Back to Journals » Infection and Drug Resistance » Volume 11
Comparative analysis of the pathogenicity between multidrug-resistant Acinetobacter baumannii clinical isolates: isolation of highly pathogenic multidrug-resistant A. baumannii and experimental therapeutics with fourth-generation cephalosporin cefozopran
Received 21 February 2018
Accepted for publication 24 May 2018
Published 10 October 2018 Volume 2018:11 Pages 1715—1722
DOI https://doi.org/10.2147/IDR.S166154
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Joachim Wink
Satoshi Nishida, Yasuo Ono
Department of Microbiology and Immunology, Teikyo University School of Medicine, Itabashi, Tokyo, Japan
Introduction: The pathogenicity of fatal-outbreak Acinetobacter baumannii isolates has not been fully investigated. This study aimed to compare the pathogenicity between A. baumannii clinical isolates, including multidrug-resistant A. baumannii (MDRA).
Materials and methods: Antibiotic susceptibility was determined by the broth microdilution method, and drug-resistant genes were characterized by PCR and sequencing. The pathogenicity of A. baumannii and antibiotic responses were evaluated using the Galleria mellonella infection model. Clinical isolates from an A. baumannii outbreak at our hospital were categorized using the pulse-field gel electrophoresis. Of the 16 isolated A. baumannii clones, 12 clones were resistant to carbapenems (meropenem and imipenem), of which 10 clones were also resistant to amikacin and ciprofloxacin (MDRAs). MDRAs had OXA-51-like β-lactamase gene harboring an insertion sequence in the promoter region and armA gene encoding 16S rRNA methyltransferase.
Results: Carbapenem- and/or amikacin-resistant A. baumannii were more pathogenic than carbapenem- and/or amikacin-sensitive A. baumannii in G. mellonella. MDRA isolate TK1033 was more virulent than other A. baumannii isolates. However, TK1033 was sensitive to the fourth-generation cephalosporin cefozopran in addition to minocycline, tigecycline, and polymyxins (colistin and polymyxins B) in vitro and in vivo in the MDRA-G. mellonella infection model.
Conclusion: Differences in pathogenicity among carbapenem-resistant A. baumannii clones are consistent with heterogeneous clinical outcomes. Strain TK1033, isolated frequently during the outbreak, was the most virulent, whereas preoutbreak isolate TK1032 was less virulent than other A. baumannii isolates. Infection by high-virulence isolates may be more prevalent during outbreaks. These strains may prove valuable for investigating MDRA virulence and novel therapeutics.
Keywords: Acinetobacter baumannii, amikacin, aminoglycoside, carbapenem, cephalosporin, cefozopran, virulence, Galleria mellonella, multidrug resistance
Introduction
The Enterococcus feacium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter sp. (ESKAPE) pathogens frequently acquire drug resistance. Nosocomial infections from multidrug-resistant (MDR) pathogens often have poor outcomes, especially in intensive care units. A. baumannii is a Gram-negative pathogen of opportunistic infection, and MDR A. baumannii (MDRA) strains are often resistant to major antibiotics such as amikacin (AMK), ciprofloxacin (CIP), and imipenem (IPM).1,2
A fatal MDRA outbreak occurred in 2010 at Teikyo University Hospital (Tokyo, Japan).3,4 In this study, we compared the pathogenicity of A. baumannii clinical isolates including MDRA strains. Despite many clinical studies, little is known regarding the virulence and microbial fitness of A. baumannii. The greater wax moth Galleria mellonella may be useful for the prospective evaluation of microbial pathogens in randomized controlled studies5 as infectivity and antibiotic responses show strong concordance with mammalian infection models,6 particularly for A. baumannii.5,7 Furthermore, this in vivo system has obvious ethical and logistical advantages over mammalian infection models. The objectives of this study were to assess the virulence and antibiotic response profiles of carbapenem-resistant A. baumannii (CRAB) isolates from this fatal outbreak using the G. mellonella host model.
Materials and methods
Ethics
The Teikyo University Animal Ethics Committee exempted this study.
Materials
Ampicillin (AMP) and kanamycin (KAN) were purchased from Meiji Seika Pharma (Tokyo, Japan). Cefepime (FEP) was purchased from Bristol-Myers Squibb (New York, NY, USA). Colistin (CST), minocycline (MIN), polymyxin B (PMB), and tetracycline (TET) were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). AMK and tigecycline (TGC) were purchased from Wako Pure Chemical Industries (Osaka, Japan). Sitafloxacin (STX) was purchased from Cimic CMO (Tokyo, Japan). Cefozopran (ZOP), doxycycline (DOX), and IPM were generously provided by Takeda Pharmaceutical (Osaka, Japan), Pfizer (New York, NY, USA), and Banyu Pharmaceutical (Tokyo, Japan), respectively. CIP, sparfloxacin (SPX), and meropenem (MEM) were generously provided by Sumitomo Dainippon Pharma (Osaka, Japan). Lysogeny broth (LB; Miller), LB ager (Miller), and Muller Hinton Broth II (MHB II) were purchased from Becton Dickinson (Franklin Lakes, NJ, USA). Cation-adjusted MHB II was prepared by adding CaCl2 and MgCl2 (Wako Pure Chemical Industries) at the final concentrations of 50 mg/L Ca2+ and 25 mg/L Mg2+ to autoclaved MHB II. Saline was prepared as 0.9% NaCl (Wako Pure Chemical Industries).
