Back to Journals » Journal of Pain Research » Volume 18
Context-Dependent Roles of Four Classes of Bioactive Lipids in Neuroglia-Mediated Regulation of Neuroinflammation
Authors Sun S, Hu S, Wang Y, Sun M, Zhao L
Received 23 April 2025
Accepted for publication 5 August 2025
Published 18 August 2025 Volume 2025:18 Pages 4139—4149
DOI https://doi.org/10.2147/JPR.S536220
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Robert B. Raffa
Shiqi Sun,1 Shuangyuan Hu,1 Yuyan Wang,1 Mingsheng Sun,1 Ling Zhao1,2
1Acupuncture and Tuina School, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan, People’s Republic of China; 2Key Laboratory of Acupuncture and Moxibustion in Prevention and Treatment of Geriatric Diseases (Chengdu University of Traditional Chinese Medicine), Ministry of Education, Chengdu, Sichuan, People’s Republic of China
Correspondence: Ling Zhao, Acupuncture and Tuina School, Chengdu University of Traditional Chinese Medicine, 1166 Liutai Avenue, Chengdu, Sichuan, People’s Republic of China, Email [email protected]
Abstract: Neuroinflammation is an adaptive immune response of the nervous system to tissue injury or microbial invasion. As innate immune cells of the central nervous system, neuroglia cells—especially astrocytes and microglia—play a critical role in maintaining neural homeostasis. Bioactive lipids, key molecules at the core of tissue immune homeostasis, regulate a variety of biological activities with precision, participating in the management of neuroinflammation, a process often mediated by glial cells. The four major classes of bioactive lipids, including classic eicosanoids, specialized pro-resolving mediators (SPMs), endocannabinoids (eCBs), and lysophospholipids/sphingolipids (LPLs/SPs), influence the regulation of neuroinflammation in a context-dependent manner by affecting the phenotypic changes and activation states of neuroglial cells, thus collectively contributing to the modulation of neuroinflammation. A comprehensive review and summary of these regulatory mechanisms will provide potential directions for future research integrating emerging single-cell lipidomics techniques and clinical trials of lipid-based therapeutics. In-depth exploration of this field holds great promise for offering novel strategies for the early diagnosis and treatment of neurological disorders.
Keywords: neuroinflammation, bioactive lipids, microglia, astrocytes
Introduction
Neuroinflammation is an adaptive immune response of the nervous system to tissue damage or microbial invasion, characterized by the release of inflammatory mediators by neuroglia cells and other immune cells to counter various harmful stimuli.1 Although transient neuroinflammation plays a protective role, prolonged and chronic neuroinflammation becomes a critical trigger for many neurological diseases. For instance, neuroinflammatory-related disorders such as Alzheimer’s disease, Parkinson’s disease, and migraines are among the most common, all of which are associated with high disability rates and impose significant economic and healthcare burdens.2–4 Therefore, exploring effective early intervention strategies to control the progression of neuroinflammation is crucial to preventing further damage.
As the innate immune cells of the central nervous system (CNS), neuroglia cells are the guardians of neural homeostasis, and their importance in the management of neuroinflammation is well-recognized. Astrocytes and microglia are the primary effector cells in neuroinflammation. They directly participate in and regulate the occurrence and progression of neuroinflammation, making them key targets in neuroinflammation research. Traditionally, lipids have been viewed as structural components of cell membranes and sources of efficient energy. However, bioactive lipids, situated at the core of tissue immune homeostasis, actually form a complex molecular network fundamentally involved in every key aspect of inflammation, including its initiation, coordination, and resolution.5 Although existing research has provided evidence linking bioactive lipids with neuroinflammatory diseases,6,7 their specific roles in the progression of inflammation from acute to chronic stages have not yet been fully elucidated.
Interestingly, an increasing body of research evidence suggests that bioactive lipids, including classic eicosanoids, specialized pro-resolving mediators (SPMs), endocannabinoids (eCBs) and lysophospholipids/sphingolipids (LPLs/SPs), may participate in the management of neuroinflammation under the mediation of neuroglia cells.8 These lipids modulate neuroinflammation by influencing the phenotypic transformation and the activation or inhibition states of neuroglial cells. With the rapid advancement of cutting-edge technologies such as single-cell multi-omics, the diversity of microglia and astrocytes in neuroinflammation is gradually being unveiled. The transcriptional programs of microglia (eg, disease-associated, interferon-responsive, lipid-droplet-enriched) and astrocytes (eg, reactive A1, neuroprotective A2) play crucial roles at various stages of inflammation.9,10 For instance, Trem2-deficient microglia exhibit characteristics of lipid-droplet-enriched microglia (LDAM),11,12 and their altered lipid metabolism is closely linked to the progression of neurodegenerative diseases. LDAM microglia modulate lipid metabolic pathways that not only intensify inflammatory responses but also regulate interactions with astrocytes.13 Furthermore, the metabolism of long-chain polyunsaturated fatty acids (PUFAs) in astrocytes plays a pivotal role in neuroinflammation, further confirming the close connection between lipid metabolism and the functional states of glial cells.14 These findings highlight the essential role of lipid metabolism in the functional transitions of microglia and astrocytes, providing new perspectives for the treatment of neuroinflammation and neurodegenerative diseases. In our team’s previous research, we have also observed a close correlation between the accumulation of lipid droplets in microglia and the onset of neuroinflammation. These results further support our hypothesis that bioactive lipids, under the regulation of neuroglial cells, particularly microglia and astrocytes, may play a key role in various stages of neuroinflammation. This review aims to explore how bioactive lipids interact with neuroglial cells and co-regulate the entire process from inflammation induction to chronic transformation. This perspective not only deepens our understanding of the mechanisms underlying neuroinflammation but also provides valuable insights for the early diagnosis and therapeutic intervention of neurodegenerative diseases.
The Complex Role of Microglia and Astrocytes in Neuroinflammation
The significance of astrocytes and microglia in neuroinflammation, along with the extensive research on these cells, makes them the focal points of our review. Numerous studies have elucidated their specific mechanisms and roles in neuroinflammation, providing a wealth of data and theoretical foundation. To ensure the depth and focus of this review, we have chosen to concentrate on astrocytes and microglia.
