Back to Journals » Infection and Drug Resistance » Volume 12
Community-associated Staphylococcus aureus PVL+ ST22 predominates in skin and soft tissue infections in Beijing, China
Authors Xiao N ![]() , Yang J, Duan N, Lu B
, Yang J, Duan N, Lu B ![]() , Wang L
, Wang L
Received 15 April 2019
Accepted for publication 25 July 2019
Published 12 August 2019 Volume 2019:12 Pages 2495—2503
DOI https://doi.org/10.2147/IDR.S212358
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Nan Xiao,1,* Jianghui Yang,2,* Ning Duan,1 Binghuai Lu,3 Lijun Wang4
1Department of Laboratory Medicine, Beijing Tsinghua Changgung Hospital, Tsinghua University, Beijing 102218, People’s Republic of China; 2Department of Pathology, Beijing Tsinghua Changgung Hospital, Tsinghua University, Beijing 102218, People’s Republic of China; 3Laboratory of Clinical Microbiology and Infectious Diseases, Department of Pulmonary and Critical Care Medicine, China-Japan Friendship Hospital; National Clinical Research Center of Respiratory Diseases, Beijing 100029, People’s Republic of China; 4Clinical Research Center, Beijing Tsinghua Changgung Hospital, School of Clinical Medicine, Tsinghua University, Beijing 102218, People’s Republic of China
*These authors contributed equally to this work
Purpose: Community-associated Staphylococcus aureus (CA S. aureus) is the most common causative pathogen of the skin and soft tissue infections (SSTIs). This study aims to determine clonal distribution, virulence factors of CA S. aureus clinical isolates from purulent SSTIs in Beijing, China.
Materials and methods: CA-S. aureus isolates were collected from 115 outpatients with purulent SSTIs from the department of dermatology from April 2015 to April 2017. Multilocus sequence typing and Staphylococcus cassette chromosome mec typing were performed to explore molecular characteristics. Phylogenetic analysis of 16S rRNA of dominant S. aureus isolates was performed using MEGA-X software. Virulence genes were detected by PCR, while biofilm formation was evaluated by a microtiter plate method. The antimicrobial susceptibility was tested by an automatic VITEK system.
Results: Forty-four CA-S. aureus isolates identified from SSTIs contain 9 methicillin-resistant S. aureus (MRSA) isolates (20.4%) and 35 methicillin-susceptible S. aureus isolates (MSSA) (79.6%). The dominant sequence types (STs) were ST22 (40.9%) and clonal complex 59 (CC59; 77.8%) in Community-associated methicillin resistant methicillin-resistant S. aureus. 27.8% of ST22 isolates were homologous to the epidemic ST22 EMRSA-15 in Europe. The prevalence of virulence genes lukS/lukF, tst-1, etA, edinA, icaA, and icaD was 50%, 93.2%, 4.5%, 4.5%, 100%, and 100%, respectively. All CC59 isolates exhibited stronger biofilm-forming capability than ST22 clones. Among the MSSA subgroup, the poor biofilm producers had significantly higher sensitivity to sulfamethoxazole/Trimethoprim.
Conclusion: The dominant epidemic clone PVL+ ST22 MSSA containing tst-1 occurs in Beijing, indicating that a PVL+ ST398 clone which was previously predominant in this district had been replaced by a new clone.
Keywords: community-associated Staphylococcus aureus, skin and soft tissue infection, biofilm formation, MLST-genotyping
Introduction
Skin and soft tissue infections (SSTIs) are common in both the outpatient and inpatient settings. Staphylococcus aureus is the most common causative pathogen of the SSTIs,1 particularly in purulent infections such as furuncles, carbuncles, cutaneous abscesses, and impetigo.
Community-associated methicillin-resistant S. aureus (CA-MRSA) has emerged as an important pathogen worldwide. The prevalence of CA-MRSA in SSTIs varies from region to region, 2.6% in China and 23% in the Middle East.2,3 In addition, the global distribution of CA-MRSA clones is heterogeneous and often characterized by a regional predominant clonal lineage.4 ST121 was previously the most common community-associated Staphylococcus aureus (CA-S. aureus) clone in SSTIs in China.2 The ST398 was then reported as the predominant clone in Jiangsu Province and Beijing, China.5,6 Since the dominant CA-MRSA clones are likely to have evolved from locally circulating community-associated methicillin-sensitive S. aureus (CA-MSSA) genotypes that are strongly associated with SSTIs,7,8 it is important to investigate the CA-S. aureus clonal structure within SSTIs around the local district.
Furthermore, virulence factors are highly associated with the pathogenesis of S. aureus invasive infections such as toxic shock syndrome and staphylococcal scalded skin syndrome (SSSS). Panton–Valentine leukocidin (PVL), for instance, is a key virulence factor. Previous studies have shown the association between PVL and severe invasive infections.9,10 Thus, it is important to characterize the virulence factors of CA-S. aureus for improving therapeutic approaches.
