Back to Journals » Journal of Inflammation Research » Volume 16
Common Variables That Influence Sepsis Mortality in Mice
Authors Garcia LF, Singh V, Mireles B, Dwivedi AK, Walker WE ![]()
Received 3 December 2022
Accepted for publication 18 February 2023
Published 14 March 2023 Volume 2023:16 Pages 1121—1134
DOI https://doi.org/10.2147/JIR.S400115
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Luiz F Garcia,1 Vishwajeet Singh,2 Blake Mireles,3 Alok Kumar Dwivedi,2– 4 Wendy E Walker1,3
1Center of Emphasis in Infectious Diseases, Department of Molecular and Translational Medicine, Paul L. Foster School of Medicine, Texas Tech University Health Sciences Center El Paso, El Paso, TX, USA; 2Biostatistics and Epidemiology Consulting Lab, Office of Research, Texas Tech University Health Sciences Center El Paso, El Paso, TX, USA; 3Graduate School of Biomedical Sciences, Texas Tech University Health Sciences Center El Paso, El Paso, TX, USA; 4Division of Biostatistics and Epidemiology, Department of Molecular and Translational Medicine, Paul L. Foster School of Medicine, Texas Tech University Health Sciences Center El Paso, El Paso, TX, USA
Correspondence: Wendy E Walker, 5001 El Paso Drive, El Paso, TX, 79905, USA, Tel +1 915 215-4268, Fax +1 915 783-1271, Email [email protected]
Introduction: Sepsis is characterized by a dysregulated host immune response to infection, leading to organ dysfunction and a high risk of death. The cecal ligation and puncture (CLP) mouse model is commonly used to study sepsis, but animal mortality rates vary between different studies. Technical factors and animal characteristics may affect this model in unanticipated ways, and if unaccounted for, may lead to serious biases in study findings. We sought to evaluate whether mouse sex, age, weight, surgeon, season of experiments, and timing of antibiotic administration influenced mortality in the CLP model.
Methods: We created a comprehensive dataset of C57BL/6J mice that had undergone CLP surgery within our lab during years 2015– 2020 from published and unpublished studies. The primary outcome was defined as the time from sepsis induction to death or termination of study (14 days). The Log rank test and Cox regression models were used to analyze the dataset. The study included 119 mice, of which 43% were female, with an average age of 12.6 weeks, an average weight of 25.3 g. 38 (32%) of the animals died.
Results: In the unadjusted analyses, experiments performed in the summer and higher weight predicted a higher risk of mortality. In the stratified Cox model by sex, summer season (adjusted hazard ratio [aHR]=5.61, p=0.004) and delayed antibiotic administration (aHR=1.46, p=0.029) were associated with mortality in males, whereas higher weight (aHR=1.52, p=0.005) significantly affected mortality in females. In addition, delayed antibiotic administration (HR=1.42, p=0.025) was associated with mortality in the non-summer seasons, but not in the summer season.
Discussion: In conclusion, some factors specific to sex and season have a significant influence on sepsis mortality in the CLP model. Consideration of these factors along with appropriate group matching or adjusted analysis is critical to minimize variability beyond the experimental conditions within a study.
Keywords: sepsis, cecal ligation and puncture, mouse, variables, sex, age, weight, season, surgeon, antibiotic timing
A Letter to the Editor has been published for this article.
A Response to Letter by Dr Joffre has been published for this article.
Introduction
Variety is the spice of life. It is crucial to incorporate diversity into our scientific studies to generalize the applicability of our results to subjects of all genders, age groups, weights, races/ethnicities, and individuals with disabilities and comorbidities.1,2 Clear guidelines have been presented for the inclusion of women and minorities in NIH-funded clinical studies involving human subjects.3 In regard to preclinical animal studies, both males and females should be included unless scientifically justified, and sex should be considered as a biological variable.4 There has also been a recent emphasis on including aged mice in research studies.5 However, the inclusion of biological variability beyond the experimental variable can make science more challenging. With more variability, larger group sizes are required to achieve adequate statistical power. To minimize variability, many scientists employ inbred strains (such as C57BL/6) that are almost genetically identical for their research. Most investigators carefully match their experimental animals for sex, and either age or weight, in order to reduce the level of biological variability. Some investigators conduct their study with only one gender, typically male, to avoid perceived effects of the estrus cycle in female mice. Additionally, many investigators use a narrow animal age range to reduce age-related variation. These restrictions may increase the power of the study to test a specific research hypothesis; however, they may limit the generalizability of the findings. Technical variation, arising from environmental fluctuations, as well as differences in the execution of experimental protocols between investigators, can add additional “noise” to the system. There is an increased risk of type I error with biased associations if the condition groups are not adequately matched or adjusted for confounders.
Sepsis is a condition that arises in the context of a severe bacterial, fungal or viral infection, where the immune response becomes dysregulated and organ damage ensues. Cecal ligation and puncture (CLP) is the most commonly used model to study this condition. However, this model lacks standardization,6,7 and sepsis survival rates can vary widely between different labs.8 Although some factors have been demonstrated to influence sepsis mortality (such as the distance of the cecum ligated, needle size and number of punctures),8 it is clear that other unknown factors are at play.
For many animal studies that have been designed to power a specific comparison, the group sizes may be too small to elucidate the effects of common lab and animal variables on sepsis survival. To explore how CLP-sepsis survival is affected by these factors, we conducted a retrospective analysis of a large set of untreated C57BL/6 mice that underwent CLP surgery in our lab. We hypothesized that multiple animal, environmental and technical characteristics are associated with sepsis mortality.
Materials and Methods
Animals
The Institutional Animal Care and Use Committee (IACUC) at Texas Tech University Health Sciences Center El Paso approved all animal experiments. The research was conducted in accordance with the guidelines presented in the federal Animal Welfare Act (AWA) and the Public Health Service (PHS) Policy on the Humane Care and Use of Animals. This study performed an analysis of 119 male and female C57BL/6J mice (6–24 weeks of age) that underwent the CLP sepsis model for various experiments, including published9–12 and unpublished studies from years 2015–2020. Animals that received any experimental treatments (such as neutralizing antibodies or inhibitors) and animals of a different genotype (knockout and transgenic mice) were excluded from the study.
