Back to Journals » Psoriasis: Targets and Therapy » Volume 15
Codon-Usage Bias and Genomic Architecture of 79 Psoriasis-Susceptibility Genes: Implications for Expression Efficiency and Therapeutic Targeting
Authors Jiang S, Chen L, Li J, Gao F ![]() , Yan X, Zhao Y
, Yan X, Zhao Y
Received 9 June 2025
Accepted for publication 6 September 2025
Published 25 September 2025 Volume 2025:15 Pages 479—499
DOI https://doi.org/10.2147/PTT.S545695
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Enzo Errichetti
Shanshan Jiang,1 Lu Chen,1 Jianghui Li,2 Fei Gao,3 Xiaoning Yan,1 Yiding Zhao1
1Department of Dermatology, Shaanxi Provincial Hospital of Chinese Medicine, Xi’an, 710003, People’s Republic of China; 2First Clinical Medical College, Shaanxi University of Chinese Medicine, Xianyang, 712046, People’s Republic of China; 3Shaoxing Academy of Biomedicine, Zhejiang Sci-Tech University, Shaoxing, 312030, People’s Republic of China
Correspondence: Xiaoning Yan, Department of Dermatology, Shaanxi Provincial Hospital of Chinese Medicine, Xihuamen Road, Xi’an, People’s Republic of China, Email [email protected] Yiding Zhao, Department of Dermatology, Shaanxi Provincial Hospital of Chinese Medicine, Xihuamen Road, Xi’an, People’s Republic of China, Email [email protected]
Introduction: Psoriasis is a chronic autoimmune skin disorder with a complex genetic basis. However, the codon usage patterns and nucleotide features of psoriasis-related genes remain unexplored, despite their potential to influence gene expression and disease progression.
Methods: We analyzed 79 psoriasis-associated genes to investigate codon usage bias (CUB) and nucleotide composition. Metrics included GC content, effective number of codons (ENC), and relative synonymous codon usage (RSCU). Evolutionary influences were assessed using correspondence analysis, parity rule 2 (PR2) plots, and neutrality plots.
Results: Functional enrichment analysis identified pathway involvement. Comparative genomic analysis evaluated differences in coding sequence and UTR lengths and GC content relative to the genome-wide background. Psoriasis-related genes showed high GC content (mean = 53.3 ± 9.3%) with a strong preference for GC-ending codons, especially at the third codon position (GC3 = 60.6 ± 16.1%). RSCU analysis revealed frequent use of GCC (alanine), CTG (leucine), and GTG (valine). While the mean ENC (46.2 ± 9.9) suggested moderate codon bias, several genes displayed strong bias (ENC < 30). Selection pressure accounted for 71% of codon usage variation, with mutation pressure contributing 29%. Functional enrichment showed significant involvement in IL-17 (FDR = 3.4× 10− 3), JAK-STAT (FDR = 3.4× 10− 3), and TNF (FDR = 8.0× 10−³) signaling pathways. These genes also tended to have shorter coding sequences and 5′UTRs and higher GC content compared to genome-wide averages.
Conclusion: In conclusion, this study reveals that psoriasis-related genes are under strong selective pressure, enriched in key inflammatory pathways, and exhibit codon and nucleotide features that may optimize expression in inflamed tissues. These insights have translational relevance for designing codon-optimized mRNAs, gene therapies, and diagnostic tools tailored to autoimmune diseases like psoriasis.
Keywords: psoriasis, candidate genes, codon usage bias, mutation pressure, natural selection
Introduction
Psoriasis is a chronic, nontransmissible, inflammatory, autoimmune disease affecting nearly 0.5–4.6% world’s population, significantly impacting quality of life and imposing a substantial societal burden.1,2 It is characterized by immune infiltration, hyperproliferation, and abnormal differentiation of keratinocytes.3 Psoriasis manifests in various forms, including guttate, plaque, inverse, palmoplantar, erythrodermic, and pustular psoriasis.4 As a multifactorial disorder, it is driven by genetic, environmental, and immune factors, with a strong genetic basis highlighted by familial recurrence and higher concordance in monozygotic twins compared to dizygotic twins.5 Genome-wide linkage studies have identified multiple risk loci related to psoriasis (PSORS 1–15), with PSORS1 located in the major histocompatibility complex region contributing 35–50% of genetic susceptibility.6 Key genes, such as HLA-C*06:02, are associated with a higher risk of psoriasis and linked to clinical characteristics.7 Other genes, including IL23R and TNFRSF1B, are involved in critical pathways like IL-23/Th17, NFκB (nuclear factor kappa B), and TNF-a signalling, which drive inflammation and disease pathogenesis.7,8 Immune-related genes often exhibit distinctive codon-usage patterns across diverse biological contexts, underscoring the relevance of examining CUB in psoriasis. Moreover, because codon optimality can shape mRNA stability and translation efficiency, inflammatory skin microenvironments may modulate the expression of psoriasis-associated genes through their codon composition.8 Understanding the genetic and molecular basis of psoriasis is important for advancing treatments and developing personalized management strategies.
Codon usage bias (CUB), the preferential use of specific synonymous codons, influences gene expression, protein folding, and function.9,10 Studying CUB in psoriasis-related genes may reveal novel regulatory mechanisms that impact disease pathways and identify genes more amenable to therapeutic intervention, thereby advancing personalized medicine approaches for psoriasis. The degeneracy of the genetic code allows multiple codons to encode the same amino acid, yet their usage varies across genes and organisms.11 This phenomenon, influenced by mutation pressure, natural selection, and translational efficiency, affects gene expression and protein production.12,13 In this study, we investigate the nucleotide composition and CUB of 79 psoriasis-related candidate genes from PSORS 1–15 loci using bioinformatic tools. We focus on translational efficiency, optimal codon usage, and gene expression patterns to uncover key factors shaping CUB in these genes. This work provides foundational insights into the molecular mechanisms underlying psoriasis and paves the way for future research into clinically relevant therapeutic strategies.
Methodology
Availability of Sequence Data
The coding sequences (CDS) of psoriasis-susceptible candidate genes having perfect start and stop codons were retrieved from the National Centre for Biotechnology Information, USA (http://www.ncbi.nlm.nih.gov/). A comprehensive list of these psoriasis susceptibility loci, candidate genes, and their GenBank accession numbers is provided in Table 1. In this study, we selected only CDs of seventy-nine genes having perfect start and stop codons and devoid of any unknown bases in the middle of the sequence.