This study is focused on investigating purulent SSTIs cases in Beijing and characterizing S. aureus by multilocus sequence typing (MLST) types, SCCmec types, and key virulence factors. To our knowledge, it is the first study to report PVL+ ST22 CA-MSSA, an epidemic lineage in Europe,11 as a predominant clone in China.
Methods
Staphylococcus aureus isolates
From April 2015 to April 2017, pathogen detections were performed for all outpatients with purulent SSTIs in the department of dermatology in a hospital located in Beijing. The bacterial identification was performed by Matrix-Assisted Laser Desorption/Ionization Time of Flight Mass Spectrometry. All S. aureus clinical isolates were maintained with tryptic soy broth (TSB) at −70°C. And they were revived by streaking onto Columbia blood agar and cultured at 35°C overnight.
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing of S. aureus isolates was performed with a commercial VITEK Compact II system. Antibiotics susceptibility profiles were determined according to Clinical Laboratory Standards Institute Performance Standards of Antimicrobial Susceptible Testing (CLSI document M100 27th edition).
MLST and SCCmec genotype
MLST analysis of S. aureus isolates was performed according to previously described procedures.12 In the MLST database (https://pubmlst.org/saureus/), the sequences of seven housekeeping genes (arcC, aroE, glpF, gmk, pta, tpi, and yqi) were compared to existing sequences by the assignment of allelic numbers. The SCCmec genotype was also identified as previously described.13
Phylogenetic analysis
ST22 clinical isolates were compared with two representatives of epidemic ST22 S. aureus strains, H-EMRSA-15 (NZ_CP007659.1:538355-539911) and HO 5096 0412 (HE681097.1:474448-476002), to construct phylogenetic tree for 16S rRNA gene using MEGA X software.14 16S rRNA sequencing of clinical S. aureus isolates was carried out by a commercial company (Beijing Ruibiotech) using a primer pair, 27F 5-AGAGTTTGATCCTGGCTCAG and 1492R 5-TACGGCTACCTTGTTACGACTT.
Detection of virulence genes
All S. aureus isolates were screened for toxic shock syndrome toxin 1 gene (tst-1), Panton–Valentine leukocidin genes (lukS/lukF), three types of Staphylococcal exfoliative toxin (ET) genes (etA, etB, etD), epidermal cell differentiation inhibitor genes (edinA and edinB), and intracellular adhesion molecule genes (icaA and icaD) by PCR with appropriate primers as previously described.15–22
Biofilm formation assay
Biofilm formation contributes to the pathogenesis of S. aureus. Quantification of biofilms was performed using a modified Microtiter plate method.23 Briefly, the bacterial isolates were cultured in TSB and incubated at 37ºC overnight. The cultures were diluted with fresh TSB to 106 CFU/mL. Two hundred microliters of the diluted solution was added to sterile 96-well plates and incubated at 37ºC for 24 hrs. The negative control wells contained TSB alone. After 24-hr incubation, broth was removed and the wells were gently washed three times with phosphate-buffered saline. After the wells were dried at room temperature, they were stained with 0.1% crystal violet. Then, the crystal violet bound to the biofilms in the wells was dissolved by 30% acetic acid. The optical density (OD) was measured at 570 nm by BioTek Synergy H1 reader. Each assay was performed in triplicate wells and was repeated three times. The cutoff of OD (ODc) was defined as three standard deviations above the mean OD of the negative control. The ability of biofilm formation was classified as follows: OD≤ODc, negative; ODc<OD≤2xODc, weak; 2xODc<OD≤4xODc, moderate; and 4xODc<OD, strong.
Statistical analysis
The correlation between the antibiotic resistance and biofilm formation was evaluated by the Pearson Chi-Square test. P-values less than 0.05 were regarded as significant. The statistic software used in our study was MedCalc 15.2.2 (Seoul, Korea).
Results
Staphylococcus aureus isolates
One hundred fifteen purulent SSTIs outpatients were enrolled in this study from April 2015 to April 2017. S. aureus was the most common pathogen, accounting for 38.3% (44/115). A total of 44 non-duplicate S. aureus isolates were recovered from abscess.
Molecular characteristics of CA-S. aureus
Of 44 CA S. aureus strains, MSSA accounted for 79.6% (n=35), and MRSA for 20.4% (n=9). All CA-MRSA harbored SCCmec Ⅳ or Ⅴ. Eleven distinct sequence types (STs) were identified among the 44 isolates (Table 1), among which the most prevalent ST22 accounted for 40.9% (n=18). Furthermore, the predominant ST among MSSA was ST22 (17/35, 48.6%), followed by ST398 (5/35, 14.3%), ST188 (4/35, 11.4%), ST121 (2/35, 5.7%), and ST5 (2/35, 5.7%) respectively. Meanwhile, the most prevalent ST within MRSA was the clonal complex 59 (CC59) including ST59 (5/9, 55.6%) and ST338 (2/9, 22.2%). Other clones were isolated sporadically.
