Back to Journals » Infection and Drug Resistance » Volume 19
Confronting blaNDM-5 in Salmonella Typhi: From Molecular Epidemiology, Resistance Mechanism to Clinical Management
Received 14 November 2025
Accepted for publication 23 March 2026
Published 16 April 2026 Volume 2026:19 581658
DOI https://doi.org/10.2147/IDR.S581658
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Hazrat Bilal
Xiaolan Guo, Jia Wu, Ke Li
Clinical Laboratory, Shenshan Medical Centre, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Shanwei, Guangdong, People’s Republic of China
Correspondence: Ke Li, Clinical Laboratory, Shenshan Medical Centre, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University, Shanwei, Guangdong, People’s Republic of China, Email [email protected]
Abstract: The emergence of Carbapenem-Resistant Salmonella enterica serovar Typhi (CR Salmonella enterica var typhi) represents a critical challenge in the prevention and control of typhoid fever. In recent years, CR Salmonella enterica var typhi strains carrying the blaNDM-5 gene have rapidly disseminated worldwide, particularly across South Asia such as India and Pakistan, severely exacerbating the clinical treatment failure rates and significant mortality. Due to the acceleration of global travel and trade, the multidrug resistant clones mainly transmitted through plasmids (such as lncX3), increasingly endanger China with historically low resistance levels. The CR Salmonella enterica var typhi strains exhibit extensive resistance to fluoroquinolones, cephalosporins, and carbapenems, compromising the effectiveness of conventional treatment regimens. Herein, efforts are made to provide a comprehensive review on the global epidemiology, underlying resistance mechanisms, the evolving challenges in clinical management. The antimicrobial resistance surveillance systems, such as whole-genome sequencing (WGS) technology, strict antibiotic management, infection control measures, and future perspectives on the combination therapies (such as meropenem combined with polymyxin) have also been discussed and outlooked. It is highly anticipated that the surveillance technologies combined with potential therapeutic pathways will witness a leap-forward development in clinical translation for public health.
Keywords: Salmonella enterica, carbapenem resistance, blaNDM-5 global dissemination
Introduction
Global Burden of Disease from Salmonella Typhi Infection
Typhoid fever, caused by Salmonella Typhi, continues to pose a significant global health threat, particularly in low- and middle-income countries (LMICs) of South Asia, Southeast Asia, and sub-Saharan Africa.1,2 The World Health Organization (WHO) has classified drug-resistant Salmonella Typhi as a high-priority pathogen, underscoring the urgency of this challenge.3 The disease burden is substantial, with an estimated 11 to 21 million cases and 128,000 to 161,000 deaths annually.2 Beyond mortality, the economic and social impacts are profound. The annual cost of illness associated with typhoid fever is considerable, encompassing direct medical expenses and indirect productivity losses. Furthermore, the disability-adjusted life years (DALYs) lost due to typhoid, particularly its drug-resistant forms, highlight its severe impact on healthy life expectancy. For instance, reports from the CDC indicate that drug-resistant infections lead to longer hospitalizations and poorer outcomes, exacerbating this burden (doi: 10.15585/mmwr.mm7207a2). The evolution of Salmonella Typhi from classical multidrug resistance (MDR) to carbapenem resistance represents a critical escalation.4,5 The failure of carbapenems, the last line of defense, marks a new era in the treatment crisis and underscores the pressing need for this review (Figure 1).6,7
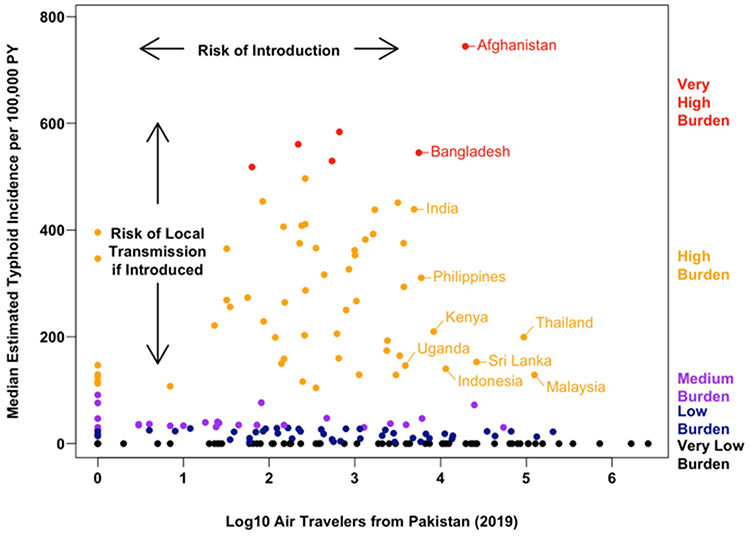 |
Figure 1 Air travel from Pakistan was used as a proxy for the risk of XDR typhoid introduction, while the estimated typhoid burden was used as a proxy for the risk of onward transmission and local outbreaks. Source data available in a separate file. Reproduced with permission. Copyright 2023, Springer Nature. |
The Global Epidemic Trend of Carbapenem-Resistant Salmonella Typhi
From the classical multidrug resistance (MDR) chloramphenicol/ampicillin/trimethoprim-sulfamethoxazole (TMP-SMX), there has been a notable evolution towards carbapenem resistance. Carbapenem antibiotics were previously regarded as the last line of defense, and their failure marks a comprehensive escalation in the treatment crisis.5 Notably, the emergence of CR Salmonella enterica var typhi represents a distinct manifestation of the carbapenem-resistant Enterobacteriaceae (CRE) crisis, posing a threat that exceeds that of typical CRE due to its combination of high pathogenicity and extensivedrug resistance.
The emergence and global spread of CR Salmonella enterica var typhi were formally confirmed in 2018 when Klemm et al used whole-genome sequencing (WGS) to identify a strain isolated from a child in Pakistan in 2016.5 The subsequent case report from Pakistan, the recognized epicenter of the CR Salmonella enterica var typhi epidemic, revealed the clinical severity of CR Salmonella enterica var typhi infection.8 Genomic surveillance has confirmed the rapid spread of the H58 lineage clone carrying the blaNDM-5 gene on IncX3 within Pakistan.4
Although other carbapenemase genes, such as blaNDM-1, blaVIM, blaKPC, blaOXA-48, and blaIMP, are frequently identified in various Enterobacteriaceae, their presence in Salmonella Typhi remains exceedingly rare. To date, the vast majority of carbapenem-resistant Salmonella Typhi isolates worldwide carry blaNDM-5, highlighting its dominant role in the current epidemiology. The sporadic reports of other carbapenemases in non-typhoidal Salmonella serovars (discussed below) underscore the potential for horizontal transfer, but they have not yet become established in Salmonella Typhi.