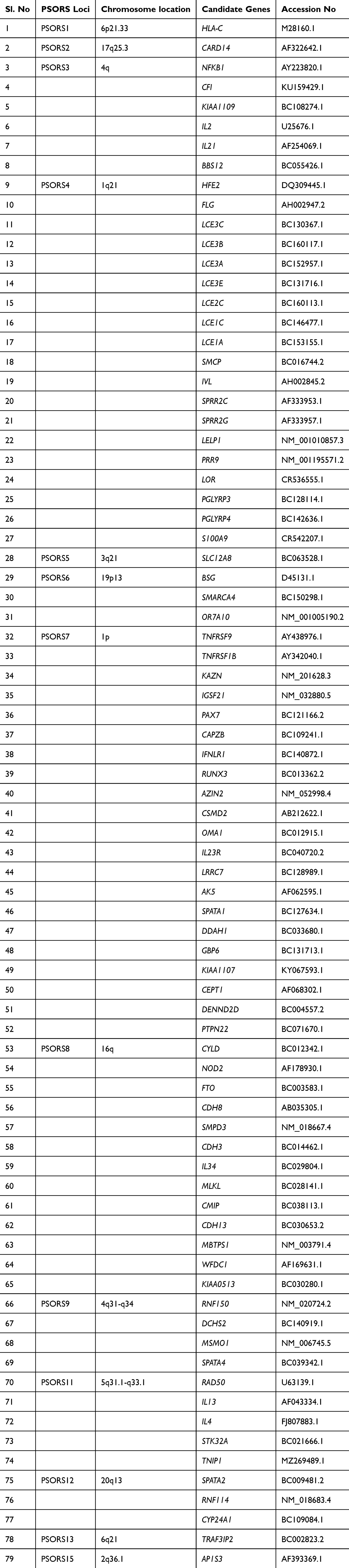 |
Table 1 List of Psoriasis Susceptibility Loci and Candidate Genes and Their Complete Coding Sequences Along with Accession Numbers Downloaded From NcbiGene Bank Database |
Nucleotide Composition
The overall percentage nucleotide composition (A%, T%, C% and G%) and the composition at the 3rd position of codon (A3%, C3%, G3%, and T3%) alongside GC content were assessed. Overall GC frequency at first (GC1%), second (GC2gh%), and third (GC3%) positions was also evaluated using MEGA (Molecular Evolutionary Genetics Analysis, Version 11) software (https://www.megasoftware.net/).14
Improved Effective Number of Codons (ENC)
The effective number of codons (ENC) is a measure used to quantify CUB within a gene or genome. It reflects the degree to which synonymous codons are used unevenly. Researchers use ENC and related measures to study codon usage patterns, evolutionary processes, and gene expression in various organisms.15,16 The ENC value ranges from 20 (indicating extreme bias, where only one codon is used per amino acid) to 61 (indicating no bias, where all synonymous codons are used equally).10 A lower ENC value indicates greater bias towards specific codons, while a higher ENC value suggests more even usage of synonymous codons. Previously, the ENC value was calculated through different mathematical equations. However, the current study evaluated the ENC values via an easily accessible online web portal at http://agnigarh.tezu.ernet.in/~ssankar/cub.php, as previously mentioned by Uddin et al.11
Relative Synonymous Codon Usage (RSCU)
Relative synonymous codon usage (RSCU) is another measure used to evaluate codon usage bias within a gene or genome. It compares the observed frequency of each synonymous codon to the expected frequency if all synonymous codons for a given amino acid were used equally.17 Researchers often use RSCU to identify preferred or optimal codons within a gene or genome, which can provide insights into translation efficiency, evolutionary processes, and gene expression regulation.18 Formerly, the sharp and li method19 was used to calculate RSCU values. However, the present study employed MEGA software (version 11.0) (https://www.megasoftware.net/) to determine RSCU values, as previously suggested by.14 RSCU values >1.0 and <1.0 signify high bias (higher frequency) and low bias (lower frequency), respectively. While the RSCU value equal to 1.0 shows no bias (equal codon preference). Similarly, the values >1.6 and <0.6 denote over- and under-presented codons, respectively.20
Correspondence Analysis (COA)
Correspondence analysis (COA) is a multivariate statistical analysis, most commonly used to understand the major trends in the codon usage pattern among genes.21,22 It allocates the codons in two axes, ie, axis1 and axis2. COA was performed using RSCU values of all coding sequences used in the study. Herein, each coding sequence is characterized as a 59-dimensional vector, where each dimension represents the RSCU value of a synonymous codon, excluding the non-synonymous codons, ie, TGG (tryptophan), ATG (methionine), and three-stop codons.11
Relative Abundance of Dinucleotide
The odd ratio value in terms of relative abundance of dinucleotide across the CDS of psoriasis-related genes was evaluated using DAMBE software.23 The odd ratio value greater than 1.23 and less than 0.78 is considered as over-represented and under-represented dinucleotide.24
Neutrality Plot Analysis
Neutrality plot analysis was used to detect the balance between mutation pressure and natural selection in influencing CUB. It was drawn between GC content at the first and second codon position (GC12) on the y-axis and GC content at the third codon position (GC3) on the x-axis. The regression coefficient closer to 0 with a narrow GC3 distribution indicates the dominant role of natural selection, while the slope of the regression line closer to 1 with a wide GC3 distribution indicates a dominant role of mutation pressure in influencing the CUB, suggesting complete neutrality.25
GRAVY Statistics
GRAVY (Grand average of hydropathy) score is calculated as the sum of the products of the frequency of each amino acid in a protein and its corresponding hydropathy index.26 A positive gravy value (> 0) signifies a hydrophobic protein, whereas a negative score (< 0) reflects the hydrophilic nature of the protein.27
Statistical Analysis
Correlation analysis was used to identify the relationship between overall nucleotide composition and each base at the 3rd position of the codon. All the statistical analyses were performed using SPSS 21 software (https://www.ibm.com/support/pages/spss-statistics-210-available-download).
Results
Nucleotide Composition in the Coding Sequence of Psoriasis-Related Susceptibility Genes
The nucleotide composition of the coding sequences (CDS) of psoriasis susceptibility candidate genes was analyzed (Table 2). Pyrimidine C showed the highest frequency (27.1±5.9%) compared to purine G (26.2±4.7%), purine A (25.4±6.0%), and pyrimidine T (21.3±4.8%). At the third codon position, C3 had the highest percentage (31.9±12.4%), followed by G3 (28.7±8.9%), T3 (20.9±8.6%), and A3 (18.5±8.7%). The CDS exhibited higher GC content (53.3±9.3%) than AT content (46.7±9.3%), with GC3% (60.6±16.1%) surpassing AT3% (39.4±16.1%). These findings suggest a preference for GC-ending codons, potentially impacting gene expression and protein function in psoriasis, particularly in genes involved in skin barrier function or immune regulation.28
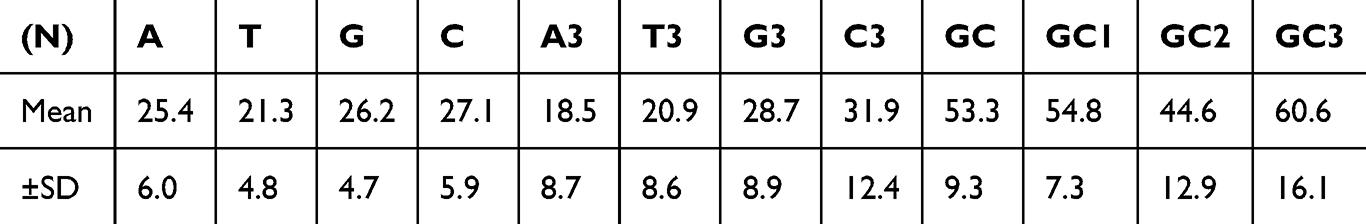 |
Table 2 Nucleotide Index (N) with Mean and Standard Deviation (SD) |
Codon Usage Bias of Candidate Genes Involved in Psoriasis Pathogenesis
The effective number of codons (ENC) was calculated to measure CUB, revealing an average ENC of 46.2 ± 9.9, indicating low codon bias compared to the standard threshold of 35.11 However, genes such as LCE3C, LCE3B, LCE3A, LCE3E, LCE1A, SPRR2C, and SPRR2G exhibited ENC values <30, suggesting high codon bias. These genes, particularly the LCE family, are associated with psoriasis susceptibility due to their role in skin barrier function. Their high codon bias suggests tightly regulated expression, which may be critical for their function in maintaining skin integrity and in the pathogenesis of psoriasis.7 Similarly, immune-related genes like IL23R, a key target for biologic therapies, showed distinct codon usage patterns that may influence expression efficiency, potentially affecting treatment outcomes.
