Back to Journals » International Journal of Nanomedicine » Volume 15
Consequences Of Long-Term Bacteria’s Exposure To Silver Nanoformulations With Different PhysicoChemical Properties
Authors Kędziora A, Wernecki M ![]() , Korzekwa K, Speruda M, Gerasymchuk Y
, Korzekwa K, Speruda M, Gerasymchuk Y ![]() , Łukowiak A
, Łukowiak A ![]() , Bugla-Płoskońska G
, Bugla-Płoskońska G
Received 15 March 2019
Accepted for publication 18 September 2019
Published 14 January 2020 Volume 2020:15 Pages 199—213
DOI https://doi.org/10.2147/IJN.S208838
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Anderson Oliveira Lobo
Anna Kędziora, 1 Maciej Wernecki, 1 Kamila Korzekwa, 1 Mateusz Speruda, 1 Yuriy Gerasymchuk, 2 Anna Łukowiak, 2 Gabriela Bugla-Płoskońska 1
1Department of Microbiology, Institute of Genetics and Microbiology, University of Wroclaw, Wroclaw, Poland; 2Trzebiatowski Institute of Low Temperature and Structure Research PAS in Wrocław, Wroclaw, Poland
Correspondence: Gabriela Bugla-Płoskońska; Anna Kędziora
Department of Microbiology, Institute of Genetics and Microbiology, University of Wroclaw, Ul. Przybyszewskiego 63-77, Wroclaw, Poland
Tel +48 71 375 63 23
Email [email protected]; [email protected]
Purpose: Resistance to antibiotics is a major problem of public health. One of the alternative therapies is silver – more and more popular because of nanotechnology development and new possibilities of usage. As a component of colloid, powder, cream, bandages, etc., nanosilver is often recommended to treat the multidrug-resistant pathogens and we can observe its overuse also outside of the clinic where different physicochemical forms of silver nanoformulations (e.g. size, shape, compounds, surface area) are introduced. In this research, we described the consequences of long-term bacteria exposure to silver nanoformulations with different physicochemical properties, including changes in genome and changes of bacterial sensitivity to silver nanoformulations and/or antibiotics. Moreover, the prevalence of exogenous resistance to silver among multidrug-resistant bacteria was determined.
Materials and Methods: Gram-negative and Gram-positive bacteria strains are described as sensitive and multidrug-resistant strains. The sensitivity of the tested bacterial strains to antibiotics was carried out with disc diffusion methods. The sensitivity of bacteria to silver nanoformulations and development of bacterial resistance to silver nanoformulations has been verified via determination of the minimal inhibitory concentrations. The presence of sil genes was verified via PCR reaction and DNA electrophoresis. The genomic and phenotypic changes have been verified via genome sequencing and bioinformatics analysis.
Results: Bacteria after long-term exposure to silver nanoformulations may change their sensitivity to silver forms and/or antibiotics, depending on the physicochemical properties of silver nanoformulations, resulting from phenotypic or genetic changes in the bacterial cell. Finally, adaptants and mutants may become more sensitive or resistant to some antibiotics than wild types.
Conclusion: Application of silver nanoformulations in the case of multiple resistance or multidrug-resistant bacterial infection can enhance or decrease their resistance to antibiotics. The usage of nanosilver in a clinic and outside of the clinic should be determined and should be under strong control. Moreover, each silver nanomaterial should be considered as a separate agent with a potential different mode of antibacterial action.
Keywords: silver, antibiotics, resistance, sil genes, mutant, adaptant
Introduction
The overuse of antibiotics has led to the increase of bacterial resistance.1 Currently, one of the most popular alternatives and often a supporting way, recommended as an antibacterial factor in medicine and in most industrial branches, is silver. Silver, among other metals, possesses biological activity and its high efficacy against a broad range of microorganisms (including fungi, Gram-negative and Gram-positive bacteria, or even viruses) is well described in the literature.2–6 Development of nanotechnology gives us new possibilities of silver usage as a new generation of silver-based products, e.g. nanoparticles (AgNPs). It is a kind of structure (with at least one dimension lower than 100 nm) which obtains unique physicochemical and biological properties due to its nanoscale.7,8 Nanotechnological modifications of silver are usually connected with the transfer of silver ions (Ag+) to metallic silver (Ag0). Despite the production of silver-based medical products (orthopedic implants, bandages, biomedical tools, or devices), the potential of antibacterial nanosilver is more exploited in industrial branches. Due to the popularity of silver nanoparticles, the commercial market is full of fabrics, clothes, cosmetics, or simple household items with AgNPs.9–11 The similarities and differences between antibacterial activity of silver ions and silver nanoformulations were reviewed by the authors.9
Molecular mechanisms of bacterial resistance to silver ions are well known, especially in Gram-negative bacteria.12,13 The first described bacterial silver resistance occurred in 1975, in Massachusetts General Hospital. Gram-negative Salmonella Typhimurium, which caused the death of several burn ward patients, was the host of the pMG101 plasmid, a 180 kb construct with specific regions involved in bacterial resistance to silver ions.12,14 This type of resistance was called “exogenous silver resistance” by Randall et al.13 The sil region covers an operon of nine sil genes, gathered in three transcriptional units (silRS, silE, and silCFBAGP), each controlled by a separate promoter.13,15 Functions of proteins encoded by sil genes were determined after comparison of their structures to homolog components of the CusCFBA and PcoE system with a chromosome-encoded efflux pump involved in the transport of copper and silver ions.13,16 Despite that, the function of the SilG protein remains unknown. The transcription of sil genes is carried out by the SilS/SilR two-component regulatory system, made of membrane histidine kinase and a response regulator.12,17 Products of silA (inner membrane substrate-binding transporter), silB (membrane fusion protein), and silC (outer membrane protein) genes form a tripartite resistance–nodulation–division (RND) efflux pump, SilCFBA – a structure spanning both membranes, responsible for ejecting silver ions out of the cell (from cytoplasm or periplasmic space).12,14 The protein encoded by the SilP gene is a P-type ATPase located in the inner membrane. It transports all of the toxic Ag+ from cytoplasm to periplasmic space, where it can be captured and delivered to SilCFBA by periplasmic chaperone – SilE.15 A similar role is performed by another protein, encoded by the silF gene. It stops the uptake of silver ions from the environment, by binding Ag+ in periplasmic space and passing them to the efflux pump.14,16 The distribution of sil genes in pMG101 plasmid is presented in Figure 1. The presence of pMG101 in bacteria is not a requirement for changing their phenotypes to silver-resistant. Randall et al13 proved that silver ions exert selective pressure on bacterial cells treated during 6 days with silver ions (Ag+), causing point mutations in the bacterial (Escherichia coli) genome, e.g. in ompR or cusS genes, leading to the loss of their function. OmpR is a transcription factor of OmpC and OmpF – two outer membrane proteins, porins with a β-barrel structure, responsible for the transport of drugs and cations such as silver ions. CusS is a histidine kinase, a component of the regulatory system of chromosome-encoded efflux pump CusCFBA.12,17 As described by Randall et al,13 the examined bacteria became 6 times more resistant to silver ions. Randall et al13 proved that there were no changes in Staphylococcus aureus sensitivity to silver ions after 42 days of exposition to Ag+. Despite that, the occurrence of a single sil gene (silE) in staphylococci is possible but with an unknown function.18 Besides silver, resistance to mercury, tellurite and a few antibiotics such as streptomycin, chloramphenicol, tetracycline, and ampicillin are encoded on the pMG101 plasmid.12