Isolation and characterization of A. baumannii
A. baumannii clinical isolates were previously collected from Teikyo University Hospital, a 1,082-bed academic hospital in Tokyo, Japan,4 and stored in glycerol at -80°C. During the MDRA outbreak from 2009 to 2010, 59 patients were infected and nine patients who had hematological diseases died as a result of the infection. A. baumannii strains were originally isolated from blood, sputum, stool, urine, bronchoalveolar lavage, pus, and pharyngeal mucus specimens. All clinical isolates of A. baumannii were identified at the species level by the rpoB sequence.8 Carbapenem resistance was confirmed by the broth microdilution method. Antibiotic susceptibility was interpreted according to Clinical and Laboratory Standards Institute (CLSI) guidelines (document M100-S22). Each isolate exhibited a unique pattern on pulse-field gel electrophoresis (PFGE) (Kawakami, in preparation, 2018). Frozen stocks of A. baumannii were streaked on LB agar and incubated at 37°C. A. baumannii isolates were grown at 37°C in LB for liquid culture in Bioshaker BR-40LF (Taitec, Tokyo, Japan).
Gene amplification and DNA sequencing
A genomic fragment was amplified by PCR using KOD FX Neo (Toyobo, Osaka, Japan). RNA polymerase β-subunit gene rpoB was amplified according to a previously described protocol.8 DNA sequences were determined by direct sequencing using a BigDye Terminator v3.1 Cycle Sequencing Kit and ABI PRISM 3100 Genetic Analyzer (Thermo Fisher Scientific, Waltham, MA, USA). Sequences were compared using NCBI BLASTN 2.7.1. Presence of the blaOXA-51-like gene was confirmed by PCR with primers OXA-51A (5′-CTAATAATTGATCTACTCAAG-3′) and OXA-51B (5′-CCAGTGGATGGATGGATAGATTATC-3′), while presence of the armA gene was determined using primes armA-F (5′-AGGTTGTTTCCATTTCTGAG-3′) and armA-R (5′-ATCTCTTCCATTCCCTTCTCC-3′).9,10
Antimicrobial activity
The minimum inhibitory concentrations (MICs, in milligram per liter) were determined by the broth microdilution technique according to CLSI guidelines (document M7-A9). The MIC was defined as the lowest antimicrobial concentration of a serial twofold dilution (0.25–256 mg/L) in cation-adjusted MHB II conferring no growth (<2 mm button or no definite turbidity) of A. baumannii on 96-well plates (BMBio, Tokyo, Japan).
Infection model
G. mellonella larvae (250–350 mg) were purchased from Live Bait Factory (Osaka, Japan). Each isolate was diluted in saline (0.9% NaCl) and 10 µL was injected into the rear right proleg of 10 randomly selected G. mellonella larvae using an insulin syringe (Terumo, Tokyo, Japan). Saline (0.9% NaCl) and injection-free groups (both n=10) were used to evaluate the effects of trauma and attrition, respectively. G. mellonella larvae were incubated at 37°C. Death was defined as unresponsiveness to touch.7 Survival curves were plotted using Kaleidagraph 4.1.4 (Synergy Software, Reading, PA, USA), and median lethal dose (LD50) values were determined by curve fitting at 24h.11
For evaluating antibacterial activity against MDRA, 10 µL of antimicrobial agents were injected into the rear left proleg of 10 G. mellonella larva infected with MDRA TK1033. Saline was used as a control of agents. Antimicrobial doses were 40 mg/kg each.
Statistical analysis
Statistical analysis was performed using Microsoft Excel 2007 for Windows (Redmond, WA, USA) and Excel Statistics 2008 (Social Survey Research Information, Tokyo, Japan). Survival plots were generated by the Kaplan-Meier method and compared among antibiotics by the log-rank test. Differences between log CFU values were tested for significance by one-way ANOVA followed by Fisher’s least significant difference tests for pair-wise comparisons. A P-value of <0.05 was considered to be statistically significant. All tests were two-tailed.
Results
Pathogenicity of A. baumannii clinical isolates in G. mellonella
Sixteen unique pulse-field gel electrophoresis (PFGE) isolates were tested in the G. mellonella infection model. Colony-forming units were used to confirm the injected inocula. Injection of A. baumannii isolates resulted in time-dependent and dose-dependent larval death, but LD50 values varied markedly (Table 1). We then compared the LD50 values among drug-sensitive, MEM/CIP-resistant, CIP/AMK-resistant, and MEM/CIP/AMK-resistant (MDRA) isolates (Figure 1). The LD50 values of drug-sensitive isolates were comparable to double-resistant isolates MEM/CIP (P=0.59) and CIP/AMK (P=0.13). In contrast, the LD50 values of MDR isolates (MEM/CIP/AMK) were significantly lower (higher virulence) than those of drug-sensitive isolates (P=0.010) and double-resistant (MEM/CIP) isolates (P=0.0027). The LD50 values of CIP/AMK-resistant isolates were significantly lower (higher virulence) than those of MEM/CIP-resistant isolates (P=0.046). However, the LD50 values of MEM/CIP/AMK-resistant isolates were comparable to double-resistant (CIP/AMK) isolates (P=0.21). These data suggest that AMK resistance is a critical determinant of A. baumannii virulence. Similarly, the LD50 values of the 10 MDRA isolates were comparable to those of the four double-resistant CIP/aminoglycoside (AMK or KAN) isolates (P=0.10).