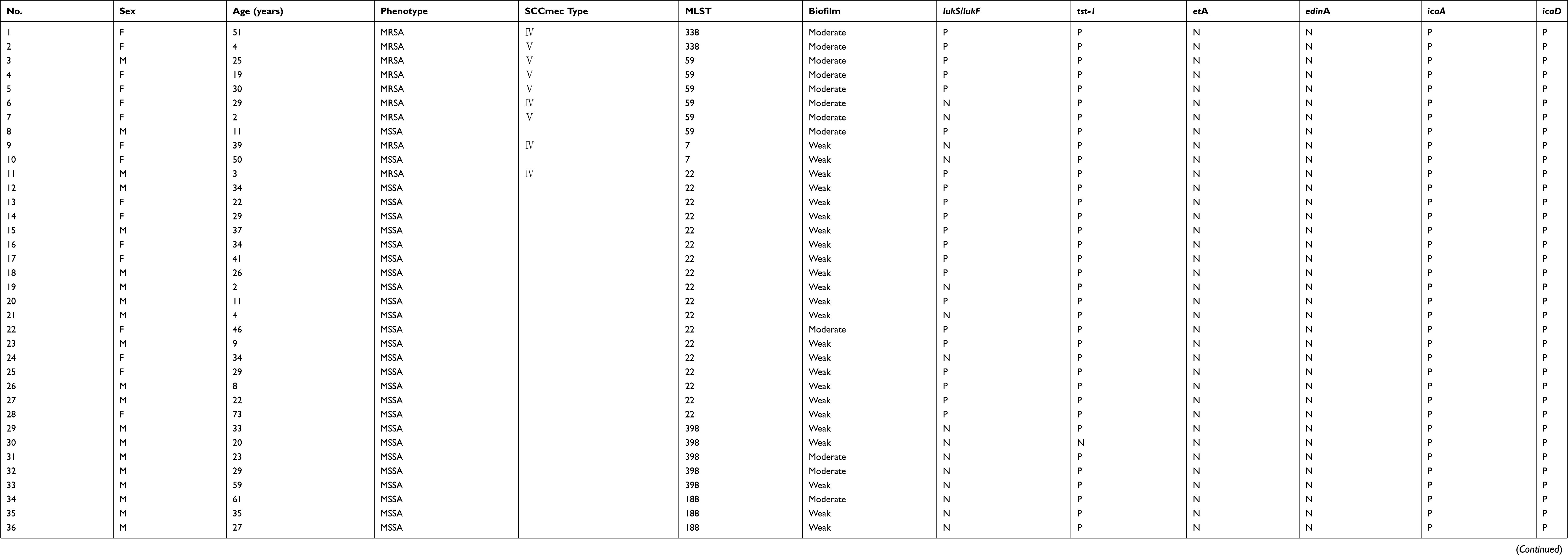 |
Table 1 MLST typing, biofilm formation, and virulence factors genes of 44 S. aureus isolations from SSTI patients |
Phylogenetic tree
Neighbor-Joining tree for 16S rRNA gene of predominant ST22 S. aureus showed two clades (Figure 1). Five out of 18 (27.8%) ST22 isolates exhibit the same evolutionary origins as the epidemic ST22 EMRSA-15 (HO 5096 0412 and H-EMRSA-15) in Europe.
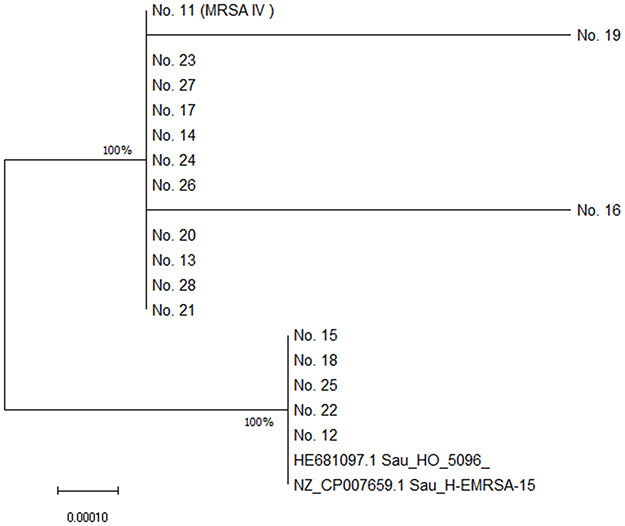 |
Figure 1 Neighbor-Joining tree of ST22 S. aureus isolates. The tree was rooted by the representative reference genomes of ST22 EMRSA-15 (HO 5096 0412 and H-EMRSA-15). The optimal tree with the sum of branch length =0.00212286 is shown. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Evolutionary analyses were conducted in MEGA X. |
Virulence genes
The prevalence of PVL positive S. aureus was 50% (22/44), and there was no significant difference (p=0.45) between MRSA (66.7%, 6/9) and MSSA (45.7%, 16/35). 83.3% (15/18) of ST22 was detected positive for lukS/lukF. All ST398, ST188, ST121, and ST5 isolates did not carry the lukS/lukF genes. Forty-one isolates, except two ST121 and one ST398, carried the tst-1 gene accounting for 93.2% (38/41). Only ST121 isolates carried etA and edinA, accounting for 4.5%. However, none were found positive for etB, etD, and edinB. All of the isolates possessed two genes icaA and icaD coding for critical intracellular adhesion molecules.