 |
Figure 1 Map view. Distribution of sil genes in the plasmid pMG101 (created with SnapGene). |
Resistance to antibiotics is a major and growing public health problem. It is estimated that every year about 25,000 patients in Europe die due to infections caused by bacterial strains resistant to antibiotics.19 The increasing percentage of isolates of bacteria, both Gram-negative and Gram-positive, resistant to many antibiotics is indicated at the same time.20
The term “multiple resistance” (MR) refers to microorganisms resistant to more than one antibiotic used in the treatment of infection.21 Bacteria described as “multidrug resistant” (MDR) are characterized by bacterial resistance to antibiotics that belong to 3 or more different classes.21 MDR strains are a group of microorganisms that are especially dangerous due to the fact that therapeutic options are limited. Data on the phenomenon of antibiotic resistance pathogens in European countries were included in the European Centre for Disease Prevention and Control (ECDC) report (November 2017) summarizing data for 2013–2016 from 30 European countries.20 According to the report, the percentage of antibiotic-resistant strains is higher in the countries of southern and south-eastern Europe in comparison to the countries of northern Europe. Over 1/3 of the isolates of Klebsiella pneumoniae in Europe are resistant to one therapeutic group, but resistance to 3 or more therapeutic groups was noticed more often. Among the E. coli in European countries, a statistically significant increase in resistance was observed in the case of using third-generation cephalosporins, fluoroquinolones, and aminoglycosides. In 2013–2016, the amount of carbapenem-resistant strains of E. coli and K. pneumoniae increased in several countries. In the years 2013–2016, the European trend of methicillin-resistant S. aureus gradually decreased, although in 10 countries the percentage of such strains still remains above 25%. In Poland, since 2013, it has remained at the level of 10–25%. As a result, standard treatments become ineffective, and infections persist and may spread.19 Bacteria producing carbapenemases (usually belonging to K. pneumoniae, Pseudomonas aeruginosa, and Acinetobacterspp.) become a significant problem. The treatment of this infection is limited to one or two antibiotics.22,23 The rapid spread of carbapenemase resistance (CPE) in Enterobacteriaceae is one of the most serious problems for the health and safety of patients. Prudent use of silver nanoformulations and widespread observance of the principles of control and prevention of infections in all health care sectors, e.g. hospitals and nursing homes, form the basis for effective interventions to prevent the selection and spread of antibiotic-resistant bacteria.24,25 Otherwise, Europe may soon be faced with epidemic hospital diseases caused by Enterobacteriaceae with extended multiple resistance (extensively drug resistant, XDR), or even strains completely resistant (pandrug resistant, PDR).26
The main purpose of this study was to check the sensitivity of bacteria to silver nanoformulations after long-term exposure, to analyze the exogenous and endogenous resistance to silver, and, finally, to compare the antibiotic susceptibility of Gram-positive and Gram-negative bacteria strains (wild types and variants with changed sensitivity after repeated exposure to silver nanoformulations).
Materials And Methods
Bacteria Strains
The wild types (19 strains altogether) and their selected variants (signed in the text as S1 V, S2 V, S7 V strains altogether) of Gram-negative strains (E. coli, K. pneumoniae, Enterobacter cloacae) and Gram-positive bacteria strains (S. aureus) belonging to MR or MDR bacteria were tested. Bacterial strains (wild type) were purchased from ATCC or provided by Dialab Medical Laboratory. The variants were selected after repeated exposure to silver nanoformulations (S0–S7), described below.
Chemicals
The media (Luria Broth [LB], Mueller Hinton Agar [MHA], Mueller Hinton Broth [MHB]) were purchased from Biocorp. Antibiotics were obtained from Oxoid. The Plasmid Mini Kit, Genomic Mini Kit, and DNA Ladder marker (0.1 µg/mL. 100–1000 bp) were purchased from A&A Biotechnology. Midori Green Advance DNA Stain (Nippon Genetics), Phusion Hot Start II High Fidelity PCR Master Mix, and 50× Tris–acetate EDTA buffer (TAE, pH 8.3) were purchased from Thermo Scientific. All of the primers were ordered in Genomed. The sequences of all used primers are summarized in Table 1 (based on the information described by Woods et al27). Silver nanoformulations S1–S6, described in detail in our previous study,5,28 were prepared by A. Kedziora. The formulations S0 and S7 were commercially available. All silver nanoformulations are summarized in Table 2.
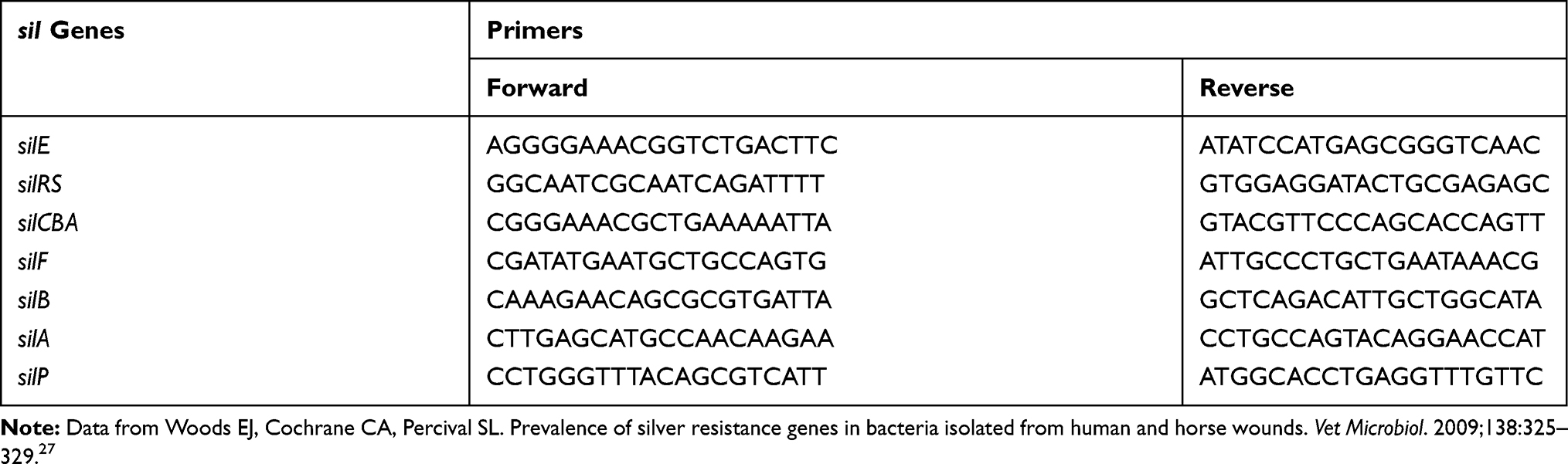 |
Table 1 Sequences Of Forward And Reverse Primers |
 |
Table 2 Silver Nanoformulations, Which The Bacteria Were Treated With |
Long-Term Exposure Of Bacteria To Silver Nanoformulations
This task was performed with long-term exposure in gradual increases of minimal inhibitory concentration (MIC) of silver nanoformulations S0–S7 (0.5 MIC, 1 MIC, and 4 MIC). Initially, the sensitivity of bacteria to silver nanoformulations was determined via MIC measurement.29 Bacteria strains were stored at −70 °C before each investigation and the standard protocol was performed to check their purity and revitalization. The inoculum was spread on the agar plate (MHA), incubated at 37 °C for 19 h, and after that used in the experiment. In the first step, the stock of silver samples was prepared (at concentrations between 2048 µg/mL and 0.03125 µg/mL, depending on the nanoformulation sample). 100 µL of each concentration was poured to hole. Next, the bacteria strains at the final concentration of 1.5x106 cfu/mL were added and incubated at 37 °C. Minimal inhibitory concentration values were read within 16–19 h. The experiment was repeated 3 times. For 0.5 MIC and 1 MIC, broth (MHB) was used; hence, in the case of 4 MIC, an agar plate (MHA) was applied. The appropriate bacteria density (cfu/mL comparable with MIC determination) was poured into the 0.5 MIC and incubated for 24 h at 37 °C in a final volume of 10 mL. After that time, 0.5 mL of overnight culture was transferred to the new broth (containing 0.5 MIC concentration of the appropriate silver nanoformulations) and incubated under the same condition. Overnight culture of bacteria was transferred to the new broth (with 1 MIC), incubated at 37 °C forr 24 h, and the steps were repeated for 1 MIC as described for 0.5 MIC. The population that adapted to the increasing concentration of silver nanoformulations grew on the agar plate (4 MIC). The MIC value was then determined again as described above and compared with that before long-term exposure.30
Analysis Of Whole Genome Sequences
Whole genome DNA was isolated with the Genomic Mini Kit according to the original instruction from 2 mL of overnight LB cultures. The product purity and concentration were verified with a Nano Photometer (Implen). Genomic libraries were prepared according to the NEBNext DNA Library Prep Master Mix Set for Illumina protocol, and sequencing (NGS) was performed at Genomed (Warsaw, Poland) using the Illumina MiSeq platform. NGS reads were trimmed with Cutadapt 1.9.1.31 De-novo genome assembly was performed with SPAdes 3.13.0,32 their quality was evaluated with QUASTt 5.0.2,33 and resulting contigs were rearranged with progressiveMauve in Mauve 2.4.034,35 using the genome of E. coli K-12 substr. MG1655 (NC_000913.3) or K. pneumoniae subsp. pneumoniae HS11286 (NC_016845.1) as a reference. Newly assembled genomes were annotated with prokka 1.12.36 Genomic mutations in adapted strains were identified and characterized with snippy 4.3.8.37 The pre-processed reads and assembled genomes have been deposited in the NCBI SRA and Genome databases accordingly under the following accession numbers: SRR9733699, SRR9733700, SRR9733697, SRR9733698, SRR9733703, SRR9733704, SRR9733701, SRR9733702, SRR9733705, SRR9733706, SRR9733707 and VLTC00000000, VLTB00000000, VLTA00000000, VLSZ00000000, VLSY00000000, VLSX00000000, VLSW00000000, VLSV00000000, VLSU00000000, VLST00000000, VLSS00000000.