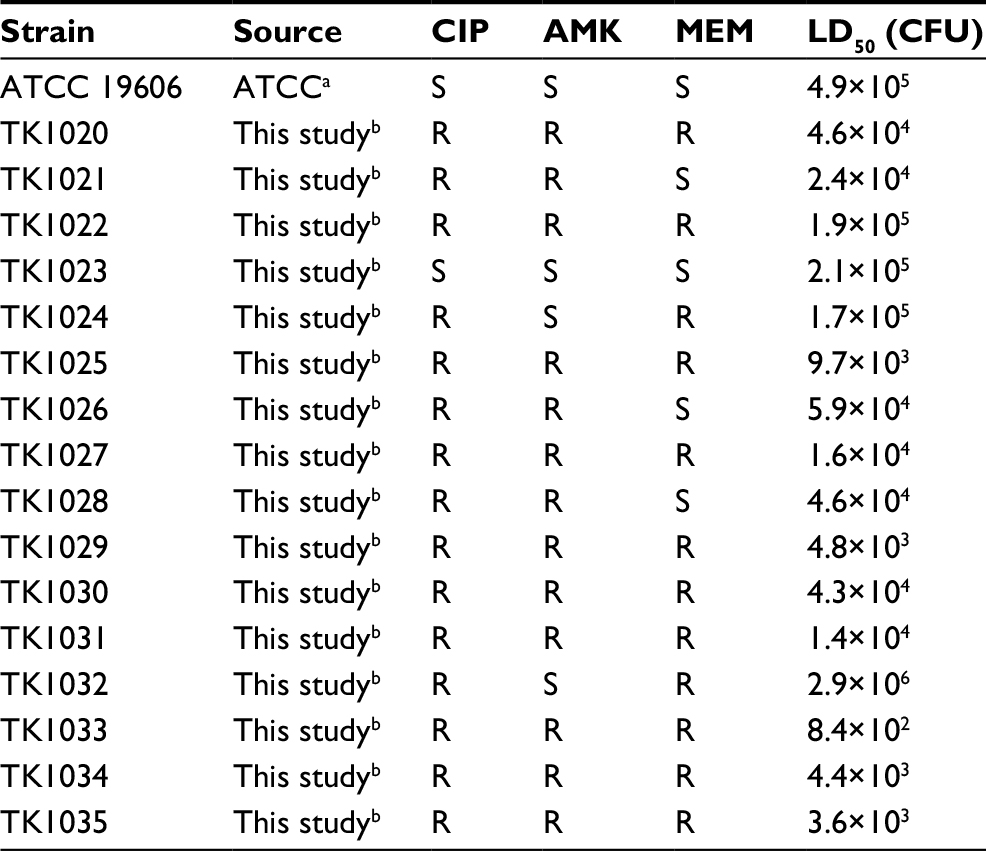 | Table 1 Summary of Acinetobacter baumannii clinical isolates Notes: R, AMK, CIP, and MEM MICs ≥32, 4, and 16 mg/L, respectively. aIsolated from urine. bIsolated from blood, sputum, stool, urine, bronchoalveolar lavage, pus, and pharyngeal mucus specimens. Abbreviations: AMK, amikacin; ATCC, American Type Culture Collection; CFU, colony-forming units; CIP, ciprofloxacin; LD50, median lethal dose; MEM, meropenem; MICs, minimum inhibitory concentrations; R, resistant, S, sensitive. |
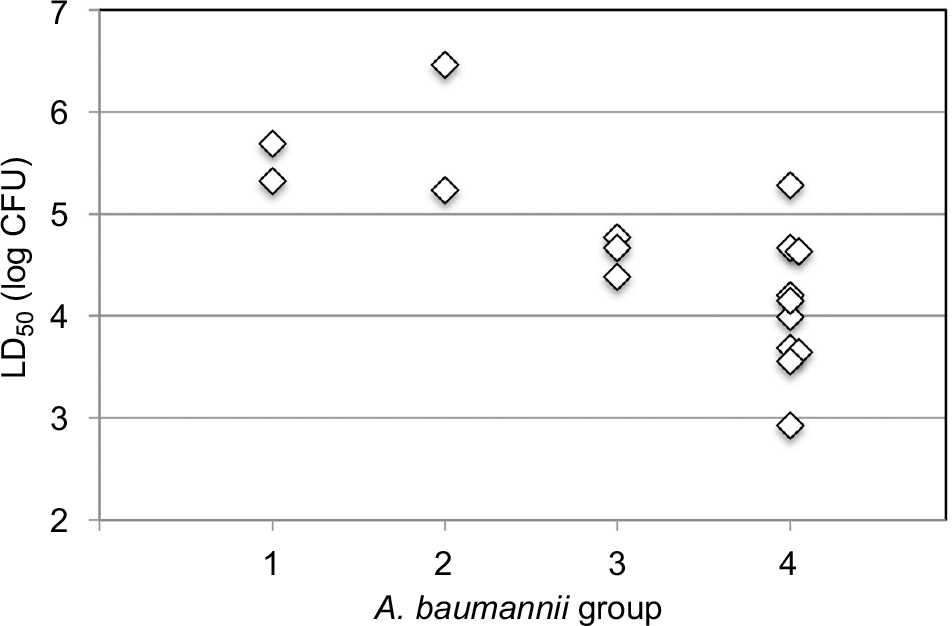 | Figure 1 LD50 values of different A. baumannii strains in G. mellonella. Notes: The LD50 values expressed in log CFU are plotted for four A. baumannii groups: (1) drug sensitive, (2) MEM/CIP resistant, (3) CIP/AMK resistant, and (4) MEM/CIP/AMK resistant (MDR). MDR isolates (MDRA) induced significantly greater mortality than the drug-sensitive group and the MEM/CIP-resistant isolates group (P=0.010 and 0.0027, respectively). Abbreviations: A. baumannii, Acinetobacter baumannii; AMK, amikacin; CIP, ciprofloxacin; G. mellonella, Galleria mellonella; LD50, median lethal dose; MEM, meropenem; MDR, multidrug resistant; MDRA, MDR A. baumannii. |
Next, we compared the survival of G. mellonella infected with AMK-sensitive and -resistant isolates. At 24 h postinoculation, the LD50 values of the 13 AMK-resistant isolates were lower than those of the four AMK-sensitive isolates (Figure 2A). Furthermore, G. mellonella larvae were killed more rapidly by the 13 AMK-resistant isolates than the four AMK-sensitive isolates (P=2.5×10−7). For instance, after 2 days, 62% of the G. mellonella larvae inoculated with AMK-sensitive isolates survived, compared to only 20% inoculated with AMK-resistant isolates (Figure 2A). The presence of the armA gene coding 16S rRNA methyltransferase was confirmed in all AMK-resistant isolates by PCR, suggesting that armA-positive isolates are generally more virulent against G. mellonella larvae than armA-negative isolates (except for the AMK-sensitive, KAN-resistant and armA-positive isolate TK1024).