Relationship Between ENC and Nucleotide Compositions
Regression analysis between ENC and nucleotide compositions showed positive coefficients for A, T, G, A3, and T3, and negative coefficients for C, C3, G3, and GC3 (Figure 1). Correlation analysis (Table 3) indicated positive and negative correlations among nucleotide compositions, suggesting that both mutation pressure and natural selection shape CUB. These patterns may influence the expression of genes critical to psoriasis pathways, such as IL-23/Th17 or TNF-α signaling, affecting disease severity or therapeutic response.
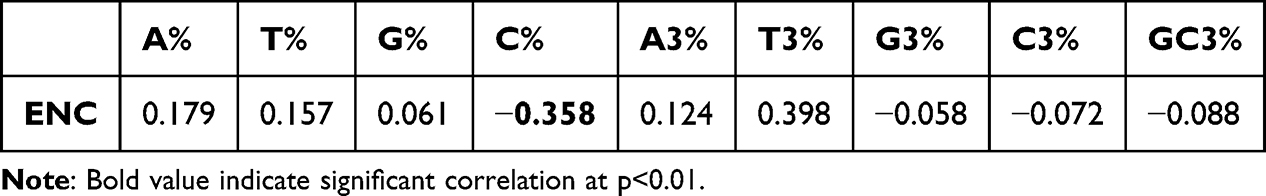 |
Table 3 Correlation Analysis Between Nc and Compositional Attributes of Candidate Genes. The Positive Correlation Between ENC and A%, T%, G%, A3%, T3% and Negative Correlation Between ENC and C%, G3%, C3% GC3% Suggested That Codon Usage Bias of Psoriasis Susceptible Candidate Genes are Influenced by Both Mutation Pressure and Translational Selection |
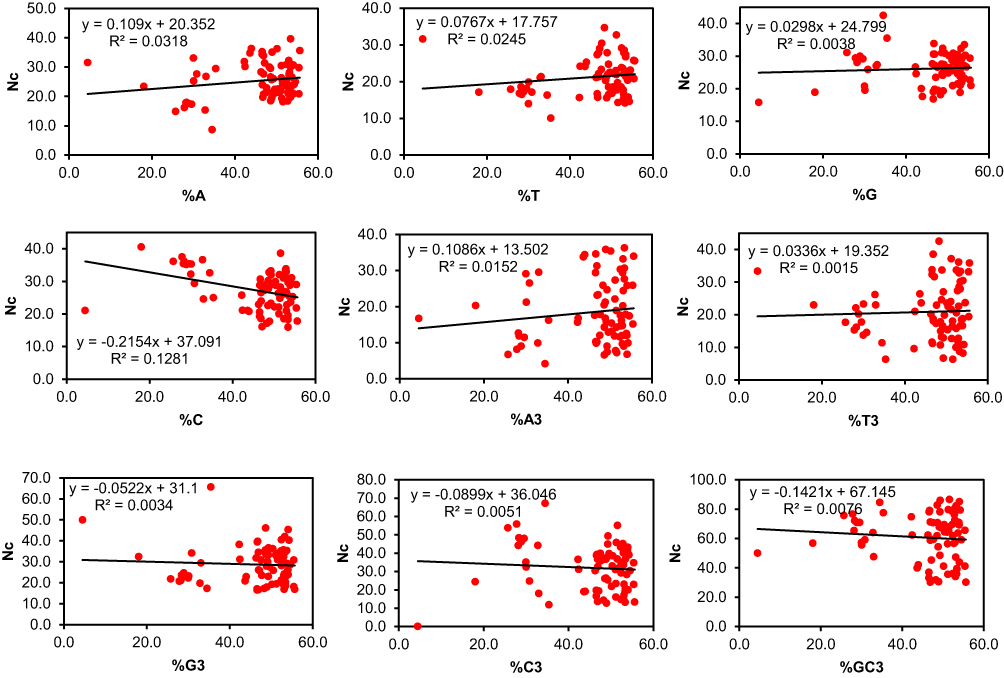 |
Figure 1 A regression plot was drawn between ENC and the Nucleotide composition. The regression coefficient was positive between ENC and A, T, G, A3, T3, while negative for C, G3, C3, and GC3. |
Relationship Between Overall Nucleotide Composition and Nucleotide Composition at the 3rd Codon Position
Correlation analysis between overall nucleotide composition (A%, T%, G%, C%, GC%) and third codon position nucleotides (A3%, T3%, G3%, C3%, GC3%) revealed significant positive correlations for homogeneous pairs (eg, T and T3, C and C3) and negative correlations for heterogeneous pairs (Table 4). These results indicate that mutation pressure and natural selection influence CUB, potentially affecting the expression of genes like IL23R, which are targeted by biologics in psoriasis treatment.
