Back to Journals » International Journal of Nanomedicine » Volume 20
Combined Hyaluronic Acid Nanobioconjugates Impair CD44-Signaling for Effective Treatment Against Obesity: A Review of Comparison with Other Actors
Authors Uti DE ![]() , Omang WA, Alum EU
, Omang WA, Alum EU ![]() , Ugwu OPC
, Ugwu OPC ![]() , Wokoma MA, Oplekwu RI, Atangwho IJ, Egbung GE
, Wokoma MA, Oplekwu RI, Atangwho IJ, Egbung GE
Received 19 March 2025
Accepted for publication 2 August 2025
Published 21 August 2025 Volume 2025:20 Pages 10101—10126
DOI https://doi.org/10.2147/IJN.S529250
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Farooq A. Shiekh
Daniel Ejim Uti,1,2 Wilson Achu Omang,3 Esther Ugo Alum,1 Okechukwu Paul-Chima Ugwu,1 Margaret Amieibi Wokoma,4 Rowland Inalegwu Oplekwu,5 Item Justin Atangwho,4 Godwin Eneji Egbung4
1Department of Research and Publications, Kampala International University, Kampala, Uganda; 2Department of Biochemistry, Faculty of Basic Medical Sciences, College of Medicine, Federal University of Health Sciences, Otukpo, Benue State, Nigeria; 3Department of Medical Laboratory Sciences, College of Health Technology, Calabar, Cross River State, Nigeria; 4Department of Biochemistry, Faculty of Basic Medical Sciences, University of Calabar, Calabar, Nigeria; 5Department of Biochemistry, Faculty of Basic Medical Sciences, Federal University of Allied Health Sciences, Enugu, Nigeria
Correspondence: Daniel Ejim Uti, Department of Biochemistry, Research and Publications, Kampala International University, P.O. Box 20000, Kampala, Uganda, Email [email protected]
Abstract: CD44, a key hyaluronic acid (HA) receptor, has emerged as a central mediator of adipose tissue inflammation, remodeling, and insulin resistance in obesity. Its overexpression in obese adipose depots promotes leukocyte infiltration, pro-inflammatory signaling, and extracellular matrix dysregulation processes that underlie metabolic dysfunction. This review explores the therapeutic relevance of targeting the HA–CD44 axis by synthesizing data primarily from preclinical studies, with emerging evidence from early clinical investigations. A narrative review methodology was employed to assess and compare therapeutic modalities, highlighting advances in molecular targeting, drug delivery systems, and metabolic interventions. We focus on two primary therapeutic classes: small molecules and nanobioconjugates. Small molecules, such as curcumin, metformin, and CD44 antagonists, offer systemic modulation and accessibility but are limited by their low tissue specificity and potential side effects. In contrast, HA-functionalized nanobioconjugates, including liposomes, PLGA nanoparticles, dendrimers, and exosomes, enable targeted delivery to adipose tissue, prolonged drug release, and reduced systemic toxicity. These nanosystems have demonstrated superior modulation of CD44 signaling, adipose inflammation, and glucose homeostasis in obesity models. Emerging strategies such as monoclonal antibodies, GLP-1 analogs, gene-editing tools (eg, CRISPR/Cas9), microbiome modulators, and brown adipose tissue (BAT) activators are also discussed. A comparative analysis indicates that nanobioconjugates offer the highest targeting precision, while small molecules remain advantageous in terms of cost and ease of administration. However, biologics and gene therapies face challenges related to delivery and scalability. Collectively, current evidence predominantly preclinical supports the HA–CD44 axis as a promising therapeutic target in obesity. Integrated approaches combining nanotechnology with molecular inhibitors and biologics could offer a multifaceted strategy for managing metabolic disease.
Keywords: adipose tissue inflammation, targeted nanomedicine, metabolic reprogramming, CD44 isoform-specific therapy, obesity-associated fibrosis
Introduction
Obesity has become one of the most serious public health issues of the 21st century, with the World Health Organization indicating that at present over 650 million adults across the world are obese.1,2 It is a multifactorial and persistent disorder wherein the deposition of adipose tissue becomes excessive, resulting in increased body mass index (BMI) and health consequences, which include type 2 diabetes, cardiovascular types of pathology, non-alcoholic fatty hepatitis, and even cancer.3–5 Obesity is not only a problem in high-income countries, and the two-fold burden of malnutrition and obesity is becoming a problem in low- and middle-income countries.6 The burden of obesity on the health and economy of the world is huge, and it poses significant consequences in healthcare and the productivity of the workforce.7 There is a need to know the underlying mechanism of obesity to design effective therapeutic strategies, given the current high rates of obesity among children and adults.
The factors that are central to the incidence and advancement of obesity are complex interaction of metabolic dysregulation and chronic low-grade inflammation.2,8 In obese individuals, adipose tissue undergoes hypertrophy and dysfunction, as a result, secretion of adipokines, cytokines and chemokines are impaired with further stimulation of systemic inflammation and insulin resistance.8–10 The major molecular mediators, which are recognized tumor necrosis factor-alpha (TNF-a), interleukin-6 (IL-6), and leptin, are highly upregulated that creates an environment favoring lipid accumulation, impaired glucose metabolism, and immune cell infiltration.11,12 These inflammatory cues create a metabolic imbalance that is sustained; hence, obesity is not only a condition of an energy imbalance, rather a condition of molecular and cellular derangements. Pathophysiology of obesity, therefore, requires a line of treatment which works on both the inflammatory and the metabolic aspects of the disease.
The hyaluronic acid (HA)-CD44 axis is one of the many molecular actors involved in obesity, which has caused much interest because of its relationship to cellular interactions, tissue remodeling, and inflammation.13,14 A cell surface glycoprotein, CD44, and the main receptor of the HA is highly expressed in adipose tissue and especially in obese conditions.14 This HA-CD44 interaction regulates the growth of adipocytes, the influx of macrophages, as well as the secretion of cytokines in the adipose microenvironment.15 Interestingly, CD 44 signaling has also been associated with many of the pathologies involved in adipose tissue inflammation, fibrosis, and insulin resistance, highlighting its suitability as a therapeutic target.16 Blocking of HA-CD44 interactions can downregulate the inflammation process and also normalize metabolic status in obese persons.1 Comprehension of this molecular interaction creates new opportunities to do therapeutic interventions to help reduce the complications of obesity.
Due to the shortcomings of the existing therapies in treating obesity with pharmacological and surgical modalities, nanotechnology is building an interesting and new horizon in its treatment.17 The use of delivery systems based on nanoparticles will help increase the bioavailability, stability, and selectivity of anti-obesity drugs in fine-tuning dysfunctional adipose tissues and important molecular circuits like the HA-CD44 axis.18–21 HA-based nanobioconjugates can be used both as delivery agents and as functional biological materials that may react with the CD44 receptor to regulate the signals elicited at the adipose tissue.22 The recent breakthroughs in nanotechnological engineering have enabled the creation of multifunctional nanoplatforms that can accomplish imaging, local delivery, and regulated release of a drug at the same time to enhance therapeutic efficacy and minimize negative side effects.23 The goal of this review is to present the current knowledge regarding HA-CD44 interactions in obesity management, examine the use of nanotechnology in the domain of improving therapeutic results, and compare nanobioconjugate strategies with other molecular and pharmacologic therapies. This review aims at shedding light on newer pathways and technologies in an effort to manage obesity in a better manner in light of a synthesis of recent findings.
Overview of CD44: Structure, Isoforms, and Functional Roles
Structure and Isoforms of CD44
CD44 is a transmembrane glycoprotein, the major receptor of hyaluronic acid (HA), mediating critical cellular adhesion, migration, proliferation, and signaling. One gene on the human chromosome 11p13 is structurally encoded.1,16 The receptor is found in many structural forms; however, through alternative splicing and post-translational modifications, it takes part in a great variety of physiological and pathological processes.24 The CD44 receptor contains three structural parts as indicated in Figure 1 namely the extracellular domain, a single-pass transmembrane domain, and the cytoplasmic tail.24 The extracellular domain has the property of binding to the ligand, especially to HA, although also to other elements of the extracellular matrix (ECM), such as osteopontin, fibronectin, collagen, and matrix metalloproteinases.24 The receptor is anchored in the cell membrane by the transmembrane region and interacts with cytoskeleton and intracellular signalling pathways by the interaction of the cytoplasmic tail with proteins, such as ankyrin and ezrin, radixin moesin (ERM) proteins.
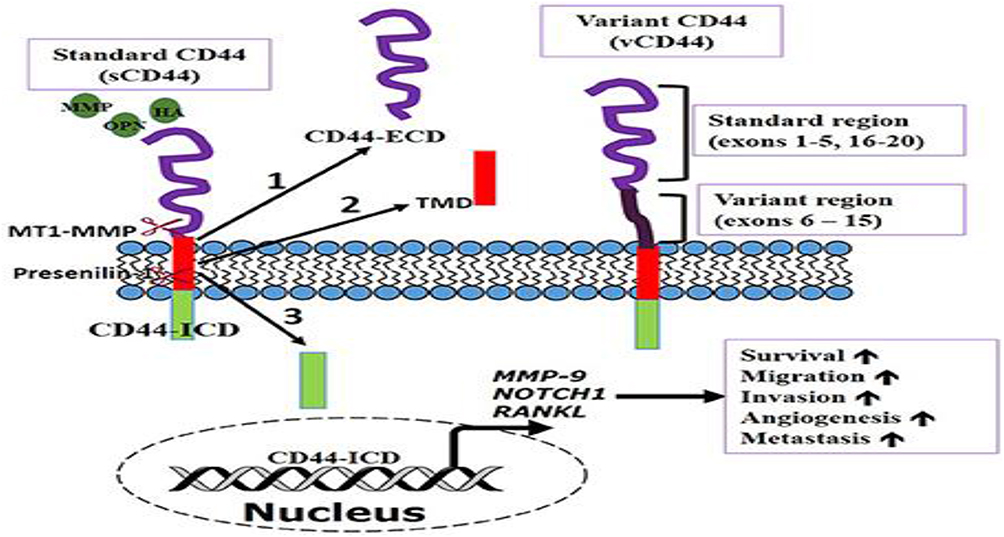 |
Figure 1 CD44 Transmembrane receptor. Adapted from Senbanjo, L.T., Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 5, (2017).25 Under a free open access license. https://creativecommons.org/licenses/by/4.0/. Interaction between CD44 and hyaluronic acid. The CD44 receptor exists in two forms: variant (CD44v) and standard (CD44s). It has four segments: an N-terminal hyaluronic acid-binding loop (BX7B), a stem region, a transmembrane domain, and a cytoplasmic C-terminal domain. Abbreviation: CD44, Cluster of Differentiation 44. |
 |
Figure 2 CD44 Expression and Signaling in Obese Adipose Tissue Microenvironment. Created in BioRender. Basajja, M. (2025) https://BioRender.com/m54hfz5. |
 |
Figure 3 Hyaluronic Acid (HA): Function and Interaction with CD44. Created in BioRender. Basajja, M. (2025) https://BioRender.com/mxfixt8. |
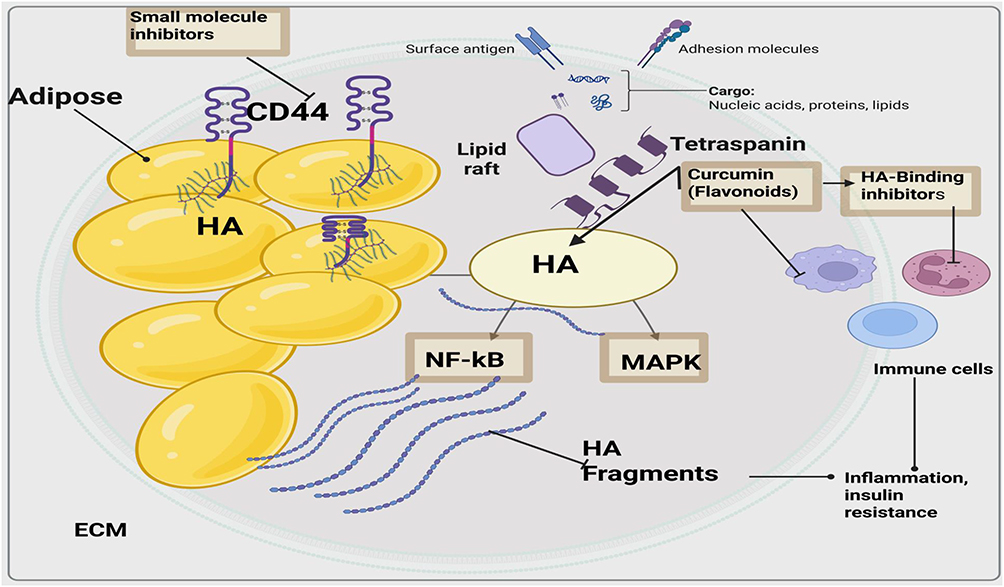 |
Figure 4 CD44 Signalling in Obesity, and small molecules intervention points. Created in BioRender. Basajja, M. (2025) https://BioRender.com/ jdyyys1. |
The diversity of CD44 is also expressed in the effect of 10 variable exons inserted into the extracellular domain, encoding possible alternative splicing to create structural variants of the isoforms.26 Because of this, a standard isoform (CD44s), including the constitutive exons solely, is produced, and several variant isoforms (CD44v) that have one or more of the variable exons added.26 Unlike other variants, the CD44s is expressed ubiquitously in most tissues and is involved in important basal cellular processes such as lymphocyte homing and wound healing.27 On the contrary, the tissue-specific expression and contextual dependence of CD44v isoforms can be found. The variants are especially predominant in epithelial cells and tend to be up-regulated in pathological situations, including inflammation and cancer.1,28 Overexpressed CD44v6 includes exons v6 and v7, interacts with receptor tyrosine kinases, such as MET and EGFR. This interaction improves the motility, invasion, and survival of tumor cells. There are other isoforms like CD44v3, which have Heparan sulfate binding sites, which act to increase growth factor cues that aid in tumor progression.26
The functional diversity of CD44 can be explained by its structurally varied isoforms. These isoforms do not only regulate various physiological processes but also can be used as cancer and other chronic diseases biomarkers and therapeutic targets. The structural peculiarities of CD44 are thus very important towards translational medicine.