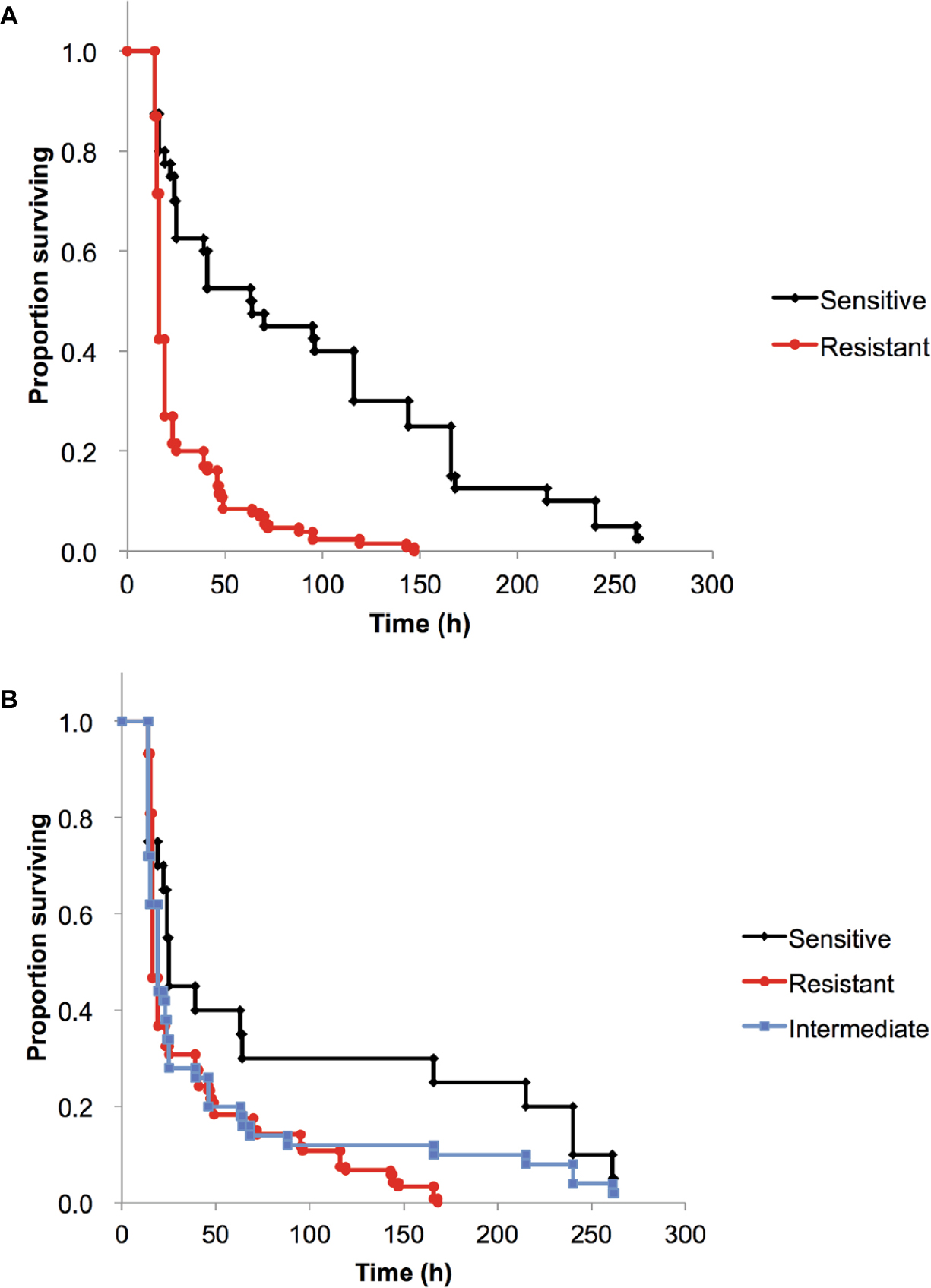 | Figure 2 Kaplan–Meier cumulative survival curves of G. mellonella infected with A. baumannii. Notes: G. mellonella larvae were infected with 1×105 CFU of each strain. Survival curves for the three drug-resistant groups were compared using the log-rank test. (A) AMK-resistant A. baumannii (MIC ≥32 mg/L) induced significantly greater mortality than AMK-sensitive A. baumannii (P=2.5×10−7). (B) CRAB (MEM MIC ≥32 mg/L) induced significantly greater mortality than CSAB (MEM MIC ≤2 mg/L) (P=0.011) and comparable mortality than A. baumannii of intermediate carbapenem resistance (MEM MIC 4–16 mg/L) (P=0.47). Abbreviations: AMK, amikacin; A. baumannii, Acinetobacter baumannii; CFU, colony-forming units; CRAB, carbapenem-resistant A. baumannii; CSAB, carbapenem-sensitive A. baumannii; G. mellonella, Galleria mellonella; MEM, meropenem; MIC, minimum inhibitory concentration. |
CRAB isolates also induced greater mortality of G. mellonella than carbapenem-sensitive A. baumannii (CSAB) isolates (P=0.011, Figure 2B). The isolates with intermediate carbapenem resistance showed virulence comparable to CRAB (P=0.47) and CSAB (P=0.13). In our study, G. mellonella larvae were killed more rapidly by blaOXA-23-negative isolates of A. baumannii than blaOXA-23-positive isolate TK1032 (Table 1). These results suggest that clinical isolates harboring the blaOXA-23 gene and those involved in an outbreak may be less virulent.
Antibiotic sensitivity of highly virulent MDRA