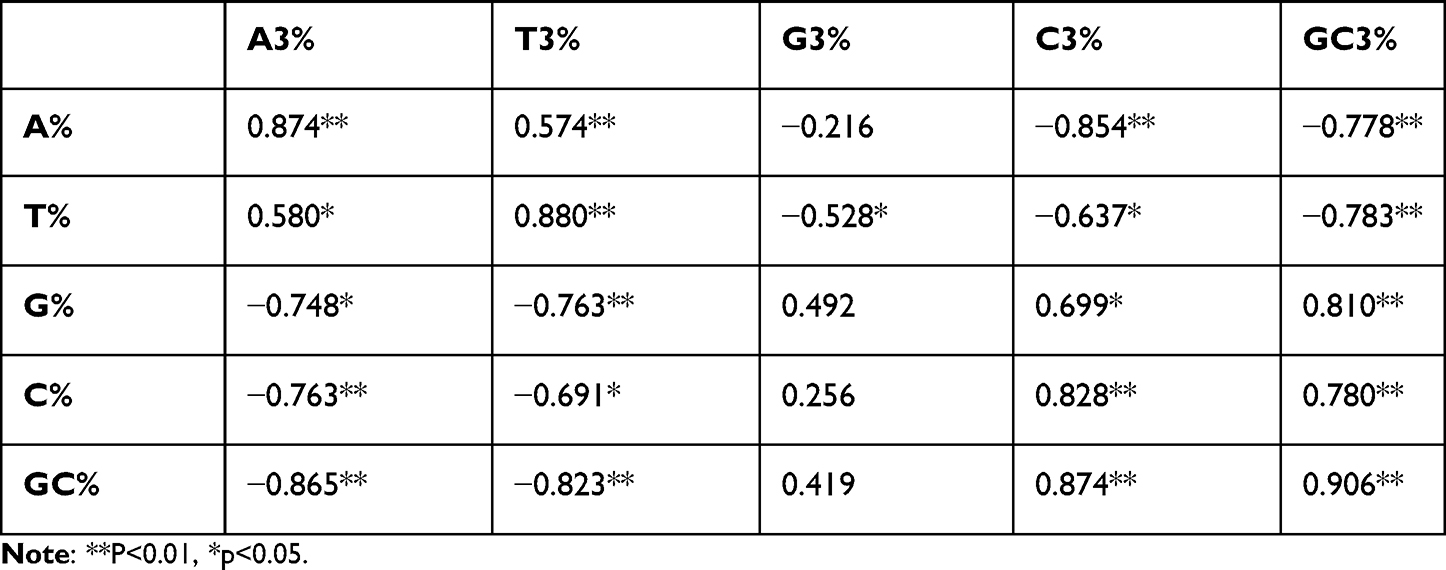 |
Table 4 Correlation Analysis Was Performed Between Composition of the Nucleotide and 3rd Position of Codon. The Significant Correlation Inferred That Compositional Constraints Under Mutation Pressure and Natural; Selection Shapes the Codon Usage of Selected Genes |
Relative Synonymous Codon Usage of Psoriasis Candidate Genes
Analysis of relative synonymous codon usage (RSCU) for 59 sense codons (excluding methionine, tryptophan, and stop codons) identified 32 preferred codons (RSCU>1.0), with C- or G-ending codons (eg, GCC, CTG, GTG) dominating over A- or T-ending codons (Table 5 and Figure 2). Overrepresented codons (RSCU>1.6) included GCC (alanine), CTG (leucine), and GTG (valine), which may enhance gene expression efficiency in psoriasis genes. These codons are prevalent in genes like LCE3B and IL23R, suggesting their tightly controlled expression may contribute to skin barrier dysfunction or immune dysregulation in psoriasis.
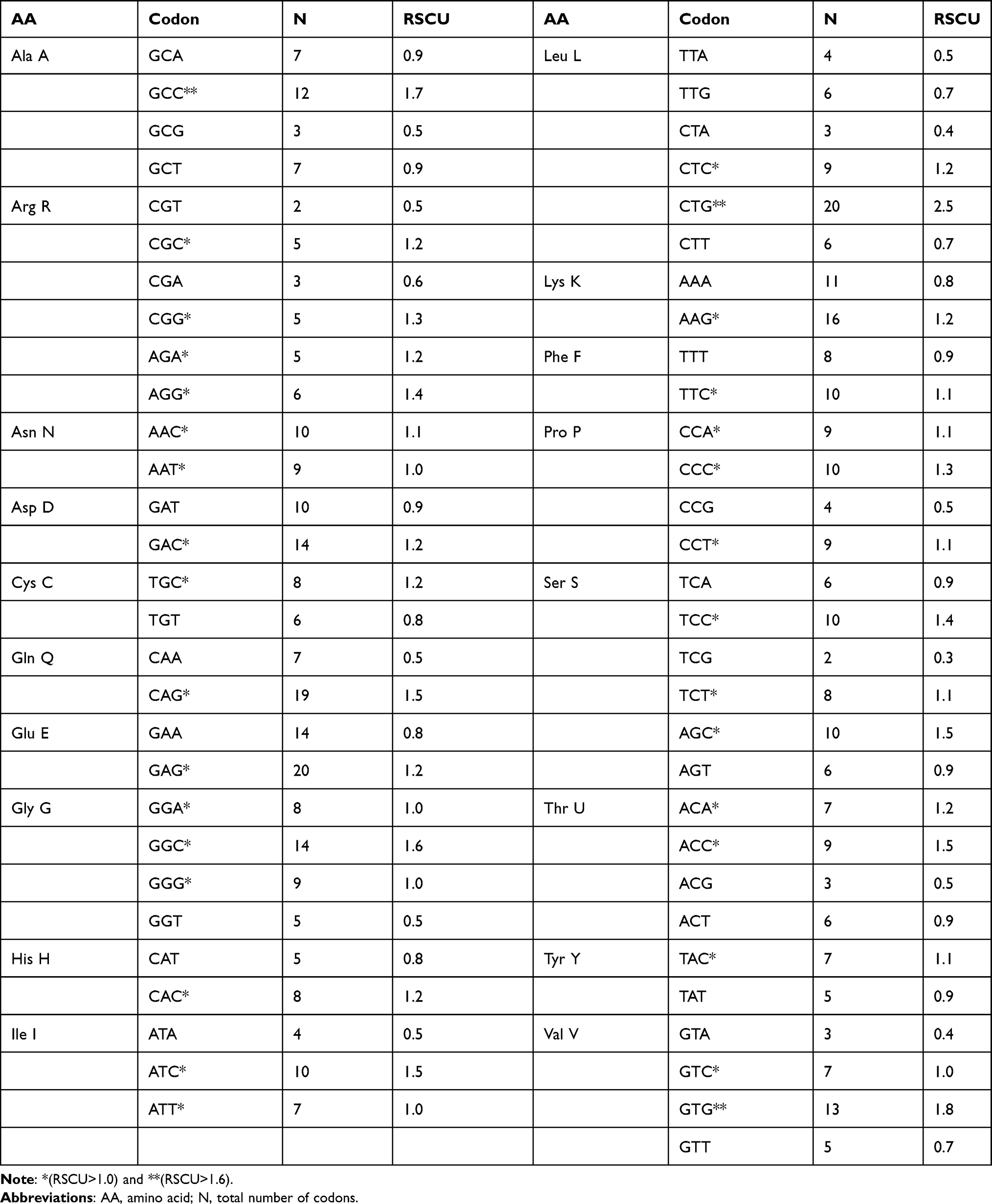 |
Table 5 Relative Synonymous Codon Usage (RSCU) Values for the 79 Psoriasis-Susceptibility Genes. RSCU > 1 Indicates Preferential Usage (Enrichment) of a Codon Within Its Synonymous Family, RSCU < 1 Indicates Under-Representation, and RSCU = 1 Denotes No Bias. The Single Amino Acid Encoding Codons Methionine and Tryptophan, and Three Stop Codons Were Excluded. TABLE 5 |
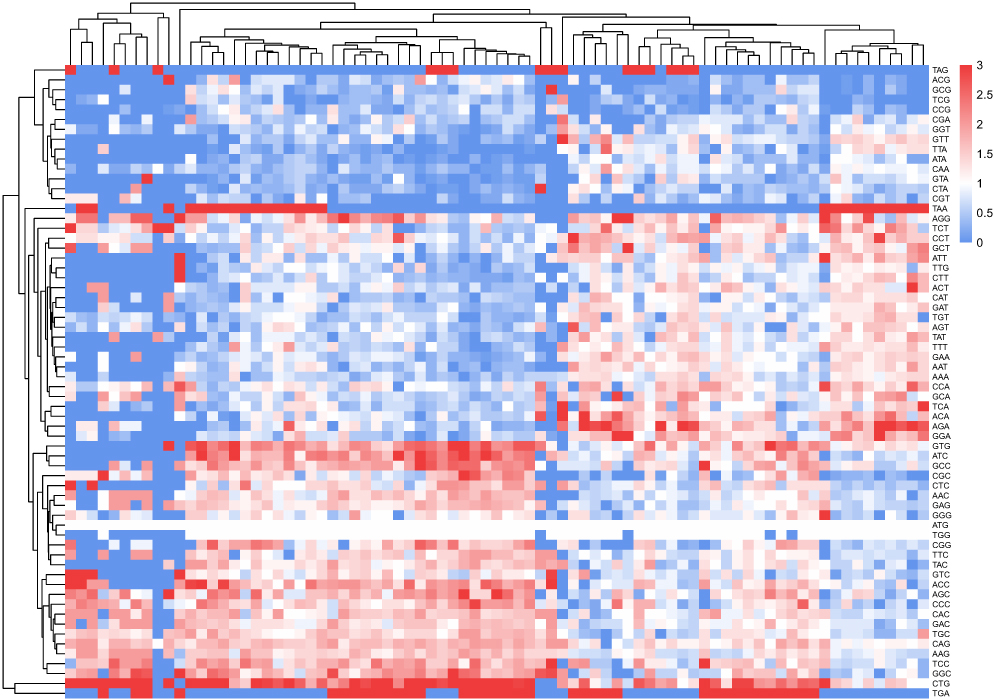 |
Figure 2 Heat map of RSCU values of each codon across the CDs of psoriasis-associated candidate genes. |
Correspondence Analysis (COA) of Psoriasis Susceptibility Genes
Correspondence analysis using RSCU values showed that codon usage varies across psoriasis genes, with Axis 1 contributing 23.76% and Axis 2 contributing 12.55% to total variation (Figure 3). Codons clustered near the axes indicate that nucleotide composition, influenced by mutation bias, correlates with CUB. This variation may affect the expression of genes in key psoriasis pathways, such as NF-κB, potentially influencing disease mechanisms.
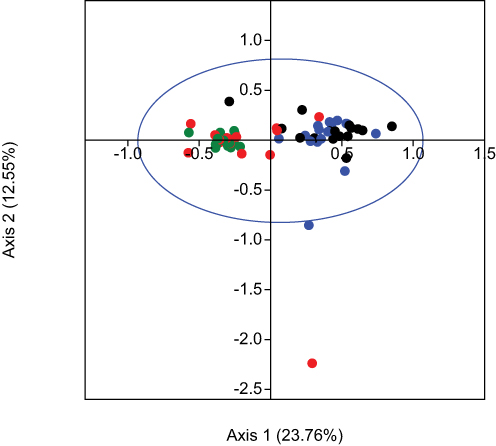 |
Figure 3 Correspondence analysis (CA) based on the RSCU values of the selected candidate genes of psoriasis. The first axis (Axis 1) and the second axis (Axis 2) values for each codon, generated by CA, were represented on a scatter plot. Axis 1 explained 23.76% of the total variance, while Axis 2 covered 12.55% of the total variance. Black, blue, red and green color indicate the codon ending with A, T, G and C, respectively. |