While local infections of CR Salmonella enterica var typhi in neighboring India are extremely rare (the first case was reported in Bangalore in 2025, belonging to the input-related H58 lineage), the region faces a continuous threat due to the backdrop of carbapenem-resistant Enterobacteriaceae epidemics and the environmental prevalence of NDM genes. The identification of homologous, blaNDM-5-harboring IncX3 plasmids in strains from both India and Pakistan underscores the ongoing risk of cross-border transmission.9–11
Other countries primarily report imported cases, involving international travelers or individuals returning from epidemic areas. For instance, countries such as the United Kingdom, the United States, and Canada have reported travel-related cases.12 In 2022, Denmark first reported an imported case involving a pregnant woman.13 This pattern of global dispersal heightens vigilance worldwide. In China, although surveillance data from 2015 to 2022 identified only two blaNDM-5-positive strains among 3695 Salmonella isolates,14 Salmonella Typhimurium (ST34 clone) carrying blaNDM-5 has been detected in retail pork, indicating that the animal-food chain may serve as a potential transmission route.15,16 Additionally, non-typhoidal Salmonella from animal sources (such as pigs and chickens) carrying carbapenemase genes have been reported in various regions worldwide, including China,15–18 Germany,19,20 and Iraq,21 indicating the potential risk of the horizontal transfer of antibiotic resistance genes to Salmonella Typhi.16,17
The Discovery of blaNDM-5 and Its Significance in Salmonella Typhi
The resistance of CR Salmonella enterica var typhi to carbapenem antibiotics primarily arises from the production of carbapenemases, with metallo-β-lactamases (MBLs) being the most prevalent. Among MBLs, the New Delhi metallo-β-lactamase (NDM) type dominates, accounting for over 96.7% of CR Salmonella enterica var typhi cases, with the NDM-5 variant being particularly common (Figure 2).22–24
 |
Figure 2 Phylogenetic analysis of Salmonella enterica strains, including Salmonella Typhimurium ST19 and ST34 isolates from NCBI, alongside local strains. The figure illustrates the relationship between different Salmonella serotypes. The blaNDM-carrying status, sequence type (ST), host, isolation time, and region are represented by squares of different colors. Note that ST19 and ST34 are characteristic of Salmonella Typhimurium. |
NDM enzymes can efficiently hydrolyze nearly all β-lactam antibiotics, including carbapenems, the last-resort treatments for multidrug-resistant typhoid. This gene encoding NDM-5 was initially identified in Salmonella Typhi isolated from a patient in Pakistan, marking a pivotal moment in the evolution of drug-resistant typhoid. It is reported that the blaNDM-5 gene often co-occurs with resistance to fluoroquinolones and cephalosporins, creating extensively drug-resistant (XDR) phenotypes that significantly elevate the risk of treatment failure and mortality.9
Genomic analysis has shown that Salmonella Typhi strains carrying blaNDM-5 typically belong to the H58 haplotype, which is currently the globally prevalent antibiotic-resistant lineage.9 Additionally, blaNDM-5 is often co-located with antibiotic resistance genes such as aac(6′)-Ia, and may be transmitted between different strains via InCX3-type plasmids, which increases the risk of its sustained dissemination in both hospital and community settings. Therefore, continuous genomic surveillance is crucial for tracking the transmission dynamics of blaNDM-5 positive Salmonella Typhi and guiding treatment strategies.9,25
The Molecular Epidemiological Characteristics of blaNDM-5 Positive Salmonella Typhi in China
Geographical Distribution and Time Trend Analysis
Since the blaNDM-5 gene was first detected in Salmonella from food sources in 2016, its detection frequency in Salmonella in China has gradually increased, involving multiple sources such as environment,17 clinical,18 poultry26 and food.15 Between 2015 and 2022, two strains of Salmonella enterica carrying the blaNDM-5 gene were isolated from a total of 3695 strains, confirming the introduction of such resistant bacteria into Chinese healthcare settings.14 Genomic analyses revealed that these clinical strains are closely related to the XDR Salmonella enterica var typhi clones responsible for outbreaks in Pakistan and India, indicating a clear risk of cross-border transmission.14,27 Currently, reports of blaNDM-5 positive Salmonella Typhi in China have primarily been are mainly concentrated in the eastern coastal regions, which aligns with the characteristics of frequent international travel and dense medical resources in these areas.14 The main detections of the blaNDM-5 gene in various Salmonella serotypes (including Typhi and Typhimurium) across China are shown in Table 1.15–18,22,24,26,28–30
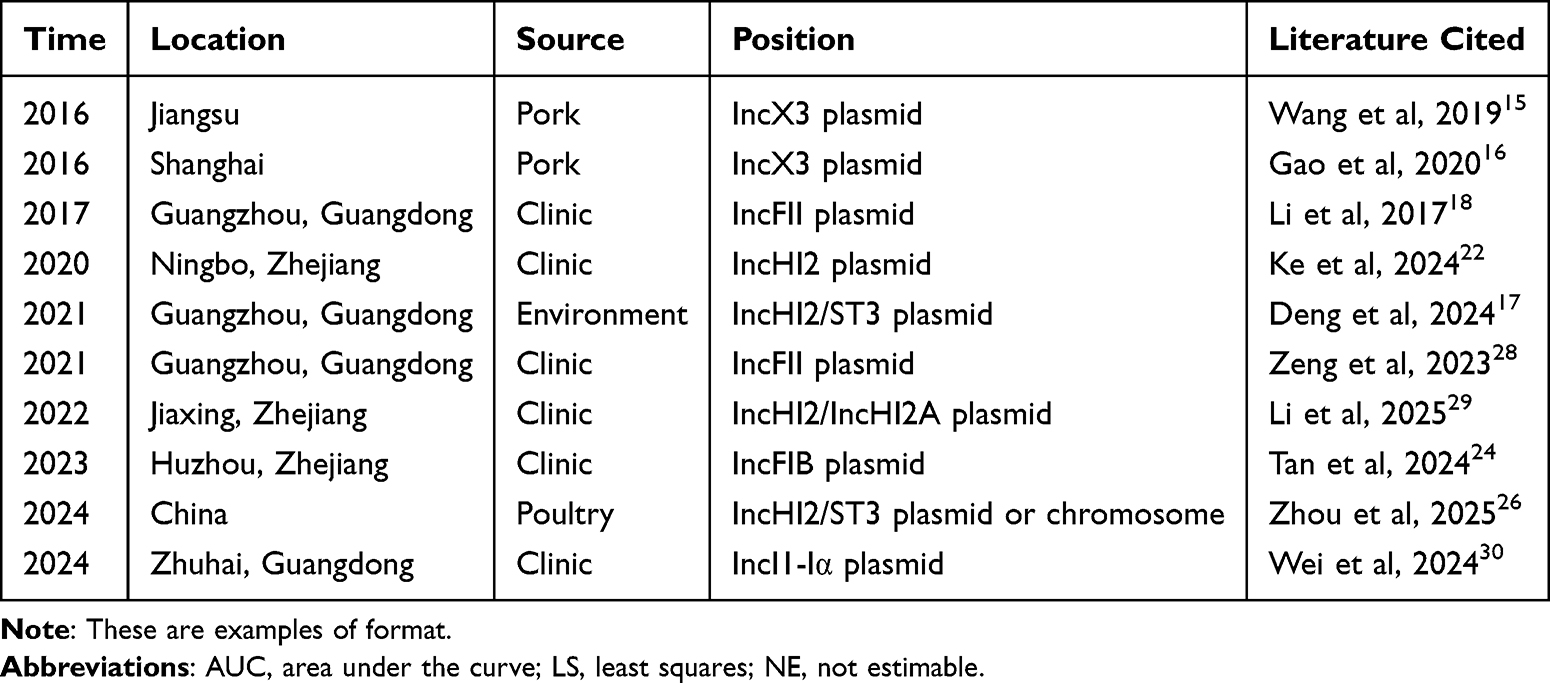 |
Table 1 The Main Detection of blaNDM-5 Gene in Salmonella in China |
MLST Typing Characteristics of the Main Epidemic Strains
Genomic analysis indicates that the blaNDM-5 positive Salmonella Typhi identified in China primarily belongs to the H58 haplotype, which is characterized by the presence of multiple drug resistance genes.31 Specifically, the ST20 strain accounts for a certain proportion of carbapenem-resistant bacterial strains in China.32 In line with global trends, these strains often carry the gyrAS83Y mutation, which leads to resistance to fluoroquinolone antibiotics.33 Notably, the Chinese isolates exhibited a high genomic similarity with the XDR Salmonella Typhi strains prevalent in the South Asian region, particularly the strains from the outbreak in Pakistan in 2016, indicating a potential common evolutionary origin.27 It is important to clarify that while this review focuses on Salmonella Typhi, comparative genomic analyses often include other serotypes. For instance, as shown in Figure 2 (adapted from Ke et al), ST19 and ST34 strains are Salmonella Typhimurium, highlighting the broader context of blaNDM-5 dissemination across different Salmonella serovars.22
Multidrug Resistance Spectrum Characteristics of Clinical Isolates