Microglia, as the innate immune cells of the CNS, primarily serve roles in immune defense and maintaining internal homeostasis. Under normal conditions, these cells remain in a resting state, but pathological stimuli can activate them, marking the onset of neuroinflammation.15 Activated microglia exhibit different functional states, which play distinct roles in the inflammatory response, including both pro-inflammatory and anti-inflammatory regulatory functions.16 However, excessive or prolonged activation can lead to the continuous release of inflammatory mediators such as pro-inflammatory cytokines and chemokines, thereby exacerbating inflammation and tissue damage.17 Therefore, regulating the activation of microglia and their release of inflammatory mediators is crucial for controlling neuroinflammation and the progression of related diseases.
Astrocytes are the most abundant cell type in the CNS, responsible for critical functions such as the synthesis and recycling of neurotransmitters, regulation of neuronal metabolism, and defense against oxidative stress.18 There is a close interaction between these cells and microglia, where the activation of microglia, through the secretion of IL-1α, TNFα, and C1q, drives a neurotoxic astrocyte profile, characterized by upregulation of complement component C3. In contrast, TGF-β and IL-10 promote the formation of a neuroprotective astrocyte phenotype. This interaction between microglia and astrocytes plays a critical role in neuroinflammation and disease progression.19–21 For example, astrocytes induced by activated microglia may exhibit various functional states, which could either exacerbate or alleviate the progression of disease. Certain astrocytes may exhibit pro-inflammatory functions in response to neural injury, while others may provide protective effects by expressing neurotrophic factors or anti-inflammatory genes.22 Therefore, a deeper understanding of the physiological and anti-inflammatory mechanisms of astrocytes and exploring regulatory strategies targeted at these cells represent an important research direction for controlling neuroinflammation.
Recent studies suggest that neuroinflammation may be caused by acute or chronic metabolic dysregulation and energy depletion, where neuroglial cells play a central role in controlling neuroinflammation by regulating the metabolic processes of neural tissues.23 Additionally, the bioactive lipids produced by neuroglial cells not only coordinate the normal physiological processes of the CNS but are also involved in various neuroinflammatory conditions and related diseases, offering new perspectives for researching their roles in these disorders.24,25
The Multifaceted Roles of Bioactive Lipids Mediated by Neuroglial Cells in Neuroinflammation
Endogenous lipids, also known as bioactive lipids, are divided into four main families based on their biochemical functions: classic eicosanoids, SPMs, eCBs and LPLs/SPs. These lipids play a crucial role in various stages of inflammation, serving different functions in the induction, resolution, and chronic transformation of inflammation.5,26 We will further explore how these lipid mediators cooperate with neuroglia cells to play these roles and participate in the management of neuroinflammation (Figure 1).
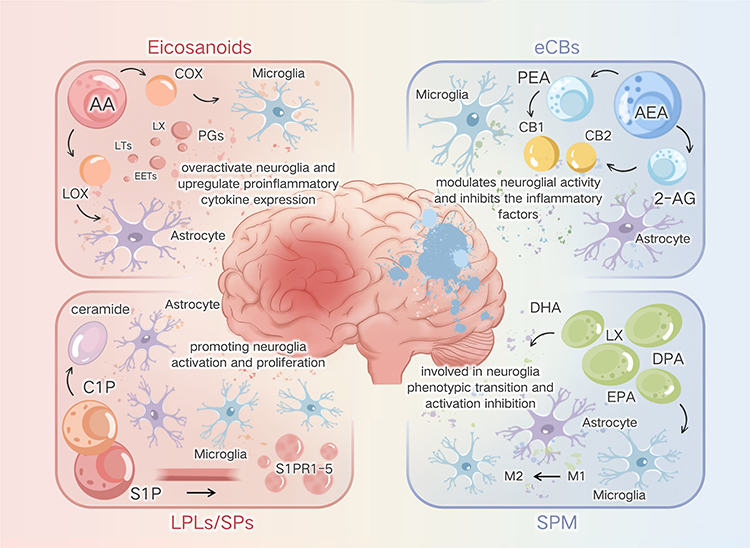 |
Figure 1 Summarizes the respective roles of bioactive lipids in resolving neuroinflammation. Classical eicosanoids act as key triggers of acute neuroinflammatory responses; SPMs are responsible for promoting homeostasis recovery after neuroinflammation; eCBs are key maintainers of tissue homeostasis post-neuroinflammation; and LPLs/sps play a crucial role in maintaining system balance during neuroinflammation. |
Classical Eicosanoids: Key Trigger of Acute Neuroinflammatory Responses
Eicosanoids are well-known bioactive lipids that are rapidly produced in response to tissue damage or infection to combat harmful stimuli. Due to their potent pro-inflammatory effects, they are considered critical triggers of acute inflammation. Among them, omega-6 PUFAs, specifically arachidonic acid (AA), serve as a common precursor for many molecules. Released from cell membrane phospholipids by phospholipase A2 (PLA2), AA is transformed by lipoxygenases (LOX) and cyclooxygenases (COX) into various bioactive molecules, examples include leukotrienes (LTs), prostaglandins (PGs), and Hydroxyeicosatetraenoic acids (HETEs). These molecules not only play roles in regulating vascular tone and pain perception but are also key in the initiation of inflammation. For instance, Prostaglandin E2 (PGE2) is one of the major metabolites of arachidonic acid through the COX pathway. Studies have demonstrated that it amplifies the inflammatory response in the central nervous system by activating downstream signaling pathways, such as EP2/NOX2/NF-κB.27 Similarly, TBs has been implicated in the regulation of blood-brain barrier permeability and the recruitment of immune cells during neuroinflammation.28 These findings provide substantial evidence supporting the critical role of arachidonic acid-derived lipid mediators in modulating neuroinflammatory processes.
In the CNS, the metabolic products of PUFAs play a crucial role in the activation of neuroglia cells, particularly in the onset of acute neuroinflammation.29 For instance, metabolites of AA, including PGE2 produced by COX, act as immunomodulators that can either trigger or reduce inflammation by modulating the COX2/PGE2 signaling pathway.30,31 Moreover, astrocytes and microglia are major sources of PGE2 and other prostaglandins within the CNS. Overexpression of PGE2 can lead to hyperactivation of microglia, exacerbating neuroinflammation.32,33 Reactive microglia release chemokines and prostaglandins that further stimulate the production of more prostaglandins through the activation of PLA2 and COX, creating a vicious cycle of inflammation.26,34
However, acute inflammation is not a constant state. As the inflammation gradually resolves, the levels of classic eicosanoids decrease and undergo a transformation in type. Taking lipoxins (LX) as an example, they typically have anti-inflammatory effects and, as SPMs, help to facilitate the resolution of subsequent inflammation and tissue repair.