Biofilm formation
The results showed that 74.3% (26/35) of isolates were weak biofilm producers, and 25.7% (9/35) MSSA isolates were moderate biofilm producers, while 77.8% (7/9) MRSA were moderate biofilm producers (Table 2). All CC59 isolates, including ST59 and ST338, exhibit moderate biofilm formation ability, irrespective of methicillin susceptibility.
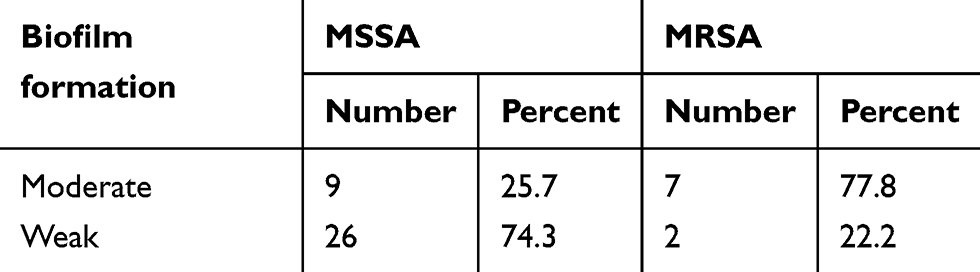 |
Table 2 Biofilm formation ability of MSSA and MRSA |
Comparison of susceptibility profiles of different biofilm producers
Antibiotics sensitivity was different between weak and moderate biofilm producers among MSSA subgroup. Within the MSSA subgroup, the sensitivity to benzylpenicillin, gentamycin, ciprofloxacin, levofloxacin, and erythromycin of weak biofilm-producing isolates were higher than that of moderate biofilm producers; however, the differences were not statistically significant. The only significant difference of antibiotic sensitivity was observed in sulfamethoxazole/trimethoprim (92.3% vs 55.6%, p=0.03) (Table 3). Although MRSA subgroup consisted of only nine isolates, the majority of them are moderate biofilm producers. All MRSA and MSSA isolates were sensitive to vancomycin, rifampicin, linezolid, moxifloxacin, and tigecycline.
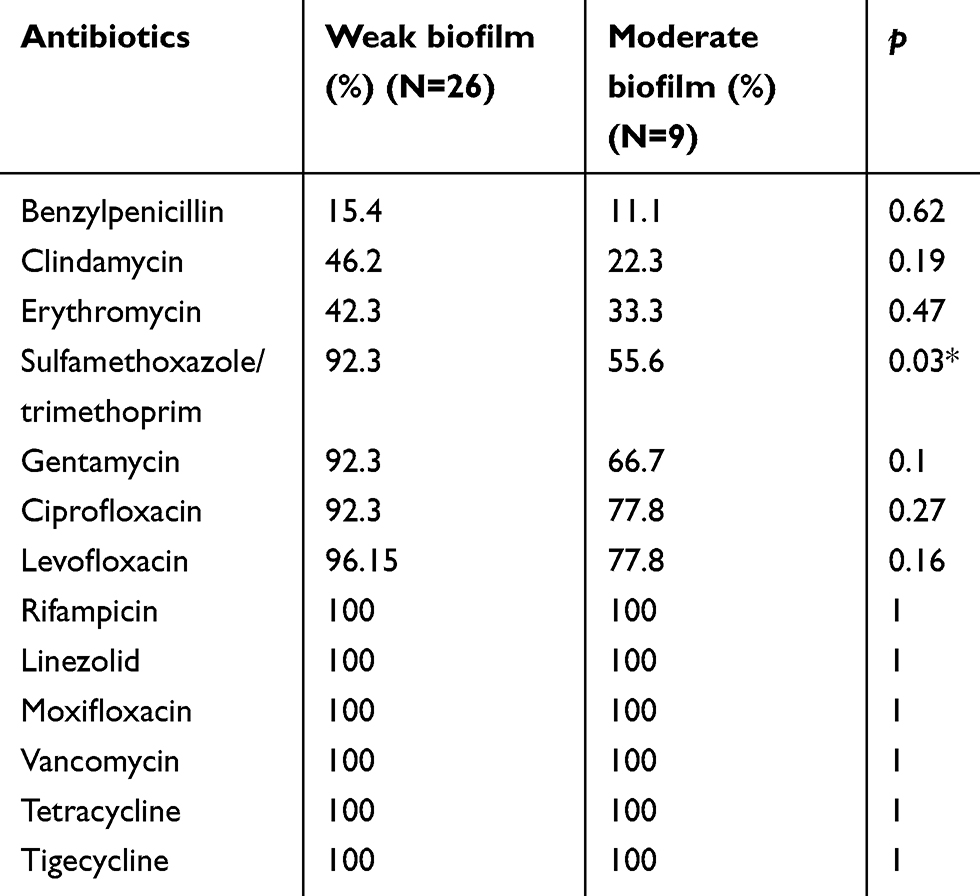 |
Table 3 The sensitivities of antibiotics between weak biofilm and moderate biofilm producers among MSSA subgroup |
Discussion
In the present research, we determined the clonal distribution, virulence factors, and biofilm formation ability of S. aureus clinical isolates from purulent SSTIs. Several findings were reported. Firstly, an endemic PVL+ ST22 clone similar to EMRSA-15 appeared in this region. Secondly, CA-MRSA accounted for 20.4% of SSTIs. The local circulating MRSA clone belongs to CC59, and all CC59 isolates have stronger biofilm formation ability compared to ST22. Finally, the majority of isolates harbored tst-1 and lukS/lukF genes.