Relative Abundance of Dinucleotides
It was reported earlier that the frequency of dinucleotides can also influence codon usage bias.29 The frequencies of 16 abundant dinucleotide odd ratio values in each CDs of psoriasis gene were calculated (Supplementary File S1) and we represented them in a heat map (Figure 4). Dinucleotide analysis revealed overrepresentation of ApG, CpA, CpT, and TpG, and underrepresentation of CpG and TpA across the psoriasis gene CDS. These patterns may influence CUB and gene expression, potentially impacting proteins critical to psoriasis pathogenesis, such as those in the epidermal differentiation complex.
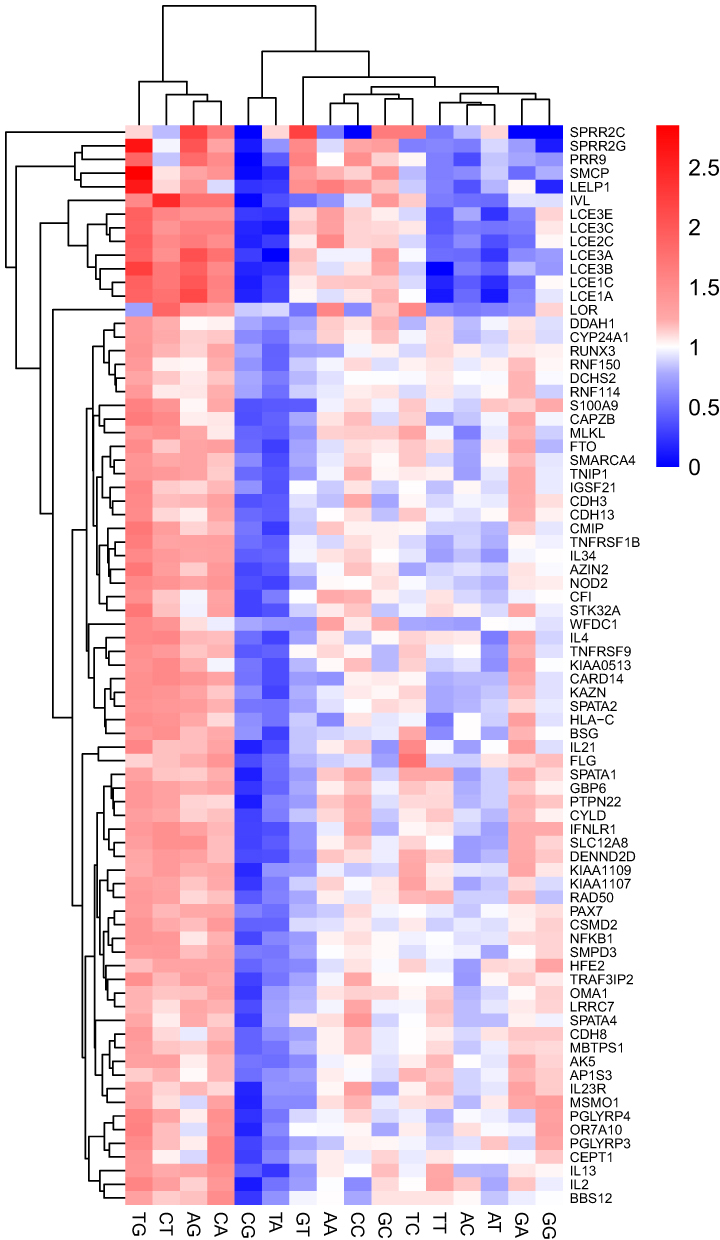 |
Figure 4 Heat map analysis of the odds ratio value in terms of the relative abundance of 16 dinucleotides in each CD of the gene. The value greater than 1.23 indicates over-represented while less than 0.78 as under-represented dinucleotide. |
Parity Plot Analysis
Parity rule 2 (PR2) plot analysis was performed to explore the impact of mutation and natural selection on the codon usage of the selected susceptibility genes of psoriasis. If the percentage of nucleotide A and T is equal to the percentage of C and G at the 3rd codon position, it indicates that mutation pressure alone affects the CUB of genes.30 On the contrary, natural selection may permit the inequality in the percentage of A and T bases to G and C.31 According to the parity rule 2 plot, no biases for mutation and natural selection occur at the centre of the coordinate (0.5, 0.5).25 However, the extent of deviation from the centre, that is, unequal distribution of nucleobases, might refer to the existence of biases for mutation and selection force.31 In our present study, we plotted the values of GC bias along the x‐axis and AT bias along the y‐axis (Figure 5), employing the parity rule 2 (PR2) bias plots. Here, we found that overall GC bias [G3/(G3 + C3)] and AT bias [A3/(A3 +T3)] were equal to 0.5, which suggests that there is no bias between mutation and selection pressure. However, disproportionate distribution towards the centre confirms the role of mutation pressure along with natural selection in the codon usage of the candidate genes of psoriasis.