Detection Of Sil Genes
To verify the exogenous silver resistance in all tested bacteria strains, the prevalence of sil genes located on pMG101 plasmid was determined in the following steps.
Plasmid Isolation
Plasmid pMG101 was isolated with the Plasmid Mini Kit according to the original instruction. The product quantity of this extraction was confirmed with the Nano Photometer (Implen). Plasmid DNA was isolated from all of the tested strains (described above), and the one from the E. coli J53 strain was used as a positive control. All bacteria strains grew on Luria Broth before DNA extraction.
Primer Preparation
In the first step, lyophilized primers were centrifuged (12,500 rpm, 2 min); next, the required volume of water (MQ) was poured to the final concentration of 100 µM. The primers were vortexed and left for 10 min at room temperature. Then, they were diluted 10 times and stored at −20 °C.
PCR Mixture
The following reagents were mixed for every reaction: 5 µL Phusion Hot Start II High Fidelity PCR Master Mix, 0.5 µL forward primer, 0.5 µL reverse primer, 1 µL plasmid, and the required volume of MQ. The final volume of all reactions was 10 µL. The positive (with plasmid from E. coli J53) and negative (sample without any of plasmid) controls were performed.
PCR Conditions
The following steps were established for the best final products of PCR: 30 sec at 98 °C (initial denaturation) followed by 30 cycles of 98 °C for 5 sec (denaturation), 60 °C for 10 sec (annealing), and 72 °C for 12 sec (elongation); and 72 °C for 10 min (final elongation). PCR was carried out on a T100™ Thermal Cycler (BioRad).
DNA Electrophoresis
5 μL of each sample and 1 μL of heavy buffer (30% glycerol stained bromophenol blue) were loaded to a 2% agarose Tris–acetate–EDTA (TAE) gel containing 0.002% Midori Green. Electrophoresis was carried out during 0.5 h at 7.5 V/cm. A 5 μL DNA marker was applied on the gel. Images of the gels were carried out on a Gel Doc XR+ (BioRad).
Antibiotic Susceptibility Testing
Sensitivity of the tested bacterial strains to antibiotics was determined with disc diffusion methods. The criteria for selection of antimicrobials were based on the EUCAST recommendation.38 Bacteria (0.5 McFarland) were inoculated and antibiotic discs were placed on the MHA plate. Plates were incubated at 37 °C during 18 h, followed by zone diameter measurement and breakpoint estimation.
Results And Discussion
The changes of bacterial sensitivity to silver nanoformulations were determined after bacteria strains' exposition to increasing concentrations of silver nanoformulations. After comparison of the primary (in wild type strains, wt) and secondary (among variants, V) MIC values, the changes of bacterial sensitivity were determined and the selection of variants was possible. The obtained MIC values are described in Tables 3 and 4. The susceptibility of bacteria depended on the kind of silver nanoformulations (their size, shape, surface area, compounds, etc.) and bacterial strains. Among some of the tested bacterial strains, the changes of sensitivity to certain silver nanoformulations were noticed. It is interesting that almost all strains changed their susceptibility level to S7, 4 of 5 E. coli changed sensitivity to S2, 2 of 3 K. pneumoniae changed sensitivity to S1 and S2, and only 1 of 6 S. aureus changed the sensitivity to S0. All of the selected variants are summarized in Table 5. To verify the reasons for changing the variants' sensitivity, whole genome sequences were analyzed. DNA from 11 of the tested wt strains and their phenotype variants (V) was isolated: K. pneumoniae ATCC 4352 wt, K. pneumoniae ATCC 4352 S1 V, K. pneumoniae ATCC 4352 S2 V, K. pneumoniae ATCC 4352 S7 V, K. pneumoniae 626 wt, K. pneumoniae 626 S1 V, K. pneumoniae 626 S2 V, K. pneumoniae 626 S7 