CD44 Expression and Activation in Obese Tissues
CD44 is highly distributed among tissues and has several isoforms incurred by alternative splicing.29 Hyaluronic acid (HA) is most prominent and well characterized among its many ligands. A CD44 protein, being significantly upregulated in adipose tissues, has become a focus of investigation not only because of its role in the promotion of constant inflammation, insulin resistance, and metabolic disorders in the obesity epidemic. Obesity is a chronic low-grade inflammatory disease and is marked by adipocyte hypertrophy, inflammatory cell infiltration, and altered extracellular matrix (ECM).5,30 In obese people, as well as in obese animal models, CD44 expression is markedly increased in white adipose tissue (WAT) as well as immune cells (ie, macrophages, T-cells, and stromal vascular fractions).31 The research studies have provided evidence of CD44 negatively associating with body mass index (BMI), leptin, as well as an indicator of systemic inflammation, TNF-alpha and IL-6.32 The presence of an increased CD44 in adipose tissue mediates the inflammatory response by improving the attraction and cycling of inflammational macrophages causing an environment that is predisposed to pro-inflammatory status, which enhances metabolic dysfunction.17
In tissues of obese individuals, the activation of CD44 is mainly due to interaction with HA and other ECM ligands, including osteopontin, fibronectin, among others.33 The HA-CD44 pathway is especially critical to regulating inflammatory signalling routes such as NF-kappa B and JNK that are involved in adipose inflammation and insulin resistance.34 CD44-based signaling also stimulates the expression of molecules associated with adhesion and chemokines that lead to adhesion and migration of leukocytes, maintaining an inflammatory microenvironment in the adipose tissue.35 In addition, the levels and functional activity of CD44 are regulated by metabolic stressors, in particular, hyperglycemia and lipotoxicity.1 These stressors increase synthesis and fragmentation of HA in obese tissues, which causes the production of low-molecular-weight HA, and the fragments are more pro-inflammatory through ligation of CD44.1 This HA-CD44 interaction plays a role in fibrosis of adipose tissue, resulting in fibroblast activation and increased collagen production, which further worsens the plasticity and metabolic homeostasis of adipose tissue.36
The role of CD44 in obesity is pathogenic, as it has been demonstrated in experimental studies.1,16,37 In CD44 knock-out or pharmacological inhibition of CD44 in obese mice decreases macrophage inflammation, and the level of insulin sensitization is enhanced.17,38 These results indicate that CD44 is not simply a marker of the adipose tissue inflammation but also a pathogenic factor of metabolic disorders observed in obesity. CD44 expression and activation significantly increase in obese adipose tissues and serve as the hub to maintain inflammation, immune cell recruitment, and ECM remodelling. The HA-CD44 axis is developed as one of the major regulators that connect the structural remodeling in the tissues with abnormalities in metabolism. The treatment option of CD44 signaling appears to have a future of reducing inflammation and insulin resistance caused by obesity and enhancing better metabolic effects. Accordingly, CD44 can be utilized as a metabolic inflammation biomarker and also can be used as a target of novel anti-obesity strategies.
Figure 2 shows the architectural view of the CD44 protein, including the nature of its structural components and the distinction outlined in the conventional and variant isoforms of the same. CD44 as a form of a type I transmembrane glycoprotein, consisting of three important regions: Extracellular Domain: The extracellular domain is more relevant in binding the ligand, particularly hyaluronic acid (HA), among other components of the extracellular matrix (ECM) such as osteopontin and collagen. The extracellular region of the variant isoforms (CD44v) has extra variable domains, which are encoded by other exons that are alternatively spliced. The variable inserts can fine-tune ligand-binding activity as well as receptor activity. Transmembrane Domain: This is a hydrophobic part of CD44 that fixes in the plasma membrane and functions as a channel through which external signals can travel to the inside of the cell. Intracellular (Cytoplasmic) Domain: The intracellular tail of CD44 binds to cytoskeletal adaptor proteins (ie, the ERM (ezrin-radixin-moesin) complex). This binding makes possible the binding of CD44 to actin filaments, which are key in downstream signaling intracellular events, cell motility, and immune responses. The figure compares, specifically, standard isoforms CD44s that have only the constitutive exons with variant isoforms (CD44v), which have one or more of the variable exons (eg, the v6 isoform, v7 isoform). Such types are mostly upregulated in pathological conditions, such as obesity, inflammation, and cancer, and they are involved in correlated processes, such as immune cell recruitment, tissue remodeling, and signaling transduction. This structural representation is critical in the setting of obesity-associated inflammation due to the emphasis that special microdomains of CD44 mediate pathological activation of the molecule through the interactions with ligands, mainly HA, ultimately resulting in the transmission of both pro-inflammatory and fibrotic messages within the adipose tissue. Figure 2 therefore, gives a background on the multifunctional behaviour of CD44, indicating how CD44 plays an important role in obesity-associated metabolic dysfunction.
CD44 in Adipose Tissue Remodeling, Inflammation, and Insulin Resistance
CD44 operates under a myriad of functions, one of which has recently been pointed out to be adipose tissue remodeling, inflammation, and insulin resistance, which are core elements in the pathophysiology of obesity and metabolic syndrome.17 The role of CD44 in adipose biology is based on the possibility to provide the adhesion of cells, their migration, immune regulation, and those points of contact between cells and the extracellular environment called the extracellular matrix (ECM).39 Remodeling of the adipose tissue is a dynamic process that entails adipocyte hypertrophy and hyperplasia, angiogenesis, extracellular matrix turnover, and infiltration by immune cells.40,41 These remodeling events are dysregulated in the setting of obesity, resulting in excessive deposition of ECM (fibrosis), tissue architecture distortion, and low-grade chronic inflammation.42 CD44 is over-expressed in adipocytes and stromal vascular cells such as pre-adipocytes, macrophages, and endothelial cells. It also interacts with HA and other components of the ECM, and this affects adipocyte differentiation as well as tissue structure.42 Research has demonstrated that there is upregulation of CD44 adipose in obesity, which helps in accumulating ECM and the development of fibrosis.43,44 This fibrotic environment also causes the adipose tissue expandability to be inefficient, which triggers euchromatic lipid preservation and institutional metabolic uneasiness.
Localised inflammation is a trademark of diseased adipose tissue and the key contributor to insulin resistance.2 CD44 plays a critical role in regulating inflammation pathways through the regulation of adhesion of leukocytes and their recruitment into adipose tissue.16 CD44 also promotes the recruitment of pro-inflammatory macrophages (the so-called M1 phenotype) in the adipose tissue during obesity, promoting the production of pro-inflammatory cytokines, eg, tumor necrosis factor-alpha (TNF-alpha) and interleukin-6 (IL-6).45 These cytokines can also interfere with the signaling of insulin on the adipocytes by binding serine kinases to phosphorylate insulin receptor substrates, which can result in impaired uptake of glucose and systemic resistance to insulin. T-cell activation and cell retention in adipose tissue also take place through CD44, which adds to chronic inflammation 2,21
Significantly, CD44 is integrated into adipocyte-immune cell crosstalk by interactions with HA. Amassed fragmented HA during the stress and remodeling of adipose tissue demonstrates an announcement of danger-associated pattern molecule (DAMP), instigating CD44 to initiate inflammatory signaling pathways, such as NF-kB and MAPK.46 This enhances this inflammatory environment, which continues to cause insulin resistance and favour metabolic decline. In mice, CD44 knock-out mice have decreased adipose tissue inflammation, enhanced insulin sensitivity, and dietary obesity resistance, which further confirms the prominent role of CD44 in the process of metabolic inflammation.16 Insulin action is also modulated by CD44 without correlation to inflammation. It either affects the expression of glucose transporter 4 (GLUT4) and insulin receptor substrate-1 (IRS-1), which are important in the process of insulin-mediated glucose uptake.16,47,48 Also, CD44 connections with receptor tyrosine kinases and integrins have the upper hand on the cascade of insulin signaling and influence insulin receptor activation and downstream phosphorylation of Akt.49 Transgression of CD44 distorts these routes, causing ineffective glucose homeostasis.
Therapeutically, the CD44 inhibition or the interaction of CD44 with HA is a viable option to mitigate the effects of obesity. CD44 inhibition with pharmacological agents suppresses the inflammatory and fibrotic state of adipose tissue, increases insulin sensitivity, and increases metabolic normalization in in vitro studies.16,50 Moreover, another form of therapy that could be developed against metabolic diseases is HA-based nanomedicines aimed at inhibiting CD44-mediated signaling. In a nutshell, CD44 plays an important role in adipose tissue modeling, inflammation, and insulin resistance. It manifests and is expressed in the adipose tissue of obese individuals, leading to the remodeling of the ECM, immigration of immune cells, and the breakdown of insulin signaling. The elucidation of the molecular mechanisms behind the functions of CD44 has opened up a new explanation to the pathophysiology of metabolic diseases and makes it possible to develop new effective therapeutic methods for tackling obesity and insulin resistance through the use of CD44 as a target.
Methodology
This article is a narrative review aimed at synthesizing existing knowledge on the role of CD44-hyaluronic acid (HA) signaling in obesity and the potential of HA-based nanobioconjugates as therapeutic agents. The review was guided by a thematic focus rather than a rigid systematic protocol, allowing for a broader, more flexible integration of relevant findings from diverse sources. To construct this review, we engaged with a wide array of scientific literature, including original research articles, reviews, preclinical studies, and theoretical discussions published in peer-reviewed journals. Emphasis was placed on recent advances, but foundational studies and landmark contributions were also considered to provide historical and conceptual context. The selected literature covers areas such as CD44 structure and isoforms, HA signaling mechanisms, obesity-associated inflammation, nanomedicine, and targeted drug delivery. Sources were identified through iterative exploration of academic journals, reference lists, and recognized scientific repositories. The aim was to highlight key mechanistic insights, therapeutic innovations, and comparative perspectives that connect CD44 signaling with metabolic dysregulation and emerging treatment modalities in obesity. This flexible, narrative approach allows for critical reflection on trends, knowledge gaps, and future directions, making it well-suited to capture the evolving and interdisciplinary nature of the topic.
Hyaluronic Acid (HA): Function and Interaction with CD44
Hyaluronic acid (HA), originally referred to as hyaluronan, is a large-sized glycosaminoglycan that has widespread locations across connective, epithelial, and nervous tissue.51,52 It consists of a similar disaccharide repeating unit, both D-glucuronic acid and N-acetyl-D-glucosamine combined through alternating 1,4 and 1,3 glycosidic interactions.17 Due to its distinct physicochemical capabilities and biological activities (Figure 3), HA has major roles in tissue hydration, structural support, signaling, and repair.17 Another important feature of the HA bioactivity is its binding to the plasma membrane receptor, CD44, which is a multifunctional transmembrane, that causes adhesions, migration, and the immune response.13 HA-CD44 bonding coordinates an immense number of cellular and physiological processes, especially in inflammatory, wound healing, cancer development, and even metabolic diseases.38
Architecture and Location of HA Structural Features and Distribution of Hyaluronic Acid
HA also contains no sulfation, and its linear structure is depicted in Figure 3 which is in contrast to other glycosaminoglycans, including heparan sulfate or chondroitin sulfate.53 Its metallic, unbranched architecture and its ability to bind water to itself help the HA form hydrated extracellular matrices that offer a mechanical scaffold and facilitate the diffusion of substances with the exchange of nutrients and wastes.53 The production of HA is dominated by three isoforms of hyaluronan synthases (HAS1, HAS2, and HAS3) in the plasma membrane.54 HA polymers are in the oligosaccharide to the high-molecular-weight (HMW) chains greater than 106 Daltons. The functional significance of this difference in MW is that HMW-HA tended to be anti-inflammatory and immunosuppressing since low-molecular-weight (LMW) fragments typically served as pro-inflammatory markers.55 Physiologically, HA is found in large amounts in the vitreous body of the eye, synovial fluid of the joints, umbilical cord, the dermis, as well as the interstitial spaces of the epithelial tissue.55 Hormonal balance between the production of its synthesis by HAS enzymes and the degradation of hyaluronidases immediately balances its concentration and molecular weight.17 Such enzymes act to degrade HA into smaller active fragments, allowing HA to code in a wide array of cellular signaling pathways (Figure 3).
The Biological Functions of Hyaluronic Acid
Because of its viscoelastic, lubricating, and signaling capability, HA can perform multifaceted biological functions. It helps to provide the connective tissues with mechanical strength as well as hydration; it is vital in the lubrication of joints as well as the elasticity of the skin.56 HA is used in most dermatological cosmetic products and wound healing preparations since in the dermis, it facilitates the proliferation of fibroblasts and synthesis of collagen. In the immune system, HA regulates the trafficking of leukocytes, antigen presentation, as well as release of cytokines, and these effects are highly dependent upon the molecular size.56 HA is essential at every phase of wound repair: it is known to facilitate blood clotting during the early stages, accelerating proliferative and migratory processes of keratinocytes, as well as angiogenesis and extracellular matrix remodeling with regard to the later phases.17,56 In addition, the HA has an effect on embryogenesis in which cell migration and tissue morphogenesis are controlled during the development of the heart and the neural tube. In cancer biology, it has also been implicated in its anti-apoptotic, mitogenic effects that facilitate the growth and other factors in cancer biology.57 HA production and signaling become abnormal and increasing cell growth and metastasis.
CD44: The Leading HA Receptor
The main cell surface receptor of HA is CD44, which is highly expressed in many cell types, such as epithelial cells, fibroblasts, lymphocytes, as well as cancer stem cells.1 Functionally, CD44 has an extracellular region, a transmembrane domain, and a cytoplasmic domain that connects with the intracellular signals and cytoskeleton. Pre-mRNA of CD44 is alternatively spliced, expressing isoforms (CD44 standard or CD44s, and variant forms CD44v), differing in extracellular domain content, and are therefore variably bound to ligands and perform different roles.16,50 HA-CD44 interaction is one of the key mediators of cell-matrix adhesion, cell migration, and signal transduction.1 When bound by HA, CD44 forms oligomers and interacts with accessory proteins and signal transduction complexes, stimulating other downstream signaling targets including Rho GTPases, PI3K/Akt, MAPK/ERK, and NF-kB.58 These cascades modulate the cytoskeletal rearrangements, cell proliferation, survival, as well as inflammatory response. CD44 and HA interactions in the immune system mediate leukocyte rolling, adhesion, and extravasation in inflammatory reactions 59
Cellular Functions of the HA-CD44 Interaction
There is a crucial involvement of the HA-CD44 axis in both pathological and physiological situations. HA is produced within the tumor microenvironment, and binding of HA to CD44 acts in cancer to enhance the epithelial–mesenchymal transition (EMT; tumor cell motility) as well as cancer stem cell maintenance.38 Robust CD44 expression is usually associated with malignant tumor phenotypes and indifference to treatments. In this regard, HA-CD44 signaling supports a pro-survival niche, by activation of Akt and ERK, which promotes chemoresistance and metastatic ability.38 The interactions between HA and CD44 in the adipose tissue magnify inflammation and insulin resistance in metabolic diseases, including obesity and diabetes type 2. The CD44 expression of adipocytes and macrophages is increased in obese conditions and adsorption of HA contributes to inflammatory cell infiltration and release of cytokines.16,58 Inhibition of the HA-CD44 pathway, has therefore, become one of the promising ways of reducing the metabolic inflammation and enhancing insulin sensitivity. In addition, HA-CD44 interactions in regenerative medicine are exploited in tissue engineering and development of drugs.60 The protein based HA gels can be synthesized to imitate the extracellular matrix and release therapeutic agents or cells to the target tissues.60 Defined targeting of nanocarrier systems to CD44 has to date demonstrated preclinical promising results, especially in cancer where CD44 is overexpressed on cancer cells.60
Hyaluronic acid (HA) is an essential marquee of the extracellular matrix that has varied and various biological functions over encoding, structure, and cellular signaling. Its binding with the CD44 receptor is one of the major mechanisms by which HA modulates the behaviour of cells in health and disease. As shown in Figure 3, the fact that HA is relatively simple in structure is deceptive of the complicated and far-reaching implications of HA in tissue biology. HA-CD44 axis is centralized to pathways of immune response, tissue regeneration, cancer pathogenesis, and metabolic deficiency. The interaction has the promise of providing insight into therapeutic breakthroughs in so-called regenerative medicine, oncology, and longstanding inflammatory states.
Recent Advances in Small Molecules Targeting CD44-HA Signalling in Obesity
CD44 hyaluronic acid (HA) signaling pathway is important in the pathophysiology of obesity especially adipose tissue inflammation, fibrosis, and metabolic dysfunction.13 It interacts with HA and especially low-molecular-weight HA (LMW-HA)), leading to macrophage and T-lymphoid cell recruitment, pro-inflammatory activations as well as insulin resistance.13 Inhibition of the pathway depicted in Figure 4 is a potential therapy against obesity and its consequences (Figure 4). A schematic representation of CD44-HA signaling pathway in adipose tissue of the obese is provided in Figure 4 and indicates multiple intervention points that may be targeted using small molecule inhibitors. By targeting this axis, the inflammatory infiltrates into tissues can be regulated, chronic inflammation can be reduced, and metabolic homeostasis can be regained.