It is worth noting that we screened a dominant clone PVL+ ST22 CA-MSSA circulating in this Beijing community, whereas previous study6 suggested the PVL+ ST398 was the most dominant clone among CA-MSSA with SSTIs around Beijing. It is likely that ST22 has replaced ST398 as the dominant lineage in Beijing. This trend deserves attention from the physicians for the following reasons. Firstly, ST22 clone has a strong ability to spread. ST22-IVh (EMRSA-15 clone) is a pandemic lineage originated from Europe, which spread rapidly through hospitals following introduction into Singapore, replacing the endemic ST239 population.24,25 Recent studies reported several outbreaks of ST22 CA-MSSA or CA-MRSA.11,26,27 In our study, the phylogeny analysis indicated local PVL+ ST22 CA-MSSA is similar to epidemic EMRSA-15, which could serve as a reservoir for CA-MRSA. Secondly, ST22 clone is highly virulent. DeLencastre et al,28 document that ST22-IV effectively invade cells in vitro and is highly pathogenic in vivo compared with ST228-I. In addition, a long-term familial infection cluster was caused by a novel PVL-positive ST22 CA-MRSA in Japan.29
The spread of CA-MRSA has become a serious problem worldwide. Our data showed the prevalence of CA-MRSA was 20.4%, which is much higher than previously reported ~4% in China.6 The prevalence of CA-MRSA was found dramatically increasing in a majority of regions.2,5,6,30 The trend would pose a serious challenge to local infection control.
As far as the clonal distribution of CA-MRSA is concerned, five CA-MRSA clones, ST1, ST30, ST80, ST59, and ST8, account for the vast majority of CA-MRSA infections worldwide.31 Different clonal lineages spread in specific regions, for instance, ST8 in Germany32 and ST59 in Taiwan.33 Several surveillance reports suggested that the predominant clone would shift across regions. A multicenter study in China between 2009 and 2011 revealed that the most prevalent sequence type was ST121 (19/51, 37.3%) among CA-MRSA with SSTIs,2 however, ST59 was found to be the most prevalent ST in China since 2012.34–36 We also found the circulating clone CC59 accounted for 77.8% CA-MRSA. Indeed, ST59 CA-MRSA has become a persistent pandemic clone causing invasive infections around China now.36
PVL is a key virulence factor of S. aureus, mainly associated with necrotic lesions in the skin or mucosa.18 The prevalence of lukS/lukF ranged from over 20% to approximately 80% of S. aureus isolates.6,30,34,37,38 We detected 50% of S. aureus harboring lukS/lukF. Particularly, 83.3% of ST22 isolates were PVL positive. Chen39 reported that ST22 tends to harbor a lukS/lukF gene compared to other ST types in China. Surprisingly, Gu5 detected no lukS/lukF gene among CA-MRSA with SSTIs in Jiangsu Province, China. So far, it is unknown whether PVL+ CA-MRSA clones arose through acquisition of the mec element from strains with a PVL+ MSSA or conversely, through acquisition of PVL phage by strains with a methicillin resistance background. Overall, we are concerned whether the PVL+ ST22 CA-MSSA clone would arise to become the dominant CA-MRSA clone around the city. Further studies are needed to uncover the possible evolution of the dominant PVL+ ST22 CA-MSSA over time in China.
TSST-1-producing S. aureus are more likely to cause complicated infections. In this study, we found the tst-1 gene was detected among 93.2% CA-S.aureus isolates. The overall prevalence of tst-1 carrying S. aureus in Iran was 21.3%, ranging from 0% to 68%.40 Moreover, a recent survey in China41 reported that only 3.9% MRSA strains harbored the tst-1 gene. Concerning the tst-1 gene, the prevalence is much higher in our study, so we presume high virulence isolates circulate around this community in Beijing.
ETs are the sole virulence factors responsible for SSSS, a disease mostly affecting neonates and children under 5 years of age. In Italy, etA positive ST5 clone S. aureus had ever caused an outbreak of skin infections in neonates.42 We found two ST121 clone strains contain both the etA gene and the edinA gene associated with epidermal hyperplasia. Yet, the etA gene positive S. aureus strains were rarely reported in China.34 Continuous monitoring ETs positive S. aureus in this community may be of interest.