V, E. coli ATCC 11229 wt, E. coli ATCC 11229 S2 V, and E. coli ATCC 11229 S7 V. The above strains were selected on the basis of the obtained results described in Table 5. For the analysis, mutations were identified in protein coding sequences (CDS), RNA coding sequences, and intergenic regions. All mutations recorded in the above genomic regions were divided into nucleotide substitutions, insertions, deletions, and other complex mutations (Table 6). The most mutations (mainly SNP) were noticed among K. pneumoniae ATCC 4352 and E. coli ATCC 11229 strains treated with S1 and S2, respectively. Definitely fewer genomic changes were indicated among K. pneumoniae 626 treated with S1 and S2, and K. pneumoniae ATCC 4352 treated with S2. In general, all mutations can be divided into conservative and nonconservative. Mutation is considered as conservative when it does not introduce any change in primary protein structure or the introduced amino acid change has score ≥0 in BLOSUM62.39 Otherwise, mutation is considered as nonconservative.40 Counts of genes harboring at least one mutation (including conservative and nonconservative) are listed in Table 6. In the case of the strains with the highest number of mutations (K. pneumoniae ATCC 4352 S1 V and E. coli ATCC 11229 S2 V), more than 1200 genes were identified with nonconservative mutations (only a few examples are shown in Table 6), while among the strains K. pneumoniae 626 S1 and S2 V and K. pneumoniae ATCC 4352 S2 V only one gene was identified, cusS and tamB appropriate. It is worth underlining that the same genes were noticed in cases of strains with a huge number of mutations. The obtained results can be divided into three clusters, where each one has a distinctive feature: a) high number of mutations (E. coli ATCC 11229 S2 V, K. pneumoniae ATCC 4352 S1 V), b) single gene mutations (K. pneumoniae 626 S1 and S2 V, and K. pneumoniae ATCC 4352 S2 V), c) no gene mutations (E. coli ATCC 11229 S7 V and, K. pneumoniae ATCC 4352 S7 V and 626 S7 V) (Table 6). The mutation frequency counts in genomic sequencing of E. coli ATCC 11229 S2 V and K. pneumoniae ATCC 4352 S1 V strains are shown in Figure 2. As indicated by these data, all recorded mutations occurred with a frequency between 82 and 100 per 100 reads, suggesting some level of heterogeneity gained in silver-treated populations, more noticeable in the case of K. pneumoniae ATCC 4352 S1 V. Therefore, this post-treatment population may be considered not as a uniform strain, but as a mix of differently adapted organisms, harboring varying sets of genomic mutations. In summary, S1 and S2 samples may be considered as silver nanoformulations that exert the highest selective pressure depending on the individual features of bacterial strains, in contrast to S7. To explain the mutation rates among obtained variants after S0–S7 exposition, the substitution counts from whole genome sequencing followed by the distribution of substitutions in coding regions and possible impact on coded protein structure are described in Table 7. Since the two variants (E. coli ATCC 11229 S2 V and K. pneumoniae ATCC 4352 S1 V) gained an extraordinarily high number of mutations, they may be premised as mutator strains.41 This observation can be explained by a possible decrease of DNA repair efficiency in obtained variants, resulting from nonconservative mutations in genes coding proteins involved in mismatchrepair (MMR): mutL (Pro389Ser, Leu418Pro) in E. coli ATCC 11229 S2 V, and mutL (Pro371Thr) and mutM (Ala148Glu) in K. pneumoniae ATCC 4352 S1 V. As MMR deficiency promotes not only quantitative, but also qualitative changes in the mutational profile, substitution rates from this experiment are compared with corresponding data obtained by Lee et al.40 In brief, the wt strain gains transitions