Recent advances in small molecule therapeutics have resulted in appearances of a number of agents that can interfere with CD44-HA interactions, or downstream signaling.61 Direct inhibition of the HA-binding domain on CD44 is one of the big means. Sulfates hyaluronal-type analogs and HA mimetics are such compounds, which act as competitive HA blockers, down-regulating pro-inflammatory signaling pathways by inhibiting NF-kappa and the mitogen-activated pathways, leading to a decreased number of released cytokines and enhanced insulin responsiveness.61 Redesigned repurposed oncology agents (example RG7356 — designed to target CD44-abundant tumors) have emerged as having promise in countering metabolic inflammation via their CD44+ immune cell removal and interruption of HA-mediated activation of immune cells.62 Using mice obesity models, RG7356 administered still showed lowered macrophage infiltration and enhanced glucose metabolism.63,64
Natural small molecules have a CD44-modulatory effect, especially those bioactive phytochemicals like curcumin, epigallocatechin gallate (EGCG) and glycyrrhizin. These mediators have been known to suppress CD44 expression levels, to disrupt HA-CD44 reactions, and to downregulate downstream inflammatory responses.65–67 As an example, curcumin lowers the expression of CD44 mRNA and inhibits the adhesion and migration of the cells using HA within the inflamed adipose tissue.66,68 The pace of discovery of new inhibitors has been further boosted by the new advances in computational drug discovery. There has been the identification of high-affinity ligands of the HA-binding site of CD44, via computer-aided screening and molecular docking protocols, and good pharmacokinetics for development.69,70 One new approach that is becoming popular is that of HA-conjugated drug delivery, whereby an overexpression of CD44 is harvested and used to deliver drugs in order to specifically zero in on medications that have anti-inflammatory properties, at the same time.71 HA-modified nanoparticles that target inhibitors or enzyme activity or anti-inflammation can specifically accumulate in CD44 inflammatory tissue, therefore, improving the antitumor activity and reducing general toxicity.71 Along with these innovations, there have been issues with regards to optimizing the specificity, bioavailability, and the safety of these small molecules. In addition, the complexity of the structural organization and the variety of isoforms of CD44 require specific strategies depending on tissue-specificities 72
Hence, the appearance of small molecule inhibitors of CD44-HA signaling is an important breakthrough in obesity medical treatment. Combined application of synthetic compounds, phytochemicals, and specific delivery vehicles has provided a potential opportunity to combat obesity-related adipose tissue inflammation and to normalize metabolism, and these have been summarized in Table 1.
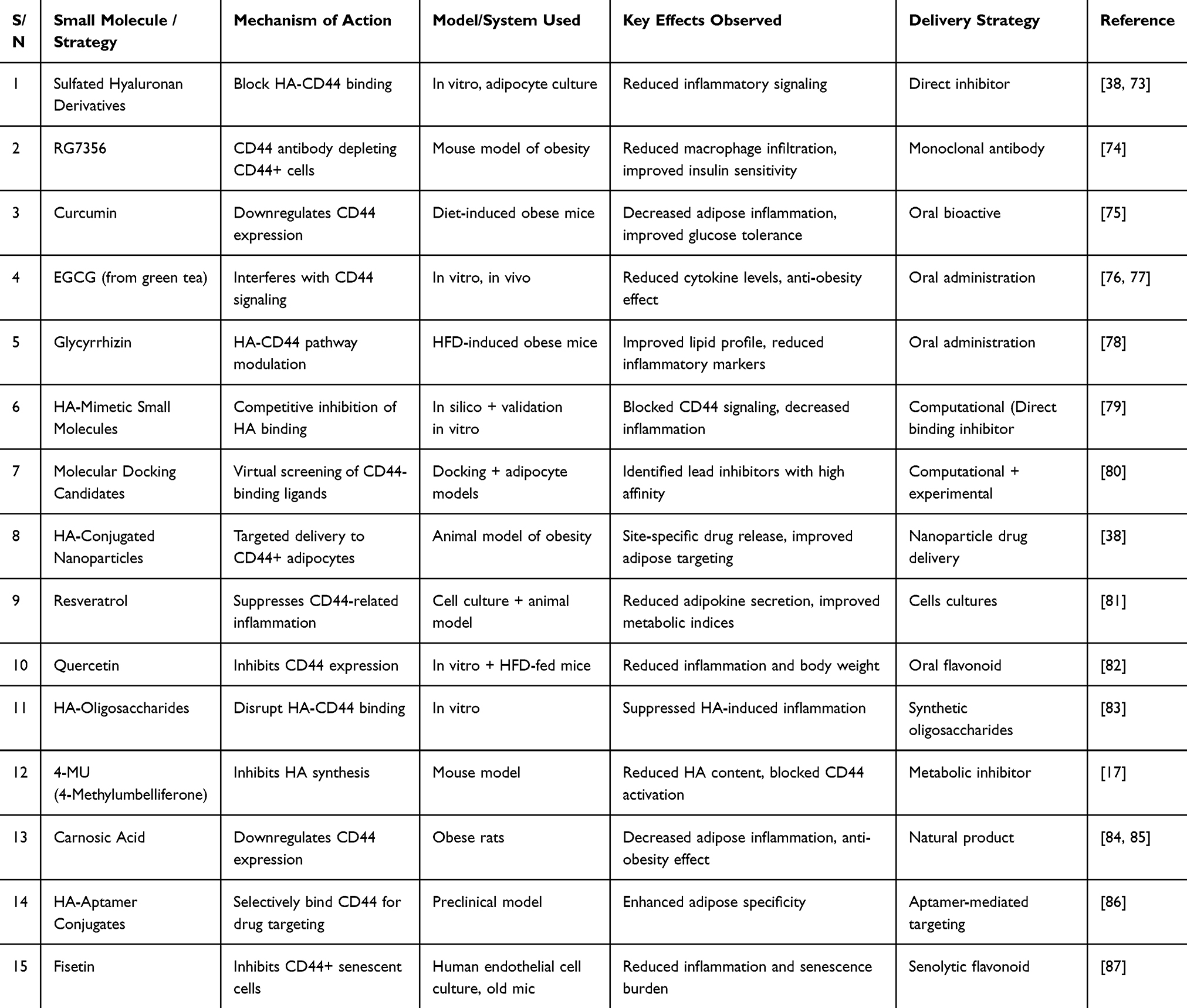 |
Table 1 Summary of Recent Advances in Small Molecules Targeting CD44-HA Signaling in Obesity |
Challenges with Small Molecules Targeting CD44-HA Signaling in Obesity
Despite the promising therapeutic potential of small molecules in modulating CD44-hyaluronic acid (HA) signaling to combat obesity and its associated metabolic complications, several key challenges hinder their clinical application and translational progress.
Structural Complexity and Isoform Specificity of CD44
CD44 exists in multiple isoforms generated via alternative splicing, including the standard form (CD44s) and variant isoforms (CD44v).88 These isoforms exhibit distinct expression profiles and biological functions in different tissues and cell types. Designing small molecules that selectively target pathologically relevant isoforms without disrupting the physiological functions of CD44 poses a significant hurdle. Isoform-specific structural data remain limited, complicating structure-based drug design.
Redundancy and Compensation in HA Receptors
While CD44 is a major HA receptor, other receptors such as RHAMM (receptor for HA-mediated motility) and LYVE-1 also bind HA and can compensate for CD44 inhibition.89 This redundancy in HA signaling pathways can reduce the efficacy of CD44-targeted interventions. Blocking CD44 alone may be insufficient to fully modulate HA signaling in adipose tissues.
Context-Dependent Role of CD44 in Obesity
The function of CD44 in adipose tissue is context-dependent and varies with obesity stage, fat depot, and systemic metabolic status. Inflammation-induced upregulation of CD44 enhances HA binding and promotes macrophage recruitment and fibrosis.90 However, basal CD44 expression may have homeostatic roles in tissue remodeling. Targeting CD44 indiscriminately could lead to unintended effects on immune surveillance or tissue repair.90
Limited Bioavailability and Stability of Small Molecules
Many small molecules targeting CD44 or interfering with HA binding suffer from poor pharmacokinetic properties, such as low oral bioavailability, rapid metabolism, and limited tissue penetration.17,91,92 These pharmacological limitations restrict their in vivo efficacy and increase the need for frequent dosing or nanocarrier-based delivery systems.
Off-Target Effects and Safety Concerns
Because CD44 is ubiquitously expressed in multiple organs and immune cells, small molecules that inhibit CD44-HA interactions risk causing systemic immune dysregulation, impaired wound healing, or hematopoietic dysfunction.38,93 Achieving target specificity while minimizing adverse effects remains a key challenge in drug development.
Incomplete Understanding of Downstream Signaling
CD44-HA signaling activates multiple downstream pathways, including PI3K/AKT, MAPK, and NF-κB, which regulate cell proliferation, inflammation, and metabolism.94 However, the exact molecular cascades specific to obesity-induced adipose dysfunction are still being elucidated. Without precise knowledge, designing pathway-specific inhibitors is difficult.
Translational Gaps From Preclinical to Clinical Models
Most data on CD44-HA inhibition come from in vitro studies or rodent models, which do not fully recapitulate the complexity of human obesity. Differences in CD44 expression patterns and immune responses between species hinder translation. Additionally, long-term safety and efficacy data in human obesity are lacking.1
Recent Advances in Nanobioconjugates Targeting CD44-HA Signalling
Recent advances in nanotechnology have produced nanobioconjugates constructed of engineered nanomaterials conjugated with biological ligands, and these compounds have transformed targeted drug delivery and molecular imaging. Nanobioconjugates against CD44-HA signaling make use of the specific interaction between HA and CD44 and enable these specific nanocarriers to deliver therapeutic or diagnostic cargo to cells that express high levels of CD44, thus reducing any systemic effects.95 Such nanosystems are liposomes, micelles, dendrimers, metal-organic frameworks (MOFs), mesoporous silica nanoparticles, exosomes, and polymeric nanoparticles, which are commonly functionalized with HA or CD44-specific antibodies (Table 2).17,96,97
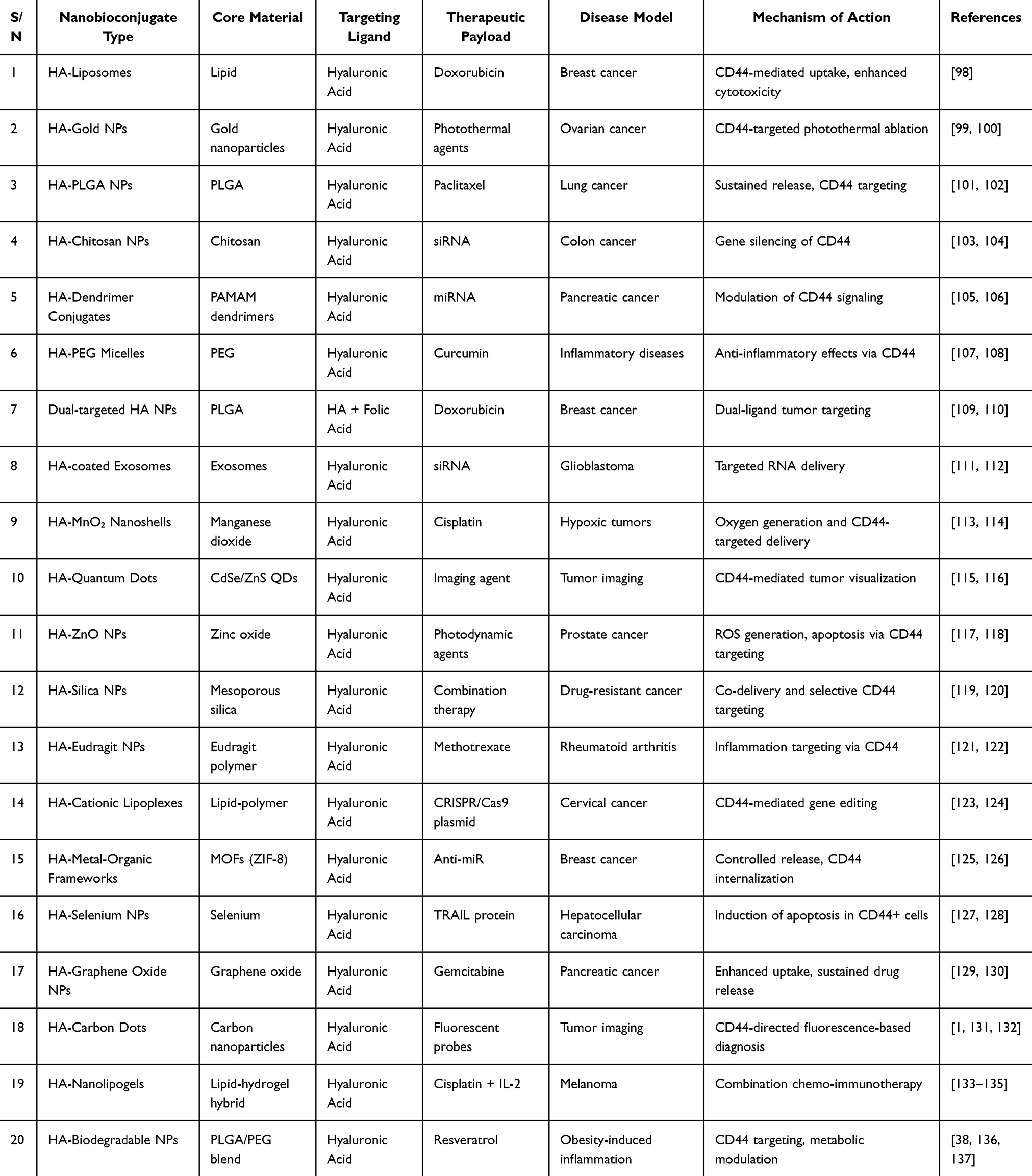 |
Table 2 A Summary of Recent Advances in Nanobioconjugates Targeting CD44-HA Signalling |
The most studied nanocarriers include derivatives of HEPES acrylate (HA) decorated liposomes and polymeric nanoparticles.23 HA-coated liposomes that contain doxorubicin or paclitaxel have been found more effective in the receptor-mediated endocytosis and accumulation of drugs both inside the cell and into the CD44-abundant tumours.17 Correspondingly, HA-conjugated PLGA (poly(lactic-co-glycolic acid)) nanoparticles have demonstrated regulated drug release and improved tumor accumulation in vivo. Besides administration of chemotherapeutics, HA-nanobioconjugates have been found useful in administering nucleic acids, including siRNA, miRNA, and CRISPR/Cas9, to silence CD44 or its downstream effectors.138,139 The method leads to the disruption of key tumorigenic pathways, halts the tumor growth, and makes the tumor more responsive to standard treatments. As an example, dendrimer and cationic liposome conjugated to HA deliver miRNA mimics and inhibitors that may allow selective manipulation of oncogenic signal transduction pathways in pancreatic, breast, and lung cancer.140
Smart nanocarriers have also led to innovations that have improved the efficiency of therapeutic precision.141 Nanoresponsive nanobioconjugates release their cargo in response to some specific environmental factor, such as pH, redox potential, or enzyme activity, and have been shown to work well in hypoxic tumor microenvironments.141 case in point is with HA-capped manganese dioxide (MnO 2) nanoparticles, which target the CD44 positive cells and, in addition, mitigate tumor hypoxia by producing oxygen to enhance combination remedies like photodynamic therapy.142 Coupled targeting approaches are also becoming common where nanocarriers become co-functionalized both using HA and a separate targeting reagent, eg, folic acid, transferrin, or RGD peptides.17 These designs increase selectivity and internalization in mixed tumors. Together with that, the use of nanocarriers, which introduce several drugs simultaneously, with the examples of using chemotherapy in combination with immunomodulators or antioxidants, provides a synergetic effect and lowers the resistance to drugs.