Clinical isolates of blaNDM-5 positive Salmonella Typhi in China exhibit an extensively drug-resistant (XDR) profile, characterized by multidrug resistance to ampicillin, ciprofloxacin, ceftriaxone, tetracycline, and meropenem, but typically remain susceptible to azithromycin and other alternative agents such as chloramphenicol and co-trimoxazole.27,34 These strains often carry aminoglycoside-modifying enzyme genes, such as aac(6′)-Ia, resulting in cross-resistance.33,34 Molecular epidemiological investigations have revealed that approximately 37.5% of the resistant strains carry the extended-spectrum β-lactamase gene blaCTX-M-15 gene, further compromising β-lactam efficacy.35 Notably, certain strains have disseminated the blaNDM-1 gene via IncFII(Yp) type plasmids, which have been detected in 46 strains of diverse sequence types across six sentinel monitoring sites, highlighting a parallel route for the spread of carbapenem resistance in Salmonella35 The primary resistance mechanisms and their genetic contexts are summarized in Figure 3.28
 |
Figure 3 The genetic surroundings of blaNDM-5 are depicted by diverse colored arrows, each denoting different open reading frames (ORFs) with the arrow’s orientation indicating transcription direction. Homogeneous areas are shown with light gray shading. Reproduced with permission. Copyright 2023, mSphere. |
The Genetic Vector Structure and Transmission Elements of blaNDM-5
The Core Genetic Structure of blaNDM-5
The blaNDM-5 gene is typically situated within a complex genetic context, characterized by its core structure, which comprises multiple insertion sequences (IS) and auxiliary genes. Notably, blaNDM-5 is frequently associated with the bleMBL (bleomycin resistance gene) and trpF (phosphoribosyl anthranilic acid isomerase gene), forming a stable gene cluster.36–38 In addition, the genetic background of blaNDM-5 often contains insertion sequences such as IS26, IS5, ISAba125 and IS3000, which play a key role in horizontal gene transfer and recombination.28,39
Plasmid Type Analysis
In popular clones of CR Salmonella enterica var typhi, the blaNDM-5 gene is primarily located on plasmids, which is a key factor for its efficient horizontal gene transfer (HGT) among different bacteria. The most common vector for the blaNDM-5 gene in Salmonella Typhi in China is the IncX3 type plasmid, which encodes a type IV secretion system (T4SS) responsible for plasmid conjugative transfer.40 Research by Gao et al has shown that the T4SS of IncX3 plasmids in certain Enterobacterales exhibits a phenomenon of apparent silencing, which may reduce the fitness burden on the host bacterium, thereby promoting the plasmid’s stable persistence and widespread dissemination within bacterial populations.40 This mechanism, observed in related species, is hypothesized to similarly contribute to the successful dissemination of IncX3 plasmids carrying blaNDM-5 in Salmonella Typhi. Additionally, blaNDM-5 has been found on IncFIA/FIB/IncFII fusion plasmids,16 with certain clinical isolates carrying as many as 5–10 copies of the blaNDM-5 gene.41,42 Notably, a study on Salmonella Typhi isolates from India identified the presence of the InCX3 plasmid carrying blaNDM-5,9 indicating the widespread distribution of this plasmid type in Asia. As stated in the literature, IncHI2-type plasmids are widely disseminated in the livestock-food chain across Asia, particularly in China,24 and the IncHI2/ST3 plasmid disseminates the blaNDM-5 gene among various Enterobacteriaceae species. IncX318 and IncFII-type plasmids have also been found to carry the blaNDM gene in human and animal/foodborne Salmonella, indicating their significant role in the cross-ecological transmission of antibiotic resistance genes.16,43 As shown in Figure 4b, there are similarities in the genetic structure of blaNDM-5 gene in different plasmids.
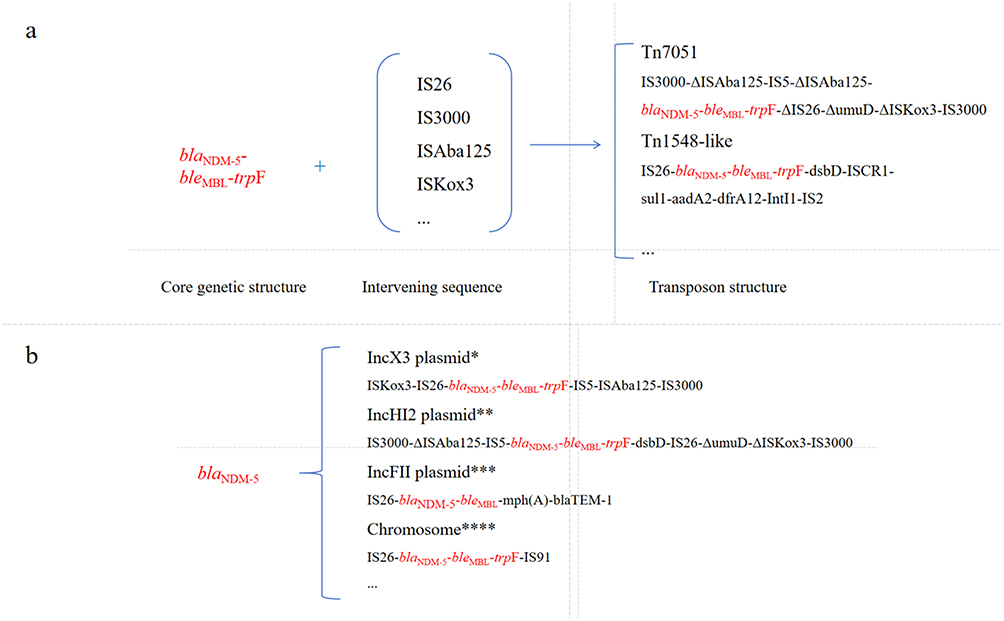 |
Figure 4 (a) Genetic structure of blaNDM-5 gene; (b) genetic structure of blaNDM-5 gene in different plasmids. * Xiang et al, 2023, Harada et al, 2024, ** Yang et al, 2022, Shin et al, 2023, *** Huang et al, 2025, ****Tang et al, 2024, Gonzalez et al, 2025. |
Structural Characteristics of Transposon Complex
The blaNDM-5 gene is commonly located within the structure of IS composite transposons.16,44 Research has shown that the blaNDM-5 region is situated on a composite transposon formed by IS elements, which includes a deletion of 366 bp in the Rep A protein-coding region. This structural variation may affect plasmid replication.45 In other Enterobacteriaceae, the dissemination of the blaNDM gene is primarily driven by the jumping of plasmid-borne transposons, followed by a restriction in plasmid exchange, which may be attributed to the adaptability of plasmids to their specific bacterial hosts.46 Experimental evidence indicates that the replicative transposition of Tn and IS elements facilitates the horizontal transfer of blaNDM and the evolution of plasmids that produce KPC-2.47 Similar composite transposon structures carrying blaNDM-5 have been identified in other Salmonella serovars (eg., Salmonella Typhimurium of porcine origin in China), reflecting the diversity of resistance element evolution. However, the primary focus of this review remains on Salmonella Typhi. Research indicates that the genetic environment of blaNDM-5 in CR Salmonella enterica var typhi is highly similar to that found in other Enterobacteriaceae, such as Escherichia coli and Klebsiella pneumoniae, strongly suggesting the interspecies transmission of antibiotic resistance genes.16 As shown in Figure 4a, the blaNDM-5 gene is often embedded in composite transposons, which are composed of multiple IS elements and auxiliary genes. For example, Tn7051 is a new type of transposon, its structure is IS3000-ΔISAba125-IS5-ΔISAba125-blaNDM-5-bleMBL-trpF-ΔIS26-ΔumuD-ΔISKox3-IS3000.37 This transposon was first discovered in IncHI2 plasmid and may play a key role in the transfer of blaNDM-5 gene from IncX3 plasmid to IncHI2 plasmid. In addition, in some IncFIB plasmids, the blaNDM-5 gene was inserted into the Tn1548-like transposon,48 and its structure was IS26-blaNDM-5-bleMBL-trpF-dsbD-ISCR1-sul1-aadA2-dfrA12-IntI1-IS2.
While the blaNDM-5 gene in Salmonella is predominantly plasmid-borne, studies in other Enterobacteriaceae have demonstrated its potential for chromosomal integration. For instance, Kong et al elucidated the chromosomal integration mechanism of blaNDM-1 in Proteus vulgaris through the SXT/R391 ICE.49 In Escherichia coli, the blaNDM-5 gene has been found inserted into the chromosome flanked by IS26 elements. Although direct evidence of chromosomal integration of blaNDM-5 in Salmonella spp.50 is currently lacking, these findings suggest a potential pathway for its stable inheritance and vertical transmission in the future, which could complement plasmid-mediated horizontal dissemination and lead to more persistent resistance reservoirs.