and transversions with comparable ratios (56% vs 44%), with a slight domination of G:C>A:T transitions. In contrast, the MMR defective strain (mutL) gains mostly transitions (over 90%) with clear dominance of A:T>G:C transitions (70%). The two mutator-like variants from this experiment, carrying spontaneous mutation in mutL, do not fully fall into any of these categories, although the transition ratio (73–75%) is notably higher than the transversion ratio (25–27%), what is a resemblance to the mutL strain, where A:T>G:C and G:C>A:T transition ratios are close to equal. Considering the effect of substitution, in both WT and mutL strains, the nonsynonymous mutation ratio outweighed the synonymous mutation ratio. Interestingly, in silver-resistant variants the trend is opposite and most substitutions do not affect coded proteins. Therefore, the mutator phenotype in E. coli ATCC 11229 S2 and K. pneumoniae ATCC 4352 cannot be explained with a single mutL dysfunction, and may be the result of more complex changes and specific selective pressure of silver nanoformulations. Other examined variants do not harbor mutations in any known gene related to DNA repair, consistently the mutator phenotype is missing in them. Table 7 illustrates the relative abundance of particular substitution types within the sample. Genotoxicity of silver nanoparticles toward Gram-positive and Gram-negative bacteria was confirmed.42 Panacek et al43 showed that Gram-negative bacteria (including E. coli strains) can develop resistance to silver nanoparticles after repeated exposure but without any genetic changes. It resulted from phenotypic changes involving inhibition of the flagellum protein production. Panacek et al43 confirmed that strains which changed sensitivity to silver nanoparticles did not change sensitivity to silver ions. Those results suggest different response of bacteria cells to various types of silver but is the opposite of studies performed by Randall et al13 that proved the genomic changes in E. coli strains after 6 days of exposure to a subinhibitory concentration of silver ions.
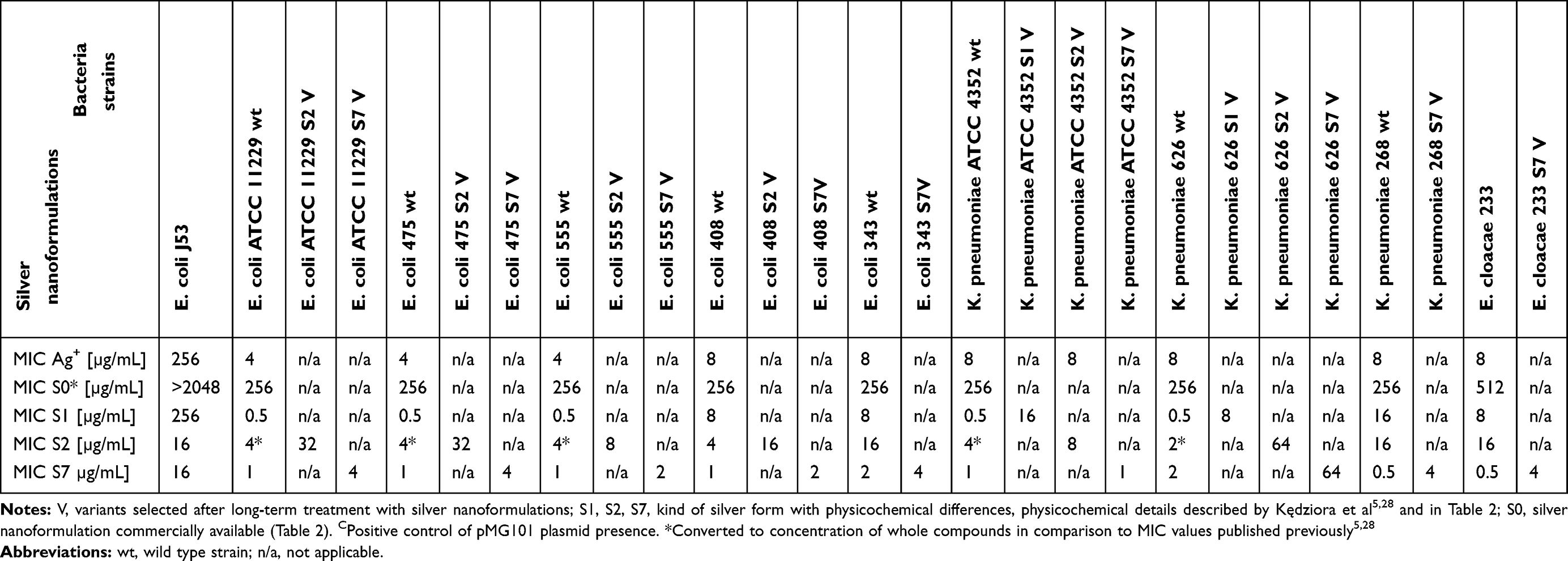 |
Table 3 MIC Of Different Silver Nanoformulations Against Tested Gram-Negative Bacteria Strains |
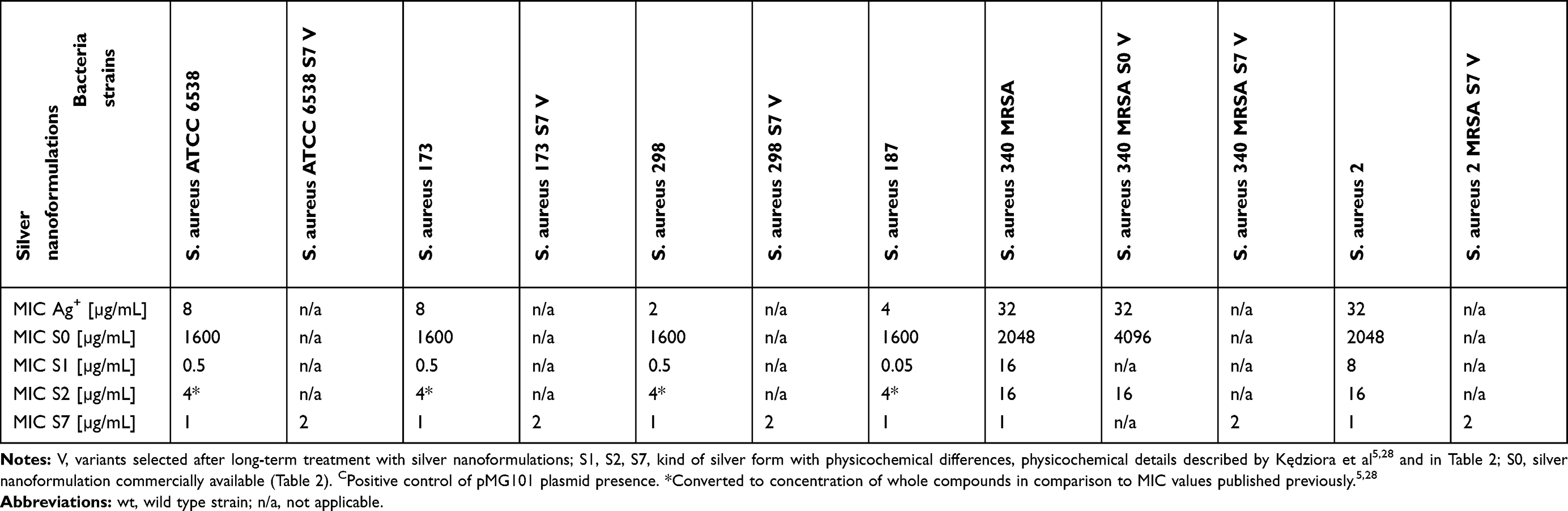 |
Table 4 MIC Of Different Silver Nanoformulations Against Tested Gram-Positive Bacteria Strains |
 |
Table 5 The Changes Of Bacterial Sensitivity To Silver Nanoformulations After Their Repeated Exposure |
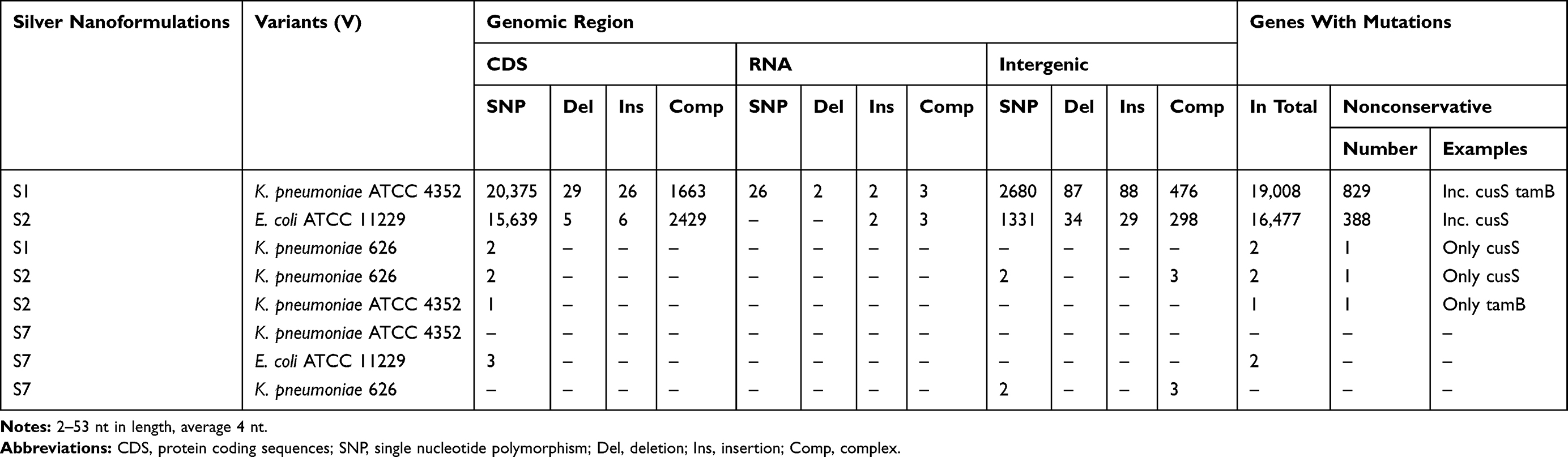 |
Table 6 Number And Distribution Of Genomic Mutations Acquired By Selected Test Variants (V) Due To Long-Term Exposure For Different Silver Nanoformulations |
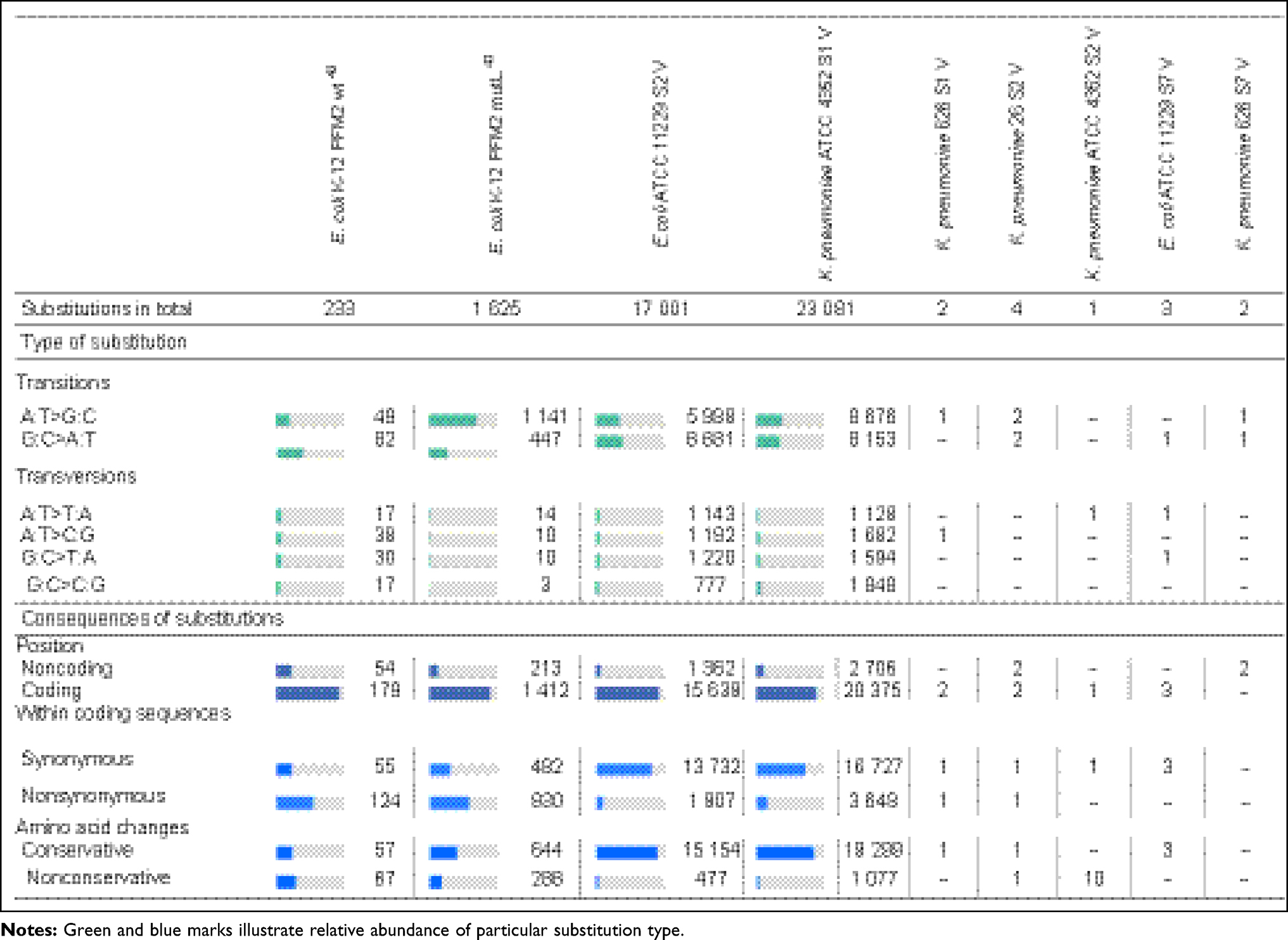 |
Table 7 Substitution Counts From Whole Genome Sequencing As Divided For Transitions And Transversions, Followed By Distribution Of Substitutions In Coding Regions And Possible Impact On Coded Protein Structure |
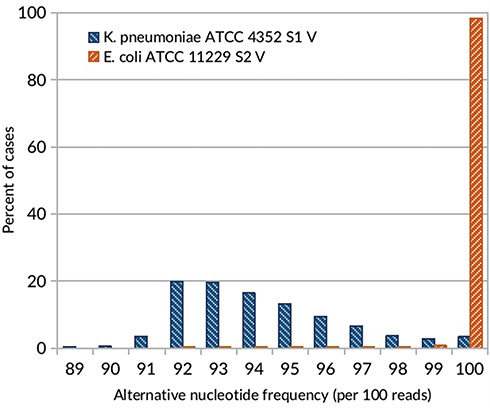 |
Figure 2 A histogram showing mutation frequency counts in genomic sequencing of Escherichia coli ATCC 11229 S2 V and Klebsiella pneumoniae ATCC 4352 S1 V strains. |
Among all of the bacteria strains tested by us, both Gram-negative and Gram-positive, the presence of sil genes, located on the plasmid pMG101, was checked. We indicated that 2 strains possess some sil genes: silE, silRS, silCBA, silF, and silBwere present in E. coli ATCC 11229 and silBin K. pneumoniae 626 (Table 8). It is worth underlining that those strains showed high sensitivity to silver ions and tested silver nanoformulations (excluding S0) (Table 3). Probably, this results from incompleteness of the pMG101 plasmid. In most of these strains the lack of all checked sil genes was confirmed. All results were visualized with electrophoresis (Figure 3).