Nanobioconjugates based on CD44-HA have more applications beyond drug/gene delivery and have entered the area of diagnostics.143 The fluorescence, photoacoustic, and PET imaging of the CD44 overexpressing tumors have been imaged using quantum dots, gold nanoparticles, and upconversion nanoparticle functionalized with HA.144,145 These diagnostic nanoconjugates give information concerning tumor localization, stage, and therapeutic response. Although these achievements are outstanding, there are still a few challenges that should be overcome prior to clinical translation.62 These include stability, scalability, biodegradability, and pharmacokinetics. In addition, long-term immunogenicity and off-target effects should be improved through rigorous preclinical studies.98 Still, as nanomaterial engineering, ligand design, and delivery methodologies advance continually, CD44-HA-targeting nanobioconjugates hold a promising avenue of precision medicine. Though current applications are mostly associated with cancer, these give an idea and knowledge of its application to obesity, and management of these diseases.
Comparative Analysis of Small Molecules vs Nanobioconjugates, and other Therapeutic Targets and Actors in Obesity
The treatment of obesity has changed and evolved past lifestyle change and bariatric surgery to small molecules and nanobioconjugates. New directions also include the emerging players, such as peptides, monoclonal antibodies, gene therapies, and gut microbiota modulators.
The Use of Small Molecules in the Treatment of Obesity: Low molecular weight compounds (<900 Da), that can either bind a protein or a receptor and modulate cellular pathways, are known as small molecules.146 They are good because of oral bioavailability, systemic distribution, and affordability. Small molecules in the management of obesity include targeting signaling processes, and these ways include AMPK activation, PPARgamma modulation, inhibition of lipogenesis, and satiety.147 Orlistat (pancreatic lipase), liraglutide (GLP-1 receptor agonist), and metformin (AMPK activator) representatives are clinically approved.148 Nevertheless, the disadvantages of the small molecules are the off-target effects, poor tissue specificity, and drug resistance.147
Nanobioconjugates: Accuracy and Virtuosity: The nanobioconjugates are complex engineered nanostructure conjugates that exhibit site-specific delivery and controlled release with targeting ligands, antibodies, peptides, or drugs.149 Nanobioconjugates, by contrast to small molecules, have the ability to package several therapeutic agents, increase the pharmacokinetics, and deliver inside the cell.150 In obesity treatment, liposomes, polymer-based nanoparticles, and dendrimers are functionalized with adipose targeting molecules (eg, CD44, HA, or integrins).17 Such nanosystems enable the modulation of adipogenesis, inflammation, and insulin sensitivity with low systemic toxicity.
Therapeutic Actors Beyond Small Molecules and Nanobioconjugates: Monoclonal antibodies and peptides present a high level of specificity but a lower immunogenicity.151,152 Peptide-based therapies are aimed at hunger signalling modulation and energy expenditure, including GLP-1 analogs, amylin mimetics, and leptin sensitizers.153 Anti-inflammatory effects of monoclonal antibodies to pro-inflammatory cytokines (such as TNF-α and IL-6) assist in reducing the inflammatory effects of obesity. There are gene editing reagents: CRISPR/Cas9 and RNA interference (siRNA, miRNA), that have potential but are hampered by delivery and efficacy over longer expression times.35 Also, one of the emerging strategies in metabolic reprogramming in obesity is the modulation of microbiota using probiotics, prebiotics, and fecal transplantation 154,155
Though small molecules are low-cost and easily accessible, they have a disadvantage because of their poor targeting capacity and increased side effects. Nanobioconjugates are characterised by higher precision, greater efficacies, and possibilities of combinatorial therapies, although manufacturing problems and regulatory issues are still a concern. Other therapeutic players, such as biologics and gene therapy, offer futuristic advantages but experience problems in delivery, stability, and cost. A comparative presentation on Small Molecules, Nanobioconjugates, and Other Therapeutic Targets and Actors in Obesity is presented in Table 3.
 |
Table 3 Comparative Summary of Small Molecules, Nanobioconjugates, and Other Therapeutic Targets and Actors in Obesity |
Potential Off-Target and Systemic Consequences of CD44-HA Signaling Blockade
Although the treatment of CD44-hyaluronic acid (HA) signaling through its therapeutic targeting is a promising method in the obesity management and, more so, attenuation of adipose tissue inflammation, hyperinsulin sensitivity, and refinement of metabolic profiles, it would be important to note the effects of inhibiting signaling on the broader range of physiology. CD44 is a highly expressed transmembrane glycoprotein present in various biological processes and different tissues and body systems.184 Through its binding with HA, which is a major component of the extracellular matrix, it controls cellular expression of a number of functions such as adhesion, proliferation, migration, and tissue repair.143
Because CD44-HA signaling inhibition is systemic, it may have undesirable effects outside the adipose tissue. CD44 is important in the activation of leukocytes, in the homing of leukocytes, and in the migration of leukocytes in the immune system.35,185 The interruption of this activity can cause impaired immune activities, susceptibility to infections, and uncontrolled inflammatory responses, which predispose people to autoimmune diseases.2,185 Epithelial integrity and wound healing are maintained by CD44-HA signaling in the epithelial cells of the skin, gastrointestinal tract, and the lung. Inhibition of this pathway may interfere with regeneration, slow healing, and predispose to tissue injury or fibrosis.186 Moreover, the CD44 is also present in the synovial lining of joints, and its interaction with HA helps in providing cartilage homeostasis as well as joint lubrication.187 When these protective mechanisms are inhibited over a long period, they can result in joint stiffness or degenerative joint problems. In addition, CD44 has been shown to play functions in the regulation of hematopoietic stem cells and maintenance of the stem cell niche. Long-term inhibition of CD44 may affect normal stem cell functionality, hematopoiesis, or disrupt progenitor cell replenishment.187,188
Considering these facts, all therapeutic interventions against CD44-HA signaling in obesity should consider tissue-specific and systemic functions of CD44. Precision ways to reduce off-target are necessary. They may incorporate fat-specific methods of delivery (eg, nanoparticles or liposomes that accumulate in tissue-selective matter), inducible or temporary methods of gene silencing, or the administration of ligand-blocking antibodies that are highly specific towards pathogenic CD44 isoforms (eg, CD44v) responsible for inflammation of obesity.
A combination of such strategies can be used to maintain the physiological functions of CD44-HA signaling in non-target tissues and to combat the pathological functions of it in adipose tissue successfully. This is a balance required in the creation of secure and effective therapies. Therefore, although CD44-HA signaling blockade has significant potential as an obesity and metabolic complications therapeutic approach, it is imperative to bear in mind that it has some systemic roles to play. The inclusion of selective delivery systems and assessing the long-term safety will play a critical role in introducing this form of treatment into clinical practice.
Role of Hyaluronic Acid (HA) Fragments in Tissue-Specific Contexts
Molecular weight is a strong determinant of the biological activity of hyaluronic acid (HA), with high molecular weight (HMW-HA, >1,000 kDa) and low molecular weight (LMW-HA, <500 kDa) fragments having stereotypically different effects in diverse tissues and in diverse pathological scenarios, and frequently antagonistic effects in the same tissue subjected to distinct pathological processes.189,190 HMW-HA is usually related to homeostatic and anti-inflammatory processes. It helps in tissue hydration and structural integrity as well as immune quiescence.190 Conversely, LMW-HA fragments, which are usually produced during tissue damage or inflammation, may act as danger-associated molecular patterns (DAMPs) that induce activation of an inflammatory signaling cascade through Toll-like receptors (TLR) and CD44.190
Recently, in adipose tissue, an example of how LMW-HA is causative in the promotion of inflammation, macrophage infiltration, and insulin resistance, and thus metabolic dysfunction as has been reported in obesity.191 On the other hand, HMW-HA seems to have protective properties as it stabilizes the extracellular matrix (ECM) and has anti-inflammatory properties. At the tumorigenesis level, LMW-HA fragments could promote tumor development through promotion of angiogenesis, cell motility, and/or cytokine production, whereas HMW-HA can serve as a physical barrier to cell migration and invasive cell spreading in the early stages of tumor development.191
The function of HA fragments is context-dependent, and the use of the HA fragment is also dichotomous, as it depends on the context in which they are used. Furthermore, complicating the interpretation of HA signaling outcome is also the tissue-specific expression of HA synthases (HAS13) and hyaluronidases (HYAL1 2) and the varied receptor interactions (CD44 variants, RHAMM, and TLRs).192 Hence, HA signaling-mediated therapeutics require prioritizing development strategies towards size-specificity and tissue of choice application of enzymatic remodeling of HA, using HA mimetic therapies, or the conjugation of a drug with HA.
Preclinical and Emerging Clinical Evidence: in vivo and in vitro Models Using HA-Nanobioconjugates in Obesity
Preclinical and emerging clinical data support the potential of hyaluronic acid (HA)-based nanobioconjugates as novel therapeutics in obesity and metabolic disorders. HA, due to its biocompatibility, biodegradability, and specific binding affinity to CD44 receptors overexpressed in inflamed adipose tissues, serves as an efficient carrier for targeted drug delivery.193 Various in vitro and in vivo studies have utilized HA and its nanoconjugated derivatives to modulate adipogenesis, inflammation, insulin resistance, and lipid accumulation 179,194
In vitro models such as 3T3-L1 adipocytes and human adipose-derived stem cells have been employed to assess the anti-adipogenic, anti-inflammatory, and insulin-sensitizing effects of HA-drug conjugates, including those loaded with metformin, resveratrol, or siRNA targeting CD44.179,195. These nanobioconjugates have demonstrated enhanced cellular uptake, reduced lipid droplet formation, and downregulation of pro-inflammatory cytokines compared to non-targeted therapies.
In vivo models, especially high-fat diet-induced obese mice or Zucker diabetic fatty rats, have further validated the efficacy of HA-based nanocarriers.196–198 Results show significant reductions in body weight, visceral fat, systemic inflammation, and improved glucose tolerance. Moreover, HA conjugates showed prolonged circulation, targeted accumulation in inflamed adipose tissue, and minimal off-target toxicity.199 Although clinical trials remain limited, early-phase evaluations indicate favorable pharmacokinetics and biosafety. Further comparative studies and long-term follow-up are necessary to validate translational success. Table 4 presents a comparative summary of select studies that highlight preclinical and clinical advancements in HA–nanobioconjugate therapies in obesity.
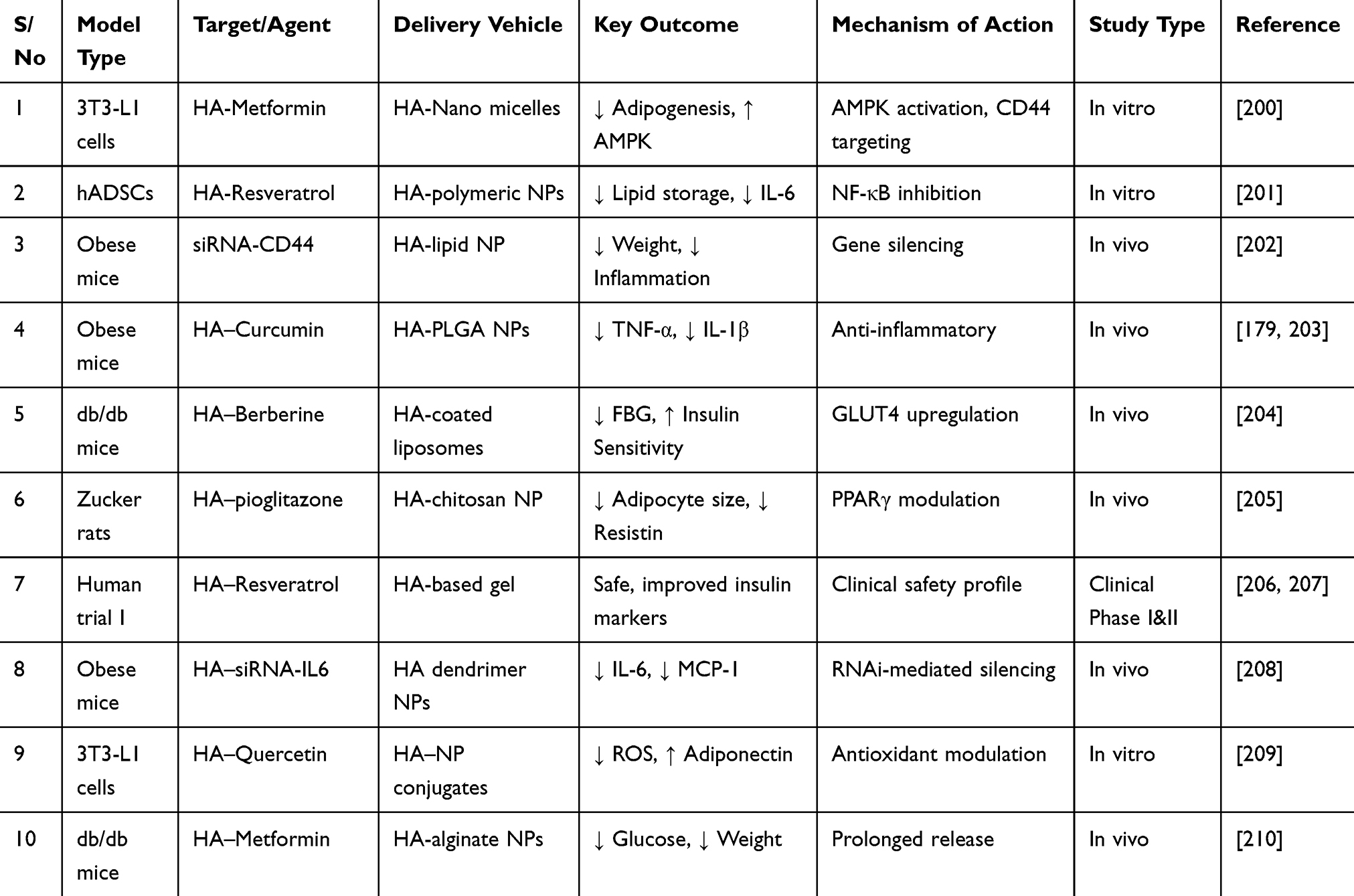 |
Table 4 Comparative Summary of Preclinical and Emerging Clinical Evidence Using HA–HA-Nanobioconjugates in Obesity |
Clinical Relevance of Targeting CD44–Hyaluronic Acid (HA) Signaling in Obesity: Small Molecules, Nanobioconjugates, and Emerging Therapies
Developments on obesity and improvement on the CD44 targeting hyaluronic acid (HA) signalling axis have become valuable information in the fight against obesity-related inflammation and abnormal metabolism.211 There is interest in cheap small-molecule inhibitors that regulate this pathway to inhibit adipose-tissue pro-inflammatory responses.91,212 These agents mediate by disrupting the CD44-HA attachment as well as inhibiting the expression of the CD44 and downstream effectors like NF-KB, JNK. Some compounds, such as flavonoids like quercetin and luteolin, and sulfated HA derivatives, have proven anti-inflammatory and insulin-sensitizing mechanisms in the preclinical models.65,212 Despite remaining primarily in the experimental phase, the small molecules present an opportunity to achieve a pharmacologically feasible and scalable way of reducing adipose tissue inflammation and bringing the metabolism of obese individuals back to balance.