C57BL/6J mice were obtained from Jackson Labs (Bar Harbor, ME) and bred in our Laboratory Animal Resource Center (LARC) facility to supply these experimental mice. Housing and care of the mice occurred under specific pathogen-free conditions in the LARC at our institution. Groups ranging from 1 to 5 mice were housed in nonsterile Optimice cages (Animal Care Systems, Centennial, CO), and given 12hr light/12hr dark cycles. Corncob bedding was supplied along with 5R53 Extruded PicoLab Rodent Diet 20 (Lab Diet, St. Louis, MO). Water was given through either water bottles or lixit drinking valves connected to an automatic watering system. The colony was routinely tested and found to be free from pathogens (including norovirus, helicobacter and other common mouse pathogens) via the use of sentinel mice, to ensure a healthy microbiome. We followed the Animal Research: Reporting In Vivo Experiments (ARRIVE) guidelines for the reporting of in vivo animal experiments in this manuscript.
Sepsis Model and Animal Monitoring
Mice underwent Cecal Ligation and Puncture (CLP) surgery in order to induce sepsis, with a goal of 25–50% mortality. Mice were first anesthetized with isoflurane (2–2.5% with an oxygen flow rate of 1.0) via a precision vaporizer. The animals were weighed and then administered Buprenorphine SR analgesic (1 mg/kg s.c.; ZooPharm, Windsor, CO) prior to surgery. The mouse abdomen was then shaved and scrubbed with betadine and ethanol three times. Puralube Vet ointment (Overland Park, KS) was applied to the eyes to avoid drying during surgery. A midline lateral incision was performed on the skin followed by the peritoneal membrane, exposing the peritoneal cavity. The cecum was then located and 1 cm of its distal portion was ligated with a silk suture. The ligated section was punctured through and through (2 holes) with a 21-gauge needle. A small drop of cecal content was squeezed out through the puncture hole and then the cecum was placed back into the upper left quadrant of the peritoneal cavity. The incision was closed with 5–0 coated Vicryl violet braid suture with RB-1 needle (Ethicon, Somerville, NJ). A continuous suture was used to close the peritoneal membrane followed by an interrupted suture for the skin. CLP surgeries were performed between the hours of 12:00PM and 5:00PM. The animals received 1 mL of Lactated Ringer’s Solution (Covetrus, Chicago, IL) s.c. immediately after surgery and 25 mg/kg imipenem/cilastatin (Primaxin, Merck, Whitehouse Station, NJ) i.p starting at 1–8hr post-surgery, and then twice daily for the following 4 days (please note, the timing of antibiotic initiation was adjusted over the seasons to maintain a mortality rate of 25–50%, according to our experimental observations). Additional doses of Buprenorphine SR were administered at 48–72h intervals for mice with continued signs of discomfort. Mouse surface temperature was measured on the sternum with a La Crosse Technology infrared thermometer, as suggested by a prior report.13 Mice that were unable to regain sternal recumbence when laid on their side were considered moribund and were euthanized. Animals that reached this humane endpoint were recorded as “dead” at the subsequent observational time point. Our model is consistent with the recent guidelines on Minimum Quality Threshold in Pre-Clinical Sepsis Studies (MQTiPSS).7
Covariates
Animal biological sex, age in weeks and weight in grams at the initiation of surgery, surgeon performing CLP surgery, season when the experiment was performed (spring/summer/fall/winter), and timing of the initiation of antibiotics in hours were considered primary covariates in this study. For survival pattern evaluation, these covariates were categorized. In addition, we collected data for the daily average temperature and humidity from September 1, 2018 to August 31 2020, as well as animal surface temperature at 18h post-CLP surgery for each mouse.
Statistical Analysis
Some of the variables in this study were modified in groups for analysis based on the sample size. The characteristics of the entire cohort were described using either mean or median with standard deviation (SD) or range, whereas categorized variables were summarized with number and percentages. We first compared all the characteristics between sexes using either an unpaired t-test or a Fisher’s exact test. The Kruskal–Wallis test was used to compare ambient temperature and humidity as well as animal hypothermia (18h post-CLP) across the four seasons.
The primary outcome was defined as the time from sepsis induction to death or truncation of study (14 days). A Kaplan–Meier curve was constructed to describe survival distribution in the entire cohort as well as by each considered factor. We categorized quantitative covariates into tertiles (T1: low tertile, T2: middle quartile and T3: highest tertile) for describing the survival pattern, otherwise the original form of quantitative data was retained in unadjusted and adjusted analyses.
We tested the normality distribution of continuous variables using the Shapiro–Wilk test. In case of any variable significantly departed from the normal distribution, we used the multivariable fractional polynomial model as recommended in our prior report,14 to assess the non-linear relationship of non-normally distributed variables in the Cox model. Since the results were unchanged with MFP analysis, we presented results with original form of quantitative variables.
All unadjusted comparisons of survival rates between groups were tested using a Log rank test. An unadjusted Cox regression with or without time-varying coefficient model was used to analyze mortality outcome for estimating effect size. We also determined interactions between considered factors on mortality. Due to the presence of significant interactions between sex and weight as well as season and timing of antibiotics, we conducted multivariable Cox regression models separately by sex and season. In the adjusted analysis, the proportionality assumption was tested using Schoenfeld residuals analyses, and the results of Cox proportional hazards models were summarized with the adjusted hazard ratio (aHR), 95% confidence interval (CI) and p-value. P values ≤0.05 were considered statistically significant. In the primary adjusted analysis, we adjusted all the considered variables. We further validated the findings by using a backward stepwise elimination approach with the probability of removal at 10%. GraphPad Prism 6 software (GraphPad, San Diego, CA) was used to generate the Kaplan–Meier curves. All other statistical analyses were conducted using STATA 17 (College Station, Texas). Forest plots were used to display findings obtained from multivariable analyses.
Power Analysis
Previous studies showed a 20% to 35% difference in survival rates with corresponding hazard ratios (HR) of 1.8 to 2.8 according to age and sex.15–19 Based on these estimates, our total sample size of 119 afforded greater than 85% power to detect biologically significant HRs of 1.5 or higher with a 30% mortality rate using a Log rank test and Cox proportional hazard regression analyses. Sample size estimation and simulations were carried out using PASS 21 (PASS 2021 Power Analysis and Sample Size Software (2021). NCSS, LLC. Kaysville, Utah, USA, ncss.com/software/pass).
Results
Description of the Animal Cohort
Table 1 describes the characteristics of the animal cohort and the variables that were analyzed for this study. The cohort was comprised of 119 mice with an average age of 12.57 weeks and an average weight of 25.28 grams. The cohort included both sexes, and 51 (42.86%) of the mice were female. The average timing of antibiotic initiation was 4.57h. The surgeries were performed during all four seasons (spring, summer, fall and winter) in the years 2015–2020. The majority of surgeries were performed in the fall (38.66%) and summer (36.97%) followed by spring (15.97%) and winter (8.4%). Five surgeons performed the CLP procedure. The animal mortality rate was 31.93% with an average time of death or study termination of 249.94h (Table 1 and Figure 1A). Age and weight differences were also observed between the sexes (Table 1). Female mice were slightly older and had a lower weight compared to male mice.