 |
Table 8 Presence Of sil Genes In MDR Bacteria Strains wt |
 |
Figure 3 Electropherograms of Escherichia coli ATCC 11229 wt and Klebsiella pneumoniae 626 wt sil genes. |
Finally, the antibiotic susceptibility of variants was checked. Results are summarized in Tables 9 and 10. Results indicated that 4 variants (E. coli 475 S2 V and E. cloacae 233 S7 V among Gram-negative bacteria and S. aureus ATCC 6538 S7 V and S. aureus 340 S7 V among Gram-positive bacteria) became more resistant to antibiotic (ciprofloxacin in the case of Gram-negative bacteria and few classes in Gram-positive strains) (Tables 9 and 10). All of them were obtained after exposition to different kinds of silver nanoformulations: S2 and S7. Remaining Gram-negative and Gram-positive variants maintained the level of antibiotic susceptibility or became more sensitive to some of the antibiotic classes. Almost all of the S7 variants became more sensitive to certain classes of antibiotics. One of the possible mechanisms of bacterial resistance to antibiotics is the inhibition of antibiotic penetration into the bacteria cell.44–48 In this case it may be related to higher envelope permeability after silver nanoformulation treatment. On the other hand, Barras et al49 reviewed that incorporation of silver into antibiotics can increase their antibacterial activity. Anuj et al50 confirmed that cationic particles of nanosilver disturb the membrane integrity in bacteria strains, increasing their permeability in E. coli for linezolid, its higher intracellular concentration, and better antibacterial efficacy. A similar study was performed by Kaur et al51 who conjugated vancomycin with silver nanoparticles and enhanced antibacterial activity against both classes of bacteria: Gram-positive S. aureus and Gram-negative E. coli. However, Mühling et al52 showed that environmental exposure of resistant bacteria to silver nanoparticles did not increase antibiotic resistance in naturally occurring strains. They proved that there is no interaction between the antibiotics and Ag. As we reviewed,9 the interaction of bacteria with silver nanoformulations and consequences depends on the physicochemical properties of silver. The size, shape, biodiversity, and active surface determine their mode of action and the cell answer.
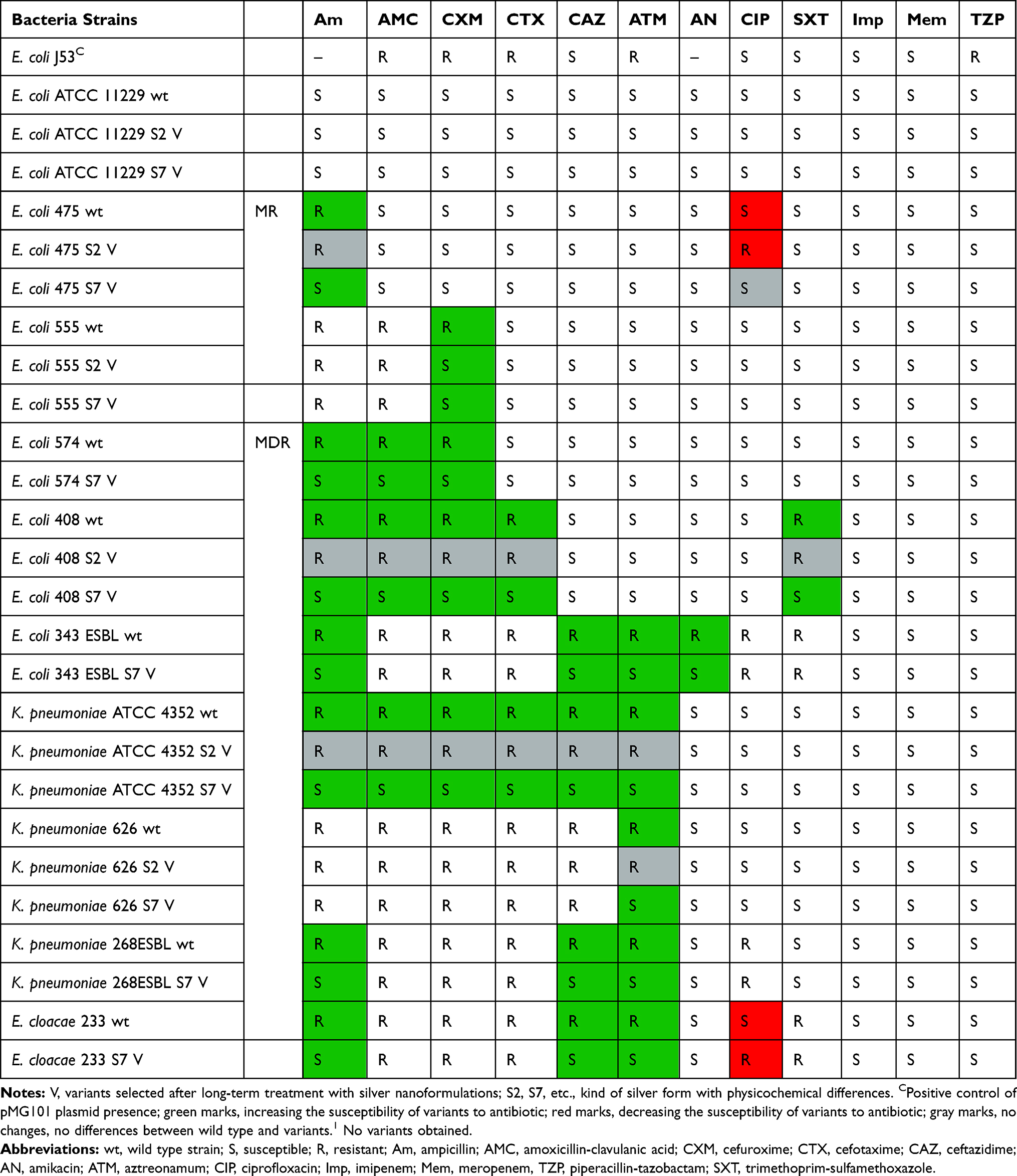 |
Table 9 Antibiotics Susceptibility Of Gram-Negative Bacteria Strains |
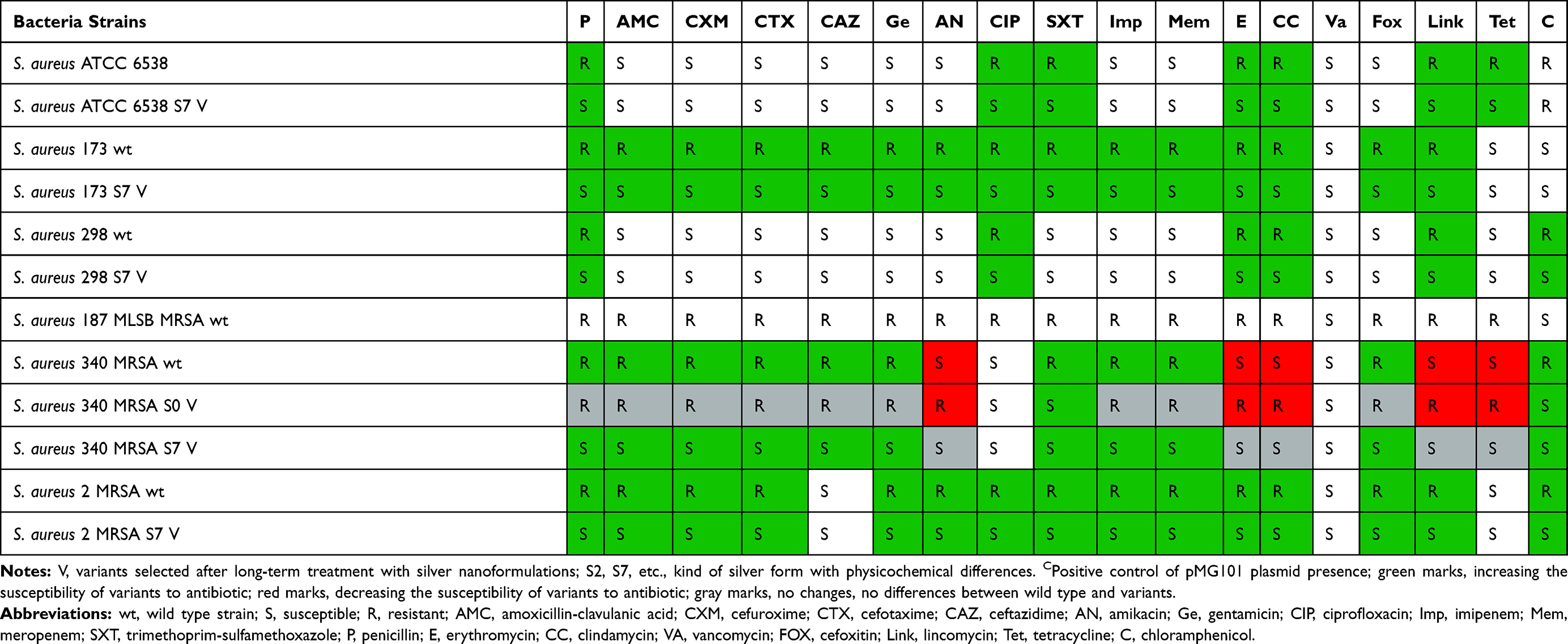 |
Table 10 Antibiotics Susceptibility Of Gram-Positive Bacteria Strains |
Conclusion
Silver nanoformulations with different physicochemical properties exert selective pressure on the bacterial strain population. Application of silver in the case of MR or MDR bacterial infection may cause different bacterial responses to antibiotics. Based on our results and the analyzed literature we may conclude that the response of bacteria cells to silver nanoparticles depends on the physicochemical properties of the nanoformulations (such as size, shape, charge, surface area, compounds) and individual features of bacteria strains (such as structural compounds and metabolism). The incorporation of silver into industrial products should be under strong control. Moreover, each silver nanomaterial should be considered as a separate agent with a potential different mode of antibacterial action.
Highlights
- Silver nanoformulations, in dependence on physicochemical properties, can exert selective pressure on the bacterial cells, decreasing the sensitivity to silver.
- Phenotypical changes, as a consequence of long-term exposure to silver nanoformulations, can change their sensitivity to antibiotics, including increasing bacterial susceptibility to some classes of antibiotics.
- Silver nanoformulations may cause mutational or phenotypical changes in dependence on physicochemical properties.