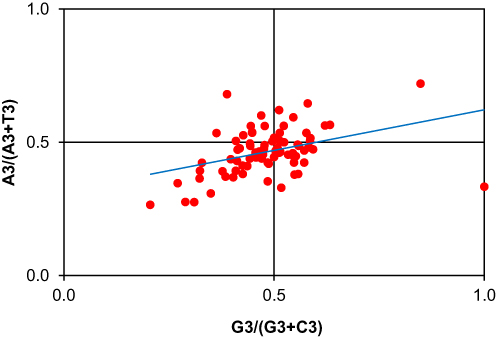 |
Figure 5 Parity plot showing the presence of AT bias [A3/(A3 + T3)] and GC bias [G3/(G3+ C3)] for the coding sequences of candidate genes of psoriasis. The centre of the plot, where the value of both coordinates is 0.5, indicates the place where there is no bias in mutation or selection rates. |
Neutrality Plot Among Psoriasis-Related Genes
The influence of mutation and natural selection on the candidate genes’ codon usage bias was assessed using a neutrality plot analysis (Figure 6). Nucleotide substitutions at the third codon position typically do not alter the encoded amino acid and are thus attributed to mutational pressure, whereas changes that result in amino acid substitutions are generally shaped by natural selection. To determine the impact of mutation pressure and natural selection in the codon usage pattern of the selected candidate genes, a neutrality plot, GC12 (average of GC1 and GC2) versus GC3 analysis was drawn. We observed a positive correlation between the values of GC12 and GC3. The slope of the linear regression was 0.290, representing that the mutation pressure attributed to 29% of the selection force in the coding sequences of psoriasis genes, while natural selection accounted for the residual 71%. Thus, the neutrality analysis indicated that natural selection dominated the forces in shaping the codon usage pattern of the psoriasis-relevant genes. The neutrality plot indicates the result of the equilibrium coefficient of mutation pressure and natural selection. Herein, points of the plot were diagonally dispersed, displaying a narrow range of GC3 distribution thus exhibiting that both mutation pressure and natural selection might affect the CUB in candidate genes.32
 |
Figure 6 Neutrality plot analysis of GC3 against GC12 for the CDs of psoriasis candidate genes. GC12 refer to the average of GC1 and GC2. The black line is the linear regression of GC12 versus GC3, with a regression coefficient of 0.290. |
Relationship Between GRAVY Values and ENC, GC, GC3s Values
Regression analysis was applied between GRAVY, ENC, GC, and GC3 values to find the association between codon usage bias and GRAVY in the codon usage patterns of the selected genes (Figure 7). Our analysis showed that GRAVY was positively correlated with GC and GC3 values, while negatively correlated with ENC values. The regression coefficient of the ENC on GRAVY was negative (−35.05), while the regression coefficient of GC% on GRAVY was positive (40.76) likely, the regression coefficient of GC3s on GRAVY was also positive (60.42). These results indicated that the general average hydropathicity is correlated with the codon usage variation in the candidate genes of psoriasis.
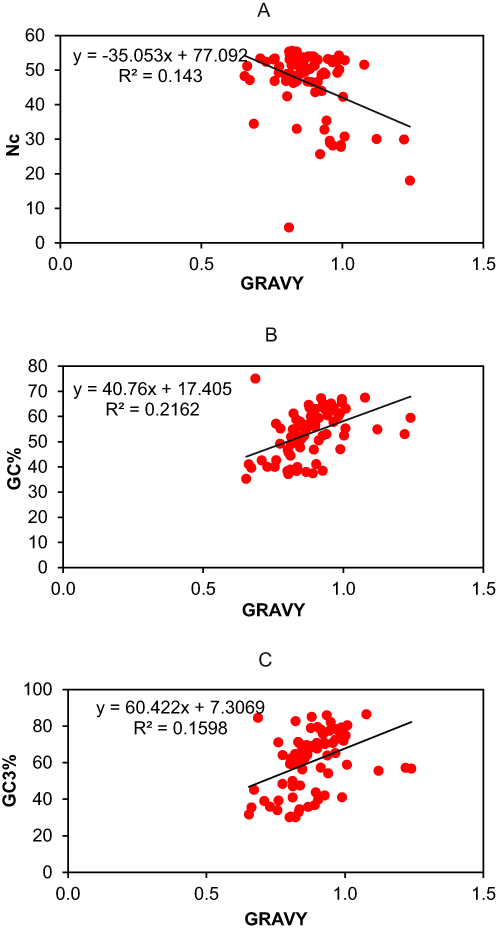 |
Figure 7 Regression analysis between (A) GRAVY and Nc, (B) GRAVY and GC%, (C) GRAVY and GC3%. The black line indicates the regression line. |
Functional and Genomic Profiling of Psoriasis-Related Genes
To gain deeper insight into the biological functions and regulatory architecture of psoriasis, we conducted functional enrichment and comparative genomic analyses of the 79 psoriasis-related genes. Using pathway enrichment analysis (Figure 8 and Table 6), we observed significant overrepresentation of these genes in several inflammation-associated pathways that are central to psoriasis pathophysiology.