Co-Localization with Other Drug Resistance Genes
Carbapenem-resistant Salmonella Typhi (CR Salmonella enterica var typhi) commonly exhibits a broad spectrum of resistance, primarily characterized by an extensively drug-resistant (XDR) phenotype, which includes resistance to ampicillin, chloramphenicol, TMP-SMX, fluoroquinolones, and third-generation cephalosporins (such as ceftriaxone), as well as carbapenems. Whole-genome sequencing analysis revealed that blaNDM-5 often coexists with other resistance genes, including blaCTX-M-15, which is the most prevalent and mediates resistance to third-generation cephalosporins.9,51 Some strains carry both blaTEM-1 and blaSHV type β-lactamase genes.10,51 Mutations in the QRDR region: gyrA (Ser83Phe, Asp87Asn/Gly) and parC (Ser80Ile) mutations lead to high levels of resistance;51,52 Plasmid-mediated resistance genes: qnrS, aac(6′)-Ib-cr,51,53 Chloromycetin resistance: catA1/b3; Sulfonamide resistance: sul1/sul2/sul3; trimethoprim resistance: dfrA7/14/27; aminoglycoside resistance: aac(3)-IV, aadA1.43,53 A strain of Salmonella Typhi isolated in India was found to simultaneously harbor blaNDM-5 and aac(6′)-Ia genes.9 This co-localization phenomenon enhances the multidrug resistance of bacteria, posing a greater challenge for clinical treatment. Additionally, blaNDM-5 positive strains typically carry the gyrAS83Y mutation associated with fluoroquinolone resistance,9 further limiting treatment options. In certain cases, the blaNDM-5 gene coexists with the mcr-1 mediated colistin resistance gene,54 resulting in an extremely drug-resistant phenotype that poses a serious threat to public health.
Molecular Mechanism of Drug Resistance Transmission
Horizontal Gene Transfer
The blaNDM-5 gene is mainly located on plasmids. These plasmids are highly transferable and can be transmitted between different bacteria through conjugation. Common plasmid types include IncX3,55 IncHI2,56 IncFI.57 Whole-genome sequencing analyses have revealed that this gene is often located on InCX3-type plasmids and co-localizes with resistance genes such as aac(6′)-Ia.45 The positioning of insertion sequences IS3 and IS30 in the 5 kbp adjacent region of the bla-ble gene cluster indicates that transposon-mediated horizontal gene transfer is an important mode of dissemination.58 The IncY plasmid p60006, identified in Indian isolates, does not carry the carbapenemase gene; however, its presence confirms the potential for plasmid-mediated resistance transmission.11 Additionally, the blaNDM-5 gene can be amplified in multiple copies through IncFIA/FIB/IncFII plasmids, and this gene dosage effect significantly enhances the resistant phenotype.41
Vertical Transmission of Chromosomes
Although the blaNDM-5 gene is mainly present in plasmids, it has also been reported that it can be integrated into bacterial chromosomes. For example, in Escherichia coli, the blaNDM-5 gene is associated with insertion sequences such as IS26 and IS91, forming the structure of “IS26-blaNDM-5-bleMBL-trpF-IS91”.59,60 This chromosomal integration further increases the stability and transmission potential of the blaNDM-5 gene. The blaNDM-5 gene carried by the chromosome can be stably inherited and co-evolve with chromosomal resistance markers such as the gyrAS83Y mutation.45 Notably, certain isolates carried antibiotic resistance plasmids; however, no antimicrobial resistance gene clusters were found on the chromosome. This suggests that the loss of plasmids may lead to phenotypic reversion (Figure 5).11,61
 |
Figure 5 Both the IncFIA/IncFIB/IncFII consensus plasmid and the characteristics of the blaNDM-5-containing translocatable unit that were characterized from isolates 1–5 are summarized. Reproduced with permission. Copyright 2022, Clin Infect Dis.41 |
Environment-Animal-Human Cross-Host Transmission
In monitoring conducted in China, two blaNDM-5 positive enteric Salmonella strains were detected among 3695 Salmonella isolates, confirming the presence of antibiotic-resistant bacteria in the environment.62 The study of the transmission patterns of the mcr-9 gene in avian-derived S. Thompson strains provides a model for understanding the interspecies dissemination of antibiotic resistance genes.63 Monitoring of carbapenem-resistant Enterobacteriaceae in Thailand revealed that identical plasmids can be transmitted between different bacterial species, confirming gene flow at the environmental-clinical interface.64 NDM-5 positive Escherichia coli, Klebsiella pneumoniae, and Citrobacter spp. isolated from environmental samples and human feces exhibited the same antibiotic resistance plasmid profile, providing direct evidence of the transmission chain of resistance.65
Advances in Diagnostic Techniques and Surveillance Methods
The effective management of CR Salmonella enterica var typhi hinges on rapid, accurate diagnosis and robust surveillance. Technological advancements have enhanced capabilities across the entire spectrum, from phenotypic confirmation to genomic epidemiology.
Phenotypic Examination
For the phenotypic detection of blaNDM-5 positive Salmonella Typhi, the Vitek ID/(Antimicrobial Susceptibility Testing) AST system is widely used for preliminary identification and to profile resistance to critical antibiotics, such as carbapenems (eg, meropenem), third-generation cephalosporins (eg, ceftriaxone), and fluoroquinolones (eg, ciprofloxacin).66,67 Notably, these strains typically exhibit a multidrug resistance pattern to ampicillin, ciprofloxacin, ceftriaxone, tetracycline, and meropenem, while remaining sensitive to azithromycin, chloramphenicol, and trimethoprim-sulfamethoxazole.65,66 Serological typing serves as a supplementary diagnostic method to confirm the serotype of Salmonella Typhi.66
Molecular Detection
Molecular detection technologies for blaNDM-5 have rapidly developed. Online applications such as Pathogenwatch can quickly identify antibiotic resistance markers in genomes, and their clustering analysis is comparable to traditional bioinformatics methods.68 Whole genome sequencing technologies, such as the Illumina platform, can simultaneously detect the presence of resistance genes such as blaNDM-5 and aac(6′)-Ia, as well as the IncX3 plasmid.9,66 In a large-scale study conducted in Singapore, whole-genome sequencing was successfully applied to the analysis of 222 carbapenem-resistant non-susceptible strains, identifying multilocus sequence typing and antibiotic resistance gene profiles.69 These technologies hold significant value for the early detection of outbreaks of NDM-5 positive strains.9,64
Whole Genome Sequencing (WGS)
Whole-genome sequencing (WGS) provides the most comprehensive information, including all known and potentially unknown resistance genes, mutations, plasmid types, serotypes, multilocus sequence typing (MLST), and single nucleotide polymorphism (SNP) typing for high-resolution phylogenetic analysis. It has become a key tool for studying the transmission of antibiotic resistance in Salmonella Typhi.
Outbreak Investigation and Source Tracking
In outbreak investigations, WGS can accurately identify the phylogenetic relationships between strains, such as the close association between an Indian isolate and the 2016 XDR outbreak strain from Pakistan.9,65
Unveil Transmission Dynamics
Through the genomic analysis of 3489 strains of Salmonella enterica serovar Typhi, researchers were able to trace the spatiotemporal transmission patterns of antibiotic-resistant strains,25,70 while studies in Malawi used WGS to find the H58 genotype similarity between cases and isolates from carriers (Figure 6). 71,72
 |
Figure 6 (a) Monthly distribution of Salmonella enterica var typhi genotypes in cases. (b) Monthly distribution of Salmonella enterica var typhi genotypes in carriers. Note that counts include all culture-positive Salmonella enterica var typhi participants and those culture-positive for other Salmonella but later identified as Salmonella enterica var typhi by WGS. Multiple introductions of multidrug-resistant typhoid associated with acute infection and asymptomatic carriage, Kenya. Reproduced with permission. Copyright 2021, eLife. |
Discover Novel Resistance Mechanisms
WGS plays an irreplaceable role in identifying novel antibiotic-resistant plasmids (such as IncFII(Yp)) and tracking cross-regional transmission chains.35,67 Large-scale studies, such as one analyzing 222 carbapenem-non-susceptible isolates in Singapore, demonstrate its power in defining population structure and resistomes.69
Clinical Treatment Challenges
Clinical Manifestations and Treatment Challenges
Carbapenem-resistant Salmonella enterica serotype Typhi (CR Salmonella enterica var typhi) causes typhoid fever. Although the cardinal symptoms (persistent high fever, relative bradycardia, rose spots, and hepatosplenomegaly) are indistinguishable from those caused by non-resistant strains, the risk of severe complications—including intestinal hemorrhage, perforation, toxic encephalopathy, and septic shock—is significantly increased. The increased severity is primarily attributed to diagnostic delays (due to insufficient sensitivity of laboratory tests) and delays in effective antimicrobial treatment.
