dkl071
73. Zhang Y, Xu D, Shi L, Li C, Yan H. Association between agr type, virulence factors, biofilm formation and antibiotic resistance of Staphylococcus aureus isolates from pork production. Front Microbiol. 2018;9:e1876. doi:10.3389/fmicb.2018.01876
74. Manandhar S, Singh A, Varma A, Pandey S, Shirvastava N. Biofilm producing clinical Staphylococcus aureus isolates augmented prevalence of antibiotic resistant cases in tertiary care hospitals of Nepal. Front Microbiol. 2018;9:e2749. doi:10.3389/fmicb.2018.02749
75. Bhattacharya S, Bir R, Majumdar T. Evaluation of multidrug resistant Staphylococcus aureus and their association with biofilm production in a Tertiary Care Hospital, Tripura, Northeast India. J Clin Diagn Microbiol. 2015;9:DC01–DC04.
76. Neopane P, Nepal HP, Shrestha R, Uehara O, Abiko Y. In vitro biofilm formation by Staphylococcus aureus isolated from wounds of hospital-admitted patients and their association with antimicrobial resistance. Int J Gen Med. 2018;11:25–32. doi:10.2147/IJGM.S153268
77. Conceicao T, Aires-de-sousa M, Füzi M, et al. Replacement of methicillin-resistant Staphylococcus aureus clones in Hungary over time: a 10-year surveillance study. Clin Microbiol Infect. 2007;13:971–979. doi:10.1111/j.1469-0691.2007.01794.x
78. Hanczvikkel A. [Multidrog-rezisztens baktériumok túlélése textíliákon: a környezeti körülmények és az antibakteriális hatóanyagok hatása] (in Hungarian). PhD thesis. University of Óbuda; 2018. Available from: http://lib.uni-obuda.hu/sites/lib.uni-obuda.hu/files/Hanczvikkel_Adrienn_ertekezes.pdf.
79. Füzi M, Bano JR, Tóth Á. Global evolution of pathogenic bacteria with extensive use of fluoroquinolone agents. Front Microbiol. 2020;11:e271. doi:10.3389/fmicb.2020.00271
80. Huang CY, Ho CF, Chen CJ, Su LH, Lin TY. Comparative molecular analysis of community-associated and healthcare-associated methicillin-resistant Staphylococcus aureus isolates from children in northern Taiwan. Clin Microbiol Infect. 2008;14:1167–1172.
81. Croes S, Deurenberg RH, Boumans ML, Beisser PS, Neef C, Stobberingh EE. Staphylococcus aureus biofilm formation at the physiologic glucose concentration depends on the S. aureus lineage. BMC Microbiol. 2009;9:e229.
82. Luther MK, Parente DM, Caffrey AR, et al. Clinical and genetic risk factors for biofilm-forming Staphylococcus aureus. Antimicrob Agents Chemother. 2018;62:e02252–e02217. doi:10.1128/AAC.02252-17
83. Recker M, Laabei M, Toleman MS, et al. Clonal differences in Staphylococcus aureus bacteraemia-associated mortality. Nat Microbiol. 2017;2:1381–1388. doi:10.1038/s41564-017-0001-x
84. Lim Y, Shin HJ, Kwon AS, Reu JH, Park G, Kim J. Predictive genetic risk markers for strong biofilm-forming Staphylococcus aureus: fnbB gene and SCCmec type III. Diagn Microbiol Infect Dis. 2013;76:539–541. doi:10.1016/j.diagmicrobio.2013.04.021
85. Da fonseca batistao DW, de Campos PA, Camilo NC, et al. Biofilm formation of Brazilian meticillin-resistant Staphylococcus aureus strains: prevalence of biofilm determinants and clonal profiles. J Med Microbiol. 2016;65:286–297. doi:10.1099/jmm.0.000228
86. Pozzi C, Waters EM, Rudkin JK, et al. Methicillin resistance alters the biofilm phenotype and attenuates virulence in Staphylococcus aureus device-associated infections. PLoS Pathog. 2012;8:e1002626. doi:10.1371/journal.ppat.1002626
87. Periasamy S, Joo HS, Duong AC, et al. How Staphylococcus aureus biofilms develop their characteristic structure. Proc Natl Acad Sci USA. 2012;109:1281–1286. doi:10.1073/pnas.1115006109
88. Roilides M, Simitsopoulou M, Katragkou A, Walsh TJ. How biofilms evade host defenses. Microbiol Spectr. 2013;3. doi:10.1128/microbiolspec.MB-0012-2014.