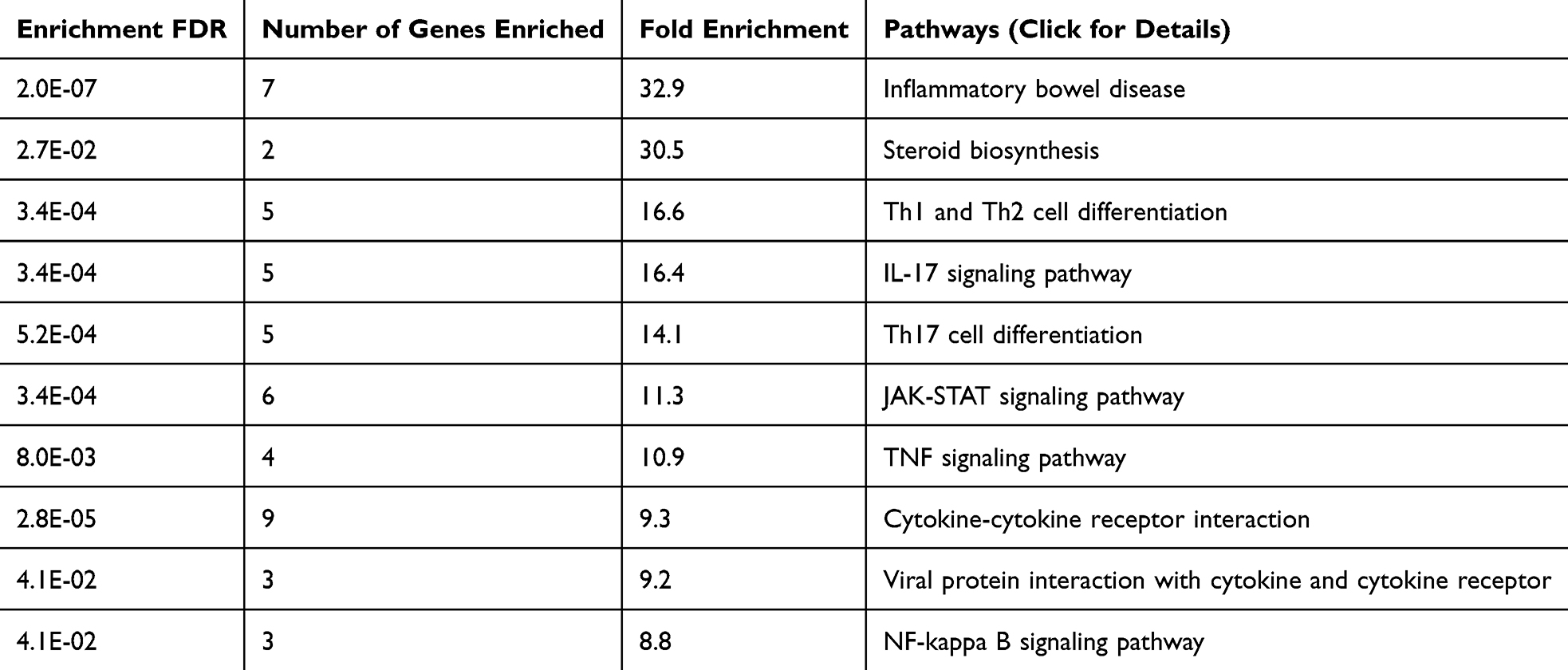 |
Table 6 Other Details of Pathway Enrichment Analysis of the Candidate Genes |
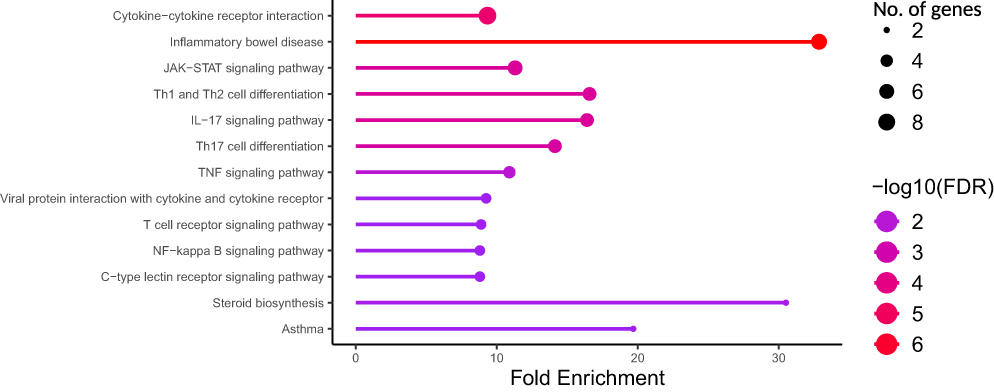 |
Figure 8 Pathway enrichment analysis of candidate genes. The figure shows significantly enriched pathways, including IL-17 signaling, JAK-STAT signaling, and TNF signaling, with their respective FDR values and fold enrichment. |
The most relevant significantly enriched pathway was the IL-17 signaling cascade (FDR = 3.4×10−3, fold enrichment = 16.4), which plays a central role in mediating keratinocyte activation, neutrophil recruitment, and sustained inflammatory responses characteristic of psoriatic plaques. This finding underscores the well-established importance of the Th17 axis in psoriasis and lends further genetic support to the effectiveness of IL-17-targeted therapies. In parallel, we observed strong enrichment in the JAK-STAT signaling pathway (FDR = 3.4×10−3, fold enrichment = 11.3), a major conduit for cytokine signaling involved in immune cell activation and proliferation. Genes in this pathway are known to contribute to the amplification of immune responses in psoriasis. Furthermore, the TNF signaling pathway (FDR = 8.0×10−³, fold enrichment = 10.9) was significantly enriched, reflecting the critical role of TNF-α as a master regulator of inflammatory cytokine networks in psoriatic skin.
To investigate whether these genes are also characterised by distinct genomic features that may influence their expression or regulation, we performed a comparative analysis of coding sequence (CDS) length, transcript length, 5’ untranslated region (5’UTR) length, 3’UTR length, and GC content. Compared to the genome-wide distribution, psoriasis-related genes exhibited a trend toward shorter coding regions and 5’UTRs, along with a higher GC content (Figure 9). While these differences did not reach statistical significance, the patterns observed may reflect selective pressures for efficient gene expression and mRNA stability in tissues affected by chronic inflammation. Notably, higher GC content has been associated with increased mRNA stability and enhanced translation, which may be particularly advantageous in the context of rapidly responding immune-related genes. The shorter 5’UTRs could also reduce regulatory complexity, enabling faster translation initiation under inflammatory stress. Together, these findings suggest that psoriasis-associated genes are not only functionally enriched in key immune pathways but also exhibit a genomic architecture that may support efficient, responsive expression in the disease context.