Epidemiological data indicate that a treatment delay of more than 72 hours can increase the risk of intestinal perforation.73 Owing to widespread resistance to frontline antibiotics (such as ampicillin, compound sulfamethoxazole, chloramphenicol), fluoroquinolones (such as ciprofloxacin), and third-generation cephalosporins (such as ceftriaxone), traditional treatment regimens have failed. The subsequent misuse of ineffective antibiotics exacerbates treatment delays, driving the increased incidence of complications (such as intestinal perforation and neurological symptoms), and elevated mortality rates, which can reach up to 26% in untreated cases.73 In a case reported from Pakistan, strains initially susceptible to carbapenems and azithromycin acquired the blaNDM-5 gene during treatment, resulting in ultimate treatment failure.74 This dynamic evolution of drug resistance poses a significant challenge to clinical treatment, particularly in resource-limited areas.75
Adverse Prognosis
Consequently, CR Salmonella enterica var typhi infection is associated with a profoundly worse prognosis compared to non-resistant or MDR/XDR (non-CR) typhoid fever. The cascade of treatment delays and failures results in markedly prolonged hospitalization, with the average length of hospital stay extended by 2 to 3 times, and a 5 to 10-fold increase in direct medical costs),12,53 critically elevating mortality rates. Although the specific mortality rates vary due to patients’ baseline conditions, severity of infection, timeliness of treatment, and effectiveness of treatment protocols, multiple studies have reported that the mortality rate associated with CR infections is elevated by 3–5 times compared to sensitive strains. For instance, a study conducted in Pakistan indicated that the mortality rate for CR infections reached 15–20%, whereas infections with sensitive strains only accounted for 1–2%.76
Therapeutic Strategies and Future Outlook
The management of CR Salmonella enterica var typhi infections necessitates a multifaceted approach, combining current clinical strategies with innovative research to overcome the challenges of extensive drug resistance.
Current Cornerstone: Combination Therapy
In the absence of reliably effective monotherapy, combination therapy has become the clinical mainstay. Intravenous administration of meropenem in combination with polymyxin has proven effective in treating infections caused by blaNDM-5 positive Salmonella Typhi.74 Notably, in numerous reported cases of multidrug-resistant Salmonella Typhi in Canada, the United States, and Europe, all isolated strains exhibited resistance to ampicillin, ceftriaxone, ciprofloxacin, and trimethoprim-sulfamethoxazole, underlying the importance of combination therapy strategies.71 However, therapy must be guided by AST, as evidenced by cases from India where strains carrying blaNDM-5 unexpectedly retained susceptibility to carbapenems, highlighting the critical need for individualized treatment plans.11
Innovative Approaches
Significant progress has been made in developing novel β-lactamase inhibitors against blaNDM-mediated carbapenem resistance. Ceftazidime-taniborbactam, a β-lactam/β-lactamase inhibitor combination under investigation, demonstrates good activity against Enterobacteriaceae expressing serine and metallo-β-lactamases.77 Silencing resistance genes, as demonstrated by the significant reduction in resistance observed upon silencing the blaNDM-1 gene in Klebsiella pneumoniae, provides a novel theoretical framework for antibacterial development.78
Novel Antimicrobial Strategies
Research is increasingly focused on innovative strategies that disarm bacterial defenses or enhance drug delivery to the site of infection. In drug delivery systems, novel macrophage-targeted mannose-modified hyaluronic acid precursor drug delivery systems have demonstrated potential for treating Salmonella Typhi infections.79 These innovative approaches provide new treatment options for infections caused by blaNDM-5 positive Salmonella Typhi.80 Recent research from Cornell University has demonstrated that rifampicin, a widely utilized antibiotic, exhibits up to 99.9% efficacy against multidrug-resistant Salmonella enterica var typhi. This therapeutic potential is attributed to its capacity to effectively disassemble the bacterium’s protective capsule.27 The bacterial capsule plays several critical roles in pathogenesis, including mediating immune evasion, facilitating host cell invasion and biofilm formation, and directly obstructing antibiotic penetration. Rifampicin effectively dismantles the Vi capsule of Salmonella enterica var typhi, compromising the pathogen’s defenses and enhancing its clearance. This mechanism not only offers a novel therapeutic approach for addressing drug-resistant typhoid fever but also has broader implications. It is highly anticipated that it could significantly broaden the application potential of rifampicin.27
Public Health Prevention and Control
Drug Resistance Monitoring Network Optimization
The emergence of blaNDM-5 positive Salmonella Typhi strains represents a significant threat to public health, underscoring the necessity for a comprehensive antimicrobial resistance monitoring network. First, it is essential to enhance the application of whole-genome sequencing technology for outbreak tracing.77 Genomic epidemiological studies enable the investigation of transmission pathways and evolutionary relationships among antibiotic-resistant strains. Additionally, it is essential to establish a comprehensive surveillance system that encompasses hospitals, communities, and the environment, with a particular focus on blaNDM-5 positive strains carrying plasmids such as InCX3.40,80 In addition, it is necessary to develop a web-based pathogen monitoring platform, such as the PathogenWatch system, to enable the rapid identification of antibiotic resistance genes and real-time comparison of public genomic data (Figure 7).68 Notably, blaNDM-5 positive bacterial strains have been identified in China,81 indicating the need to strengthen regional monitoring cooperation and establish a cross-regional information-sharing mechanism for antibiotic-resistant strains.
 |
Figure 7 The distribution of ciprofloxacin minimum inhibitory concentration (MIC) values, genotypic AMR predictions, and abundance differences of genetic mechanism combinations in Salmonella enterica var typhi isolates using PathogenWatch system are summarized. Reproduced with permission. Copyright 2021, Springer Nature. |
Infection Prevention and Control (IPC)
Comprehensive multi-layered measures should be adopted for the prevention and control of infections caused by blaNDM-5 positive Salmonella Typhi. In medical institutions, strict contact isolation measures should be implemented, particularly for patients from endemic areas, such as India and Pakistan.31,82 Given that this strain can be transmitted through the environment-animal-human pathway,65 it is necessary to enhance environmental disinfection in hospitals and manage medical waste. In terms of community prevention and control, it is essential to improve health infrastructure, particularly in areas where typhoid fever is endemic, to reduce the risk of fecal-oral transmission.83 International travelers are recommended to receive a typhoid vaccine before visiting popular areas and to enhance health monitoring after returning home.83 In addition, rapid diagnostic technologies, such as molecular detection methods, should be established to achieve early identification of the blaNDM-5 gene, providing a basis for timely preventive and control measures (Figure 8).40
 |
Figure 8 Both the IncX3 plasmid carrying blaNDM and the regulatory processes of T4SS-mediated conjugation (including hppX3-dependent repression in State 1 and actX/virBR-triggered activation in State 2) are summarized. Reproduced with permission Copyright 2025, Oxford Academic. |
Antibiotic Management (AMS)
In the face of blaNDM-5-mediated carbapenem resistance, stringent antibiotic management strategies must be implemented. Firstly, the clinical use of carbapenems should be restricted, reserving them as a last resort for the treatment of multidrug-resistant typhoid fever.84 Furthermore, it is recommended that precision medication plans be developed based on drug susceptibility results. For example, in the case of blaNDM-5 positive strains, a treatment regimen combining meropenem and polymyxin may be considered.84 In popular areas, it is essential to establish a monitoring system for antibiotic use, with a particular focus on currently effective drugs, such as azithromycin.31,85 In addition, it is essential to enhance the training of healthcare professionals to improve their ability to identify drug-resistant typhoid and standardize their awareness of medication use. Finally, the development of novel β-lactamase inhibitors should be encouraged to provide more options for clinical treatment.85 Notably, antibiotic management strategies should be combined with vaccination measures to form a comprehensive prevention and control system.83
Conclusion and Future Perspectives
Carbapenem-resistant Salmonella Typhi, particularly strains carrying the blaNDM-5 gene, represents an escalating global public health threat due to its high pathogenicity, extensively drug-resistant profile, and transregional transmission capacity. The ongoing spread is driven by horizontal gene transfer via epidemic plasmids such as IncX3 and IncHI2, often in conjunction with resistance to fluoroquinolones and cephalosporins, severely limiting treatment options. Although combination regimens like meropenem with polymyxin offer temporary efficacy, treatment failure and mortality remain high, underscoring the urgent need for novel therapeutics and robust antimicrobial stewardship. Sustainable control demands an integrated approach encompassing genomic surveillance, rapid diagnostics, and strengthened infection prevention. Crucially, global collaboration is indispensable to coordinate surveillance, share data, and accelerate the development of innovative interventions, thereby fortifying defenses against this emerging challenge in the post-antibiotic era.