Nanobioconjugates offer the next generation of precision targeting and treatment of inflammation of adipose tissue by targeting CD44-expressing cells. CD44-overexpressing adipocytes and macrophages may be targets of selectively delivering the therapeutic agents using engineered nanoparticles, including liposomes functionalized with HA, polymeric carriers, and dendrimers.150,213 Such systems enhance the solubility of drugs, promote tissue specificity, and minimize off-target toxicities. And recent experiments have also indicated that HA-coated nanocarriers loaded with any kind of anti-inflammatory agent possess the capability of decreasing the inflammation and insulin resistance in the case of obese models.214 Also, the RNA-based payloads in the form of siRNAs against CD44 or other inflammatory mediators have been delivered successfully via these nanoplatforms, with demonstration of encouraging results in experimental obesity.
New treatments also increase the CD44-HA axis of targeting obesity. CD44 monoclonal antibodies are being investigated to ensure interference with downstream pro-inflammatory signaling. In the same vein, the high specificity and stability of peptidomimetics and aptamers that are specifically supposed to inhibit HA binding were also observed.211 There is also the use of gene silencing methods such as the antisense polynucleotide and CRISPR-based approaches to selectively suppress CD44 expression in adipose tissue.215 Moreover, some bioactive dietary compounds and microbiota-focused, indirectly interceded interventions may exert an informative impact on CD44 signaling and provide adjunctive advantages. Though these novel therapies are still in their development phases, the possibility of their synergy with known metabolic therapies is an important clinical implication for the ability to be used as a CD44 HA pathway therapy target in obese patients.
Summary
This review provides an in-depth overview of the CD44-Hyaluronic Acid (HA) signalling axis in obesity as well as the new targeted–based therapeutic strategies to modulate it. CD44, which is a transmembrane glycoprotein, binds to HA to modulate the adipocyte function, invasion of immune cells, extracellular remodeling, and insulin signaling. The axis dysregulation substantially adds to adipose tissue inflammation, hypertrophy, fibrosis, and systemic metabolic aberrations that are typical of obesity and obesity-related conditions, type 2 diabetes, and cardiovascular disease, among others.
The two major therapeutic approaches reviewed are the small-molecule inhibitors and nanobioconjugates. The ease of production, oral bioavailability, and low cost of production are mentioned as advantages of small molecules such as the CD44 antagonists, hyaluronidase inhibitors, and HA biosynthesis modulators. Nevertheless, they are usually restricted due to systemic toxicity, lack of specificity, and development of drug resistance. Nanobioconjugates, however, represent a next-generation solution by engineering nanoparticles that can then be conjugated to biomolecules such as HA that would allow delivery of a drug to cells that overexpress the CD44 protein (targeted delivery), sustained release, and diminished off-targeting. Due to their high precision and efficacy, preclinical models reveal the superior performance of these nanocarriers, but there is still a hurdle to overcome: large-scale production, the immune barrier, and regulatory bottlenecks.
Noteworthy, the review also presents a comparative framework in Table 3, in which strengths, limitations, targeting strategies, and clinical status of small molecules, nanobioconjugates, and other therapeutic actors towards treating obesity have been outlined. The comparative analysis extends the horizon of the review by placing the approaches to CD44 in the context of an overall therapeutic environment of intervention, which conceives of things like dietary regulation, anti-inflammatory agents, adipokine modulators, gene-editing approaches, and so on. In sum, the review highlights a promising therapeutic potential of targeting the CD44HA signal pathway, including the combination of nanotechnology and molecular inhibitors, and further researches should focus on complex and personalized approaches, which are consistently striving to find the shape of the emerging approaches to obesity as a multifactorial metabolic condition.
Future Perspectives and Directions
The CD44–HA signaling pathway presents a compelling and multifaceted target for obesity intervention. Looking ahead, several strategic directions can elevate the translational potential of CD44-targeted therapies:
Personalized Therapy and Biomarker Development
Future research should focus on identifying and validating biomarkers linked to CD44 expression and HA metabolism across various adipose depots (WAT, BAT, VAT). These biomarkers can guide personalized therapy for obesity subtypes and monitor therapeutic efficacy.
Combinatorial Approaches
Integrating nanobioconjugates with small-molecule inhibitors could harness synergistic effects, balancing the rapid action of small molecules with the targeted precision of nanocarriers. Such hybrid approaches could optimize drug delivery, reduce dosing frequency, and minimize side effects.
CRISPR and Gene Editing Technologies
Gene editing techniques targeting CD44 or HA-synthesizing enzymes (eg, HAS2 or HAS3) offer promising strategies to modulate the signaling pathway at its genomic roots, potentially offering long-term correction of obesity-related dysregulations.
Advanced Nanoplatforms and AI Integration
Continued development of multifunctional, stimuli-responsive, and dual-ligand nanobioconjugates is essential. Artificial intelligence and machine learning can optimize nanocarrier design, predict biological interactions, and tailor therapy to patient-specific data.
Bridging Preclinical and Clinical Gaps
While preclinical evidence is promising, more well-designed, long-term, and ethnically diverse clinical trials are needed to validate the efficacy and safety of CD44-targeted interventions in humans.
Regulatory and Manufacturing Innovation
Scaling up nanobioconjugate production with consistency and biocompatibility must be a priority. Collaboration between academic institutions, biotech firms, and regulatory agencies can accelerate approval pathways for these advanced therapeutics.
Conclusion
This review highlights that the CD44hyaluronic acid (HA) axis has a central role in adipose tissue inflammation, remodeling, and insulin resistance, which are major processes that fuel obesity and its metabolic sequelae. The preclinical evidence suggests that modulating the CD44-signalling, especially by influencing the HA ligand, has a lot of promising therapeutic impacts. Of the strategies described here, HA-functionalized nanobioconjugates are the most specific, direct drugs towards the inflamed adipose tissues, and exhibit minimal systemic toxicity, compared to the administration of small molecule drugs. Although small molecules (metformin, curcumin, and CD44 antagonists) are relatively easy to administer systemically, they lack therapeutic specificity. Conversely, nanobioconjugates, albeit CD44-targeted versions in particular, allow site-selective, prolonged, and combinatorial modes of delivery, a particular advantage, given that obesity pathophysiology is complex.
Comparative analysis with other emerging modalities, including monoclonal antibodies, GLP-1 analogs, gene editing tools, and microbiome modulators, suggests that integrated strategies combining nanotechnology with molecular inhibition may offer the most effective path forward. However, challenges related to delivery specificity, long-term safety, and clinical translation remain. Altogether, the CD44–HA axis represents a viable and innovative target in obesity therapeutics. Future directions should focus on refining delivery systems, validating tissue-specific biomarkers, and advancing clinical trials to translate these findings into personalized and effective obesity interventions.
Data Sharing Statement
All data used are fully presented in the manuscript.
Acknowledgment
The graphical abstract was created in BioRender. Basajja, M. (2025) https://BioRender.com/0g2capd.
Author Contributions
Conceptualization: DEU, IJA, EUA, EGE, writing—original draft preparation: DEU, WAO, MAW, RIO, OPCU, writing—review and editing, all authors, and all the authors have read and agreed to the final version of the manuscript.
Funding
This review received no external funding or grants from any agency or organization.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Yadegar A, Pakpour S, Ibrahim FF, et al. Beneficial effects of fecal microbiota transplantation in recurrent Clostridioides difficile infection. Cell Host Microbe. 2023;31(5):695–711. doi:10.1016/j.chom.2023.03.019
2. Uti DE, Atangwho IJ, Omang WA, et al. Cytokines as key players in obesity low grade inflammation and related complications. Obesity Medicine. 2025;54:100585. doi:10.1016/j.obmed.2025.100585
3. Ahmed B, Konje JC. The epidemiology of obesity in reproduction. Best Pract Res Clin Obstet Gynaecol. 2023;89:102342. doi:10.1016/j.bpobgyn.2023.102342
4. Aloo SO, Barathikannan K, Oh D-H. Polyphenol-rich fermented hempseed ethanol extracts improve obesity, oxidative stress, and neural health in high-glucose diet-induced Caenorhabditis elegans. Food Chemistry. 2024;21:101233. doi:10.1016/j.fochx.2024.101233
5. Alum EU. Metabolic memory in obesity: can early-life interventions reverse lifelong risks? Obesity Medicine. 2025;55:100610. doi:10.1016/j.obmed.2025.100610
6. Umoru GU, Atangwho IJ, David-Oku E, et al. Modulation of Lipogenesis by Tetracarpidium conophorum Nuts via SREBP-1/ACCA-1/FASN Inhibition in Monosodium-Glutamate-Induced Obesity in Rats. Natural Product Communications. 2025;20:1934578X251344035. doi:10.1177/1934578X251344035
7. Alum EU, Ejemot-Nwadiaro RI, Betiang PA, Basajja M, Uti DE. Obesity and Climate Change: a Two-way Street with Global Health Implications. Obesity Medicine. 2025;100623. doi:10.1016/j.obmed.2025.100623.
8. Bensussen A, Torres-Magallanes JA, Roces De Álvarez-Buylla E. Molecular tracking of insulin resistance and inflammation development on visceral adipose tissue. Front. Immunol. 2023;14:1014778. doi:10.3389/fimmu.2023.1014778
9. Anguita-Ruiz A, Bustos-Aibar M, Plaza-Díaz J, et al. Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction. Int J Mol Sci. 2021;22:2756. doi:10.3390/ijms22052756
10. Bartelt A, Widenmaier SB, Schlein C, et al. Brown adipose tissue thermogenic adaptation requires Nrf1-mediated proteasomal activity. Nat Med. 2018;24(3):292–303. doi:10.1038/nm.4481
11. Umoru GU, Atangwho IJ, David‐Oku E, et al. Tetracarpidium conophorum nuts (African walnuts) up‐regulated adiponectin and PPAR‐γ expressions with reciprocal suppression of TNF‐α gene in obesity. J Cell Mol Med. 2024;28:e70086. doi:10.1111/jcmm.70086
12. Uti DE, Atangwho IJ, Eyong EU, et al. African walnuts (Tetracarpidium conophorum) modulate hepatic lipid accumulation in obesity via reciprocal actions on HMG-CoA reductase and paraoxonase. Endocrine, Metabolic & Immune Disorders-Drug Targets (Formerly Current Drug Targets-Immune. Endocrine & Metabolic Disorders). 2020;20:365–379.
13. Antonio CR, Trídico LA. The importance of interaction between hyaluronic acid and CD44 receptor. S&cd. 2021;13. doi:10.5935/scd1984-8773.2021130006.
14. Liu Z, Hou P, Fang J, et al. Hyaluronic acid metabolism and chemotherapy resistance: recent advances and therapeutic potential. Mol Oncol. 2024;18:2087–2106. doi:10.1002/1878-0261.13551
15. Yildiz R, Ganbold K, Sparman NZR, Rajbhandari P. Immune Regulatory Crosstalk in Adipose Tissue Thermogenesis. Comprehensive Physiology. 2025;15:e70001. doi:10.1002/cph4.70001
16. Weng X, Jiang H, Walker DJ, et al. Deletion of CD44 promotes adipogenesis by regulating PPARγ and cell cycle-related pathways. Journal of Endocrinology. 2024;262. doi:10.1530/JOE-24-0079.
17. Bemark M, Pitcher MJ, Dionisi C, Spencer J. Gut-associated lymphoid tissue: a microbiota-driven hub of B cell immunity. Trends in Immunology. 2024;45:211. doi:10.1016/j.it.2024.01.006
18. Alghamdi MA, Fallica AN, Virzì N, Kesharwani P, Pittalà V, Greish K. The Promise of Nanotechnology in Personalized Medicine. J Pers Med. 2022;12:673. doi:10.3390/jpm12050673
19. Azmi NAN, Elgharbawy AAM. Advances in Medical Applications: the Quest of Green Nanomaterials. In: Shanker U, Hussain CM, Rani M, editors. Handbook of Green and Sustainable Nanotechnology: Fundamentals, Developments and Applications. Cham: Springer International Publishing; 2023:1889–1909.
20. Chehelgerdi M, Chehelgerdi M, Allela OQB, et al. Progressing nanotechnology to improve targeted cancer treatment: overcoming hurdles in its clinical implementation. Mol Cancer. 2023;22:169. doi:10.1186/s12943-023-01865-0
21. Uti DE, Atangwho IJ, Alum EU, et al. Antioxidants in cancer therapy mitigating lipid peroxidation without compromising treatment through nanotechnology. Discover Nano. 2025;20(70). doi:10.1186/s11671-025-04248-0
22. Du S, Guan Y, Xie A, et al. Extracellular vesicles: a rising star for therapeutics and drug delivery. Journal of Nanobiotechnology. 2023;21:231. doi:10.1186/s12951-023-01973-5
23. Kurul F, Turkmen H, Cetin AE, Topkaya SN. Nanomedicine: how nanomaterials are transforming drug delivery, bio-imaging, and diagnosis. Next Nanotechnology. 2025;7:100129. doi:10.1016/j.nxnano.2024.100129
24. Skandalis SS. CD44 Intracellular Domain: a Long Tale of a Short Tail. Cancers (Basel). 2023;15:5041. doi:10.3390/cancers15205041
25. Senbanjo LT, Chellaiah MA. CD44: a Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front Cell Dev Biol. 2017;5. doi:10.3389/fcell.2017.00018.
26. Yanova M, Stepanova E, Maltseva D, Tonevitsky A. CD44 variant exons induce chemoresistance by modulating cell death pathways. Front Cell Dev Biol. 2025;13. doi:10.3389/fcell.2025.1508577.
27. Mosoane B, McCabe M, Jackson BS, Dlamini Z. CD44 Variant Expression in Follicular Cell-Derived Thyroid Cancers: implications for Overcoming Multidrug Resistance. Molecules. 2025;30:1899. doi:10.3390/molecules30091899
28. Ziranu P, Pretta A, Aimola V, et al. CD44: a New Prognostic Marker in Colorectal Cancer? Cancers. 2024;16:1569. doi:10.3390/cancers16081569
29. Maltseva D, Tonevitsky A. RNA-binding proteins regulating the CD44 alternative splicing. Front. Mol. Biosci. 2023;10. doi:10.3389/fmolb.2023.1326148.
30. Bays HE, Kirkpatrick C, Maki KC, et al. Obesity, dyslipidemia, and cardiovascular disease: a joint expert review from the Obesity Medicine Association and the National Lipid Association 2024. Obesity Pillars. 2024;10:100108. doi:10.1016/j.obpill.2024.100108
31. Cottam MA, Caslin HL, Winn NC, Hasty AH. Multiomics reveals persistence of obesity-associated immune cell phenotypes in adipose tissue during weight loss and weight regain in mice. Nat Commun. 2022;13:2950. doi:10.1038/s41467-022-30646-4
32. Fernández-Vallejo B, Monteagudo FJ, Romero L, et al. Cross-Sectional Analysis of IL-6, TNF-α, Adiponectin, Leptin, and Klotho Serum Levels in Relation to BMI Among Overweight and Obese Children Aged 10–14 in La Rioja, Spain. Children. 2025;12:89. doi:10.3390/children12010089
33. Xu H, Niu M, Yuan X, Wu K, Liu A. CD44 as a tumor biomarker and therapeutic target. Experimental Hematology & Oncology. 2020;9(36). doi:10.1186/s40164-020-00192-0
34. Yu W, Sun S, Yan Y, Zhou H, Liu Z, Fu Q. The role of short-chain fatty acid in metabolic syndrome and its complications: focusing on immunity and inflammation. Front. Immunol. 2025;16. doi:10.3389/fimmu.2025.1519925.
35. Wu S, Tan Y, Li F, Han Y, Zhang S, Lin X. CD44: a cancer stem cell marker and therapeutic target in leukemia treatment. Frontiers in Immunology. 2024;15:1354992. doi:10.3389/fimmu.2024.1354992
36. Bou Matar D, Zhra M, Nassar WK, et al. Adipose tissue dysfunction disrupts metabolic homeostasis: mechanisms linking fat dysregulation to disease. Front Endocrinol. 2025;16. doi:10.3389/fendo.2025.1592683.