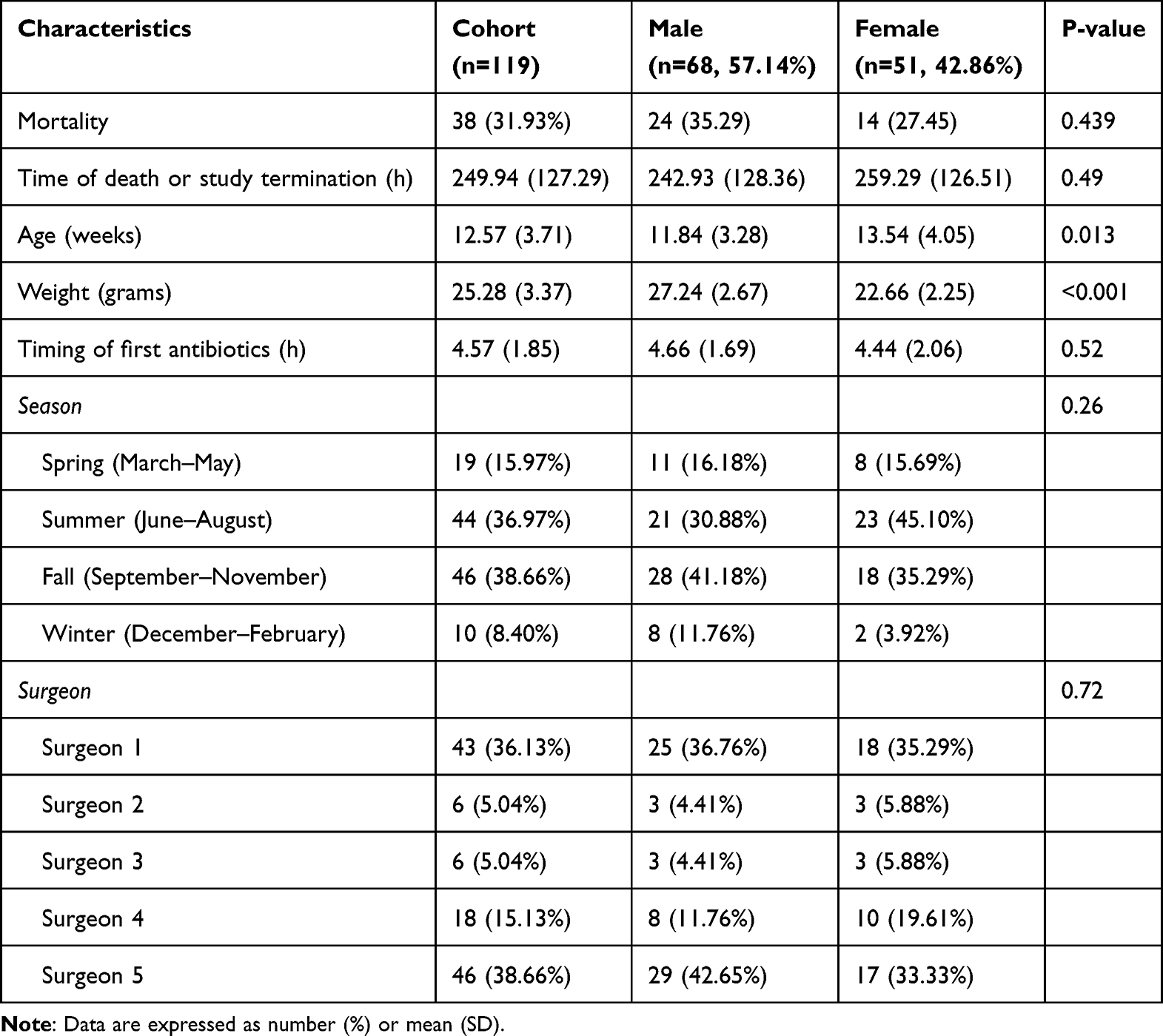 |
Table 1 Characteristics of the Animal Cohort |
 |
Figure 1 Animal survival by sex and age. We analyzed animal survival in a cohort of 119 C57BL/6 mice that underwent CLP. (A) Survival of the entire cohort, (B) Survival in females vs males, (C) Age distribution of mice in the entire cohort, (D) Survival of mice by age groups. Graphs show Kaplan–Meier plots for animal survival, and individual values for mouse age. Survival data were compared via the Log rank test. |
The Impact of Biological Sex on Sepsis Survival
We first analyzed the impact of sex on CLP sepsis survival (Figure 1B). We observed a trend towards more rapid mortality during the acute phase (0–72h post-surgery) in female vs male mice, and a trend towards a higher mortality rate in the later phase (after 72h) in male vs female mice. However, these differences were not statistically significant (HRfemale=0.79, p=0.49).
The Impact of Age on Sepsis Survival
Next, we analyzed the impact of animal age on sepsis survival. The distribution of animal ages in our cohort is shown in Figure 1C. We observed a trend towards a slightly greater mortality rate in the highest tertile (age >13.12 weeks) in comparison to the middle and lowest tertiles (aged 6.1–13.11 weeks; Figure 1D). However, these differences were not statistically significant (HR=1.06, p=0.15).
The Impact of Weight on Sepsis Survival
We next examined the impact of animal weight. We observed a higher mortality in the highest tertile (>27.01 grams) in comparison to the other two tertiles (Figure 2A). However, these differences were only statistically significant at borderline (HR=1.10, p=0.052). As mentioned, on average, the male mice in the study were found to be significantly heavier than the female mice (p<0.001; Figure 2B). Males averaged at about 27 grams and females at about 23 grams. Due to the presence of an interaction effect between weight and sex, we analyzed the effect of weight within each sex individually. We could see again that there was a trend towards higher mortality in the male mice in the highest tertile (>28.70 grams) in comparison to the other two tertiles (Figure 2C); however, this effect was not statistically significant (HRmale=1.05, p=0.548). Higher weight was found to significantly affect mortality in female mice (HRfemale=1.44, p=0.003; Figure 2D).
 |
Figure 2 Animal survival by weight. (A) Survival of all mice by weight groups, (B) Weight of male vs female mice, (C) Survival by weight groups in male mice, (D) Survival by weight groups in female mice. Graphs show Kaplan–Meier plots for animal survival, and individual values with the median for animal weight. Survival data were compared via the Log rank test. Weight distributions were compared via an unpaired t-test. |
The Impact of the Animal Surgeon on Sepsis Survival
For this cohort of animals, five different surgeons performed the CLP procedure. Although some variability in time to mortality and mortality incidence was observed between surgeons (Figure 3), the difference was not statistically significant (p=0.547). Notably, the highest and lowest mortality rates were achieved for surgeons that only operated limited numbers (n=6 mice each), and this most likely reflects a sampling effect.