Next, we evaluated the antibiotic sensitivity of the MDRA isolate TK1033 (Table 2). TK1033 was resistant to aminoglycosides such as KAN and AMK. TK1033 was also resistant to the TET antibiotics TET and DOX but sensitive to the TET antibiotics MIN and TGC. Similarly, TK1033 was sensitive to the fourth-generation quinolone antibiotic STX but resistant to the second-generation quinolone CIP and the third-generation quinolone SPX. TK1033 was also sensitive to the polymyxins PMB and CST. TK1033 was resistant to the fourth-generation cephalosporin FEP but sensitive to another fourth-generation cephalosporin, ZOP.
 | Table 2 MIC values (mg/L) of the Acinetobacter baumannii clinical isolates in this study Abbreviations: AMK, amikacin; AMP, ampicillin; CIP, ciprofloxacin; CST, colistin; DOX, doxycycline; FEP, cefepime; IPM, imipenem; KAN, kanamycin; MEM, meropenem; MIC, minimum inhibitory concentration; MIN, minocycline; PMB, polymyxin B; SPX, sparfloxacin; STX, sitafloxacin; TET, tetracycline; TGC, tigecycline; ZOP, cefozopran. |
Antibacterial treatment of MDRA with the fourth-generation cephalosporin ZOP
It has previously been shown that appropriate antimicrobials can successfully treat G. mellonella infected with A. baumannii.7 To determine whether the G. mellonella-MDRA infection model can be used to study the in vivo activity of antibacterial agents against our clinical isolates, we investigated the effects of single-dose (40 mg/kg) ZOP, CST, PMB, MIN, TGC, and STX on MDRA-infected G. mellonella larvae (Figure 3). Initially, we infected G. mellonella larvae with the MDRA strain TK1033. Treatment with antibiotics that had in vitro activity against this A. baumannii strain significantly prolonged the survival of G. mellonella larvae. Thus, the G. mellonella-MDRA infection model can be used as an alternative to mammalian systems for the assessment of antibacterial agent efficacy.