SPM: Key Players in Promoting Homeostasis Recovery After Neuroinflammation
During the resolution phase of inflammation, previously accumulated eicosanoids are transformed into newly discovered SPMs. These SPMs play a pivotal role in resolving inflammation. They not only induce the cessation of white blood cell infiltration and recruitment but also enhance the phagocytosis of apoptotic cells and debris by macrophage. Furthermore, they suppress pro-inflammatory cytokines and stimulate the production of anti-inflammatory mediators, thereby facilitating tissue regeneration and healings.35 These SPMs are primarily produced from omega-6 AA and omega-3 fatty acids such as eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), and docosapentaenoic acid (DPA). For instance, lipoxins such as LXA4 and LXB4, derived from AA, were among the first SPMs identified. They effectively mitigate inflammation and initiate tissue repair by inhibiting inflammatory responses and restoring tissue homeostasis.36 EPA serves as a precursor for the E-series resolvins (such as RvE1-E4), which regulate inflammatory responses by modulating key immune cells.37 Similarly, SPMs derived from DHA and DPA play crucial roles in removing cellular debris and reducing the production of pro-inflammatory mediators while enhancing the production of anti-inflammatory agents.38 Despite their diverse biosynthetic pathways and structures, these SPMs share the common feature of actively promoting physiological homeostasis during inflammation.39
Research indicates that astrocytes and microglia are the primary target cells for SPMs, whose ability to modulate the activation of these neuroglia cells underscores their pivotal role in regulating neuroinflammation.40 Thus, SPMs not only reduce the production of pro-inflammatory factors but also enhance neuroprotection and neuronal survival.40 Particularly, astrocytes, which express all the enzymes and receptors necessary for SPM biosynthesis, such as the lipoxin receptor ALX/FPR2, play a crucial role in suppressing astrocyte activation and reducing the production of pro-inflammatory factors.41,42 Combined treatments with RvE1 and LXA4 have been shown to reverse neuroinflammation by modulating the activation of both microglia and astrocytes.43 Furthermore, the anti-inflammatory and neuroprotective effects of omega-3 polyunsaturated fatty acids, such as DHA and EPA, have been shown to inhibit the overactivation of microglia and astrocytes, while promoting their shift toward an anti-inflammatory phenotype. This modulation of glial cell activity, along with the reduction of pro-inflammatory cytokines in the brain, contributes to the alleviation of neuroinflammation.44–46 SPMs mediated by microglia, such as RvD1 and MaR1, enhance the phagocytic activity of these cells, participating in the clearance of debris and dead cells in the brain, thus maintaining cerebral homeostasis and aiding in repair.23 Therefore, neuroglia cell-induced activity of SPMs is thought to be a key component in maintaining brain homeostasis and contributing to repair in the context of neuroinflammation.47
However, if the injurious stimuli persist or are not timely resolved by SPMs, the inflammation may evolve into a chronic state, complicating the response to damage-induced changes involving two other families of bioactive lipids.
eCBs: Key Maintainer of Tissue Homeostasis After Neuroinflammation
Under both acute and chronic pathological conditions, the differential expression of eCBs and cannabinoid receptors across various tissues reveals their complex role in the inflammatory process. eCBs, including anandamide (AEA), 2-arachidonoylglycerol (2-AG), and palmitoylethanolamide (PEA), are bioactive lipids derived from AA metabolism. Together with their associated enzymes and receptors, they form the eCB system, which plays a crucial role in maintaining homeostasis under physiological and pathological conditions. Particularly following acute injurious stimuli, levels of eCBs rapidly increase, activating anti-inflammatory and protective mechanisms. The functions of eCBs are primarily carried out by its family members—AEA, 2-AG, and PEA—where AEA and PEA mainly exert anti-inflammatory effects, while 2-AG has both pro-inflammatory and anti-inflammatory properties.5 Under inflammatory conditions, AEA and 2-AG are produced on demand and bind to and activate type 1 and type 2 cannabinoid receptors (CB1 and CB2).48,49 This receptor activity can reduce the release of neurotransmitters and affect the transmission of inflammatory signals in the CNS, thereby attempting to restore stability before disturbances occur.50,51
At the cellular level, neuroglial cells, particularly microglia and astrocytes, are important sources for the synthesis and release of eCBs.52 Under steady-state conditions, microglia synthesize 2-AG and AEA and express CB1 and CB2 receptors. When these cells are activated, the synthesis of eCBs increases, and the expression of CB2 receptors is upregulated, promoting the production of neuroprotective factors and reducing the release of pro-inflammatory factors, thus driving microglia towards a protective phenotype.53,54 Simultaneously, the CB1 receptor in astrocytes affects cell function by modulating calcium ion mobilization, while the CB2 receptor plays a protective role by inhibiting the release of inflammatory factors.55,56 The 2-AG produced in astrocytes acts as a signaling mediator interacting with microglia, also helping to terminate neuroglial-mediated inflammatory responses.57
However, long-term inflammation can disrupt this physiological balance, leading to the overactivation of eCBs receptors and potential interactions with other receptors, thus maintaining an inflammatory state in the body. The changes under chronic inflammation are often considered a result of eCB signal degradation in neuroglial cells.58 Nevertheless, the regulatory function of the eCB system is undoubtedly beneficial for maintaining homeostasis. It coordinates the inflammatory response of nearby cells through complex interactions with multiple receptors or targets, thereby controlling the homeostasis after neuroinflammation. Moreover, the plasticity of this system makes the timely activation or inhibition of eCBs an effective strategy for intervening and controlling neuroinflammation.