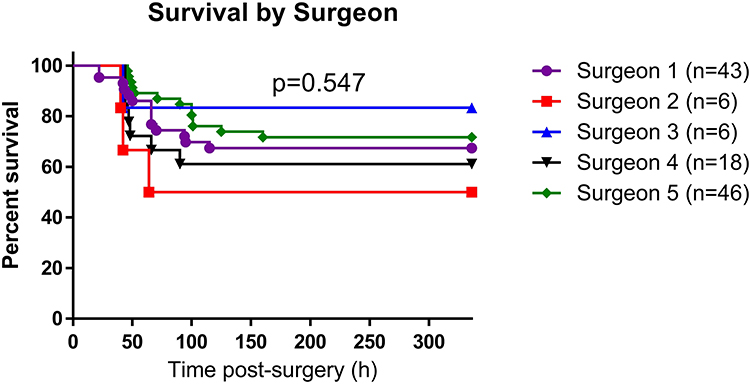 |
Figure 3 Animal survival by surgeon. Graph shows Kaplan–Meier plot of animal survival. Data were compared via the Log rank test. |
Seasonal Effects on Sepsis Survival
Next, we considered whether there were seasonal differences in sepsis survival. We observed a trend towards higher mortality rates in animals that underwent CLP in the summer months (Figure 4A), although this variability did not reach a statistically significant level (HRsummer=1.86, p=0.056). As described in our prior reports,9–12 we noticed these period fluctuations in the severity of our sepsis model, and adjusted the timing of the first dose of antibiotics to maintain a mortality rate of 25–50%. The antibiotic was given earliest in the summer season (Figure 4B); nevertheless, survival rates remained worst during this season. We also considered if seasonal effects occurred in male vs female mice. Among the male mice, we observed a substantially higher mortality rate in the summer versus the other seasons (Figure 4C) and this effect was statistically significant (HRmale=2.61, p=0.02). In contrast, female mice showed only a slight trend towards increasing mortality in the summer vs the other seasons (Figure 4D) and this difference was not statistically significant (HRfemale=1.31, p=0.614).
 |
Figure 4 Animal survival by season. (A) Survival of all mice by season, (B) Timing of first antibiotic dose by season, (C) Survival in summer vs other seasons for male mice, (D) Survival in summer vs other seasons for female mice. Graphs show Kaplan–Meier plots for animal survival, and individual values with the median for the timing of first antibiotic dose. Survival data were compared via the Log rank test. Timing of the first antibiotic dose was compared via the Kruskal–Wallis test. |
The light cycle within our LARC facility is set to 12h light/12h dark across the seasons, and the temperature and humidity are regulated to maintain the animals’ well-being. However, we considered that fluctuations might still occur within the regulated range. To test this, we obtained the facility data for the daily average temperature and humidity in our animal housing room from September 2018 to August 2020. We observed small variations in daily temperature between the seasons, with the highest median value in the summer and lowest in the winter (Supplemental Figure 1A), and these minimum differences achieved a borderline statistical significance (p=0.0526). However, we observed notable variations in the humidity between the seasons, with the highest median value in the summer and lowest in the winter (Supplemental Figure 1B), and this effect was statistically significant (p<0.0001). We considered that higher ambient temperatures in the summer might prevent the mice from developing natural protective hypothermia, leading to the higher sepsis rates in this season. To test this, we compared the mouse surface temperatures at 18h post-surgery for animals that had undergone CLP in each season. However, we found that the median mouse surface temperatures (18h post-CLP) were comparable for animals that underwent CLP in the summer vs winter, and both were lower than the temperature of animals that underwent CLP in the spring or fall (Supplemental Figure 1C); these differences were significant (p=0.0059). These data refute the notion that attenuated hypothermia mediates the higher mortality rates observed in the summer.
The Impact of Antibiotic Timing on Sepsis Survival
We did not observe any overall trend in animal mortality according to the timing of the first antibiotic dose (Figure 5, HR=0.97, p=0.765). However, due to adjustment of timing of antibiotic administration, particularly in the summer season, there was an interaction between season and timing of antibiotic administration on mortality. Delayed antibiotic administration was associated with increased mortality in the non-summer season (HR=1.42, p=0.025) but not in the summer season.
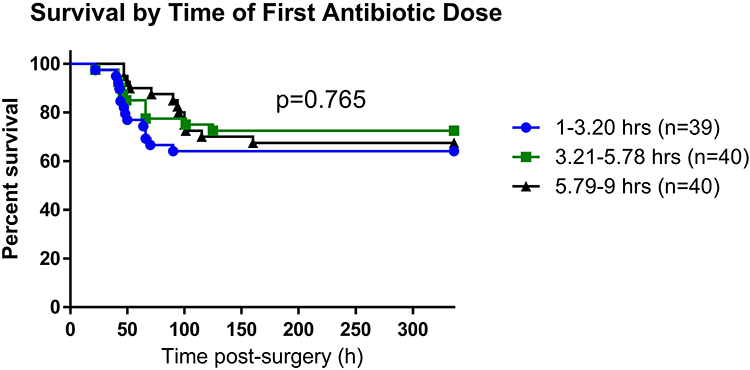 |
Figure 5 Animal survival according to antibiotic timing. Graph shows Kaplan–Meier plot of animal survival. Data were compared via the Log rank test. |
Adjusted Effect of Each Variable on Sepsis Survival
Due to significant interaction of sex with season and weight, we performed an adjusted analysis for each sex separately (Figure 6). In the adjusted analysis, summer season (aHR=5.6, p=0.004) and timing of antibiotic administration (aHR=1.46, p=0.029) were associated with mortality in male mice. However, only higher weight was associated with increased mortality rate (aHR=1.52, p=0.005) in female mice. We also conducted adjusted analysis separately for summer and non-summer seasons (Figure 7), due to the presence of a significant interaction between season and timing of antibiotic administration. Increased timing of antibiotic administration was associated with higher mortality rate (aHR=1.46, p=0.02) in the non-summer season only. Results were unchanged after applying Cox analysis with a backward elimination approach.
 |
Figure 6 Factors associated with mortality using Cox analysis by sex. Figure shows a forest plot, along with the adjusted hazard ratio (HR) and P value for each factor associated with mortality in male and female mice. The horizontal line shows a 95% confidence interval (CI) associated with each HR for each factor. Data were analyzed with Cox regression analysis. |
 |
Figure 7 Factors associated with mortality using Cox analysis by season. Figure shows a forest plot, along with the adjusted hazard ratio (HR) and P value for each factor associated with mortality for non-summer and summer seasons. The horizontal line shows a 95% confidence interval (CI) associated with each HR for each factor. Data were analyzed with Cox regression analysis. |
Discussion
It is well known that sepsis survival rates vary in the CLP model when comparing different studies, even when the experimental model is described in a very similar fashion. The research presented here highlights some of the common factors that may drive this heterogeneity. Interestingly, we identified multiple factors that were specific to animal sex. Among males, summer season and increased elapsed time to antibiotic administration were associated with mortality. In contrast, higher weight significantly affected mortality among females. In addition, increased elapsed time to antibiotic administration was strongly associated with mortality in non-summer seasons.