Biofilm formation is another primary virulence factor of S. aureus.43 Different strains possess different adhesins and differ in their ability to produce biofilms. In this study, the majority of ST22 clones had poor biofilm formation ability, whereas all CC59 strains can produce robust biofilms. CA-MRSA isolates have stronger biofilm formation ability than CA-MSSA, 77.8% vs 25.7%. Therefore, the biofilm formation ability is likely related to the genetic background or the clonal types. In fact, it is necessary to validate the hypothesis with a larger size of samples. The intracellular adhesion molecules are the main mechanism of biofilm formation. All of the isolates harbored the icaA and icaD genes in our study, which implies poor biofilm producers of ST22 would have potential to become strong biofilm producers. The bacterial biofilm communities are surrounded by exopolysaccharides matrix.44 Within this environment, bacteria develop polymicrobial interactions45 and increase antibiotics resistance.46 And some researchers reported a variable biofilm-producing ability of different MRSA isolates.47 According to our analysis, the moderate biofilm producers had a significantly lower sensitivity to Sulfamethoxazole/Trimethoprim. In addition moderate biofilm producers also exhibited lower, although not significantly, sensitivities against several other antibiotics including benzylpenicillin, gentamycin, ciprofloxacin, levofloxacin, and erythromycin. Furthermore, there were no differences in the sensitivities to many other antibiotics including rifampicin, linezolid, moxifloxacin, vancomycin, tetracycline, and tigecycline between different biofilm producers.
Conclusion
In summary, we documented a prevalent community epidemic PVL+ ST22 CA-MSSA which had previously described as a sporadic clone in Beijing, China.6 The changing epidemiology of CA-S. aureus necessitates further surveillance to inform mechanisms of evolution, empiric treatment guidelines, and prevention strategies.
Ethics approval
The ethics committee of Beijing Tsinghua Changgung Hospital approved the study procedure. The reference number of the approval is 19167-0-01.
Abbreviations
CA-S. aureus, community-associated Staphylococcus aureus; SSTIs, skin and soft tissue infections; CA-MSSA, community-associated methicillin-susceptible S.aureus; CA-MRSA, community-associated methicillin-resistant S. aureus; MLST, multilocus sequence typing; PVL, Panton-Valentine Leukocidin; ica, intracellular adhesion molecules; tst-1, toxic shock syndrome toxin-1; etA, exfoliative toxin A; edinA, epidermal cell differentiation inhibitor A; ST, sequence types; CC, clonal complex; TSS, toxic shock syndrome; SSSS, Staphylococcal scalded skin syndrome.
Acknowledgment
This study was supported by Beijing Municipal Administration of Hospitals’ Youth Programme. The funding bodies had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript. We also thank the Clinical Research Center of Beijing Tsinghua Changgung Hospital for research facilities.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Tong SY, Davis JS, Eichenberger E, Holland TL, Fowler VG
2. Liu Y, Xu Z, Yang Z, Sun J, Ma L. Characterization of community-associated Staphylococcus aureus from skin and soft-tissue infections: a multicenter study in China. Emerg Microbes Infect. 2016;5(12):e127. doi:10.1038/emi.2016.119
3. Al Jalaf M, Fadali H, Alanee R, et al. Methicillin resistant Staphylococcus aureus in emergency department patients in the United Arab Emirates. BMC Emerg Med. 2018;18(1):12. doi:10.1186/s12873-018-0164-7
4. DeLeo FR, Otto M, Kreiswirth BN, Chambers HF. Community-associated meticillin-resistant Staphylococcus aureus. Lancet. 2010;375(9725):1557–1568. doi:10.1016/S0140-6736(09)61999-1
5. Gu FF, Chen Y, Dong DP, et al. Molecular epidemiology of Staphylococcus aureus among Patients with skin and soft tissue infections in two Chinese hospitals. Chin Med J (Engl). 2016;129(19):2319–2324. doi:10.4103/0366-6999.190673
6. Zhao C, Liu Y, Zhao M, et al. Characterization of community acquired Staphylococcus aureus associated with skin and soft tissue infection in Beijing: high prevalence of PVL+ ST398. PLoS One. 2012;7(6):e38577. doi:10.1371/journal.pone.0038577
7. Baines SL, Howden BP, Heffernan H, et al. Rapid emergence and evolution of Staphylococcus aureus clones harboring fusC-containing staphylococcal cassette chromosome elements. Antimicrob Agents Chemother. 2016;60(4):2359–2365. doi:10.1128/AAC.03020-15
8. Stegger M, Wirth T, Andersen PS, et al. Origin and evolution of European community-acquired methicillin-resistant Staphylococcus aureus. mBio. 2014;5(5):e01044–e01014. doi:10.1128/mBio.01044-14
9. Yonezawa R, Kuwana T, Kawamura K, Inamo Y. Invasive community-acquired methicillin-resistant staphylococcus aureus in a Japanese girl with disseminating multiple organ infection: a case report and review of Japanese pediatric cases. Case Rep Pediatr. 2015;2015:291025.