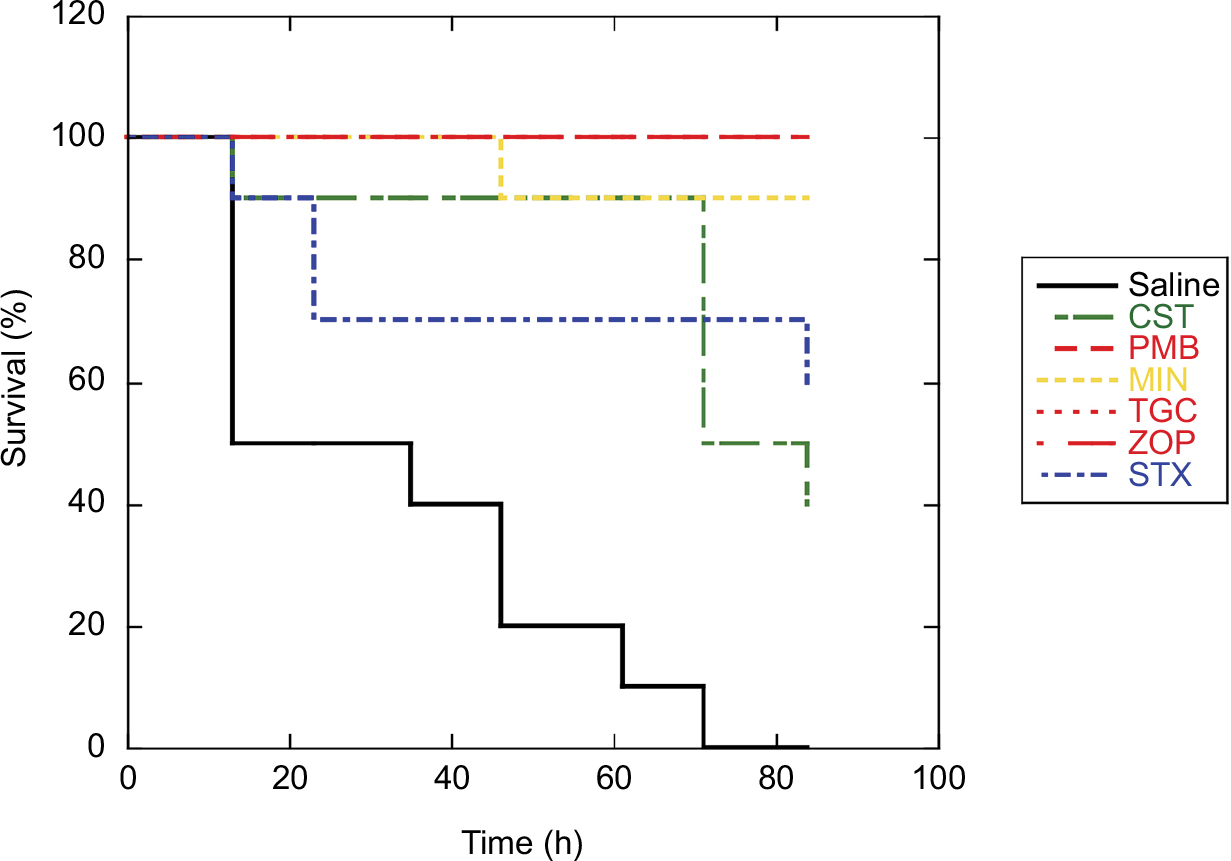 | Figure 3 Antibacterial activities against MDRA TK1033. Notes: After infection with MDRA TK1033 (5×103 CFU), ZOP, STX, MIN, TGC, PMB, and CST (40 mg/kg each), antibiotics to which the strain was susceptible, significantly prolonged the survival of infected G. mellonella larvae (P<0.01 compared to saline-treated control infected larvae). Abbreviations: CFU, colony-forming units; CST, colistin; G. mellonella, Galleria mellonella; MDRA, multidrug-resistant Acinetobacter baumannii; MIN, minocycline; PMB, polymyxin B; STX, sitafloxacin; TGC, tigecycline; ZOP, cefozopran. |
Discussion
The pathogenicity of A. baumannii clinical strains has been studied using G. mellonella. The US military clinical isolate AB5075 exhibited a 100-fold lower LD50 in the G. mellonella infection model than strain American Type Culture Collection (ATCC) 19606.12 Another clinical strain, ATCC 17978, exhibited a twofold lower LD50 than ATCC 19606. In contrast, the epidemiological outbreak strains AYE and ACICU presented about twofold higher LD50 values than ATCC 17978.13 Therefore, A. baumannii strains show a broad range of virulence values and resistance profiles in vitro that do not always correspond with clinical outcome. Thus, the clinical relevance of different strain characteristics in epidemiological outbreaks is uncertain. In this study, we selected clinical isolates previously classified at our institution to analyze the relevance of individual resistance spectra to clinical virulence. MDR (MEM/CIP/AMK) isolates of A. baumannii appeared to be more virulent than double-resistant (MEM/CIP and CIP/AMK) isolates (Figure 1). In particular, TK1033 exhibited a 500-fold lower LD50 in the G. mellonella infection model than strain ATCC 19606 (Table 1). Isolates with the same PFGE pattern as the highly virulent strain TK1033 were most frequently isolated during the outbreak, suggesting clinically significant infectivity and pathogenicity to humans. However, extrapolation of these results to human virulence will require additional study using a mouse infection model. Our previous report has suggested that the lipopolysaccharide (LPS) subtype of the outbreak strain is also a possible virulence factor. Indeed, the LPS from TK1033 induced greater expression of proinflammatory cytokine genes in human polymorphonuclear leukocytes than that from a drug-sensitive A. baumannii in vitro.14 Some studies have reported hypervirulent strains in epidemic outbreak isolates and have analyzed the pathogenicity in animal infection models.12,15–17 The horizontal clonal spread of hypervirulent strains may contribute to epidemic outbreaks and poor clinical outcomes. Further study is warranted to elucidate virulence in animal models and clinical outcomes.