There have been mixed reports on how sex affects sepsis in clinical studies, as well as rodent sepsis models (for the sake of brevity, these are summarized in two supplemental tables: Supplemental Table 1 for clinical studies and Supplemental Table 2 for animal studies). Most of these studies reported improved survival in females, although others reported equal outcomes or improved survival in males; and sex hormones were implicated to play a role in this effect (see Supplemental Tables 1 and 2). A recent report highlighted the need to re-examine the impact of sex on sepsis outcomes in the context of antibiotic treatment.17 The results presented in our study suggest a trend towards more rapid mortality during the acute phase (0–72h) in female vs male mice, paired with higher rates of chronic mortality (after 72h) in male vs female mice. This mirrors the findings reported in our recent report on aged mice.20 However, even with the large number of animals included in our cohort, the independent effect of animal sex on sepsis mortality was not statistically significant. In addition to considering the overall effects of sex, the phenotype of female mice could potentially vary according to the estrus cycle. A prior study found no significant differences in sepsis mortality among female mice according to the estrus stage; however, white blood cell and monocyte counts were significantly higher in metestrus vs estrus mice at 12h post-surgery, and there was a trend towards greater mortality in the metestrus group.21 Therefore, the effects of the estrus stage on sepsis mortality may warrant further study.
It is common to use a narrow age range of mice in scientific studies, often in the range of 6–8 or 10–12 weeks, reflecting adolescent or young adult cohorts, respectively.22 However, many laboratories including our own use a broader range of ages. This study included animals aged 6–24 weeks of age. Although it is not possible to make a strict comparison, this roughly equates to an age range of adolescents to 30 years old people.23 Within this cohort, we observed a trend toward higher mortality rates in the tertile of oldest mice (age ≥13.12 weeks) in comparison to the two tertiles consisting of younger mice, however this effect was not statistically significant. Therefore, within this range of adolescent to mature adult mice, age may have a minimal effect on sepsis mortality. In contrast, many clinical reports and animal studies have shown that advanced age is associated with higher sepsis mortality rates (for the sake of brevity, these are summarized in Supplemental Table 3). Aged mice exhibit differences in the local cytokine response to sepsis, an impeded ability to return to immune homeostasis after sepsis, an altered metabolomic response, and increased vascular oxidative damage, endothelial dysfunction and coagulopathy: all of these alterations likely contribute to the age-related exacerbation of sepsis outcomes (see Supplemental Table 3). Further supporting this notion, our own recent report showed that aged C57BL/6 mice exhibited a 90% mortality rate20 when subject to the same sepsis model described here. Please note that we did not include those aged animals in the current analysis. This was because of the large age gap between those aged mice and the animals included in this study, and because the use of aged mice in research is not a “common variable” but an intentional decision, due to the high cost and practical aspects associated with their use.
Weight affects many aspects of animal physiology. Obesity can induce chronic inflammation and oxidative stress,24 and is associated with increased risk for many serious diseases and health conditions. Two animal studies examined the phenotype of obese mice during sepsis. Following CLP, obese mice had an increased inflammatory response and exacerbated microvascular dysfunction, in comparison to lean mice.25,26 However, less is known about the impact of weight on sepsis among non-obese animals, and our research provides novel information on this topic. We found a positive association between animal weight and sepsis mortality, specifically in female mice, among a cohort of non-obese mice. One possibility is that fluid dosing explains the worse outcomes of the heavier female mice in our study, because each animal received 1 mL lactated ringer’s solution after surgery, rather than a weight-based amount of fluids; this requires future exploration. Another possibility is that increased weight promotes inflammation in females, even below the range of obesity. Further research is warranted to examine this.
On our planet Earth, the length of daylight, temperature, humidity and other factors vary across the four seasons, and this has a substantial impact on our physiology. A retrospective analysis identified seasonal variations in the incidence of nosocomial infections in climate-controlled hospitalized patients, and found an increased incidence of bacterial infections in the warmer months, with a stronger effect for gram-negative bacteria.27 Interestingly, a prior report showed that within a biobank of human specimens collected in the United Kingdom, multiple immune parameters are affected by season.28 In particular, C-reactive protein and neutrophil counts peaked in the winter, while lymphocyte counts peaked in the spring.28 Regarding sepsis models, many labs anecdotally report seasonal variations in the outcomes of their CLP model. In our own lab, we observed periodic changes in the performance of our CLP model that seemed to have a large effect on the sepsis survival rates. To attenuate this effect, we adjusted the timing of antibiotic initiation, attempting to maintain mortality rates of about 25–50% (similar to the survival rates observed in patients with severe sepsis). Despite this, we found a higher rate of sepsis mortality in male animals that underwent CLP in the summer months vs the other seasons. In agreement with our results, a prior report by Kiank et al conducted in Germany found that mice subject to colon ascendens stent peritonitis (CASP) in the summer or autumn had an increased risk of septic death in comparison to animals where CASP was induced in the winter or spring.29 This phenotype was associated with a more pronounced inflammatory cytokine response.29 In contrast to our findings, a prior report by Lewis et al analyzed 115 historic mice from their lab and found no seasonal differences in sepsis mortality.30 We predict that the difference between the results of our study and Kiank et al29 vs those of Lewis et al30 may stem from differences in the conditions within our cities and our animal facilities.