LPLs/SPs: Key to Maintaining Complex System Balance in Neuroinflammation
This class of bioactive lipids, based on glycerol or sphingolipid structures and combined with molecules like ethanolamine, choline, and ceramide, displays a high degree of molecular diversity and complexity. Lysophospholipids, as key signaling molecules, control the distribution, transport, and activation of immune cells, thus regulating adaptive immune responses and influencing the progression and resolution of inflammation.59 Additionally, sphingosine products such as sphingosine 1-phosphate (S1P) and ceramide 1-phosphate (C1P), generated through phosphorylation by sphingosine kinases (SphKs) and ceramide kinase (CerK), play significant pro-inflammatory roles and are primary active sphingolipids.60,61
S1P is one of the most well-known bioactive lipids of the family, playing a central role in inflammation through its interaction with five homologous receptors (S1PR1-5) on the S1P-S1PR axis.62,63 Moreover, S1P is a critical mediator for the transport of lymphocytes between lymphoid and non-lymphoid tissues, especially crucial in the trafficking of adaptive (T and B) cells.64 Thus, if T and B cells are triggers for certain chronic inflammations, S1P indirectly promotes the development of chronic inflammation by regulating their transport. During inflammation, S1P also alters the morphology of microglia and astrocytes, promoting their proliferation and migration and increasing the synthesis of pro-inflammatory factors, thereby exacerbating the inflammatory characteristics of neuroglia cells and perpetuating inflammation.65 The complex functions of S1P are notably evident in its interaction with the SphK1 pathway. SphK1 enhances the secretion of anti-inflammatory bioactive lipids, such as SPMs, through the COX2 pathway, which is crucial for maintaining the homeostasis of the nervous system. Elevated levels of SphK1 enhance COX2’s anti-inflammatory activity and promote the synthesis of SPMs, aiding in maintaining overall system balance. However, an imbalance between S1P and SphK1 levels may activate neuroglia cells, leading to the initiation of neuroinflammatory responses.47 Therefore, the dynamic balance between S1P and the SphK1 pathway plays a key role in controlling and maintaining the internal homeostatic environment.
The role of C1P is similarly complex. On one hand, it directly enhances the activity of classical eicosanoids, promotes the production of pro-inflammatory cytokines, and exacerbates both acute and chronic inflammatory responses. On the other hand, C1P has also been shown to negatively regulate certain pro-inflammatory factors.66,67 In the context of neuroinflammation, although an increase in ceramide levels can induce the expression of inflammatory molecules and activate astrocytes and microglia,68 short-chain ceramides in microglia exhibit inhibitory effects on certain cytokines and chemokines, highlighting its function dependent on acyl chain length and cell type.69
Overall, lysophospholipids and sphingolipids play a crucial role in maintaining biological homeostasis. Although their specific functions in neuroinflammation remain unclear, partly due to our incomplete understanding of their metabolic mechanisms, several studies have reported that S1P significantly enhances the release of pro-inflammatory cytokines. In contrast, pharmacological blockade of S1P receptors completely suppresses the release of inflammatory mediators such as TNF-α and IL-6.70 Moreover, the application of antagonists targeting lysophospholipid and sphingolipid-related receptors has been shown to be an effective strategy for inhibiting the chronic progression of inflammation.71 Therefore, while further research is required to fully elucidate the true role of these bioactive lipids in neuroinflammation, existing studies have already identified them as key bioactive lipid mediators in diseases driven by chronic inflammation.
Conclusions
Over the past many years, lipids have often been simply viewed as components of cell membranes or substances for energy storage. However, with the advancement of scientific research, modern science has revealed that lipid mediators play an important and crucial role in autonomously coordinating a wide range of biological activities and maintaining organismal homeostasis. Current research not only shows that specific lipid mediators are involved in regulating each stage of inflammation but also that these bioactive lipid families coexist and collaborate at different stages of inflammation, with complex interactions between family members (Table 1).
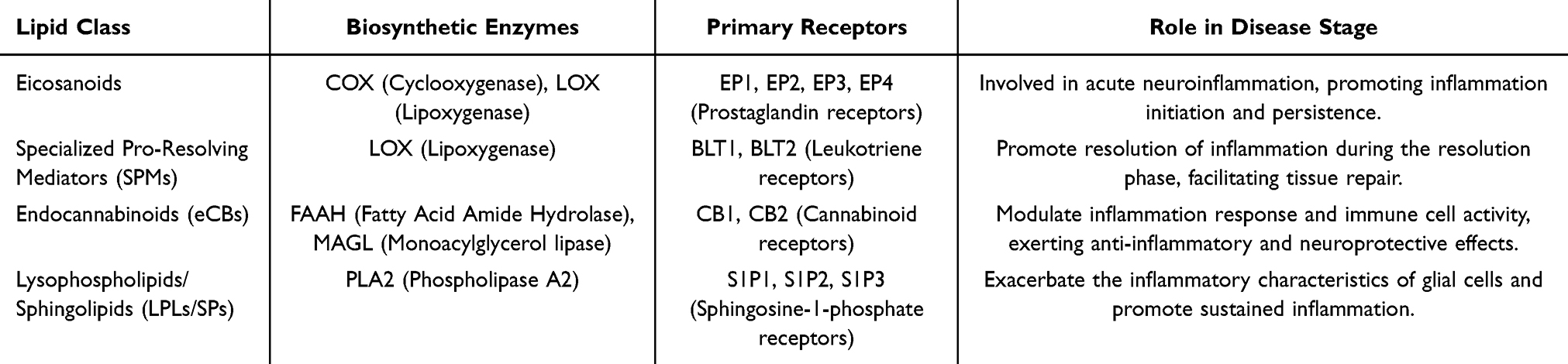 |
Table 1 Summary of Bioactive Lipids and Their Roles in Glial Cell Modulation |
Particularly noteworthy is the increasing amount of evidence that bioactive lipids play a vital role in signal transduction mediated by neuroglia cell in neuroinflammation. These studies not only highlight the potential value of treatments targeting these lipid mechanisms in addressing neuroinflammation caused by their disturbances but also provide a theoretical basis for developing new treatment strategies. Taking SPMs as an example, studies have shown that during the course of self-limited inflammation, extracellular vesicles (EVs) isolated from inflammatory exudates contain detectable levels of SPM precursors such as 14-HDHA and 17-HDHA. These molecules increase during the early phase of inflammation, decline at its peak, and accumulate again during the resolution phase.23 Based on these findings, lipidomic profiling of brain-derived EVs from patients with neurological diseases may help identify specific defects in resolution pathways. This knowledge could pave the way for the development of personalized