- Development of bacterial resistance to silver nanoformulations depends on their physicochemical properties and individual features of the bacteria cell (e.g. cell structure, cell compounds, and metabolism)
Acknowledgments
The authors thank Dialab for providing clinically strains for this study, Dorota Kiewra (PhD) and Aleksandra Czułowska (MSc) from Department of Microbial Ecology and Environmental Protection for making Thermal Cycler available, and Paulina Domagała for the laboratory support.
Funding
This work was partially supported by the National Science Centre [grants number 2017/01/X/NZ1/00765 and UMO-2016/23/B/ST5/02480]; the special-purpose grant of the Ministry of Science and High Education to carry out research or development work and tasks related to the development of young scientists and doctoral participants [grant number 1215/M/IGM/15 and 0420/2559/18].
Disclosure
Dr Anna Kędziora and Dr Gabriela Bugla-Płoskońska report grants from University of Wroclaw. The authors report no other conflicts of interest in this work.
References
1. Wright GD. Antibiotic resistance in the environment: a link to the clinic? Curr Opin Microbiol. 2010;13:589–594. doi:10.1016/j.mib.2010.08.005
2. Gaikwad S, Ingle A, Gade A, et al. Antiviral activity of mycosynthesized silver nanoparticles against herpes simplex virus and human parainfluenza virus type 3. Int J Nanomedicine. 2013;8:4303–4314. doi:10.2147/IJN.S50070
3. Rahisuddin AL-TSA, Khan Z, Manzoor N. Biosynthesis of silver nanoparticles and its antibacterial and antifungal activities towards gram-positive, gram-negative bacterial strains and different species of Candida fungus. Bioprocess Biosyst Eng. 2015;38:1773–1781. doi:10.1007/s00449-015-1418-3
4. Selvaraj M, Pandurangan P, Ramasami N, Rajendran SB, Sangilimuthu SN, Perumal P. Highly potential antifungal activity of quantum-sized silver nanoparticles against Candida albicans. Appl Biochem Biotechnol. 2014;173:55–66. doi:10.1007/s12010-014-0782-9
5. Kedziora A, Strek W, Kepinski L, Bugla-Ploskonska G, Doroszkiewicz W. Synthesis and antibacterial activity of novel titanium dioxide doped with silver. J Sol-Gel Sci Technol. 2012;62:79–86. doi:10.1007/s10971-012-2688-8
6. Kędziora A, Krzyżewska E, Dudek B, Bugla-Płoskońska G. The participation of outer membrane proteins in the bacterial sensitivity to nanosilver. Postepy Hig Med Dosw. 2016;70:610–617. doi:10.5604/17322693.1205005
7. Thiel J, Pakstis L, Buzby S, et al. Antibacterial properties of silver-doped titania. Small. 2007;3:799–803. doi:10.1002/(ISSN)1613-6829
8. Recommendation of European Union 2011/696/UE. Official Journal of the European Union L 275, 20.10.2011.
9. Kędziora A, Speruda M, Krzyżewska E, Rybka J, Łukowiak A, Similarities B-PG. Differences between silver ions and silver in nanoforms as antibacterial agents. Int J Mol Sci. 2018;19:444. doi:10.3390/ijms19020444
10. McGillicuddy E, Murray I, Kavanagh S, et al. Silver nanoparticles in the environment: sources, detection and ecotoxicology. Sci Total Environ. 2017;575:231–246. doi:10.1016/j.scitotenv.2016.10.041
11. Rónavári A, Kovács D, Igaz N. Biological activity of green-synthesized silver nanoparticles depends on the applied natural extracts: a comprehensive study. Int J Nanomedicine. 2017;12:871–883. doi:10.2147/IJN
12. Silver S. Bacterial silver resistance: molecular biology and uses and misuses of silver compounds. FEMS Microbiol Rev. 2003;27:341–353. doi:10.1016/S0168-6445(03)00047-0
13. Randall CP, Gupta A, Jackson N, Busse D, O’Neill AJ. Silver resistance in Gram-negative bacteria: a dissection of endogenous and exogenous mechanisms. J Antimicrob Chemother. 2015;70:1037–1046.
14. Silver S, Gupta A, Matsui K, Lo JF. Resistance to Ag(I) cations in bacteria: environments, genes and proteins. Met Based Drugs. 1999;6:315–320.