Within Public Health Service-approved animal facilities in the USA (and the equivalent in other countries), LARC facilities maintain carefully controlled environments to ensure the welfare of our animals. The light cycle is typically set to 12h light/12 dark across the four seasons, and the temperature and humidity are regulated within an approved range. Despite this, there is likely to be some variation in the temperature and humidity in all LARC facilities across the four seasons, due to natural variations in the outdoor temperature and humidity. In our facility, we observed that during the summer season, the median temperature was slightly higher, and the median humidity was notably higher. The ambient temperature and humidity affect many aspects of animal physiology with potential downstream effects on sepsis. A recent report highlighted the importance of considering the effects of ambient temperature in animal sepsis models, particularly because the “room temperature” that is typically used for animal housing falls below the thermoneutral conditions for a mouse, potentially inducing cold-stress.31 Two reports have specifically investigated the effect of ambient temperature on mouse sepsis models. Ganeshan et al found that higher ambient temperature (30°C vs 22°C) increased animal mortality in a model of E. coli-induced bacteremia.32 Contrasting with this report, Carpenter et al found that higher ambient temperatures (30°C vs 22°C) improved survival of male mice after CLP.33 The reasons for this are likely complex. Body temperature affects many aspects of the immune response including antigen presentation, leukocyte trafficking and activation.34 High ambient temperatures reduced levels of serum corticosterone in mice, and increased immune cell proinflammatory cytokine production in response to LPS.35 Higher ambient temperatures were also shown to alter the gut microbiome of mice.35 Notably in our report, we considered that the higher ambient temperature in the summer might prevent hypothermia following CLP; however, we found that the animal surface temperature was lower in the summer seasons, refuting this notion. Regarding the effects of humidity, additional evaporative loss of water at low humidity levels and differential water intake according to the temperature and humidity may affect sepsis survival. These could potentially affect males and females to a different extent due to their body size and composition.
Rapid administration of broad-spectrum antibiotics (within 1 hour of recognition) is a standard treatment for patients suspected of sepsis. However, it is important to note that patients typically do not present at the clinic until they are significantly ill, which may be hours or days after the initial infection. Rapid administration of antibiotics was shown to reduce sepsis mortality in clinical studies, including a retrospective analysis36 and a single-center cohort study.37 However, these results are not unequivocal as a subsequent meta-analysis showed no significant mortality benefit of rapid antibiotic administration following recognition of sepsis.38 A prior animal study showed improved sepsis survival when antibiotics were administered to mice early vs late after CLP.39 Our results also suggest that antibiotic timing influences sepsis mortality, but only in male mice. Additionally, the effects of antibiotics were only significant in non-summer seasons. This is probably due to the fact that summer season increased animal mortality in a manner that was not amenable to antibiotic rescue. Finally, our data reveal some trends toward differences in sepsis survival between different surgeons; however, these were not statistically significant. Nevertheless, it is clear that careful training of lab staff is crucial to minimize variation within a laboratory.
Germane to this study, many other factors significantly influence sepsis mortality in the CLP model. Prior reports have demonstrated that CLP sepsis mortality is significantly affected by the length of the cecal ligation,40,41 the needle size,42,43 the number of punctures,43 the fed vs fasted state,43 and the use of fluid resuscitation and antibiotics.44–46 As well, individual mouse strains exhibit different levels of sensitivity to CLP-induced sepsis, and outbred strains may perform differently to inbred strains.47–49 Variations in the CLP sepsis phenotype may occur between mice bred in the investigator’s laboratory animal resource center vs animals purchased from a commercial supplier. A prior report showed that mice purchased from two different commercial vendors exhibited different mortality rates, resulting from differences in the microbiome that affect the host immune response.50 Additionally, the anesthetics47 and analgesics51 administered to surgical animals can affect sepsis mortality. Finally, our own lab showed that the timing of CLP surgery affects sepsis survival, with greater mortality rates when infection was induced in the nighttime vs during the day.52,53
Conclusion
We believe that the inclusion of diversity along with proper reporting is critical in both animal and human research. To protect animal welfare and minimize experimental costs, we must also strive to reduce the number of animals required for our research studies. We propose that these goals can be balanced by including biological variability within our research (eg, including mice of both genders, and animals with a range of ages and weights) while minimizing technical variability. In this context, appropriate matching of experimental groups is important for scientific rigor; otherwise, adjusted analysis typically with larger sample sizes is required to yield unbiased and accurate findings. In fact, adjusted analysis is often required for animal studies due to multiple confounders, as detailed in our prior reports.54,55 In addition, appropriate stratification based on modifiable characteristics such as sex or season may be useful in some experimental studies. Our study suggests that for the CLP model, careful matching of animal weight is important, especially among females. Technical variability could potentially be reduced by identifying the factors that drive seasonal variations, and improving environmental homogeneity within the LARC facilities throughout the year. Additionally, careful training in surgical technique can minimize the variation between laboratory personnel. Reporting bias in sex and age has been observed in mouse models,56 and our manuscript highlights that there is need to assess reporting biases related to these other factors in preclinical animal sepsis studies, including animal weight. Finally, the ARRIVE guidelines57 provide a recommended list of details that should be included in any manuscript describing animal research, including animal species, strain, sex, age, weight and other details. Careful reporting of these items will enable the research community to better interpret published findings.
Abbreviations
ARRIVE, Animal Research: Reporting In Vivo Experiments; aHR, adjusted hazard ratio; BMI, body mass index; CASP, colon ascendens stent peritonitis; CI, confidence interval; CLP, cecal ligation and puncture; HR, hazard ratio; IACUC, The Institutional Animal Care and Use Committee; LARC, Laboratory Animal Care Resource Center; MQTiPSS, Minimum Quality Threshold in Pre-Clinical Sepsis Studies.
Acknowledgments
Experimental studies were supported by a SCCM-Weil grant from the Society of Critical Care Medicine, a Shock Society Research Fellowship for Early Career Investigators, a seed grant from TTUHSC El Paso and a grant from the El Paso Community Foundation (all awarded to WEW). WEW and AKD were supported by an NHLBI R15 grant 1R15HL159554-01A1 (awarded to WEW) during the data analysis and manuscript preparation. The authors thank Erica Heipertz, Dinesh Goswami, Devender Kumar and Alejandra Gallegos for generating data for the manuscript, and the veterinarian and laboratory animal resource center staff at TTUHSC El Paso for caring for our animals.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Editors, Rubin, E. Striving for diversity in research studies. N Engl J Med. 2021;385(15):1429–1430.
2. Liu KA, Mager NAD. Women’s involvement in clinical trials: historical perspective and future implications. Pharm Pract. 2016;14(1):708.
3. Kelty M, Bates A, Pinn VW. National Institutes of Health policy on the inclusion of women and minorities as subjects in clinical research. In: Principles and Practice of Clinical Research. Elsevier; 2007:129–142.