nanomedicines, in which SPMs—either extracted from EVs of healthy individuals or synthetically produced—are delivered to patients for targeted therapeutic intervention. Moreover, the plasticity of eCBs plays a pivotal role in orchestrating the resolution of inflammation. Timely activation or inhibition of their signaling pathways represents a promising strategy for modulating neuroinflammatory processes. Emerging evidence indicates that during chronic CNS inflammation, astrocytes exhibit not only elevated levels of ceramides but also a marked increase in ceramide-derived glycosphingolipids, which collectively contribute to the progression of neuroinflammation. These findings suggest that ceramides may serve as potential therapeutic targets for mitigating persistent neuroinflammation. Furthermore, the detection of secreted ceramides holds promise as a novel biomarker for assessing neuroinflammatory status and supporting the clinical diagnosis of various neurological disorders. Recent studies have reported a specific elevation of certain ceramide species in the cerebrospinal fluid of patients with multiple sclerosis, and ceramide profiling in plasma has also demonstrated potential for identifying disease risk. The utility of ceramides as predictive biomarkers has thus been increasingly supported by accumulating clinical evidence.69
Using migraine as an example, which is the focus of our research team, neuroinflammation is a key mechanism in migraine attacks, involving complex interactions between neurons, blood vessels, neuroglia cells, and immune cells. Bioactive lipids play a bridging role in this process. For instance, PGE2 promotes pain signal transmission and vasodilation through EP receptors, while endogenous cannabinoids inhibit pain and inflammation through CB1 and CB2 receptors. Abnormal lipid metabolism may lead to a persistent state of neuroinflammation, and these abnormal lipid metabolites may be key pathological factors in chronic migraine (CM) patients. An increasing number of clinical studies have revealed a strong correlation between lipid metabolic profiles and the risk of migraine.72 Interventions targeting n-3 and n-6 fatty acids have been shown to modulate bioactive mediators associated with the pathogenesis of headaches, resulting in a reduction in both the frequency and severity of migraine attacks.73 Moreover, bioactive lipids, such as cannabinoids, have gained growing attention for their potential clinical value in migraine treatment.74 EPA and DHA are also considered promising clinical therapeutic agents for migraine prevention.75 Additionally, based on the lipid metabolic abnormalities and energy homeostasis disruptions observed in migraine patients, assessing lipid changes in plasma and cerebrospinal fluid could emerge as a promising direction for precision diagnosis and treatment of migraine.76 Analyzing the lipid metabolic profile of CM patients can not only elucidate the mechanisms of migraine chronification but also help reveal related comorbidities. Using common comorbidities of CM as examples, such as cognitive impairment and mood disorders, these conditions may be associated with persistent neuroinflammation, neuronal damage, and abnormal neural circuit function. Research reports indicate that bioactive lipids, such as ECS, play a crucial role in regulating cognitive function, suggesting that chronic migraine may interfere with bioactive lipids, thus playing a key role in the management of cognitive function and mood.77,78 In the future, combining advanced techniques such as mass spectrometry and single-cell sequencing will enable more precise mapping of lipid metabolism networks and their changes at different pathological stages, providing a theoretical basis for exploring new lipid metabolic enzymes and receptors as therapeutic targets. This will effectively control migraine onset and its chronicity, as well as advance research on migraine comorbidities. At the same time, with the development of cutting-edge technologies such as imaging mass spectrometry (eg, lipid imaging mass spectrometry to visualize lipid distribution in brain tissues in real-time), biosensor probes (eg, lipid probes based on nanotechnology for monitoring dynamic interactions between lipids and receptors), and CRISPR-based lipid enzyme screening (eg, using CRISPR-Cas9 for high-throughput screening of specific lipid metabolic enzymes), these methods are poised to bring revolutionary advancements to the field of lipid metabolism and drive the next wave of research discoveries. Therefore, future research should not only focus on the mechanisms of lipid metabolism but also actively utilize these innovative technologies to promote precision treatment for migraines and other neurological disorders. Additionally, this review focuses on neuroglia cells, key participants in the migraine process. Overactivation and persistent inflammatory responses of neuroglia cells may be important factors leading to neuropathological changes and chronic pain in CM. Therefore, exploring the mechanisms of specific bioactive lipids in neuroglia cells activation and signal transduction can help reveal whether their abnormal metabolism is a core driver of persistent inflammation in CM. Studying the metabolic pathways and regulatory mechanisms of these lipids in neuroglia cells will provide new insights for developing targeted therapeutic strategies.
Thus, by comprehensively discussing how bioactive lipids participate in the management of neuroinflammation mediated by neuroglia cells, we aim to incorporate these lipids into research and treatment strategies for neuroinflammation. Future studies could focus on how these lipids spontaneously participate in inhibiting neuroinflammation or regulating homeostasis, developing cutting-edge methods for early diagnosis and treatment of neurological diseases. This not only brings new therapeutic options to the field of neuroscience but also provides more precise strategies for disease management.5