10. Rajova J, Pantucek R, Petras P, Varbanovova I, Maslanova I, Benes J. Necrotizing pneumonia due to clonally diverse Staphylococcus aureus strains producing Panton-Valentine leukocidin: the Czech experience. Epidemiol Infect. 2016;144(3):507–515. doi:10.1017/S0950268815001521
11. Tinelli M, Monaco M, Vimercati M, Ceraminiello A, Pantosti A. Methicillin-susceptible Staphylococcus aureus in skin and soft tissue infections, Northern Italy. Emerg Infect Dis. 2009;15(2):250–257. doi:10.3201/eid1502.080010
12. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38(3):1008–1015.
13. Kondo Y, Ito T, Ma XX, et al. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob Agents Chemother. 2007;51(1):264–274. doi:10.1128/AAC.00165-06
14. Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35(6):1547–1549. doi:10.1093/molbev/msy096
15. Yan X, Gu S, Shi Y, Cui X, Wen S, Ge J. The effect of emodin on Staphylococcus aureus strains in planktonic form and biofilm formation in vitro. Arch Microbiol. 2017;199(9):1267–1275. doi:10.1007/s00203-017-1396-8
16. Vasudevan P, Nair MK, Annamalai T, Venkitanarayanan KS. Phenotypic and genotypic characterization of bovine mastitis isolates of Staphylococcus aureus for biofilm formation. Vet Microbiol. 2003;92(1–2):179–185.
17. Boyle-Vavra S, Daum RS. Community-acquired methicillin-resistant Staphylococcus aureus: the role of Panton-Valentine leukocidin. Lab Invest. 2007;87(1):3–9. doi:10.1038/labinvest.3700501
18. G L, Piémont Y, Godail-Gamot F, et al. Involvement of Panton-Valentine leukocidin–producing staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis. 1999;29:1128–1132. doi:10.1086/313461
19. Becker K, Roth R, Peters G. Rapid and specific detection of toxigenic Staphylococcus aureus: use of two multiplex PCR enzyme immunoassays for amplification and hybridization of staphylococcal enterotoxin genes, exfoliative toxin genes, and toxic shock syndrome toxin 1 gene. J Clin Microbiol. 1998;36(9):2548–2553.
20. Aung MS, Urushibara N, Kawaguchiya M, et al. Virulence factors and genetic characteristics of methicillin-resistant and -susceptible Staphylococcus aureus isolates in Myanmar. Microb Drug Resist. 2011;17(4):525–535. doi:10.1089/mdr.2011.0061
21. Jarraud S, Mougel C, Thioulouse J, et al. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun. 2002;70(2):631–641. doi:10.1128/iai.70.2.631-641.2002
22. Yamaguchi T, Nishifuji K, Sasaki M, et al. Identification of the Staphylococcus aureus etd pathogenicity island which encodes a novel exfoliative toxin, ETD, and EDIN-B. Infect Immun. 2002;70(10):5835–5845. doi:10.1128/iai.70.10.5835-5845.2002
23. Stepanovic S, Vukovic D, Dakic I, Savic B, Svabic-Vlahovic M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J Microbiol Methods. 2000;175–179. doi:10.1016/S0167-7012(00)00122-6
24. Htun HL, Kyaw WM, de Sessions PF, et al. Methicillin-resistant Staphylococcus aureus colonisation: epidemiological and molecular characteristics in an acute-care tertiary hospital in Singapore. Epidemiol Infect. 2018;146(14):1785–1792. doi:10.1017/S0950268818001966
25. Hsu LY, Harris SR, Chlebowicz MA, et al. Evolutionary dynamics of methicillin-resistant Staphylococcus aureus within a healthcare system. Genome Biol. 2015;16:81. doi:10.1186/s13059-015-0667-4
26. Manoharan A, Zhang L, Poojary A, Bhandarkar L, Koppikar G, Robinson DA. An outbreak of post-partum breast abscesses in Mumbai, India caused by ST22-MRSA-IV: genetic characteristics and epidemiological implications. Epidemiol Infect. 2012;140(10):1809–1812. doi:10.1017/S0950268812000593
27. Chang Q, Abuelaish I, Biber A, et al. Genomic epidemiology of meticillin-resistant Staphylococcus aureus ST22 widespread in communities of the Gaza Strip, 2009. Euro Surveill. 2018;23(34). doi:10.2807/1560-7917.ES.2018.23.34.1700592
28. de Lencastre H, Baldan R, Testa F, et al. Factors contributing to epidemic MRSA clones replacement in a hospital setting. PLoS One. 2012;7(8):e43153. doi:10.1371/journal.pone.0043153
29. Yamamoto T, Takano T, Yabe S, et al. Super-sticky familial infections caused by Panton-Valentine leukocidin-positive ST22 community-acquired methicillin-resistant Staphylococcus aureus in Japan. J Infect Chemother. 2012;18(2):187–198. doi:10.1007/s10156-011-0316-0
30. Nurjadi D, Fleck R, Lindner A, et al. Import of community-associated, methicillin-resistant Staphylococcus aureus to Europe through skin and soft-tissue infection in intercontinental travellers, 2011-2016. Clin Microbiol Infect. 2019;25(6):739–746.