37. Musale V, Wasserman DH, Kang L. Extracellular matrix remodelling in obesity and metabolic disorders. Life Metabolism. 2023;2:load021. doi:10.1093/lifemeta/load021
38. Cheng Y, Liang S, Zhang S, Hui X. Thermogenic Fat as a New Obesity Management Tool: from Pharmaceutical Reagents to Cell Therapies. Biomedicines. 2024;12:1474. doi:10.3390/biomedicines12071474
39. Mastrogiovanni M, Donnadieu E, Pathak R, Di Bartolo V. Subverting Attachment to Prevent Attacking: alteration of Effector Immune Cell Migration and Adhesion as a Key Mechanism of Tumor Immune Evasion. Biology. 2024;13:860. doi:10.3390/biology13110860
40. Ye J, Gao C, Liang Y, Hou Z, Shi Y, Wang Y. Characteristic and fate determination of adipose precursors during adipose tissue remodeling. Cell Regeneration. 2023;12(13). doi:10.1186/s13619-023-00157-8
41. Zhao JY, Zhou LJ, Ma KL, Hao R, Li M. MHO or MUO? White adipose tissue remodeling. Obesity Reviews. 2024;25:e13691. doi:10.1111/obr.13691
42. Palacios-Marin I, Serra D, Jimenez-Chillarón J, Herrero L, Todorčević M. Adipose Tissue Dynamics: cellular and Lipid Turnover in Health and Disease. Nutrients. 2023;15:3968. doi:10.3390/nu15183968
43. Gliniak CM, Pedersen L, Scherer PE. Adipose tissue fibrosis: the unwanted houseguest invited by obesity. Journal of Endocrinology. 2023;259:e230180. doi:10.1530/JOE-23-0180
44. Oxford Academic. Extracellular matrix remodelling in obesity and metabolic disorders|Life Metabolism. Available from: https://academic.oup.com/lifemeta/article/2/4/load021/7180395.
45. Chen C, Zhao S, Karnad A, Freeman JW. The biology and role of CD44 in cancer progression: therapeutic implications. Journal of Hematology & Oncology. 2018;11:64. doi:10.1186/s13045-018-0605-5
46. Hachiya R, Tanaka M, Itoh M, Suganami T. Molecular mechanism of crosstalk between immune and metabolic systems in metabolic syndrome. Inflammation and Regeneration. 2022;42(13). doi:10.1186/s41232-022-00198-7
47. Chang Y-C, Chan M-H, Yang Y-F, Li C-H, Hsiao M. Glucose transporter 4: insulin response mastermind, glycolysis catalyst and treatment direction for cancer progression. Cancer Letters. 2023;563:216179. doi:10.1016/j.canlet.2023.216179
48. Ayten M, Straub T, Kaplan L, Hauck SM, Grosche A, Koch SF. CD44 signaling in Müller cells impacts photoreceptor function and survival in healthy and diseased retinas. Journal of Neuroinflammation. 2024;21:190. doi:10.1186/s12974-024-03175-8
49. Di X, Gao X, Peng L, et al. Cellular mechanotransduction in health and diseases: from molecular mechanism to therapeutic targets. Sig Transduct Target Ther. 2023;8:282. doi:10.1038/s41392-023-01501-9
50. Mehner L-M, Munoz-Sagredo L, Sonnentag SJ, Treffert SM, Orian-Rousseau V. Targeting CD44 and other pleiotropic co-receptors as a means for broad inhibition of tumor growth and metastasis. Clin Exp Metastasis. 2024;41:599–611. doi:10.1007/s10585-024-10292-4
51. Agarwal G, Prasad KV,K, Bhaduri SB. Biosynthesis of Hyaluronic acid polymer: dissecting the role of sub structural elements of hyaluronan synthase. Sci Rep. 2019;9:12510. doi:10.1038/s41598-019-48878-8
52. Hayward SL, Wilson CL, Kidambi S. Hyaluronic acid-conjugated liposome nanoparticles for targeted delivery to CD44 overexpressing glioblastoma cells. Oncotarget. 2016;7:34158–34171. doi:10.18632/oncotarget.8926
53. Sepuru KM, Rajarathnam K. Structural basis of chemokine interactions with heparan sulfate, chondroitin sulfate, and dermatan sulfate. J Biol Chem. 2019;294:15650–15661. doi:10.1074/jbc.RA119.009879
54. Wang J, Wu Z, Cao L, Long F. Differential Regulation of Hyaluronan Synthesis by Three Isoforms of Hyaluronan Synthases in Mammalian Cells. Biomolecules. 2024;14:1567. doi:10.3390/biom14121567
55. Buckley C, Murphy EJ, Montgomery TR, Major I. Hyaluronic Acid: a Review of the Drug Delivery Capabilities of This Naturally Occurring Polysaccharide. Polymers. 2022;14:3442. doi:10.3390/polym14173442
56. Ye H, Zhang R, Zhang C, Xia Y, Jin L. Advances in hyaluronic acid: bioactivity, complexed biomaterials and biological application: a review. Asian Journal of Surgery. 2025;48:49–61. doi:10.1016/j.asjsur.2024.08.100
57. Salih ARC, Farooqi HMU, Amin H, Karn PR, Meghani N, Nagendran S. Hyaluronic acid: comprehensive review of a multifunctional biopolymer. Future Journal of Pharmaceutical Sciences. 2024;10:63. doi:10.1186/s43094-024-00636-y
58. Li G, Fang X, Liu Y, et al. Lipid Regulatory Element Interact with CD44 on Mitochondrial Bioenergetics in Bovine Adipocyte Differentiation and Lipometabolism. J. Agric. Food Chem. 2024;72:17481–17498. doi:10.1021/acs.jafc.4c02434
59. McDonald B, Kubes P. Interactions between CD44 and Hyaluronan in Leukocyte Trafficking. Front Immunol. 2015;6:68. doi:10.3389/fimmu.2015.00068
60. Collins MN, Zamboni F, Serafin A, Ren G, Thanusha AV, Culebras M. The Role of Hyaluronic Acid in Tissue Engineering. In: Polysaccharides of Microbial Origin. Cham: Springer; 2022:1063–1116.
61. Kotla NG, Bonam SR, Rasala S, et al. Recent advances and prospects of hyaluronan as a multifunctional therapeutic system. Journal of Controlled Release. 2021;336:598–620. doi:10.1016/j.jconrel.2021.07.002
62. Wang B, Hu S, Teng Y, et al. Current advance of nanotechnology in diagnosis and treatment for malignant tumors. Sig Transduct Target Ther. 2024;9:1–65. doi:10.1038/s41392-024-01889-y
63. Zhao M, Okunishi K, Bu Y, et al. Targeting activin receptor–like kinase 7 ameliorates adiposity and associated metabolic disorders. JCI Insight. 2023;8. doi:10.1172/jci.insight.161229.
64. Zhang Z, Peng Z, Wang R, Guo X, Gao J. Metabolomic analysis reveals macrophage metabolic reprogramming and polarization under different nutritional cues. Clinica Chimica Acta. 2024;560:119735. doi:10.1016/j.cca.2024.119735
65. Uti DE, Atangwho IJ, Alum EU, Egba SI, Ugwu OP-C, Ikechukwu GC. Natural Antidiabetic Agents: current Evidence and Development Pathways from Medicinal Plants to Clinical use. Natural Product Communications. 2025;20. doi:10.1177/1934578X251323393.
66. El-Saadony MT, Yang T, Korma SA, et al. Impacts of turmeric and its principal bioactive curcumin on human health: pharmaceutical, medicinal, and food applications: a comprehensive review. Front Nutr. 2023;9:1040259. doi:10.3389/fnut.2022.1040259
67. Okutani A, Morikawa S, Maeda K. Green Tea Catechin Epigallocatechin Gallate Inhibits Vegetative Cell Outgrowth and Expression of Beta-Lactamase Genes in Penicillin-Resistant Bacillus anthracis Strain PCr. Pathogens. 2024;13:699. doi:10.3390/pathogens13080699
68. Zhang D, Fu M, Gao S-H, Liu J-L. Curcumin and Diabetes: a Systematic Review. Evid Based Complement Alternat Med. 2013;636053. doi:10.1155/2013/636053
69. Cavasotto CN, Aucar MG, Adler NS. Computational chemistry in drug lead discovery and design. International Journal of Quantum Chemistry. 2019;119:e25678. doi:10.1002/qua.25678
70. Okoh OS, Yakubu A, Adegboyega AE, et al. Identification of some bioactive compounds from Trignonella foenumgraecum as possible inhibitors of PPARϒ for diabetes treatment through molecular docking studies, pharmacophore modelling and ADMET profiling: an in-silico study. PLoS One. 2023;18:e0284210. doi:10.1371/journal.pone.0284210
71. Umar AK, Limpikirati PK, Rivai B, Ardiansah I, Sriwidodo S, Luckanagul JA. Complexed hyaluronic acid-based nanoparticles in cancer therapy and diagnosis: research trends by natural language processing. Heliyon. 2025;11:e41246. doi:10.1016/j.heliyon.2024.e41246
72. Beach MA, Nayanathara U, Gao Y, et al. Polymeric Nanoparticles for Drug Delivery. Chem Rev. 2024;124:5505–5616. doi:10.1021/acs.chemrev.3c00705
73. Espejo-Román JM, Rubio-Ruiz B, Cano-Cortés V, et al. Selective Anticancer Therapy Based on a HA-CD44 Interaction Inhibitor Loaded on Polymeric Nanoparticles. Pharmaceutics. 2022;14:788. doi:10.3390/pharmaceutics14040788
74. Vugts DJ, Heuveling DA, Stigter-van Walsum M, et al. Preclinical evaluation of 89Zr-labeled anti-CD44 monoclonal antibody RG7356 in mice and cynomolgus monkeys. MAbs. 2014;6:567–575. doi:10.4161/mabs.27415
75. Cho J, Park E. Curcumin ameliorates the effects of high-fat diet-induced obesity via activating the DNA repair response. Br J Nutr. 2025;1–10. doi:10.1017/S000711452510353X
76. Farooqi AA, Pinheiro M, Granja A, et al. EGCG Mediated Targeting of Deregulated Signaling Pathways and Non-Coding RNAs in Different Cancers: focus on JAK/STAT, Wnt/β-Catenin, TGF/SMAD, NOTCH, SHH/GLI, and TRAIL Mediated Signaling Pathways. Cancers (Basel). 2020;12:951. doi:10.3390/cancers12040951
77. Sharifi-Rad M, Pezzani R, Redaelli M, et al. Preclinical Activities of Epigallocatechin Gallate in Signaling Pathways in Cancer. Molecules. 2020;25:467. doi:10.3390/molecules25030467
78. Abo El-Magd NF, El-Mesery M, El-Karef A, El-Shishtawy MM. Glycyrrhizin ameliorates high fat diet-induced obesity in rats by activating NrF2 pathway. Life Sci. 2018;193:159–170. doi:10.1016/j.lfs.2017.11.005
79. Aljabal G, Teh A-H, Yap BK. In Silico Prediction and Biophysical Validation of Novel 14-3-3σ Homodimer Stabilizers. J. Chem. Inf. Model. 2023;63:5619–5630. doi:10.1021/acs.jcim.3c00791
80. Mandujano-Lázaro G, Torres-Rojas MF, Ramírez-Moreno E, Marchat LA. Virtual screening combined with molecular docking for the !identification of new anti-adipogenic compounds. Sci Prog. 2025;108:00368504251320313. doi:10.1177/00368504251320313
81. Tiryaki S, Erol A, Ergün MO, Tiryaki S, Erol A, Ergün MO. Resveratrol Supplementation Attenuates Excessive Inflammation and Helps Restore Impaired Restitution in an Intestinal Epithelial Cell Culture Model. The Journal of Pediatric Research. 2022. doi:10.4274/jpr.galenos.2022.68552
82. Fang L, Gao D, Wang T, Zhao H, Zhang Y, Wang S. From nature to clinic: quercetin’s role in breast cancer immunomodulation. Front. Immunol. 2024;15:1483459. doi:10.3389/fimmu.2024.1483459
83. Espejo-Román JM, Rubio-Ruiz B, Chayah-Ghaddab M, et al. N-aryltetrahydroisoquinoline derivatives as HA-CD44 interaction inhibitors: design, synthesis, computational studies, and antitumor effect. European Journal of Medicinal Chemistry. 2023;258:115570. doi:10.1016/j.ejmech.2023.115570
84. Park M, Sung M. Carnosic acid attenuates obesity‐induced glucose intolerance and hepatic fat accumulation by modulating genes of lipid metabolism in
85. Liao C-H, Hung H-C, Lai C-N, et al. Carnosic acid and rosemary extract reversed the lipid accumulation induced by bisphenol A in the 3T3-L1 preadipocytes and C57BL/6J mice via SIRT1/FoxO1 pathway. Food and Chemical Toxicology. 2023;179:113996. doi:10.1016/j.fct.2023.113996
86. Pęcak A, Skalniak Ł, Pels K, et al. Anti-CD44 DNA Aptamers Selectively Target Cancer Cells. Nucleic Acid Ther. 2020;30:289–298. doi:10.1089/nat.2019.0833
87. Mahoney SA, Venkatasubramanian R, Darrah MA, et al. Intermittent supplementation with fisetin improves arterial function in old mice by decreasing cellular senescence. Aging Cell. 2024;23:e14060. doi:10.1111/acel.14060
88. Boman BM, V Viswanathan, COB Facey, et al. The v8-10 variant isoform of CD44 is selectively expressed in the normal human colonic stem cell niche and frequently is overexpressed in colon carcinomas during tumor development. Cancer Biology & Therapy. 2023;24:2195363. doi:10.1080/15384047.2023.2195363
89. Tarullo SE, He Y, Daughters C, et al. Receptor for hyaluronan-mediated motility (RHAMM) defines an invasive niche associated with tumor progression and predicts poor outcomes in breast cancer patients. The Journal of Pathology. 2023;260:289–303. doi:10.1002/path.6082
90. Sakers A, De Siqueira MK, Seale P, Villanueva CJ. Adipose-tissue plasticity in health and disease. Cell. 2022;185:419–446. doi:10.1016/j.cell.2021.12.016
91. Mukherjee S, Joshi V, Reddy KP, Singh N, Das P, Datta P. Biopharmaceutical and pharmacokinetic attributes to drive nanoformulations of small molecule tyrosine kinase inhibitors. Asian Journal of Pharmaceutical Sciences. 2024;19:100980. doi:10.1016/j.ajps.2024.100980
92. Almawash S. Oral Bioavailability Enhancement of Anti-Cancer Drugs Through Lipid Polymer Hybrid Nanoparticles. Pharmaceutics. 2025;17:381. doi:10.3390/pharmaceutics17030381
93. Zhang F, Xia Y, Su J, et al. Neutrophil diversity and function in health and disease. Sig Transduct Target Ther. 2024;9:343. doi:10.1038/s41392-024-02049-y
94. Ouhtit A, Rizeq B, Saleh HA, Rahman MM, Zayed H. Novel CD44-downstream signaling pathways mediating breast tumor invasion. Int J Biol Sci. 2018;14:1782–1790. doi:10.7150/ijbs.23586
95. Andreana I, N Zoratto, C Di Meo, et al. An overview of hyaluronic-acid nanoparticles for cancer cell targeted drug delivery. Expert Opinion on Drug Delivery. 2025;2025:1–18. doi:10.1080/17425247.2025.2515266