Abbreviations
CNS, central nervous system; SPMs, specialized pro-resolving mediators; eCBs, endocannabinoids; LPLs, lysophospholipids; SPs, sphingolipids; LDAM, lipid-droplet-enriched microglia; PUFAs, polyunsaturated fatty acids; AA, arachidonic acid; PLA2, phospholipase A2; LOX, lipoxygenases; COX, cyclooxygenases; LTs, leukotrienes; PGs, prostaglandins; HETEs, Hydroxyeicosatetraenoic acids; LX, lipoxins; EPA, eicosapentaenoic acid; DHA, docosahexaenoic acid; DPA, docosapentaenoic acid; AEA, anandamide; 2AG, 2-arachidonoylglycerol; PEA, palmitoylethanolamide; S1P, sphingosine 1-phosphate; C1P, ceramide 1-phosphate; SphKs, sphingosine kinases; CerK, ceramide kinase; EVs, extracellular vesicles; CM, chronic migraine.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the China National Natural Science Foundation (Nos.82274664, 82004486).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gorji A. Neuroinflammation: the pathogenic mechanism of neurological disorders. Int J Mol Sci. 2022;23(10):5744. doi:10.3390/ijms23105744
2. Kursun O, Yemisci M, van den Maagdenberg AMJM, et al. Migraine and neuroinflammation: the inflammasome perspective. J Headache Pain. 2021;22(1):55. doi:10.1186/s10194-021-01271-1
3. Marogianni C, Sokratous M, Dardiotis E, et al. Neurodegeneration and inflammation-an interesting interplay in Parkinson’s disease. Int J Mol Sci. 2020;21(22):8421. doi:10.3390/ijms21228421
4. Nichols MR, St-Pierre MK, Wendeln AC, et al. Inflammatory mechanisms in neurodegeneration. J Neurochem. 2019;149(5):562–581. doi:10.1111/jnc.14674
5. Chiurchiù V, Leuti A, Maccarrone M. Bioactive lipids and chronic inflammation: managing the fire within. Front Immunol. 2018;9:38. doi:10.3389/fimmu.2018.00038
6. David S, López-Vales R. Bioactive lipid mediators in the initiation and resolution of inflammation after spinal cord injury. Neuroscience. 2021;466:273–297. doi:10.1016/j.neuroscience.2021.04.026
7. Chiurchiù V, Tiberi M, Matteocci A, et al. Lipidomics of bioactive lipids in Alzheimer’s and Parkinson’s diseases: where are we? Int J Mol Sci. 2022;23(11):6235. doi:10.3390/ijms23116235
8. Chauhan G, Roy K, Kumar G, et al. Distinct influence of COX-1 and COX-2 on neuroinflammatory response and associated cognitive deficits during high altitude hypoxia. Neuropharmacology. 2019;146:138–148. doi:10.1016/j.neuropharm.2018.11.026
9. Godeanu S, Cătălin B. The complementary role of morphology in understanding microglial functional heterogeneity. Int J Mol Sci. 2025;26(8):3811. doi:10.3390/ijms26083811
10. Ma H, Li H, Zhang Y, et al. Microglia exhibit distinct heterogeneity rather than M1/M2 polarization within the early stage of acute ischemic stroke. Aging Dis. 2023;14(6):2284–2302. doi:10.14336/AD.2023.0505
11. Li RY, Qin Q, Yang HC, et al. TREM2 in the pathogenesis of AD: a lipid metabolism regulator and potential metabolic therapeutic target. Mol Neurodegener. 2022;17(1):40. doi:10.1186/s13024-022-00542-y
12. Keren-Shaul H, Spinrad A, Weiner A, et al. A unique microglia type associated with restricting development of Alzheimer’s disease. Cell. 2017;169(7):1276–1290.e17. doi:10.1016/j.cell.2017.05.018
13. Marschallinger J, Iram T, Zardeneta M, et al. Lipid-droplet-accumulating microglia represent a dysfunctional and proinflammatory state in the aging brain. Nat Neurosci. 2020;23(2):194–208. doi:10.1038/s41593-019-0566-1
14. Zgorzynska E, Dziedzic B, Markiewicz M, et al. Omega-3 PUFAs suppress IL-1β-induced hyperactivity of immunoproteasomes in astrocytes. Int J Mol Sci. 2021;22(11):5410. doi:10.3390/ijms22115410
15. Woodburn SC, Bollinger JL, Wohleb ES. The semantics of microglia activation: neuroinflammation, homeostasis, and stress. J Neuroinflammation. 2021;18(1):258. doi:10.1186/s12974-021-02309-6
16. Kwon HS, Koh SH. Neuroinflammation in neurodegenerative disorders: the roles of microglia and astrocytes. Transl Neurodegener. 2020;9(1):42. doi:10.1186/s40035-020-00221-2
17. Singh D. Astrocytic and microglial cells as the modulators of neuroinflammation in Alzheimer’s disease. J Neuroinflammation. 2022;19:206. doi:10.1186/s12974-022-02565-0
18. Patani R, Hardingham GE, Liddelow SA. Functional roles of reactive astrocytes in neuroinflammation and neurodegeneration. Nat Rev Neurol. 2023;19(7):395–409. doi:10.1038/s41582-023-00822-1
19. Crisci I, Bonzano S, Nicolas Z, et al. Tamoxifen exerts direct and microglia-mediated effects preventing neuroinflammatory changes in the adult mouse hippocampal neurogenic niche. Glia. 2024;72(7):1273–1289. doi:10.1002/glia.24526
20. Linnerbauer M, Wheeler MA, Quintana FJ. Astrocyte crosstalk in CNS inflammation. Neuron. 2020;108(4):608–622. doi:10.1016/j.neuron.2020.08.012
21. Liddelow SA, Guttenplan KA, Clarke LE, et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541(7638):481–487. doi:10.1038/nature21029
22. Fan YY, Huo J. A1/A2 astrocytes in central nervous system injuries and diseases: angels or devils? Neurochem Int. 2021;148:105080. doi:10.1016/j.neuint.2021.105080
23. Valente M, Dentoni M, Bellizzi F, et al. Specialized pro-resolving mediators in neuroinflammation: overview of studies and perspectives of clinical applications. Molecules. 2022;27(15):4836. doi:10.3390/molecules27154836
24. Talarico G, Trebbastoni A, Bruno G, et al. Modulation of the cannabinoid system: a new perspective for the treatment of the Alzheimer’s disease. Curr Neuropharmacol. 2019;17(2):176–183. doi:10.2174/1570159X16666180702144644