Acknowledgments
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Disclosure
The authors declare that there is no conflict of interest.
References
1. Meiring JE, Shakya M, Khanam F, et al. Burden of enteric fever at three urban sites in Africa and Asia: a multicentre population-based study. Lancet Glob Health. 2021;9(12):e1688–16. doi:10.1016/s2214-109x(21)00370-3
2. World Health Organization. Typhoid vaccines: WHO position paper, March 2018 – recommendations. Vaccine. 2019;37(2):214–216. doi:10.1016/j.vaccine.2018.04.022
3. World Health Organization. WHO bacterial priority pathogens list, 2024. 2024.
4. Nizamuddin S, Khan EA, Chattaway MA, Godbole G. Case of carbapenem-resistant Salmonella Typhi infection, Pakistan, 2022. Emerg Infect Dis. 2023;29(11):2395–2397. doi:10.3201/eid2911.230499
5. Klemm EJ, Shakoor S, Page AJ, et al. Emergence of an extensively drug-resistant Salmonella enterica serovar Typhi clone harboring a promiscuous plasmid encoding resistance to fluoroquinolones and third-generation cephalosporins. Am Soc Microbiol. 2018;9(1):e00105–00118. doi:10.1128/mBio
6. Octavia S, Chew KL, Lin RTP, Teo JWP. Azithromycin-Resistant Salmonella enterica Serovar Typhi AcrB-R717Q/L, Singapore. Emerg Infect Dis. 2021;27(2):624–627. doi:10.3201/eid2702.203874
7. Walker J, Chaguza C, Grubaugh ND, et al. Assessing the global risk of typhoid outbreaks caused by extensively drug resistant Salmonella Typhi. Nat Commun. 2023;14(1):6502. doi:10.1038/s41467-023-42353-9
8. Azhar AB, Khalid A, Shah S. The implications of extensive drug-resistant typhoid fever: a case report. Cureus. 2019;11(6):e5032. doi:10.7759/cureus.5032
9. Vasanthaiah S, Takey P, Selvam PK, et al. Genomic perspectives on NDM Salmonella Typhi, and a case report from India. Infection. 2025;53(5):2053–2059. doi:10.1007/s15010-025-02546-4
10. Porwal R, Gopalakrishnan R, Rajesh N, Ramasubramanian V. Carbapenem resistant Gram-negative bacteremia in an Indian intensive care unit: a review of the clinical profile and treatment outcome of 50 patients. Indian J Crit Care Med. 2014;18(11):750–753. doi:10.4103/0972-5229.144021
11. Sah R, Donovan S, Seth-Smith HMB, et al. A novel lineage of ceftriaxone-resistant Salmonella Typhi from India that is closely related to XDR S. typhi found in Pakistan. Clin Infect Dis. 2019;71(5):1327–1330. doi:10.1093/cid/ciz1204/5681708
12. Chatham-Stephens K, Medalla F, Hughes M, et al. Emergence of extensively drug-resistant Salmonella Typhi infections among travelers to or from Pakistan - United States, 2016–2018. Morb Mortal Weekly Rep. 2019;68(1):11–13. doi:10.15585/mmwr.mm6801a3
13. Engsbro AL, Riis Jespersen HS, Goldschmidt MI, et al. Ceftriaxone-resistant Salmonella enterica serotype Typhi in a pregnant traveller returning from Karachi, Pakistan to Denmark, 2019. Eurosurveillance. 2019;24(21):1900289. doi:10.2807/1560-7917.Es.2019.24.21.1900289
14. Wu Y, Jiang T, Bao D, et al. Global population structure and genomic surveillance framework of carbapenem-resistant Salmonella enterica. Drug Resist Updates. 2023;68:100953. doi:10.1016/j.drup.2023.100953
15. Wang Z, He J, Li Q, et al. First detection of NDM-5-positive Salmonella enterica Serovar Typhimurium isolated from retail Pork in China. Microb Drug Resist. 2019;26(5):434–437. doi:10.1089/mdr.2019.0323
16. Gao Y, Wen J, Wang S, et al. Plasmid-encoded blaNDM-5 gene that confers high-level carbapenem resistance in Salmonella Typhimurium of Pork Origin. Infect Drug Resist. 2020;13:1485–1490. doi:10.2147/idr.S249357
17. Deng L, Lv L-C, Tu J, et al. Clonal spread of blaNDM-1-carrying Salmonella enterica serovar Typhimurium clone ST34 and wide spread of IncHI2/ST3-blaNDM-5 plasmid in China. J Antimicrob Chemother. 2024;79(8):1900–1909. doi:10.1093/jac/dkae178
18. Li X, Jiang Y, Wu K, et al. Whole-genome sequencing identification of a multidrug-resistant Salmonella enterica serovar Typhimurium strain carrying bla NDM-5 from Guangdong, China. Infect Genet Evol. 2017;55:195–198. doi:10.1016/j.meegid.2017.09.005
19. Fischer J, Schmoger S, Jahn S, Helmuth R, Guerra B. NDM-1 carbapenemase-producing Salmonella enterica subsp. enterica serovar Corvallis isolated from a wild bird in Germany. J Antimicrob Chemother. 2013;68(12):2954–2956. doi:10.1093/jac/dkt260
20. Borowiak M, Szabo I, Baumann B, et al. VIM-1-producing Salmonella Infantis isolated from swine and minced pork meat in Germany. J Antimicrob Chemother. 2017;72(7):2131–2133. doi:10.1093/jac/dkx101
21. Kanaan MHG, Khalil ZK, Khashan HT, Ghasemian A. Occurrence of virulence factors and carbapenemase genes in Salmonella enterica serovar Enteritidis isolated from chicken meat and egg samples in Iraq. BMC Microbiol. 2022;22(1):279. doi:10.1186/s12866-022-02696-7
22. Ke Y, Zhu Z, Lu W, et al. Emerging blaNDM-positive Salmonella enterica in Chinese pediatric infections. Microbiol Spectrum. 2024;12(12):e0148524. doi:10.1128/spectrum.01485-24
23. Huang F, Guo G, Feng L, Cai T, Huang X. Genomic insights into a clinical Salmonella Typhimurium isolate carrying plasmid-mediated blaNDM-5. J Global Antimicrob Resist. 2024;40:90–95. doi:10.1016/j.jgar.2024.11.009
24. Tan S, Li X, Lu B, et al. Genomic insights into the first emergence of blaNDM-5-carrying carbapenem-resistant Salmonella enterica Serovar London Strain in China. Infect Drug Resist. 2024;17:1781–1790. doi:10.2147/idr.S458625
25. Da Silva KE, Tanmoy AM, Pragasam AK, et al. The international and intercontinental spread and expansion of antimicrobial-resistant Salmonella Typhi: a genomic epidemiology study. Lancet Microbe. 2022;3(8):e567–e577. doi:10.1016/s2666-5247(22)00093-3
26. Zhou Y, Wu K, Lin H, et al. Whole genome sequencing of Salmonella in poultry from China reveals the presence of blaNDM-5 in different serotypes. Poult Sci. 2025;104(11):105647. doi:10.1016/j.psj.2025.105647
27. Lee GY, Song J. Rifampin mediated decapsulation and enhanced bacterial clearance: a new mechanism of rifamycins. eBioMedicine. 2025;117:105787. doi:10.1016/j.ebiom.2025.105787
28. Zeng S, Huang Y, Zhang X, Fu L, Sun Z, Li X. Molecular characterization of IncFII plasmid carrying bla NDM-5 in a Salmonella enterica serovar Typhimurium ST34 clinical isolate in China. mSphere. 2023;8(6):e0048023. doi:10.1128/msphere.00480-23
29. Li P, Yuan Y, Yan Y, et al. Characterization of blaNDM–5-carrying plasmids in two clinical Salmonella isolates from Jiaxing city, China. Front Microbiol. 2025;16:1620907. doi:10.3389/fmicb.2025.1620907
30. Wei J, Shen S, Zhang Q, et al. Emergence of a clinical Salmonella enterica serovar 1,4,[5], 12: i:-isolate, ST3606, in China with susceptibility decrease to ceftazidime-avibactam carrying a novel blaCTX-M-261 variant and a blaNDM-5. Eur J Clin Microbiol Infect Dis. 2024;43(5):829–840. doi:10.1007/s10096-024-04765-3