4. Miller LR, Marks C, Becker JB, et al. Considering sex as a biological variable in preclinical research. FASEB J. 2017;31(1):29–34. doi:10.1096/fj.201600781r
5. Jackson SJ, Andrews N, Ball D, et al. Does age matter? The impact of rodent age on study outcomes. Lab Anim. 2017;51(2):160–169. doi:10.1177/0023677216653984
6. Remick DG, Ayala A, Chaudry IH, et al. Premise for standardized sepsis models. Shock. 2019;51(1):4–9. doi:10.1097/SHK.0000000000001164
7. Osuchowski MF, Ayala A, Bahrami S, et al. Minimum quality threshold in pre-clinical sepsis studies (MQTiPSS): an international expert consensus initiative for improvement of animal modeling in sepsis. Intensive Care Med Exp. 2018;6(1):26. doi:10.1186/s40635-018-0189-y
8. Lewis AJ, Seymour CW, Rosengart MR. Current murine models of sepsis. Surg Infect. 2016;17(4):385–393. doi:10.1089/sur.2016.021
9. Goswami DG, Garcia LF, Dodoo C, et al. Evaluating the timeliness and specificity of CD69, CD64 and CD25 as biomarkers of sepsis in MICE. Shock. 2021;55(4):507–518. doi:10.1097/SHK.0000000000001650
10. Goswami DG, Rubio AJ, Mata J, Munoz S, Gallegos A, Walker WE. Large peritoneal macrophages and transitional premonocytes promote survival during abdominal sepsis. ImmunoHorizons. 2021;5(12):994–1007. doi:10.4049/immunohorizons.2100086
11. Heipertz EL, Harper J, Goswami DG, et al. IRF3 signaling within the mouse stroma influences sepsis pathogenesis. J Immunol. 2021;206(2):398–409. doi:10.4049/jimmunol.1900217
12. Heipertz EL, Harper J, Walker WE. STING and TRIF contribute to mouse sepsis, depending on severity of the disease model. Shock. 2017;47(5):621–631. doi:10.1097/SHK.0000000000000771
13. Laitano O, Van Steenbergen D, Mattingly AJ, et al. Xiphoid surface temperature predicts mortality in a murine model of septic shock. Shock. 2018;50(2):226–232. doi:10.1097/SHK.0000000000001007
14. Dwivedi AK. How to write statistical analysis section in medical research. J Investig Med. 2022;2022:jim-2022–002479.
15. Zellweger R, Wichmann MW, Ayala A, Stein S, DeMaso CM, Chaudry IH. Females in proestrus state maintain splenic immune functions and tolerate sepsis better than males. Crit Care Med. 1997;25(1):106–110. doi:10.1097/00003246-199701000-00021
16. Hoogland IC, Houbolt C, van Westerloo DJ, van Gool WA, van de Beek D. Systemic inflammation and microglial activation: systematic review of animal experiments. J Neuroinflammation. 2015;12:114. doi:10.1186/s12974-015-0332-6
17. Zhang M, Montroy J, Sharma R, et al. The effects of biological sex on sepsis treatments in animal models: a systematic review and a narrative elaboration on sex- and gender-dependent differences in sepsis. Crit Care Explorations. 2021;3(6):e0433–e0433. doi:10.1097/CCE.0000000000000433
18. Diodato MD, Knoferl MW, Schwacha MG, Bland KI, Chaudry IH. Gender differences in the inflammatory response and survival following haemorrhage and subsequent sepsis. Cytokine. 2001;14(3):162–169. doi:10.1006/cyto.2001.0861
19. Angele MK, Frantz MC, Chaudry IH. Gender and sex hormones influence the response to trauma and sepsis: potential therapeutic approaches. Clinics. 2006;61(5):479–488. doi:10.1590/S1807-59322006000500017
20. Goswami DG, Walker WE. Aged IRF3-KO mice are protected from sepsis. J Inflamm Res. 2021;14:5757–5767. doi:10.2147/JIR.S335203
21. Kennedy LH, Hwang H, Wolfe AM, Hauptman J, Nemzek-Hamlin JA. Effects of buprenorphine and estrous cycle in a murine model of cecal ligation and puncture. Comp Med. 2014;64(4):270–282.
22. Brust V, Schindler PM, Lewejohann L. Lifetime development of behavioural phenotype in the house mouse (Mus musculus). Front Zool. 2015;12(S1):1–14. doi:10.1186/1742-9994-12-S1-S17
23. Dutta S, Sengupta P. Men and mice: relating their ages. Life Sci. 2016;152:244–248. doi:10.1016/j.lfs.2015.10.025
24. Fernández-Sánchez A, Madrigal-Santillán E, Bautista M, et al. Inflammation, oxidative stress, and obesity. Int J Mol Sci. 2011;12(5):3117–3132. doi:10.3390/ijms12053117
25. Vachharajani V, Russell JM, Scott KL, et al. Obesity exacerbates sepsis-induced inflammation and microvascular dysfunction in mouse brain. Microcirculation. 2005;12(2):183–194. doi:10.1080/10739680590904982
26. Singer G, Stokes KY, Terao S, Granger DN. Sepsis-induced intestinal microvascular and inflammatory responses in obese mice. Shock. 2009;31(3):275–279. doi:10.1097/SHK.0b013e3181834ab3
27. Schwab F, Gastmeier P, Hoffmann P, Meyer E. Summer, sun and sepsis-The influence of outside temperature on nosocomial bloodstream infections: a cohort study and review of the literature. PLoS One. 2020;15(6):e0234656–e0234656. doi:10.1371/journal.pone.0234656
28. Wyse C, O’Malley G, Coogan AN, McConkey S, Smith DJ. Seasonal and daytime variation in multiple immune parameters in humans: evidence from 329,261 participants of the UK Biobank cohort. iScience. 2021;24(4):102255. doi:10.1016/j.isci.2021.102255
29. Kiank C, Koerner P, Keler W, et al. Seasonal variations in inflammatory responses to sepsis and stress in mice*. Crit Care Med. 2007;35(10):2352–2358. doi:10.1097/01.CCM.0000282078.80187.7F
30. Lewis A, Zuckerbraun B, Griepentrog J, Zhang X, Rosengart M. Reducing animal use with a biotelemetry-enhanced murine model of sepsis. Sci Rep. 2017;7(1):6622. doi:10.1038/s41598-017-05497-5
31. Helbing DL, Stabenow LK, Bauer R. Mouse sepsis models: don’t forget ambient temperature! Intensive Care Med Exp. 2022;10(1):29. doi:10.1186/s40635-022-00457-4
32. Ganeshan K, Nikkanen J, Man K, et al. Energetic trade-offs and hypometabolic states promote disease tolerance. Cell. 2019;177(2):399–413.e312. doi:10.1016/j.cell.2019.01.050
33. Carpenter KC, Zhou Y, Hakenjos JM, Fry CD, Nemzek JA. Thermoneutral housing temperature improves survival in a murine model of polymicrobial peritonitis. Shock. 2020;54(5):688–696. doi:10.1097/SHK.0000000000001551
34. Appenheimer MM, Evans SS. Chapter 24 - temperature and adaptive immunity. In: Romanovsky AA, editor. Handbook of Clinical Neurology. Vol. 156. Elsevier; 2018:397–415.