25. Kim C, Livne-Bar I, Gronert K, et al. Fair-weather friends: evidence of lipoxin dysregulation in neurodegeneration. Mol Nutr Food Res. 2020;64(4):e1801076. doi:10.1002/mnfr.201801076
26. Gorica E, Calderone V. Arachidonic acid derivatives and neuroinflammation. CNS Neurol Disord Drug Targets. 2022;21(2):118–129. doi:10.2174/1871527320666210208130412
27. Gao L, Shi H, Sherchan P, et al. Inhibition of lysophosphatidic acid receptor 1 attenuates neuroinflammation via PGE2/EP2/NOX2 signalling and improves the outcome of intracerebral haemorrhage in mice. Brain Behav Immun. 2021;91:615–626. doi:10.1016/j.bbi.2020.09.032
28. Mrowetz H, Kotob MH, Forster J, et al. Leukotriene signaling as molecular correlate for cognitive heterogeneity in aging: an exploratory study. Front Aging Neurosci. 2023;15:1140708. doi:10.3389/fnagi.2023.1140708
29. Klawonn AM, Fritz M, Castany S, et al. Microglial activation elicits a negative affective state through prostaglandin-mediated modulation of striatal neurons. Immunity. 2021;54(2):225–234.e6. doi:10.1016/j.immuni.2020.12.016
30. Saliba SW, Gläser F, Deckers A, et al. Effects of a novel GPR55 antagonist on the arachidonic acid cascade in LPS-activated primary microglial cells. Int J Mol Sci. 2021;22(5):2503. doi:10.3390/ijms22052503
31. Ji H, Zhang Y, Chen C, et al. D-dopachrome tautomerase activates COX2/PGE2 pathway of astrocytes to mediate inflammation following spinal cord injury. J Neuroinflammation. 2021;18(1):130. doi:10.1186/s12974-021-02186-z
32. Sluter MN, Li Q, Yasmen N, et al. The inducible prostaglandin E synthase (mPGES-1) in neuroinflammatory disorders. Exp Biol Med. 2023;248(9):811–819. doi:10.1177/15353702231179926
33. Xia Y, Xiao Y, Wang ZH, et al. Bacteroides Fragilis in the gut microbiomes of Alzheimer’s disease activates microglia and triggers pathogenesis in neuronal C/EBPβ transgenic mice. Nat Commun. 2023;14(1):5471. doi:10.1038/s41467-023-41283-w
34. Ghazanfari N, van Waarde A, Dierckx RAJO, et al. Is cyclooxygenase-1 involved in neuroinflammation? J Neurosci Res. 2021;99(11):2976–2998. doi:10.1002/jnr.24934
35. Serhan CN, Chiang N, Dalli J. New pro-resolving n-3 mediators bridge resolution of infectious inflammation to tissue regeneration. Mol Aspect Med. 2018;64:1–17. doi:10.1016/j.mam.2017.08.002
36. Leuti A, Maccarrone M, Chiurchiù V. Proresolving lipid mediators: endogenous modulators of oxidative stress. Oxid Med Cell Longev. 2019;2019:8107265. doi:10.1155/2019/8107265
37. Vik A, Hansen TV. Stereoselective syntheses and biological activities of E-series resolvins. Org Biomol Chem. 2021;19(4):705–721. doi:10.1039/D0OB02218G
38. Flak MB, Koenis DS, Sobrino A, et al. GPR101 mediates the pro-resolving actions of RvD5n-3 DPA in arthritis and infections. J Clin Invest. 2020;130(1):359–373. doi:10.1172/JCI131609
39. Serhan CN, Gupta SK, Perretti M, et al. The Atlas of Inflammation Resolution (AIR). Mol Aspect Med. 2020;74:100894. doi:10.1016/j.mam.2020.100894
40. Teixeira-Santos L, Martins S, Sousa T, et al. The pro-resolving lipid mediator Maresin 1 ameliorates pain responses and neuroinflammation in the spared nerve injury-induced neuropathic pain: a study in male and female mice. PLoS One. 2023;18(6):e0287392. doi:10.1371/journal.pone.0287392
41. Bisicchia E, Sasso V, Catanzaro G, et al. Resolvin D1 halts remote neuroinflammation and improves functional recovery after focal brain damage via ALX/FPR2 receptor-regulated MicroRNAs. Mol Neurobiol. 2018;55(8):6894–6905. doi:10.1007/s12035-018-0889-z
42. Ren YZ, Zhang BZ, Zhao XJ, et al. Resolvin D1 ameliorates cognitive impairment following traumatic brain injury via protecting astrocytic mitochondria. J Neurochem. 2020;154(5):530–546. doi:10.1111/jnc.14962
43. Kantarci A, Aytan N, Palaska I, et al. Combined administration of resolvin E1 and lipoxin A4 resolves inflammation in a murine model of Alzheimer’s disease. Exp Neurol. 2018;300:111–120. doi:10.1016/j.expneurol.2017.11.005
44. Dong Y, Xu M, Kalueff AV, et al. Dietary eicosapentaenoic acid normalizes hippocampal omega-3 and 6 polyunsaturated fatty acid profile, attenuates glial activation and regulates BDNF function in a rodent model of neuroinflammation induced by central interleukin-1β administration. Eur J Nutr. 2018;57(5):1781–1791. doi:10.1007/s00394-017-1462-7
45. Xia J, Yang L, Huang C, et al. Omega-3 polyunsaturated fatty acid eicosapentaenoic acid or docosahexaenoic acid improved ageing-associated cognitive decline by regulating glial polarization. Mar Drugs. 2023;21(7):398. doi:10.3390/md21070398
46. Wang X, Xiao A, Yang Y, et al. DHA and EPA prevent seizure and depression-like behavior by inhibiting ferroptosis and neuroinflammation via different mode-of-actions in a pentylenetetrazole-induced kindling model in mice. Mol Nutr Food Res. 2022;66(22):e2200275. doi:10.1002/mnfr.202200275
47. Lee JY, Han SH, Park MH, et al. Neuronal SphK1 acetylates COX2 and contributes to pathogenesis in a model of Alzheimer’s Disease. Nat Commun. 2018;9(1):1479. doi:10.1038/s41467-018-03674-2
48. da Silva Fiorin F, Do Espírito Santo CC, Do Nascimento RS, et al. Capsaicin-sensitive fibers mediate periorbital allodynia and activation of inflammatory cells after traumatic brain injury in rats: involvement of TRPV1 channels in post-traumatic headache. Neuropharmacology. 2020;176:108215. doi:10.1016/j.neuropharm.2020.108215
49. Wolf J, Urits I, Orhurhu V, et al. The role of the cannabinoid system in pain control: basic and clinical implications. Curr Pain Headache Rep. 2020;24(7):35. doi:10.1007/s11916-020-00873-9
50. Baggelaar MP, Maccarrone M, van der Stelt M. 2-Arachidonoylglycerol: a signaling lipid with manifold actions in the brain. Prog Lipid Res. 2018;71:1–17. doi:10.1016/j.plipres.2018.05.002