31. Carey ME, Thi Nguyen TN, Tran DHN, et al. The origins of haplotype 58 (H58) Salmonella enterica serovar Typhi. Commun Biol. 2024;7(1):775. doi:10.1038/s42003-024-06451-8
32. Shi Q, Hu H, Yu Q, et al. Chromosomal integration and plasmid fusion occurring in ST20 carbapenem-resistant Klebsiella pneumoniae isolates coharboring blaNDM-1 and blaIMP-4 induce resistance transmission and fitness variation. Emerging Microbes Infect. 2024;13(1):2339942. doi:10.1080/22221751.2024.2339942
33. De Pascale G, Cortegiani A, Rinaldi M, et al. Incidence of hospital-acquired infections due to carbapenem-resistant Enterobacterales and Pseudomonas aeruginosa in critically ill patients in Italy: a multicentre prospective cohort study. Critical Care. 2025;29(1):32. doi:10.1186/s13054-025-05266-1
34. Iovleva A, Mustapha MM, Griffith MP, et al. Carbapenem-resistant acinetobacter baumannii in U.S. Hospitals: diversification of circulating lineages and antimicrobial resistance. mBio. 2022;13(2):e0275921. doi:10.1128/mbio.02759-21
35. Carlos CC, Masim MAL, Lagrada ML, et al. Genome sequencing identifies previously unrecognized Klebsiella pneumoniae outbreaks in neonatal intensive care units in the Philippines. Clin Infect Dis. 2021;73(Supplement_4):S316–S324. doi:10.1093/cid/ciab776
36. Han D, Ma S, He C, Yang Y, Li P, Lu L. Unveiling the genetic architecture and transmission dynamics of a novel multidrug-resistant plasmid harboring blaNDM-5 in E. coli ST167: implications for antibiotic resistance management. BMC Microbiol. 2024;24(1):178. doi:10.1186/s12866-024-03333-1
37. Lv L-C, Lu -Y-Y, Gao X, et al. Characterization of NDM-5-producing Enterobacteriaceae isolates from retail grass carp (Ctenopharyngodon idella) and evidence of bla NDM-5-bearing IncHI2 plasmid transfer between ducks and fish. Zool Res. 2022;43(2):255–264. doi:10.24272/j.issn.2095-8137.2021.426
38. Shi Y, Yang Y, Song Y, Zhu Y, Zhao G, Tang B. Characterisation of blaNDM-5-bearing IncHI2 plasmid from Escherichia fergusonii in China. J Global Antimicrob Resist. 2025;42:222–228. doi:10.1016/j.jgar.2025.03.008
39. Tian X, Zhang L, Li C, Xia D, Ying J. Genome sequence of a sequence type 1 NDM-5-producing carbapenem-resistant Klebsiella pneumoniae in China. J Global Antimicrob Resist. 2024;38:271–274. doi:10.1016/j.jgar.2024.05.001
40. Gao Y, Xie N, Ma T, et al. VirBR counter-silences HppX3 to promote conjugation of blaNDM-IncX3 plasmids. Nucleic Acids Res. 2025;53(5):gkaf182. doi:10.1093/nar/gkaf182
41. Simner PJ, Mostafa HH, Bergman Y, et al. Progressive development of cefiderocol resistance in Escherichia coli during therapy is associated with an increase in blaNDM-5 copy number and gene expression. Clin Infect Dis. 2022;75(1):47–54. doi:10.1093/cid/ciab888/6383178
42. Bano S, Khatoon A, Quareshi U, Ul-Haq Z, Karim A. Pan-genome analysis and drug repurposing strategies for extensively drug-resistant Salmonella Typhi: subtractive genomics and e-pharmacophore approaches. Int J Biol Macromol. 2025;291:139003. doi:10.1016/j.ijbiomac.2024.139003
43. Su LH, Wu TL, Chiu CH. Development of carbapenem resistance during therapy for non-typhoid Salmonella infection. Clin Microbiol Infect. 2012;18(4):E91–94. doi:10.1111/j.1469-0691.2012.03767.x
44. Fernández J, Guerra B, Rodicio M. Resistance to carbapenems in non-typhoidal Salmonella enterica serovars from humans, animals and food. Vet Sci. 2018;5(2):40. doi:10.3390/vetsci5020040
45. Prah I, Ayibieke A, Mahazu S, et al. Emergence of oxacillinase-181 carbapenemase-producing diarrheagenic Escherichia coli in Ghana. Emerging Microbes Infect. 2021;10(1):865–873. doi:10.1080/22221751.2021.1920342
46. Acman M, Wang R, van Dorp L, et al. Role of mobile genetic elements in the global dissemination of the carbapenem resistance gene blaNDM. Nat Commun. 2022;13(1):1131. doi:10.1038/s41467-022-28819-2
47. Humphrey S, Fillol-Salom A, Quiles-Puchalt N, et al. Bacterial chromosomal mobility via lateral transduction exceeds that of classical mobile genetic elements. Nat Commun. 2021;12(1):6509. doi:10.1038/s41467-021-26004-5
48. Pedersen T, Sekyere JO, Govinden U, Moodley K, Sivertsen A. Spread of plasmid-encoded NDM-1 and GES-5 carbapenemases among extensively drug-resistant and pandrug-resistant clinical enterobacteriaceae in Durban, South Africa. Antimicrob Agents Chemother. 2018;62(5):10–128. doi:10.1128/AAC.02178-17
49. Kong L-H, Xiang R, Wang Y-L, et al. Integration of the blaNDM-1 carbapenemase gene into a novel SXT/R391 integrative and conjugative element in Proteus vulgaris. J Antimicrob Chemother. 2020;75(6):1439–1442. doi:10.1093/jac/dkaa068
50. Sattler J, Noster J, Stelzer Y, et al. OXA-48-like carbapenemases in Proteus mirabili– novel genetic environments and a challenge for detection. Emerging Microbes Infect. 2024;13(1):2353310. doi:10.1080/22221751.2024.2353310
51. Veeraraghavan B, Pragasam AK, Bakthavatchalam YD, Ralph R. Typhoid fever: issues in laboratory detection, treatment options & concerns in management in developing countries. Future Sci OA. 2018;4(6):FSO312. doi:10.4155/fsoa-2018-0003
52. Qamar FN, Yousafzai MT, Khalid M, et al. Outbreak investigation of ceftriaxone-resistant Salmonella enterica serotype Typhi and its risk factors among the general population in Hyderabad, Pakistan: a matched case-control study. Lancet Infect Dis. 2018;18(12):1368–1376. doi:10.1016/s1473-3099(18)30483-3
53. Wang W, Liu F, Peng Z, Li F, Ma A. Complete genome sequence of Salmonella enterica subsp. enterica Serovar Indiana C629, a carbapenem-resistant bacterium isolated from chicken carcass in China. Genome Announc. 2016;4(4):e00662–00616. doi:10.1128/genomeA.00662-16
54. Yang C, Chen K, Ye L, Heng H, Chan EWC, Chen S. Genetic and drug susceptibility profiles of mcr-1-bearing foodborne Salmonella strains collected in Shenzhen, China during the period 2014–2017. Microbiol Res. 2022;265:127211. doi:10.1016/j.micres.2022.127211
55. Harada S, Aoki K, Nomura Y, et al. Transmission of global clones of NDM-producing Enterobacterales and interspecies spread of IncX3 plasmid harbouring blaNDM-5 in Tokyo. J Global Antimicrob Resist. 2024;38:309–316. doi:10.1016/j.jgar.2024.06.020
56. Yang C, Han J, Berglund B, et al. Dissemination of blaNDM-5 and mcr-8.1 in carbapenem-resistant Klebsiella pneumoniae and Klebsiella quasipneumoniae in an animal breeding area in Eastern China. Front Microbiol. 2022;13:1030490. doi:10.3389/fmicb.2022.1030490
57. Huang R, Gao W, Sun Y, et al. Genomic characterization of carbapenemase-producing Klebsiella pneumoniae ST895 isolates from canine origins through whole-genome sequencing analysis. Microorganisms. 2025;13(2):332. doi:10.3390/microorganisms13020332