Recently, Lemos et al (2014)18 conducted a systematic review and meta-analysis of carbapenem resistance and mortality in patients with A. baumannii infection. CRAB-infected patients showed significantly higher mortality risk than CSAB-infected patients, although there was a substantial variation in outcome. A case-control study of mortality following CRAB bacteraemia suggested a clonal subgroup of A. baumannii as a predictor of mortality.19 Therefore, differences in virulence among CRAB clones may partly contribute to the heterogeneous results in clinical studies. The results of the present study appear consistent with those of previous clinical studies, suggesting that carriage of carbapenem-resistance genes is associated with organism virulence, although with some variance (Figure 2B). While these studies demonstrated increased mortality among patients with CRAB vs CSAB bloodstream infections on bivariate inspection, this does not exclude the possibility that patient comorbidities drive poor outcomes or that patients with more comorbidities are more likely to be infected by CRAB.
In our study, G. mellonella larvae were killed more rapidly by blaOXA-23-negative isolates of A. baumannii than blaOXA-23-positive isolate of A. baumannii. These results suggest that clinical isolates harboring the blaOXA-23 gene and those involved in an outbreak may be less virulent, consistent with a previous study regarding clinical isolates harboring the blaOXA-40 gene and those involved in an outbreak.5 However, another study reported a highly virulent isolate, AB5075, harboring blaOXA-23 and involved in an outbreak.12 Hence, the impact of carbapenem resistance on mortality due to A. baumannii infection remains uncertain.
In contrast to blaOXA-23-positive isolate, AMK-resistant isolates of A. baumannii killed G. mellonella larvae more rapidly than AMK-sensitive isolates (Figure 2A). These results suggest that clinical isolates harboring the armA gene coding 16S rRNA methyltransferase and those involved in an outbreak may be more virulent. The impact of AMK resistance and/or the armA gene on mortality due to A. baumannii infection warrants further study in light of this possibility.
In this study, we utilized G. mellonella to compare the pathogenicity of A. baumannii clinical isolates and to assess the efficacy of antibiotic treatment against a highly pathogenic MDRA. Very little is known about the pathogenicity of MDRA, so this infection model may serve as a useful tool for future research. Interestingly, we observed an association between the pathogenicity of MDRA in G. mellonella and that observed in humans. Therefore, this model may also facilitate the study of novel compounds against MDRA with the ethical and logistical advantages over mammalian models. Mammalian infection models should be used to confirm our results before human studies in the future.
The third-generation cephalosporins are widely used for the empiric therapy of serious nosocomial infections. However, expression of β-lactamase (eg, AmpC) may confer resistance. Fourth-generation cephalosporins have potent activity against Gram-negative bacteria including those producing AmpC β-lactamase.20,21 ZOP is a fourth-generation cephalosporin approved for various bacterial infections in Japan.22 The highly virulent MDRA TK1033 isolate was more sensitive to ZOP than FEP (Table 2). TK1033 was also sensitive to MIN and TGC, both of which are used for empiric therapy of MDRA infection.23–27 Intravenous MIN is particularly promising due to its improved safety profile.25–27 ZOP and/or MIN were used for empiric therapy during the MDRA outbreak in our hospital. Antibacterial activities of ZOP and MIN in vitro and in vivo were confirmed in this study (Table 2 and Figure 3). Re-evaluation and use of fourth-generation cephalosporins and MIN may help reduce the use of carbapenems, resulting in reduced carbapenem-resistance for antimicrobial stewardship.
Acknowledgments
This work was supported by the Japan Society for the Promotion of Science (JSPS) (KAKENHI 24591490 and 17K10032 to YO and 18K10030 to SN). We thank members in the Department of Microbiology and Immunology, and Private University Research Branding Project (MEXT) at Teikyo University School of Medicine for helpful discussion and technical assistance.
Disclosure
The authors report no conflicts of interest in this work.