51. Levine A, Liktor-Busa E, Lipinski AA, et al. Sex differences in the expression of the endocannabinoid system within V1M cortex and PAG of Sprague Dawley rats. Biol Sex Differ. 2021;12(1):60. doi:10.1186/s13293-021-00402-2
52. Covelo A, Eraso-Pichot A, Fernández-Moncada I, et al. CB1R-dependent regulation of astrocyte physiology and astrocyte-neuron interactions. Neuropharmacology. 2021;195:108678. doi:10.1016/j.neuropharm.2021.108678
53. Mecha M, Feliú A, Machín I, et al. 2-AG limits Theiler’s virus induced acute neuroinflammation by modulating microglia and promoting MDSCs. Glia. 2018;66(7):1447–1463. doi:10.1002/glia.23317
54. Duffy SS, Hayes JP, Fiore NT, et al. The cannabinoid system and microglia in health and disease. Neuropharmacology. 2021;190:108555. doi:10.1016/j.neuropharm.2021.108555
55. Kamaruzzaman MA, Romli MH, Abas R, et al. Regulatory role of the endocannabinoid system on glial cells toward cognitive function in Alzheimer’s disease: a systematic review and meta-analysis of animal studies. Front Pharmacol. 2023;14:1053680. doi:10.3389/fphar.2023.1053680
56. Bernal-Chico A, Tepavcevic V, Manterola A, et al. Endocannabinoid signaling in brain diseases: emerging relevance of glial cells. Glia. 2023;71(1):103–126. doi:10.1002/glia.24172
57. Chen C. Inhibiting degradation of 2-arachidonoylglycerol as a therapeutic strategy for neurodegenerative diseases. Pharmacol Ther. 2023;244:108394. doi:10.1016/j.pharmthera.2023.108394
58. De Filippis D, D’Amico A, Cipriano M, et al; Endocannabinoid Research Group. Levels of endocannabinoids and palmitoylethanolamide and their pharmacological manipulation in chronic granulomatous inflammation in rats. Pharmacol Res. 2010;61(4):321–328. doi:10.1016/j.phrs.2009.11.005
59. Yaginuma S, Omi J, Kano K, et al. Lysophospholipids and their producing enzymes: their pathological roles and potential as pathological biomarkers. Pharmacol Ther. 2023;246:108415. doi:10.1016/j.pharmthera.2023.108415
60. Langeslag M, Kress M. The ceramide-S1P pathway as a druggable target to alleviate peripheral neuropathic pain. Expert Opin Ther Targets. 2020;24(9):869–884. doi:10.1080/14728222.2020.1787989
61. Nishino S, Yamashita H, Tamori M, et al. Translocation and activation of sphingosine kinase 1 by ceramide-1-phosphate. J Cell Biochem. 2019;120(4):5396–5408. doi:10.1002/jcb.27818
62. Chen Z, Doyle TM, Luongo L, et al. Sphingosine-1-phosphate receptor 1 activation in astrocytes contributes to neuropathic pain. Proc Natl Acad Sci USA. 2019;116(21):10557–10562. doi:10.1073/pnas.1820466116
63. Doyle TM, Chen Z, Durante M, et al. Activation of Sphingosine-1-Phosphate receptor 1 in the spinal cord produces mechanohypersensitivity through the activation of inflammasome and IL-1β pathway. J Pain. 2019;20(8):956–964. doi:10.1016/j.jpain.2019.02.007
64. Kano K, Aoki J, Hla T. Lysophospholipid mediators in health and disease. Annu Rev Pathol. 2022;17:459–483. doi:10.1146/annurev-pathol-050420-025929
65. Ayub M, Jin HK, Bae JS. Novelty of sphingolipids in the central nervous system physiology and disease: focusing on the sphingolipid hypothesis of neuroinflammation and neurodegeneration. Int J Mol Sci. 2021;22(14):7353. doi:10.3390/ijms22147353
66. Presa N, Gomez-Larrauri A, Dominguez-Herrera A, et al. Novel signaling aspects of ceramide 1-phosphate. Biochim Biophys Acta Mol Cell Biol Lipids. 2020;1865(4):158630. doi:10.1016/j.bbalip.2020.158630
67. Simanshu DK, Kamlekar RK, Wijesinghe DS, et al. Non-vesicular trafficking by a ceramide-1-phosphate transfer protein regulates eicosanoids. Nature. 2013;500(7463):463–467. doi:10.1038/nature12332
68. Lee JY, Jin HK, Bae JS. Sphingolipids in neuroinflammation: a potential target for diagnosis and therapy. BMB Rep. 2020;53(1):28–34. doi:10.5483/BMBRep.2020.53.1.278
69. de Wit NM, den Hoedt S, Martinez-Martinez P, et al. Astrocytic ceramide as possible indicator of neuroinflammation. J Neuroinflammation. 2019;16(1):48. doi:10.1186/s12974-019-1436-1
70. Terlizzi M, Colarusso C, Somma P, et al. S1P-induced TNF-α and IL-6 release from PBMCs exacerbates lung cancer-associated inflammation. Cells. 2022;11(16):2524. doi:10.3390/cells11162524
71. Yang Y, Liu Y, Tang H, et al. FTY720 suppresses pathogenic retinal müller cell activation and chronic progression by inhibiting the mTOR/NF-κB signaling pathway and regulating autophagy. Curr Eye Res. 2024;49(8):862–871. doi:10.1080/02713683.2024.2337301
72. Hong P, Han L, Wan Y. Mendelian randomization study of lipid metabolism characteristics and migraine risk. Eur J Pain. 2024;28(6):978–986. doi:10.1002/ejp.2235
73. Ramsden CE, Zamora D, Faurot KR, et al. Dietary alteration of n-3 and n-6 fatty acids for headache reduction in adults with migraine: randomized controlled trial. BMJ. 2021;374:n1448. doi:10.1136/bmj.n1448
74. Zorrilla E, Della Pietra A, Russo AF. Interplay between cannabinoids and the neuroimmune system in migraine. J Headache Pain. 2024;25(1):178. doi:10.1186/s10194-024-01883-3
75. Tseng PT, Zeng BY, Chen JJ, et al. High dosage omega-3 fatty acids outperform existing pharmacological options for migraine prophylaxis: a network meta-analysis. Adv Nutr. 2024;15(2):100163. doi:10.1016/j.advnut.2023.100163
76. Castor K, Dawlaty J, Arakaki X, et al. Plasma lipolysis and changes in plasma and cerebrospinal fluid signaling lipids reveal abnormal lipid metabolism in chronic migraine. Front Mol Neurosci. 2021;14:691733. doi:10.3389/fnmol.2021.691733
77. de Melo Reis RA, Isaac AR, Freitas HR, et al. Quality of life and a surveillant endocannabinoid system. Front Neurosci. 2021;15:747229. doi:10.3389/fnins.2021.747229
78. Rezende B, Alencar AKN, de Bem GF, et al. Endocannabinoid system: chemical characteristics and biological activity. Pharmaceuticals. 2023;16(2):148. doi:10.3390/ph16020148
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.