35. Giles DA, Moreno-Fernandez ME, Stankiewicz TE, et al. Thermoneutral housing exacerbates nonalcoholic fatty liver disease in mice and allows for sex-independent disease modeling. Nat Med. 2017;23(7):829–838. doi:10.1038/nm.4346
36. Ferrer R, Martin-Loeches I, Phillips G, et al. Empiric antibiotic treatment reduces mortality in severe sepsis and septic shock from the first hour: results from a guideline-based performance improvement program. Crit Care Med. 2014;42(8):1749–1755. doi:10.1097/CCM.0000000000000330
37. Gaieski DF, Mikkelsen ME, Band RA, et al. Impact of time to antibiotics on survival in patients with severe sepsis or septic shock in whom early goal-directed therapy was initiated in the emergency department. Crit Care Med. 2010;38(4):1045–1053. doi:10.1097/CCM.0b013e3181cc4824
38. Sterling SA, Miller WR, Pryor J, Puskarich MA, Jones AE. The impact of timing of antibiotics on outcomes in severe sepsis and septic shock: a systematic review and meta-analysis. Crit Care Med. 2015;43(9):1907–1915. doi:10.1097/CCM.0000000000001142
39. Vyas D, Javadi P, DiPasco PJ, Buchman TG, Hotchkiss RS, Coopersmith CM. Early antibiotic administration but not antibody therapy directed against IL-6 improves survival in septic mice predicted to die on basis of high IL-6 levels. Am J Physiol. 2005;289(4):R1048–R1053. doi:10.1152/ajpregu.00312.2005
40. Singleton K, Wischmeyer P. Distance of cecum ligated influences mortality, tumor necrosis factor-alpha and interleukin-6 expression following cecal ligation and puncture in the rat. Eur Surg Res. 2003;35(6):486–491. doi:10.1159/000073387
41. Ruiz S, Vardon-Bounes F, Merlet-Dupuy V, et al. Sepsis modeling in mice: ligation length is a major severity factor in cecal ligation and puncture. Intensive Care Med Exp. 2016;4(1):22. doi:10.1186/s40635-016-0096-z
42. Ebong S, Call D, Nemzek J, Bolgos G, Newcomb D, Remick D. Immunopathologic alterations in murine models of sepsis of increasing severity. Infect Immun. 1999;67(12):6603–6610. doi:10.1128/IAI.67.12.6603-6610.1999
43. Wichterman KA, Baue AE, Chaudry IH. Sepsis and septic shock—a review of laboratory models and a proposal. J Surg Res. 1980;29(2):189–201. doi:10.1016/0022-4804(80)90037-2
44. Newcomb D, Bolgos G, Green L, Remick DG. Antibiotic treatment influences outcome in murine sepsis: mediators of increased morbidity. Shock. 1998;10(2):110–117. doi:10.1097/00024382-199808000-00005
45. Zanotti-Cavazzoni SL, Goldfarb RD. Animal models of sepsis. Crit Care Clin. 2009;25(4):703–719. doi:10.1016/j.ccc.2009.08.005
46. Iskander KN, Vaickus M, Duffy ER, Remick DG. Shorter duration of post-operative antibiotics for cecal ligation and puncture does not increase inflammation or mortality. PLoS One. 2016;11(9):e0163005. doi:10.1371/journal.pone.0163005
47. Imai T, Takahashi K, Masuo F, Goto F. Anaesthesia affects outcome of sepsis in mice. Can J Anaesth. 1998;45(4):360–366. doi:10.1007/BF03012029
48. Doi K, Leelahavanichkul A, Yuen PST, Star RA. Animal models of sepsis and sepsis-induced kidney injury. J Clin Invest. 2009;119(10):2868–2878. doi:10.1172/JCI39421
49. Zhang J, Liu ZG, Luo YW, et al. C57BL/6小鼠与 BALB/c 小鼠对脓毒症反应的差异 [Differences in the response to sepsis between C57BL/6 and BALB/c mice]. Nan Fang Yi Ke Da Xue Xue Bao. 2010;30(5):973–975. Chinese.
50. Fay KT, Klingensmith NJ, Chen CW, et al. The gut microbiome alters immunophenotype and survival from sepsis. FASEB J. 2019;33(10):11258–11269. doi:10.1096/fj.201802188R
51. Cotroneo TM, Hugunin KM, Shuster KA, Hwang HJ, Kakaraparthi BN, Nemzek-Hamlin JA. Effects of buprenorphine on a cecal ligation and puncture model in C57BL/6 mice. J Am Assoc Lab Anim Sci. 2012;51(3):357–365.
52. Silver AC, Arjona A, Walker WE, Fikrig E. The circadian clock controls toll-like receptor 9-mediated innate and adaptive immunity. Immunity. 2012;36(2):251–261. doi:10.1016/j.immuni.2011.12.017
53. Heipertz EL, Harper J, Lopez CA, Fikrig E, Hughes ME, Walker WE. Circadian rhythms influence the severity of sepsis in mice via a TLR2-dependent, leukocyte-intrinsic mechanism. J Immunol. 2018;201:193–201. doi:10.4049/jimmunol.1701677
54. Dwivedi AK. How to write statistical analysis section in medical research. J Investig Med. 2022;70:1759–1770. doi:10.1136/jim-2022-002479
55. Dwivedi AK, Shukla R. Evidence‐based statistical analysis and methods in biomedical research (SAMBR) checklists according to design features. Cancer Rep. 2020;3(4):e1211. doi:10.1002/cnr2.1211
56. Flórez-Vargas O, Brass A, Karystianis G, et al. Bias in the reporting of sex and age in biomedical research on mouse models. eLife. 2016;5:e13615. doi:10.7554/eLife.13615
57. Percie du Sert N, Hurst V, Ahluwalia A, et al. The ARRIVE guidelines 2.0: updated guidelines for reporting animal research. J Cereb Blood Flow Metab. 2020;40(9):1769–1777. doi:10.1177/0271678X20943823
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
C-Terminal Fibronectin Exerts Beneficial Effects in Reducing Tissue Damage and Modulating Macrophage Function in a Murine Septic Model
Geng H, Wu Y, Chen Y
Journal of Inflammation Research 2023, 16:1509-1521
Published Date: 10 April 2023
A Novel Retinal Nerve Fiber Layer Biomarker of Amyotrophic Lateral Sclerosis (ALS) Identified Using Longitudinal in vivo Ocular Imaging
Khorrami F, Gupta N, Zhou X, Liang Y, Yucel YH
Eye and Brain 2025, 17:69-79
Published Date: 2 July 2025