31. Diep BA, Otto M. The role of virulence determinants in community-associated MRSA pathogenesis. Trends Microbiol. 2008;16(8):361–369. doi:10.1016/j.tim.2008.05.002
32. Klein S, Menz MD, Zanger P, Heeg K, Nurjadi D. Increase in prevalence of PVL and clonal shift in community onset -MRSA causing skin and soft tissue infections in the Rhine-Neckar-region, Germany, 2012-2016. Int J Antimicrob Agents. 2019;53(3):261–267.
33. Tsai MS, Chen CJ, Lin TY, Huang YC. Nasal methicillin-resistant Staphylococcus aureus colonization among otherwise healthy children aged between 2 months and 5 years in northern Taiwan, 2005-2010. J Microbiol Immunol Infect. 2018;51(6):756–762. doi:10.1016/j.jmii.2017.07.014
34. Wang X, Li X, Liu W, Huang W, Fu Q, Li M. Molecular characteristic and virulence gene profiles of community-associated methicillin-resistant Staphylococcus aureus isolates from pediatric patients in Shanghai, China. Front Microbiol. 2016;7:1818. doi:10.3389/fmicb.2016.01818
35. Liu X, Liang J, Jiang Y, et al. Molecular characteristics of community-acquired methicillin-resistant Staphylococcus aureus strains isolated from outpatients with skin and soft tissue infections in Wuhan, China. Pathog Dis. 2016;74(4):ftw026. doi:10.1093/femspd/ftw026
36. Wang X, Liu Q, Zhang H, et al. Molecular characteristics of community-associated Staphylococcus aureus isolates from pediatric patients with bloodstream infections between 2012 and 2017 in Shanghai, China. Front Microbiol. 2018;9:1211. doi:10.3389/fmicb.2018.01211
37. Haque N, Aung MS, Paul SK, et al. Molecular epidemiological characterization of methicillin-susceptible and -resistant Staphylococcus aureus isolated from skin and soft tissue infections in Bangladesh. Microb Drug Resist. 2019;25(2):241–250.
38. Jiang W, Zhou Z, Zhang K, Yu Y. Epidemiological investigation of community-acquired Staphylococcus aureus infection. Genet Mol Res. 2013;12(4):6923–6930. doi:10.4238/2013.December.19.11
39. Chen Y, Liu Z, Duo L, et al. Characterization of Staphylococcus aureus from distinct geographic locations in China: an increasing prevalence of spa-t030 and SCCmec type III. PLoS One. 2014;9(4):e96255. doi:10.1371/journal.pone.0096255
40. Shahini Shams-Abadi M, Halaji M, Hoseini-Alfatemi SM, Gholipour A, Mojtahedi A, Sedigh Ebrahim-Saraie H. Epidemiology of toxic shock syndrome toxin-1 harboring Staphylococcus aureus obtained from clinical samples in Iran: a systematic review and meta-analysis. Annali Igiene. 2018;30(5):391–400. doi:10.7416/ai.2018.2239
41. Liang Y, Tu C, Tan C, et al. Antimicrobial resistance, virulence genes profiling and molecular relatedness of methicillin-resistant Staphylococcus aureus strains isolated from hospitalized patients in Guangdong Province, China. Infect Drug Resist. 2019;12:447–459. doi:10.2147/IDR.S192611
42. Pimentel de Araujo F, Tinelli M, Battisti A, et al. An outbreak of skin infections in neonates due to a Staphylococcus aureus strain producing the exfoliative toxin A. Infection. 2018;46(1):49–54. doi:10.1007/s15010-017-1084-2
43. Khoramian B, Jabalameli F, Niasari-Naslaji A, Taherikalani M, Emaneini M. Comparison of virulence factors and biofilm formation among Staphylococcus aureus strains isolated from human and bovine infections. Microb Pathog. 2015;88:73–77. doi:10.1016/j.micpath.2015.08.007
44. Lopez D, Vlamakis H, Kolter R. Biofilms. Cold Spring Harb Perspect Biol. 2010;2(7):a000398. doi:10.1101/cshperspect.a000398
45. Antunes AL, Trentin DS, Bonfanti JW, et al. Application of a feasible method for determination of biofilm antimicrobial susceptibility in staphylococci. APMIS. 2010;118(11):873–877. doi:10.1111/j.1600-0463.2010.02681.x
46. Fux CA, Costerton JW, Stewart PS, Stoodley P. Survival strategies of infectious biofilms. Trends Microbiol. 2005;13(1):34–40. doi:10.1016/j.tim.2004.11.010
47. Li T, Lu H, Wang X, et al. Molecular characteristics of Staphylococcus aureus causing bovine mastitis between 2014 and 2015. Front Cell Infect Microbiol. 2017;7:127. doi:10.3389/fcimb.2017.00517
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.