89. Archer NK, Mazaitis MJ, Costerton JW, Leid JG, Powers ME, Shirtliff ME. Staphylococcus aureus biofilms: properties, regulation and roles in human disease. Virulence. 2011;2:445–459. doi:10.4161/viru.2.5.17724
90. Chua SL, Ding Y, Liu Y, et al. Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels. Open Biol. 2016;6:e160162. doi:10.1098/rsob.160162
91. Garcia LG, Lemaire S, Kahl BC, et al. Antibiotic activity against small-colony variants of Staphylococcus aureus: review of in vitro, animal and clinical data. J Antimicrob Chemother. 2013;68:1455–1464. doi:10.1093/jac/dkt072
92. Burián K, Endrész V, Deák J, et al. Transcriptome analysis indicates an enhanced activation of adaptive and innate immunity by chlamydia-infected murine epithelial cells treated with interferon γ. J Infect Dis. 2010;9:1405–1414. doi:10.1086/656526
93. Balogh EP, Faludi I, Virók DP, Endrész V, Burián K. Chlamydophila pneumoniae induces production of the defensin-like MIG/CXCL9, which has in vitro anti-chlamydial activity. Int J Med Microbiol. 2011;301:252–259. doi:10.1016/j.ijmm.2010.08.020
94. Sugimoto S, Sato F, Miyakawa R, et al. Broad impact of extracellular DNA on biofilm formation by clinically isolated Methicillin-resistant and -sensitive strains of Staphylococcus aureus. Sci Rep. 2018;8:e2554. doi:10.1038/s41598-018-20485-z
95. Cramton SE, Gerke C, Schnell NF, Nichols WW, Gotz F. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation. Infect Immun. 1999;67:5427–5433. doi:10.1128/IAI.67.10.5427-5433.1999
96. Racenis K, Kroica J, Rezevska D, et al. S. aureus colonization, biofilm production, and phage susceptibility in peritoneal dialysis patients. Antibiotics. 2020;9:e582. doi:10.3390/antibiotics9090582
97. Jain A, Agarwal A. Biofilm production, a marker of pathogenic potential of colonizing and commensal staphylococci. J Microbiol Methods. 2009;76:88–92. doi:10.1016/j.mimet.2008.09.017
98. Boles BR, Horswill AR. Agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008;4:e1000052. doi:10.1371/journal.ppat.1000052
99. Moretro T, Hermansen L, Holck AL, Sidhu MS, Rudi K, Langrrud S. Biofilm formation and the presence of the intercellular adhesion locus ica among staphylococci from food and food processing environments. Appl Environ Microbiol. 2003;69:5648–5655. doi:10.1128/AEM.69.9.5648-5655.2003
100. O’Neill E, Pozzi C, Houston P, et al. Association between methicillin susceptibility and biofilm regulation in Staphylococcus aureus isolates from device-related infections. J Clin Microbiol. 2007;45:1379–1388. doi:10.1128/JCM.02280-06
101. O’Gara JP. ica and beyond: biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol Lett. 2014;270:179–188. doi:10.1111/j.1574-6968.2007.00688.x
102. Aricola CR, Baldassarri L, Montanaro L. Presence of icaA and icaD genes and slime production in a collection of staphylococcal strains from catheter-associated infections. J Clin Microbiol. 2001;39:2151–2156. doi:10.1128/JCM.39.6.2151-2156.2001
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.