96. Anwar DM, Hedeya HY, Ghozlan SH, Ewas BM, Khattab SN. Surface-modified lipid-based nanocarriers as a pivotal delivery approach for cancer therapy: application and recent advances in targeted cancer treatment. Beni-Suef University Journal of Basic and Applied Sciences. 2024;13(106). doi:10.1186/s43088-024-00566-x
97. Mohite P, Singh S, Pawar A, et al. Lipid-based oral formulation in capsules to improve the delivery of poorly water-soluble drugs. Front Drug Deliv. 2023;3. doi:10.3389/fddev.2023.1232012
98. Sarvepalli S, Pasika SR, Verma V, et al. A Review on the Stability Challenges of Advanced Biologic Therapeutics. Pharmaceutics. 2025;17:550. doi:10.3390/pharmaceutics17050550
99. He W, Yang F, Chen K, Zeng Q. Targeted gold nanoparticles for ovarian cancer (Review). Oncol Lett. 2024;28:589. doi:10.3892/ol.2024.14723
100. Aggarwal R, Sheikh A, Akhtar M, et al. Understanding gold nanoparticles and their attributes in ovarian cancer therapy. Molecular Cancer. 2025;24:88. doi:10.1186/s12943-025-02280-3
101. Zhang D, Liu L, Wang J, et al. Drug-loaded PEG-PLGA nanoparticles for cancer treatment. Front Pharmacol. 2022;13:990505. doi:10.3389/fphar.2022.990505
102. Wu J, Deng C, Meng F, Zhang J, Sun H, Zhong Z. Hyaluronic acid coated PLGA nanoparticulate docetaxel effectively targets and suppresses orthotopic human lung cancer. Journal of Controlled Release. 2017;259:76–82. doi:10.1016/j.jconrel.2016.12.024
103. Tirella A, Kloc-Muniak K, Good L, et al. CD44 targeted delivery of siRNA by using HA-decorated nanotechnologies for KRAS silencing in cancer treatment. Int J Pharm. 2019;561:114–123. doi:10.1016/j.ijpharm.2019.02.032
104. Puluhulawa LE, Joni IM, Elamin KM, Mohammed AFA, Muchtaridi M, Wathoni N. Chitosan–Hyaluronic Acid Nanoparticles for Active Targeting in Cancer Therapy. Polymers. 2022;14:3410. doi:10.3390/polym14163410
105. Zhang C-Y, Gu K, Chi MY, et al. The application progress of PAMAM dendrimer in cancer imaging and treatment. J Biomater Sci Polym Ed. 2025;2025:1–38. doi:10.1080/09205063.2025.2497623
106. Kesharwani P, Xie L, Banerjee S, et al. Hyaluronic acid-conjugated polyamidoamine dendrimers for targeted delivery of 3,4-difluorobenzylidene curcumin to CD44 overexpressing pancreatic cancer cells. Colloids Surf B Biointerfaces. 2015;136:413–423. doi:10.1016/j.colsurfb.2015.09.043
107. Andreana I, Bincoletto V, Ricci C, et al. Smart hyaluronated micelles to enhance a gemcitabine prodrug efficacy. Journal of Drug Delivery Science and Technology. 2025;104:106518. doi:10.1016/j.jddst.2024.106518
108. Ji P, Wang L, Chen Y, Wang S, Wu Z, Qi X. Hyaluronic acid hydrophilic surface rehabilitating curcumin nanocrystals for targeted breast cancer treatment with prolonged biodistribution. Biomater. Sci. 2019;8:462–472. doi:10.1039/C9BM01605H
109. Jurczyk M, Jelonek K, Musiał-Kulik M, Beberok A, Wrześniok D, Kasperczyk J. Single- versus Dual-Targeted Nanoparticles with Folic Acid and Biotin for Anticancer Drug Delivery. Pharmaceutics. 2021;13:326. doi:10.3390/pharmaceutics13030326
110. Lu J, Zhao W, Huang Y, et al. Targeted Delivery of Doxorubicin by Folic Acid-Decorated Dual Functional Nanocarrier. Mol. Pharmaceutics. 2014;11:4164–4178. doi:10.1021/mp500389v
111. Hao X, Wang S, Wang L, et al. Exosomes as drug delivery systems in glioma immunotherapy. J Nanobiotechnology. 2024;22:340. doi:10.1186/s12951-024-02611-4
112. Mirzaei S, Mahabady MK, Zabolian A, et al. Small interfering RNA (siRNA) to target genes and molecular pathways in glioblastoma therapy: current status with an emphasis on delivery systems. Life Sci. 2021;275:119368. doi:10.1016/j.lfs.2021.119368
113. Li X, Wu Y, Zhang R, Bai W, Ye T, Wang S. Oxygen-Based Nanocarriers to Modulate Tumor Hypoxia for Ameliorated Anti-Tumor Therapy: fabrications, Properties, and Future Directions. Front Mol Biosci. 2021;8:683519. doi:10.3389/fmolb.2021.683519
114. Zhou X, He C, Liu M, et al. Self-assembly of hyaluronic acid-mediated tumor-targeting theranostic nanoparticles. Biomater. Sci. 2021;9:2221–2229. doi:10.1039/D0BM01855D
115. Dirheimer L, Pons T, Marchal F, Bezdetnaya L. Quantum Dots Mediated Imaging and Phototherapy in Cancer Spheroid Models: state of the Art and Perspectives. Pharmaceutics. 2022;14:2136. doi:10.3390/pharmaceutics14102136
116. Yang HY, Jang M-S, Sun XS, et al. CD44-mediated tumor homing of hyaluronic acid nanogels for hypoxia-activated photodynamic therapy against tumor. Colloids Surf B Biointerfaces. 2023;228:113395. doi:10.1016/j.colsurfb.2023.113395
117. Chianese G, Fasolino I, Tramontano C, et al. ROS-Generating Hyaluronic Acid-Modified Zirconium Dioxide-Acetylacetonate Nanoparticles as a Theranostic Platform for the Treatment of Osteosarcoma. Nanomaterial. 2022;13(54). doi:10.3390/nano13010054
118. Yassin MT, Al-Otibi FO, Al-Sahli SA, El-Wetidy MS, Mohamed S. Metal Oxide Nanoparticles as Efficient Nanocarriers for Targeted Cancer Therapy: addressing Chemotherapy-Induced Disabilities. Cancers. 2024;16:4234. doi:10.3390/cancers16244234
119. Lu J, Luo B, Chen Z, et al. Host-guest fabrication of dual-responsive hyaluronic acid/mesoporous silica nanoparticle based drug delivery system for targeted cancer therapy. International Journal of Biological Macromolecules. 2020;146:363–373. doi:10.1016/j.ijbiomac.2019.12.265
120. Chen K, Chang C, Liu Z, et al. Hyaluronic acid targeted and pH-responsive nanocarriers based on hollow mesoporous silica nanoparticles for chemo-photodynamic combination therapy. Colloids and Surfaces B: Biointerfaces. 2020;194:111166. doi:10.1016/j.colsurfb.2020.111166
121. Zhong S, Liu P, Ding J, Zhou W. Hyaluronic Acid-Coated MTX-PEI Nanoparticles for Targeted Rheumatoid Arthritis Therapy. Crystals. 2021;11:321. doi:10.3390/cryst11040321
122. Gao J, Li J, Luo Z, Wang H, Ma Z. Nanoparticle-Based Drug Delivery Systems for Inflammatory Bowel Disease Treatment. Drug Des Devel Ther. 2024;18:2921–2949. doi:10.2147/DDDT.S461977
123. Yin X, Harmancey R, McPherson DD, Kim H, Huang S-L. Liposome-Based Carriers for CRISPR Genome Editing. Int J Mol Sci. 2023;24:12844. doi:10.3390/ijms241612844
124. Zhang J, Yang X, Chang Z, Zhu W, Ma Y, He H. Polymeric nanocarriers for therapeutic gene delivery. Asian Journal of Pharmaceutical Sciences. 2025;20:101015. doi:10.1016/j.ajps.2025.101015
125. Wang Y, Tang Y, Guo L, et al. Recent advances in zeolitic imidazolate frameworks as drug delivery systems for cancer therapy. Asian Journal of Pharmaceutical Sciences. 2025;20:101017. doi:10.1016/j.ajps.2025.101017
126. Wang Y, M Zeng, T Fan, et al. Biomimetic ZIF-8 Nanoparticles: a Novel Approach for Biomimetic Drug Delivery Systems. Int J Nanomed. 2024;19:5523–5544. doi:10.2147/IJN.S462480
127. He C, Zhou J, Wu X, et al. Hyaluronic acid-zein shell-core biopolymer nanoparticles enhance hepatocellular carcinoma therapy of celastrol via CD44-mediated cellular uptake. Int J Biol Macromol. 2024;281:136096. doi:10.1016/j.ijbiomac.2024.136096
128. Nag S, Kar S, Mishra S, et al. Unveiling Green Synthesis and Biomedical Theranostic paradigms of Selenium Nanoparticles (SeNPs) - A state-of-the-art comprehensive update. Int J Pharm. 2024;662:124535. doi:10.1016/j.ijpharm.2024.124535
129. Mai S, Inkielewicz-Stepniak I. Graphene Oxide Nanoparticles and Organoids: a Prospective Advanced Model for Pancreatic Cancer Research. Int J Mol Sci. 2024;25:1066. doi:10.3390/ijms25021066
130. Kearns O, Camisasca A, Giordani S. Hyaluronic Acid-Conjugated Carbon Nanomaterials for Enhanced Tumour Targeting Ability. Molecules. 2021;27(48):1. doi:10.3390/molecules27010048
131. González M, Romero MP. Surface-Modified Carbon Dots for Cancer Therapy: integrating Diagnostic and Therapeutic Applications. Int J Nanomedicine. 2025;20:7715–7741. doi:10.2147/IJN.S508181
132. Mohammadi T, Gheybalizadeh H, Rahimpour E, Soleymani J, Shafiei-Irannejad V. Advanced photoluminescent nanomaterials for targeted bioimaging of cancer cells. Heliyon. 2025;11:e41566. doi:10.1016/j.heliyon.2024.e41566
133. Wang C, Shi X, Song H, et al. Polymer-lipid hybrid nanovesicle-enabled combination of immunogenic chemotherapy and RNAi-mediated PD-L1 knockdown elicits antitumor immunity against melanoma. Biomaterials. 2021;268:120579. doi:10.1016/j.biomaterials.2020.120579
134. Z Zhang, Z Zhang, W Zeng, et al. A hyaluronic acid-based dual-functional hydrogel microneedle system for sequential melanoma ablation and skin regeneration. Int J Biological Macromolecules. 2024;283:138039. doi:10.1016/j.ijbiomac.2024.138039
135. Chen H-A, Lu Y-J, Dash BS, Chao Y-K, Chen J-P. Hyaluronic Acid-Modified Cisplatin-Encapsulated Poly(Lactic-co-Glycolic Acid) Magnetic Nanoparticles for Dual-Targeted NIR-Responsive Chemo-Photothermal Combination Cancer Therapy. Pharmaceutics. 2023;15:290. doi:10.3390/pharmaceutics15010290
136. Hlaing SP, Cao J, Lee J, et al. Hyaluronic Acid-Conjugated PLGA Nanoparticles Alleviate Ulcerative Colitis via CD44-Mediated Dual Targeting to Inflamed Colitis Tissue and Macrophages. Pharmaceutics. 2022;14:2118. doi:10.3390/pharmaceutics14102118
137. Li W, Lin J, Zhou J, et al. Hyaluronic acid-functionalized DDAB/PLGA nanoparticles for improved oral delivery of magnolol in the treatment of ulcerative colitis. International Journal of Pharmaceutics. 2024;653:123878. doi:10.1016/j.ijpharm.2024.123878
138. Gazzano E, Buondonno I, Marengo A, et al. Hyaluronated liposomes containing H2S-releasing doxorubicin are effective against P-glycoprotein-positive/doxorubicin-resistant osteosarcoma cells and xenografts. Cancer Lett. 2019;456:29–39. doi:10.1016/j.canlet.2019.04.029
139. Yang B, Mao Y, Zhang Y, et al. HA-Coated PLGA Nanoparticles Loaded with Apigenin for Colon Cancer with High Expression of CD44. Molecules. 2023;28:7565. doi:10.3390/molecules28227565
140. Menon A, Abd-Aziz N, Khalid K, Poh CL, Naidu R. miRNA: a Promising Therapeutic Target in Cancer. Int J Mol Sci. 2022;23:11502. doi:10.3390/ijms231911502
141. Kaushik A, Khan S, Pharasi N, Mani S. Dual pH and ultrasound responsive nanocarriers: a smart approach in cancer theranostics. Journal of Drug Delivery Science and Technology. 2024;95:105560. doi:10.1016/j.jddst.2024.105560
142. Bonet-Aleta J, Calzada-Funes J, Hueso JL. Manganese oxide nano-platforms in cancer therapy: recent advances on the development of synergistic strategies targeting the tumor microenvironment. Applied Materials Today. 2022;29:101628. doi:10.1016/j.apmt.2022.101628
143. Rios de la Rosa JM, Pingrajai P, Pelliccia M, et al. Binding and Internalization in Receptor-Targeted Carriers: the Complex Role of CD44 in the Uptake of Hyaluronic Acid-Based Nanoparticles (siRNA Delivery). Adv Healthc Mater. 2019;8:e1901182. doi:10.1002/adhm.201901182
144. Hu Z, Xu C, Liang Y, Liu T, Tian H, Zhang Y. Multifunctional drug delivery nanoparticles based on MIL-100 (Fe) for photoacoustic imaging-guided synergistic chemodynamic/chemo/photothermal breast cancer therapy. Materials & Design. 2022;223:111132. doi:10.1016/j.matdes.2022.111132
145. Magadani R, Ndinteh DT, Roux S, et al. Cytotoxic Effects of Lecaniodiscus Cupanioides (Planch.) Extract and Triterpenoids-derived Gold Nanoparticles On MCF-7 Breast Cancer Cell Lines. Anticancer Agents Med Chem. 2025;25:841–850. doi:10.2174/0118715206325529241004064307
146. Petersen J, Ludwig MQ, Juozaityte V, et al. GLP-1-directed NMDA receptor antagonism for obesity treatment. Nature. 2024;629:1133–1141. doi:10.1038/s41586-024-07419-8
147. Ashour MM, Mabrouk M, Aboelnasr MA, Beherei HH, Tohamy KM, Das DB. Anti-Obesity Drug Delivery Systems: recent Progress and Challenges. Pharmaceutics. 2023;15:2635. doi:10.3390/pharmaceutics15112635
148. Lu J, Liu P, Cai M, et al. Recent progress in the pharmacotherapy for obesity. European Journal of Pharmacology. 2025;1002:177850. doi:10.1016/j.ejphar.2025.177850
149. Zhang W, Taheri-Ledari R, Ganjali F, et al. Nanoscale bioconjugates: a review of the structural attributes of drug-loaded nanocarrier conjugates for selective cancer therapy. Heliyon. 2022;8:e09577. doi:10.1016/j.heliyon.2022.e09577
150. Cajigas S, Orozco J. Nanobioconjugates for Signal Amplification in Electrochemical Biosensing. Molecules. 2020;25:3542. doi:10.3390/molecules25153542
151. Aboul-Ella H, Gohar A, Ali AA, et al. Monoclonal antibodies: from magic bullet to precision weapon. Mol Biomed. 2024;5:47. doi:10.1186/s43556-024-00210-1
152. Ebrahimi SB, Samanta D. Engineering protein-based therapeutics through structural and chemical design. Nat Commun. 2023;14:2411. doi:10.1038/s41467-023-38039-x
153. Baggio LL, Drucker DJ. Glucagon-like peptide-1 receptor co-agonists for treating metabolic disease. Mol Metab. 2020;46:101090. doi:10.1016/j.molmet.2020.101090
154. Kardan R, Hemmati J, Nazari M, et al. Novel therapeutic strategy for obesity through the gut microbiota-brain axis: a review article. Caspian J Intern Med. 2024;15:215–227. doi:10.22088/cjim.15.2.215