15. Finley PJ, Norton R, Austin C, Mitchell A, Zank S, Durham P. Unprecedented silver resistance in clinically isolated enterobacteriaceae: major implications for burn and wound management. Antimicrob Agents Chemother. 2015;59:4734–4741. doi:10.1128/AAC.00026-15
16. Asiani KR, Williams H, Bird L, et al. SilE is an intrinsically disordered periplasmic “molecular sponge” involved in bacterial silver resistance. Mol Microbiol. 2016;101:731–742. doi:10.1111/mmi.2016.101.issue-5
17. Randall CP, Oyama LB, Bostock JM, Chopra I, O’Neill AJ. The silver cation (Ag+): antistaphylococcal activity, mode of action and resistance studies. J Antimicrob Chemother. 2013;68:131–138. doi:10.1093/jac/dks372
18. Cochrane C, Loh J, Percival S, Williams N, Woods E. Silver resistance in MRSA isolated from wound and nasal sources in humans and animals. Int Wound J. 2009;6:32–37. doi:10.1111/j.1742-481X.2008.00563.x
19. WHO GAP AMR Newsletter No.32: Implementation of the global action plan on antimicrobial resistance, November 2017.
20. European Centre for Disease Prevention and Control. Surveillance and disease data for antimicrobial resistance. Available from: https://ecdc.europa.eu/en/antimicrobial-resistance/surveillance-and-disease-data.
21. Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18:268–281. doi:10.1111/j.1469-0691.2011.03570.x
22. Nordmann P, Poirel L. The difficult-to-control spread of carbapenemase producers among enterobacteriaceae worldwide. Clin Microbiol Infect. 2014;20:821–830. doi:10.1111/1469-0691.12719
23. Nordmann P. Carbapenemase-producing Enterobacteriaceae: overview of a major public health challenge. Med Mal Infect. 2014;44:51–56. doi:10.1016/j.medmal.2013.11.007
24. Klevens RM, Morrison MA, Nadle J, et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA. 2007;298:1763–1771. doi:10.1001/jama.298.15.1763
25. Lindsay JA. S. aureus evolution: lineages and Mobile Genetic Elements (MGEs). In: Lindsay JA, editor. Staphylococcus: Molecular Genetics. Norfolk, UK: Caister Academic Press; 2008:45–69.
26. Glasner C, Albiger B, Buist G, et al. Carbapenemase-producing enterobacteriaceae in Europe: a survey among national experts from 39 countries, February 2013. Euro Surveill. 2013;18. doi:10.2807/1560-7917.ES2013.18.28.20525
27. Woods EJ, Cochrane CA, Percival SL. Prevalence of silver resistance genes in bacteria isolated from human and horse wounds. Vet Microbiol. 2009;138:325–329. doi:10.1016/j.vetmic.2009.03.023
28. Kedziora A, Korzekwa K, Strek W, Pawlak A, Doroszkiewicz W, Bugla-Ploskonska G. Silver nanofilms as a therapeutic agent for killing Escherichia coli and certain ESKAPE pathogens. Curr Microbiol. 2016;73:139–147. doi:10.1007/s00284-016-1034-8
29. National Committee for Clinical Laboratory Standards. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically approved standard. Seventh edition. M7-A5. 2006. 26(2).
30. Kędziora A, Speruda M, Krzyżewska E, et al. Outer membrane proteins of bacteria as change’s marker of sensitivity to silver nanoforms. Acta Biochim Pol. 2015;62(suppl. 2):131.
31. Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal. 2011;17(1):10. doi:10.14806/ej.17.1
32. Bankevich A, Nurk S, Antipov D, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19:455–477. doi:10.1089/cmb.2012.0021
33. Gurevich A, Saveliev V, Vyahhi N, Tesler G. QUAST: quality assessment tool for genome assemblies. Bioinformatics. 2013;29:1072–1075. doi:10.1093/bioinformatics/btt086
34. Darling AC, Mau B, Blattner FR, NT P. Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004;14:1394–1403. doi:10.1101/gr.2289704
35. Darling AE, Mau B, Perna NT. progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS One. 2010;5:e11147. doi:10.1371/journal.pone.0011147
36. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30:2068–2069. doi:10.1093/bioinformatics/btu153
37. Seemann T Snippy: rapid haploid variant calling and core genome alignment. Available from: https://github.com/tseemann/snippy.
38. European Committee on Antimicrobial Susceptibility Testing Breakpoint tables for interpretation of MICs and zone diameters (version 8.1). Valid from May 15, 2018.
39. Henikoff S, Henikoff JG. Amino acid substitution matrices from protein blocks. Proc Natl Acad Sci USA. 1992;89:10915–10919. doi:10.1073/pnas.89.22.10915
40. Lee H, Popodi E, Tang H, Foster PL. Rate and molecular spectrum of spontaneous mutations in the bacterium Escherichia coli as determined by whole-genome sequencing. Proc Natl Acad Sci USA. 2012;109:E2774–E2783. doi:10.1073/pnas.1210309109
41. Foster PL, Lee H, Popodi E, Townes JP, Tang H. Determinants of spontaneous mutation in the bacterium Escherichia coli as revealed by whole-genome sequencing. Proc Natl Acad Sci USA. 2015;112:E5990–E5999. doi:10.1073/pnas.1512136112
42. Muthukrishnan L, Nanda A. Geno-toxic study of silver bio-nanoparticles toward gram-positive and gram-negative clinical isolates. J Pharm Res. 2013;6:725–729.
43. Panacek A, Kvitek L, Smekalova M, et al. Bacterial resistance to silver nanoparticles and how to overcome it. Nat Nanotechnol. 2018;13:65–71. doi:10.1038/s41565-017-0013-y
44. Tang SS, Apisarnthanarak A, Hsu LY. Mechanisms of β-lactam antimicrobial resistance and epidemiology of major community- and healthcare-associated multidrug-resistant bacteria. Adv Drug Deliv Rev. 2014;78:3–13. doi:10.1016/j.addr.2014.08.003
45. Rodriguez-Martinez JM, Machuca J, Cano ME, et al. Plasmid-mediated quinolone resistance: two decades on. Drug Resist Updat. 2016;29:13–29. doi:10.1016/j.drup.2016.09.001
46. Davies J, Wright GD. Bacterial resistance to aminoglycoside antibiotics. Trends Microbiol. 1997;5:234–240. doi:10.1016/S0966-842X(97)01033-0
47. Kester M, Vrana K. Treatment of Infectious Diseases. In: Kester M, Karpa K, Vrana K, editors. Elsevier’s Integrated Review Pharmacology.
48. Pyorala S, Baptiste KE, Catry B, et al. Macrolides and lincosamides in cattle and pigs: use and development of antimicrobial resistance. Vet J. 2014;200:230–239. doi:10.1016/j.tvjl.2014.02.028
49. Barras F, Aussel L, Ezraty B. Silver and antibiotic, new facts to an old story. Antibiotics (Basel). 2018;7. doi:10.3390/antibiotics7030077
50. Anuj SA, Gajera HP, Hirpara DG, Golakiya BA. Bacterial membrane destabilization with cationic particles of nano-silver to combat efflux-mediated antibiotic resistance in gram-negative bacteria. Life Sci. 2019;230:178–187. doi:10.1016/j.lfs.2019.05.072
51. Kaur A, Preet A, Kumar V, Kumar R, Kumar R. Synergetic effect of vancomycin loaded silver nanoparticles for enhanced antibacterial activity. Colloids Surf B Biointerfaces. 2019;176:62–69. doi:10.1016/j.colsurfb.2018.12.043
52. Mühling M, Bradford A, Readman JW, Somerfield PJ, Handy RD. An investigation into the effects of silver nanoparticles on antibiotic resistance of naturally occurring bacteria in an estuarine sediment. Mar Environ Res. 2009;68:278–283. doi:10.1016/j.marenvres.2009.07.001
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.