58. Yao S, Yu J, Zhang T, et al. Comprehensive analysis of distribution characteristics and horizontal gene transfer elements of blaNDM-1-carrying bacteria. Sci Total Environ. 2024;946:173907. doi:10.1016/j.scitotenv.2024.173907
59. Tang C, Shen S, Yang W, et al. Dynamic evolution of ceftazidime-avibactam resistance from a single patient through the IncX3_NDM-5 plasmid transfer and blaKPC mutation. Int J Antimicrob Agents. 2024;64(2):107228. doi:10.1016/j.ijantimicag.2024.107228
60. Gomez SA, Martino F, Sanz MB, et al. Plasmid diversity in Klebsiella pneumoniae ST307 co-producing KPC plus NDM recovered during the COVID-19 pandemic. Infect Genet Evol. 2025;133:105795. doi:10.1016/j.meegid.2025.105795
61. Yin D, Lin Y, Li Z, et al. Characterization of a novel NDM-5-harboring plasmid from a carbapenem-resistant Escherichia coli isolate from China. Infect Drug Resist. 2020;13:3929–3935. doi:10.2147/idr.S277997
62. Kiffer CRV, Rezende TFT, Costa-Nobre DT, et al. A 7-year Brazilian national perspective on plasmid-mediated carbapenem resistance in enterobacterales, pseudomonas aeruginosa, and acinetobacter baumannii complex and the impact of the coronavirus disease 2019 pandemic on their occurrence. Clin Infect Dis. 2023;77(Supplement_1):S29–S37. doi:10.1093/cid/ciad260
63. Sheng H, Suo J, Wang X, et al. Global prevalence and transmission of the mcr-9 in Salmonella: a genomic study with insights from Salmonella enterica serovar Thompson isolated from poultry food in China. Food Res Int. 2025;202:115763. doi:10.1016/j.foodres.2025.115763
64. Abe R, Akeda Y, Sakamoto N, et al. A nationwide plasmidome surveillance in Thailand reveals a limited variety of new Delhi metallo-β-lactamase-producing carbapenem-resistant clones and spreading plasmids. J Clin Microbiol. 2022;60(12):e0108022. doi:10.1128/jcm.01080-22
65. Zhao Q, Berglund B, Zou H, et al. Dissemination of blaNDM-5 via IncX3 plasmids in carbapenem-resistant Enterobacteriaceae among humans and in the environment in an intensive vegetable cultivation area in eastern China. Environ Pollut. 2021;273:116370. doi:10.1016/j.envpol.2020.116370
66. Dyson ZA, Ashton PM, Khanam F, et al. Pathogen diversity and antimicrobial resistance transmission of Salmonella enterica serovars Typhi and Paratyphi A in Bangladesh, Nepal, and Malawi: a genomic epidemiological study. Lancet Microbe. 2024;5(8):100841. doi:10.1016/s2666-5247(24)00047-8
67. Jiang Y, Jiang M, Lu X, et al. Genomic epidemiology and antimicrobial resistance reveal local transmission dynamics of enteric fever in Shenzhen, one of the mega cities in China. J Infect. 2025;90(4):106469. doi:10.1016/j.jinf.2025.106469
68. Argimón S, Yeats CA, Goater RJ, et al. A global resource for genomic predictions of antimicrobial resistance and surveillance of Salmonella Typhi at pathogenwatch. Nat Commun. 2021;12(1):2879. doi:10.1038/s41467-021-23091-2
69. Teo JQM, Tang CY, Lim JC, et al. Genomic characterization of carbapenem-non-susceptible Pseudomonas aeruginosa in Singapore. Emerging Microbes Infect. 2021;10(1):1706–1716. doi:10.1080/22221751.2021.1968318
70. Salamzade R, Manson AL, Walker BJ, et al. Inter-species geographic signatures for tracing horizontal gene transfer and long-term persistence of carbapenem resistance. Genome Med. 2022;14(1):37. doi:10.1186/s13073-022-01040-y
71. Posen HJ, Wong W, Farrar DS, et al. Travel-associated extensively drug-resistant typhoid fever: a case series to inform management in non-endemic regions. J Travel Med. 2023;30(1):taac086. doi:10.1093/jtm/taac086
72. Kariuki S, Dyson ZA, Mbae C, et al. Multiple introductions of multidrug-resistant typhoid associated with acute infection and asymptomatic carriage, Kenya. eLife. 2021;10:e67852. doi:10.7554/eLife.67852
73. Wain J, Hendriksen RS, Mikoleit ML, Keddy KH, Ochiai RL. Typhoid fever. Lancet. 2015;385(9973):1136–1145. doi:10.1016/s0140-6736(13)62708-7
74. Petrin S, Tiengo A, Longo A, et al. Uncommon Salmonella infantis variants with incomplete antigenic formula in the poultry food chain, Italy. Emerg Infect Dis. 2024;30(4):795–799. doi:10.3201/eid3004.231074
75. Mularoni A, Cona A, Campanella M, et al. Donor-derived carbapenem-resistant gram-negative bacterial infections in solid organ transplant recipients: active surveillance enhances recipient safety. Am J Transplantation. 2024;24(6):1046–1056. doi:10.1016/j.ajt.2024.02.005
76. Hoşbul T, Aydoğan CN, Kaya S, et al. Salmonella ve Shigella Klinik İzolatlarında Azitromisin Minimum İnhibitör Konsantrasyon Değerlerinin ve Karbapenem Direncinin Araştırılması. Mikrobiyol Bul. 2021;55(4):480–491. doi:10.5578/mb.20219702
77. Wagenlehner FM, Gasink LB, McGovern PC, et al. Cefepime–taniborbactam in complicated urinary tract infection. New Engl J Med. 2024;390(20):1937–1939. doi:10.1056/NEJMc2403590
78. Ding L, Yang Z, Sun B. Understanding blaNDM-1 gene regulation in CRKP infections: toward novel antimicrobial strategies for hospital-acquired pneumonia. Mol Med. 2024;30(1):29. doi:10.1186/s10020-024-00794-y
79. Arshad R, Arshad MS, Malik A, et al. Mannosylated preactivated hyaluronic acid-based nanostructures for bacterial infection treatment. Int J Biol Macromol. 2023;242(Pt 1):124741. doi:10.1016/j.ijbiomac.2023.124741
80. Heo Y-A. Imipenem/Cilastatin/Relebactam: a Review in Gram-Negative Bacterial Infections. Drugs. 2021;81(3):377–388. doi:10.1007/s40265-021-01471-8
81. Manesh A, Meltzer E, Jin C, et al. Typhoid and paratyphoid fever: a clinical seminar. J Travel Med. 2021;28(3):taab012. doi:10.1093/jtm/taab012
82. Ray STJ, Fuller CE, Ahmadu A, et al. Aetiologies, neuroradiological features, and risk factors for mortality and long-term neurosequelae of febrile coma in Malawian children: a prospective cohort study. Lancet Glob Health. 2025;13(6):e1057–e1071. doi:10.1016/s2214-109x(25)00054-3
83. Thilliez G, Mashe T, Chaibva BV, et al. Population structure of Salmonella enterica Typhi in Harare, Zimbabwe (2012–19) before typhoid conjugate vaccine roll-out: a genomic epidemiology study. Lancet Microbe. 2023;4(12):e1005–e1014. doi:10.1016/s2666-5247(23)00214-8
84. Bai Y, Liu W, Liu Z, et al. Multi-omics analysis of glycolytic reprogramming and ROS dynamics in host-specific responses to Salmonella Typhi infection in mice. J Adv Res. 2025;S2090-1232(25):00345. doi:10.1016/j.jare.2025.05.027
85. Carey ME, Jain R, Yousuf M, et al. Spontaneous emergence of azithromycin resistance in independent lineages of Salmonella Typhi in Northern India. Clin Infect Dis. 2021;72(5):e120–e127. doi:10.1093/cid/ciaa1773
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.