References
Munoz-Price LS, Weinstein RA. Acinetobacter infection. N Engl J Med. 2008;358(12):1271–1281. | ||
Peleg AY, Seifert H, Paterson DL. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev. 2008;21(3):538–582. | ||
Yuji K, Oiso G, Matsumura T, Murashige N, Kami M. Police investigation into multidrug-resistant Acinetobacter baumannii outbreak in Japan. Clin Infect Dis. 2011;52(3):422. | ||
Alshahni MM, Asahara M, Kawakami S, et al. Genotyping of Acinetobacter baumannii strains isolated at a Japanese hospital over five years using targeted next-generation sequencing. J Infect Chemother. 2015;21(7):512–515. | ||
Esterly JS, Mclaughlin MM, Malczynski M, Qi C, Zembower TR, Scheetz MH. Pathogenicity of clinical Acinetobacter baumannii isolates in a Galleria mellonella host model according to blaOXA-40 gene and epidemiological outbreak status. Antimicrob Agents Chemother. 2014;58(2):1240–1242. | ||
Jander G, Rahme LG, Ausubel FM. Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J Bacteriol. 2000;182(13):3843–3845. | ||
Peleg AY, Jara S, Monga D, Eliopoulos GM, Moellering RC, Mylonakis E. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrob Agents Chemother. 2009;53(6):2605–2609. | ||
La Scola B, Gundi VA, Khamis A, Raoult D. Sequencing of the rpoB gene and flanking spacers for molecular identification of Acinetobacter species. J Clin Microbiol. 2006;44(3):827–832. | ||
Héritier C, Poirel L, Fournier PE, Claverie JM, Raoult D, Nordmann P. Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49(10):4174-4179. | ||
Yamane K, Wachino J, Doi Y, Kurokawa H, Arakawa Y. Global spread of multiple aminoglycoside resistance genes. Emerg Infect Dis. 2005;11(6):951–953. | ||
Nishida S, Ishii M, Nishiyama Y, Abe S, Ono Y, Sekimizu K. Lactobacillus paraplantarum 11-1 isolated from rice bran pickles activated innate immunity and improved survival in a silkworm bacterial infection model. Front Microbiol. 2017;8:436. | ||
Jacobs AC, Thompson MG, Black CC, et al. AB5075, a highly virulent isolate of Acinetobacter baumannii, as a model strain for the evaluation of pathogenesis and antimicrobial treatments. MBio. 2014;5(3):e01076–14. | ||
Antunes LC, Imperi F, Carattoli A, Visca P. Deciphering the multifactorial nature of Acinetobacter baumannii pathogenicity. PLoS One. 2011;6(8):e22674. | ||
Ubagai T, Nakano R, Nakano A, Kamoshida G, Ono Y. Gene expression analysis in human polymorphonuclear leukocytes stimulated by LPSs from nosocomial opportunistic pathogens. Innate Immun. 2015;21(8):802–812. | ||
Harris G, Kuo Lee R, Lam CK, et al. A mouse model of Acinetobacter baumannii-associated pneumonia using a clinically isolated hypervirulent strain. Antimicrob Agents Chemother. 2013;57(8):3601–3613. | ||
Bruhn KW, Pantapalangkoor P, Nielsen T, et al. Host fate is rapidly determined by innate effector-microbial interactions during Acinetobacter baumannii bacteremia. J Infect Dis. 2015;211(8):1296–1305. | ||
Jones CL, Clancy M, Honnold C, et al. Fatal outbreak of an emerging clone of extensively drug-resistant Acinetobacter baumannii with enhanced virulence. Clin Infect Dis. 2015;61(2):145–154. | ||
Lemos EV, de La Hoz FP, Einarson TR, et al. Carbapenem resistance and mortality in patients with Acinetobacter baumannii infection: systematic review and meta-analysis. Clin Microbiol Infect. 2014;20(5):416–423. | ||
Nutman A, Glick R, Temkin E, et al. A case-control study to identify predictors of 14-day mortality following carbapenem-resistant Acinetobacter baumannii bacteraemia. Clin Microbiol Infect. 2014;20(12): O1028–O1034. | ||
Bryskier A. New concepts in the field of cephalosporins: C-3’ quaternary ammonium cephems (Group IV). Clin Microbiol Infect. 1997;3:S1–S6. | ||
Garau J, Wilson W, Wood M, Carlet J. Fourth-generation cephalosporins: a review of in vitro activity, pharmacokinetics, pharmacodynamics and clinical utility. Clin Microbiol Infect. 1997;3:S87–S101. | ||
Iwahi T, Okonogi K, Yamazaki T, et al. In vitro and in vivo activities of SCE-2787, a new parenteral cephalosporin with a broad antibacterial spectrum. Antimicrob Agents Chemother. 1992;36(7):1358–1366. | ||
Karageorgopoulos DE, Kelesidis T, Kelesidis I, Falagas ME. Tigecycline for the treatment of multidrug-resistant (including carbapenem-resistant) Acinetobacter infections: a review of the scientific evidence. J Antimicrob Chemother. 2008;62(1):45–55. | ||
Gordon NC, Wareham DW. A review of clinical and microbiological outcomes following treatment of infections involving multidrug-resistant Acinetobacter baumannii with tigecycline. J Antimicrob Chemother. 2009;63(4):775–780. | ||
Ritchie DJ, Garavaglia-Wilson A. A review of intravenous minocycline for treatment of multidrug-resistant Acinetobacter infections. Clin Infect Dis. 2014;59(suppl 6):S374–S380. | ||
Colton B, Mcconeghy KW, Schreckenberger PC, Danziger LH. I.V. minocycline revisited for infections caused by multidrug-resistant organisms. Am J Health Syst Pharm. 2016;73(5):279–285. | ||
Lashinsky JN, Henig O, Pogue JM, Kaye KS. Minocycline for the treatment of multidrug and extensively drug-resistant A. baumannii: a review. Infect Dis Ther. 2017;6(2):199–211. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.