155. Zhang Z, Mocanu V, Cai C, et al. Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review. Nutrients. 2019;11:2291. doi:10.3390/nu11102291
156. Subramaniyan V, Hanim YU. Role of pancreatic lipase inhibition in obesity treatment: mechanisms and challenges towards current insights and future directions. Int J Obes (Lond). 2025;49:492–506. doi:10.1038/s41366-025-01729-1
157. Dutta S, Shah RB, Singhal S, et al. Metformin: a Review of Potential Mechanism and Therapeutic Utility Beyond Diabetes. Drug Des Devel Ther. 2023;17:1907–1932. doi:10.2147/DDDT.S409373
158. Jangra A, B Babu, S Divakar, et al. An in-depth review of PPARγ modulators as anti-diabetes therapeutics. Drug Metabolism Reviews. 2025;2025:1–27. doi:10.1080/03602532.2025.2508152
159. Reiss AB, Gulkarov S, Lau R, et al. Weight Reduction with GLP-1 Agonists and Paths for Discontinuation While Maintaining Weight Loss. Biomolecules. 2025;15:408. doi:10.3390/biom15030408
160. Ghusn W, Hurtado MD. Glucagon-like Receptor-1 agonists for obesity: weight loss outcomes, tolerability, side effects, and risks. Obesity Pillars. 2024;12:100127. doi:10.1016/j.obpill.2024.100127
161. Lombardo D, Kiselev MA. Methods of Liposomes Preparation: formation and Control Factors of Versatile Nanocarriers for Biomedical and Nanomedicine Application. Pharmaceutics. 2022;14:543. doi:10.3390/pharmaceutics14030543
162. Yang J, Zeng H, Luo Y, et al. Recent Applications of PLGA in Drug Delivery Systems. Polymers. 2024;16:2606. doi:10.3390/polym16182606
163. Kaurav M, Ruhi S, Al-Goshae HA, et al. Dendrimer: an update on recent developments and future opportunities for the brain tumors diagnosis and treatment. Front Pharmacol. 2023;14:1159131. doi:10.3389/fphar.2023.1159131
164. Tatode A, Agrawal PR, Taksande J, et al. Role of folate receptor and CD44 in targeting of docetaxel and paclitaxel fabricated conjugates for efficient cancer therapy. Journal of Medicine, Surgery, and Public Health. 2025;5:100163. doi:10.1016/j.glmedi.2024.100163
165. Sargazi S, Arshad R, Ghamari R, et al. siRNA‐based nanotherapeutics as emerging modalities for immune‐mediated diseases: a preliminary review. Cell Biol Int. 2022;46:1320–1344. doi:10.1002/cbin.11841
166. Gabel M, Knauss A, Fischer D, Neurath MF, Weigmann B. Surface Design Options in Polymer- and Lipid-Based siRNA Nanoparticles Using Antibodies. Int J Mol Sci. 2022;23:13929. doi:10.3390/ijms232213929
167. Volčanšek Š, Koceva A, Jensterle M, Janež A, Muzurović E. Amylin: from Mode of Action to Future Clinical Potential in Diabetes and Obesity. Diabetes Ther. 2025;16:1207–1227. doi:10.1007/s13300-025-01733-8
168. Wu S, Zhou Y. Monoclonal antibody precision therapy targeting inflammation for bipolar disorder: a narrative review. Ther Adv Psychopharmacol. 2024;14:20451253241227772. doi:10.1177/20451253241227772
169. Khan MS, Qureshi N, Khan R, Son Y-O, Maqbool T. CRISPR/Cas9-Based therapeutics as a promising strategy for management of Alzheimer’s disease: progress and prospects. Front Cell Neurosci. 2025;19:1578138. doi:10.3389/fncel.2025.1578138
170. Selvamani S, Mehta V, Ali El Enshasy H, et al. Efficacy of Probiotics-Based Interventions as Therapy for Inflammatory Bowel Disease: a Recent Update. Saudi J Biol Sci. 2022;29:3546–3567. doi:10.1016/j.sjbs.2022.02.044
171. Hitch TCA, Hall LJ, Walsh SK, et al. Microbiome-based interventions to modulate gut ecology and the immune system. Mucosal Immunol. 2022;15:1095–1113. doi:10.1038/s41385-022-00564-1
172. Ji J, Jin W, Liu S, Jiao Z, Li X. Probiotics, prebiotics, and postbiotics in health and disease. MedComm. 2023;4:e420. doi:10.1002/mco2.420
173. Ali S, Hamayun M, Siraj M, Khan SA, Kim H-Y, Lee B. Recent advances in prebiotics: classification, mechanisms, and health applications. Future Foods. 2025;12:100680. doi:10.1016/j.fufo.2025.100680
174. Kim KO, Gluck M. Fecal Microbiota Transplantation: an Update on Clinical Practice. Clinical Endoscopy. 2019;52:137. doi:10.5946/ce.2019.009
175. Lin A, Jiang A, Huang L, et al. From chaos to order: optimizing fecal microbiota transplantation for enhanced immune checkpoint inhibitors efficacy. Gut Microbes. 2025;2025:1.
176. Bai L, Yu L, Ran M, et al. Harnessing the Potential of Exosomes in Therapeutic Interventions for Brain Disorders. Int J Mol Sci. 2025;26:2491. doi:10.3390/ijms26062491
177. Casado ME, Collado-Pérez R, Frago LM, Barrios V. Recent Advances in the Knowledge of the Mechanisms of Leptin Physiology and Actions in Neurological and Metabolic Pathologies. Int J Mol Sci. 2023;24:1422. doi:10.3390/ijms24021422
178. Recinella L, Orlando G, Ferrante C, Chiavaroli A, Brunetti L, Leone S. Adipokines: new Potential Therapeutic Target for Obesity and Metabolic, Rheumatic, and Cardiovascular Diseases. Frontiers in Physiology. 2020;11:578966. doi:10.3389/fphys.2020.578966
179. Kunnumakkara AB, Hegde M, Parama D, et al. Role of Turmeric and Curcumin in Prevention and Treatment of Chronic Diseases: lessons Learned from Clinical Trials. ACS Pharmacol Transl Sci. 2023;6:447–518. doi:10.1021/acsptsci.2c00012
180. Prapaharan B, Lea M, Beaudry JL. Weighing in on the role of brown adipose tissue for treatment of obesity. J. Pharm. Pharm. Sci. 2024;27:13157. doi:10.3389/jpps.2024.13157
181. Kim J, Li J, Wei J, et al. Regulatory T Cell Metabolism: a Promising Therapeutic Target for Cancer Treatment? Immune Netw. 2025;25:e13. doi:10.4110/in.2025.25.e13
182. Morchón-Araujo D, Catani G, Mirallas O, et al. Emerging Immunotherapy Targets in Early Drug Development. Int J Mol Sci. 2025;26:5394. doi:10.3390/ijms26115394
183. Aderinto N, Abdulbasit MO, Tangmi ADE, Okesanya JO, Mubarak JM. Unveiling the growing significance of metabolism in modulating immune cell function: exploring mechanisms and implications; a review. Ann Med Surg (Lond). 2023;85:5511–5522. doi:10.1097/MS9.0000000000001308
184. Didamson OC, Chandran R, Abrahamse H. Chapter 25 - Biomarkers and signaling pathways of esophageal cancer stem cells: promising targets to limit the metastatic potential and stemness. In: Pathak S, Banerjee A editors. Cancer Stem Cells and Signaling Pathways. Academic Press. 2024. 439–455.
185. Schleh MW, Caslin HL, Garcia JN, et al. Metaflammation in obesity and its therapeutic targeting. Sci Transl Med. 2023;15:eadf9382. doi:10.1126/scitranslmed.adf9382
186. Liu J, Xiao Q, Xiao J, et al. Wnt/β-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Sig Transduct Target Ther. 2022;7(3). doi:10.1038/s41392-021-00762-6
187. Qadri MM. Targeting CD44 Receptor Pathways in Degenerative Joint Diseases: involvement of Proteoglycan-4 (PRG4). Pharmaceuticals (Basel). 2023;16:1425. doi:10.3390/ph16101425
188. Gupta RC, Lall R, Srivastava A, Sinha A. Hyaluronic Acid: molecular Mechanisms and Therapeutic Trajectory. Front Vet Sci. 2019;6(192). doi:10.3389/fvets.2019.00192
189. Chistyakov DV, Astakhova AA, Azbukina NV, Goriainov SV, Chistyakov VV, Sergeeva MG. High and Low Molecular Weight Hyaluronic Acid Differentially Influences Oxylipins Synthesis in Course of Neuroinflammation. Int J Mol Sci. 2019;20:3894. doi:10.3390/ijms20163894
190. Snetkov P, Zakharova K, Morozkina S, Olekhnovich R, Uspenskaya M. Hyaluronic Acid: the Influence of Molecular Weight on Structural, Physical, Physico-Chemical, and Degradable Properties of Biopolymer. Polymers. 2020;12:1800. doi:10.3390/polym12081800
191. Kawai T, Autieri MV, Scalia R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol. 2021;320:C375–C391. doi:10.1152/ajpcell.00379.2020
192. Liu Y, Li L, Wang L, et al. ‘Two-faces’ of hyaluronan, a dynamic barometer of disease progression in tumor microenvironment. Discov Onc. 2023;14:11. doi:10.1007/s12672-023-00618-1
193. Sarkar K, Bank S, Chatterjee A, et al. Hyaluronic acid-graphene oxide quantum dots nanoconjugate as dual purpose drug delivery and therapeutic agent in meta-inflammation. Journal of Nanobiotechnology. 2023;21:246. doi:10.1186/s12951-023-02015-w
194. Al-Jaber H, Mohamed NA, Govindharajan VK, et al. In Vitro and In Vivo Validation of GATA-3 Suppression for Induction of Adipogenesis and Improving Insulin Sensitivity. Int J Mol Sci. 2022; 23: 11142. doi:10.3390/ijms231911142
195. Odeniyi IA, Ahmed B, Anbiah B, et al. An improved in vitro 3T3-L1 adipocyte model of inflammation and insulin resistance. Adipocyte. 2024. 13:2414919. doi:10.1080/21623945.2024.2414919
196. Holmes A, Coppey LJ, Davidson EP, Yorek MA. Rat Models of Diet-Induced Obesity and High Fat/Low Dose Streptozotocin Type 2 Diabetes: effect of Reversal of High Fat Diet Compared to Treatment with Enalapril or Menhaden Oil on Glucose Utilization and Neuropathic Endpoints. J Diabetes Res. 2015;2015:307285. doi:10.1155/2015/307285
197. Sadikan MZ, Abdul Nasir NA, Lambuk L, et al. Diabetic retinopathy: a comprehensive update on in vivo, in vitro and ex vivo experimental models. BMC Ophthalmol. 2023;23:421. doi:10.1186/s12886-023-03155-1
198. Stott NL, Marino JS. High Fat Rodent Models of Type 2 Diabetes: from Rodent to Human. Nutrients. 2020;12:3650. doi:10.3390/nu12123650
199. Huang D, Deng M, Kuang S. Polymeric carriers for controlled drug delivery in obesity treatment. Trends Endocrinol Metab. 2019;30:974–989. doi:10.1016/j.tem.2019.09.004
200. Chen D, Wang Y, Wu K, Wang X. Dual Effects of Metformin on Adipogenic Differentiation of 3T3-L1 Preadipocyte in AMPK-Dependent and Independent Manners. Int J Mol Sci. 2018;19:1547. doi:10.3390/ijms19061547
201. Sikur N, Böröczky C, Paszternák A, et al. Resveratrol and Its Derivatives Diminish Lipid Accumulation in Adipocytes In Vitro—Mechanism of Action and Structure–Activity Relationship. Nutrients. 2024;16:3869. doi:10.3390/nu16223869
202. Ding Y, Chen Q-B, Xu H, et al. siRNA nanoparticle targeting Usp20 lowers lipid levels and ameliorates metabolic syndrome in mice. J Lipid Res. 2024;65:100626. doi:10.1016/j.jlr.2024.100626
203. Khursheed R, Singh SK, Wadhwa S, et al. A sojourn into therapeutic and nutraceutical potential of curcumin and its novel drug delivery system: current achievements and future perspectives. South African Journal of Botany. 2022;149:944–962. doi:10.1016/j.sajb.2022.04.021
204. Zhang X, Zhang Y, Zhou M, et al. DPHC From Alpinia officinarum Ameliorates Oxidative Stress and Insulin Resistance via Activation of Nrf2/ARE Pathway in db/db Mice and High Glucose-Treated HepG2 Cells. Front. Pharmacol. 2022;12. doi:10.3389/fphar.2021.792977
205. Matsuura N, Asano C, Nagasawa K, et al. Effects of pioglitazone on cardiac and adipose tissue pathology in rats with metabolic syndrome. Int J Cardiol. 2015;179:360–369. doi:10.1016/j.ijcard.2014.11.099
206. Ćućuz V, Cvejić J, Gojković-Bukarica L. Clinical trials of resveratrol efficacy and safety. Vojnosanitetski Pregled. 2022;79:613–618.
207. Ortiz MG. Effect of Resveratrol Administration on Metabolic Syndrome, Insulin Sensitivity and Insulin Secretion. 2020.
208. Sharma D, Arora S, Banerjee A, Singh J. Improved insulin sensitivity in obese-diabetic mice via chitosan Nanomicelles mediated silencing of pro-inflammatory Adipocytokines. Nanomedicine: Nanotechnology, Biology and Medicine. 2021;33:102357. doi:10.1016/j.nano.2020.102357
209. Hong SY, Ha AW, Kim W. Effects of quercetin on cell differentiation and adipogenesis in 3T3-L1 adipocytes. Nutr Res Pract. 2021;15:444–455. doi:10.4162/nrp.2021.15.4.444
210. Gu C, Loube J, Lee R, et al. Metformin Alleviates Airway Hyperresponsiveness in a Mouse Model of Diet-Induced Obesity. Front. Physiol. 2022;13. doi:10.3389/fphys.2022.883275.
211. Ruiz-Ojeda FJ, Méndez-Gutiérrez A, Aguilera CM, Plaza-Díaz J. Extracellular Matrix Remodeling of Adipose Tissue in Obesity and Metabolic Diseases. Int J Mol Sci. 2019;20:4888. doi:10.3390/ijms20194888
212. Lee SC, Ma JSY, Kim MS, et al. A PSMA-targeted bispecific antibody for prostate cancer driven by a small-molecule targeting ligand. Sci Adv. 2021;7:eabi8193. doi:10.1126/sciadv.abi8193
213. Wu Y, Ma J, Chen J, et al. Ablation of CD44 Attenuates Adipogenesis in White Adipocytes via the Tryptophan 5-Hydroxylase 2/5-Hydroxytryptamine Axis to Protect Mice from High-Fat Diet–Induced Obesity. The American Journal of Pathology. 2025;195:247–264. doi:10.1016/j.ajpath.2024.10.005
214. Goyal S, Thirumal D, Rana J, et al. Chitosan based nanocarriers as a promising tool in treatment and management of inflammatory diseases. Carbohydrate Polymer Technologies and Applications. 2024;7:100442. doi:10.1016/j.carpta.2024.100442
215. Lee S. Gene Silencing in Neurogenetics: a Comprehensive Guide, Available from: https://www.numberanalytics.com/blog/ultimate-guide-gene-silencing-neurogenetics.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.