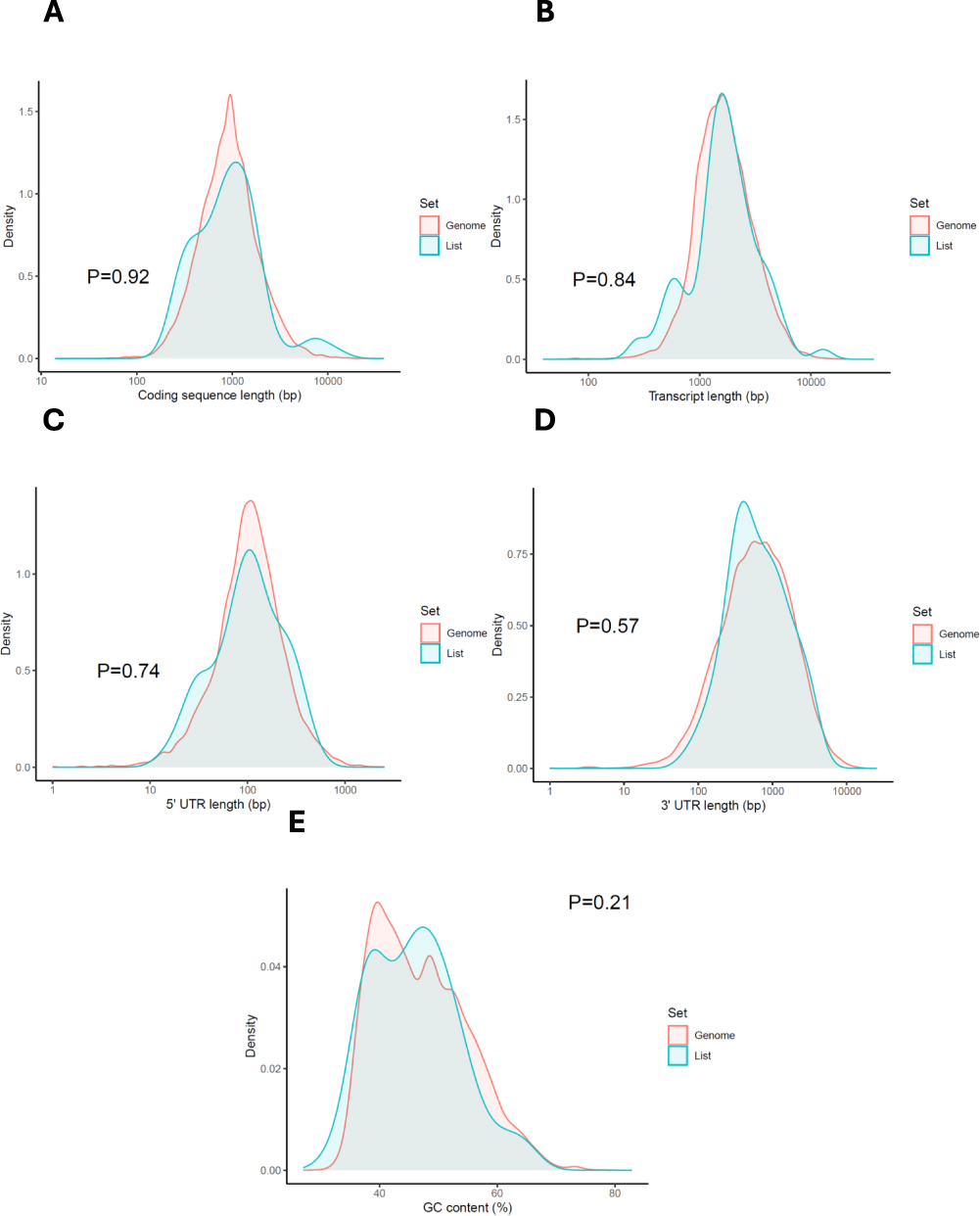 |
Figure 9 The characteristics of the candidate genes compared with the rest of the genome. Chi-squared and Student’s t-tests are run to see if the genes have special characteristics when compared with all the other genes within the genome (A–E). The “list” here denotes 79 psoriasis-related genes. |
Discussion
Several theories, including neutral theory and selection-mutation-drift theory, explain codon usage bias (CUB), which influences gene expression, protein folding, and function.33 Selection theory suggests that natural selection favours certain codons because they are translated more efficiently or accurately.17 In highly expressed genes, selection for optimal codons that match abundant tRNAs can improve translation speed and reduce errors.34 Similarly, different codons may have different mutation rates, leading to a biased codon usage pattern.35 Several other factors, such as gene length, gene expression, protein properties like aromaticity, hydrophobicity, and environmental stress, are also found to affect the CUB.36–38 Herein, the present study was conducted to explore the compositional constraints and patterns of codon usage in the impact of two major evolutionary forces, mutation and natural selection, in the psoriasis susceptible genes.
Nucleotide composition and the third position of codon play an important role in influencing the CUB of the genome.39 In our study, nucleobase pyrimidine C was found in higher frequency compared to purine G, while purine, in contrast to pyrimidine T. Similarly, C3 percentage (31.9±12.4%) was also the highest relative to G3 (28.7±8.9%), T3 (20.9±8.6%), and A3 (18.5±8.7%). Moreover, an overall greater percentage of GC content (53.3±9.3%) was noted in comparison to AT content (46.7±9.3%) in the gene CDS. Likewise, GC3% was also higher (60.6±16.1%), than that of AT3% (39.4±16.1%). These findings align with the previous studies that have also reported the majority of C or G ending codons (higher GC content than AT) and greater GC3 scores, indicating a high GC bias in oral cancer and human tumor suppressor genes.10,28 Studies also described that a higher GC3 percentage, G and C bases at the third position in the codon, is a crucial indicator of codon usage bias that is linked with enhanced gene expression.40 Besides, GC content is also supposed to influence the bendability, thermostability, and ability to convert the B form of DNA to the Z form. It is also found to be involved in the active process of transcription because it can keep the coding region in an open chromatin state.41 Presently, our study showed an average ENC value of 46.2 ± 9.9, higher than the 35 (standard value), indicating a low codon bias in the psoriasis gene. Similar findings were reported for genes associated with ovarian cancer, rotavirus VP7, and human papillomavirus type 51.11,42,43
On the contrary, a few psoriasis candidate genes, ie, LCE3C, LCE3B, LCE3A, LCE3E, LCE1A, SPRR2C, and SPRR2G exhibited a lower ENC value <30, suggesting high codon bias. Subsequently, a substantial correlation (p<0.001) was noted between overall nucleotide composition and the third position of codon in all CDS, indicating that the nucleotide composition bias affected by mutation pressure and translational selection majorly contributes to outlining the codon usage patterns of psoriasis susceptibility genes.44 Besides, a correlation between ENCs and base composition was observed, indicating that compositional constraints affect CUB. Relative synonymous codon usage value is used to determine over-represented (RSCU>1.6), under-represented (RSCU<0.6), more frequently used (RSCU>1.0) and less frequently used (RSCU<1.0) codons in the coding sequence of the genes.45,46 Our findings revealed thirty-two preferred codons (A-ending 4, T-ending 4, G-ending 8 and C-ending 16) used more frequently (RSCU>1.0). Consistent with these results, similar findings were documented for the EPB41L3 gene across various mammalian species, where 31 codons were reported to be frequently used, mainly with G/C ending.47 Besides, Over-represented codons (RSCU>1.6) were also found in the CDs of psoriasis genes, ie, GCC (alanine), CTG (leucine) and GTG (valine). These over-represented and under-represented codons play an important role in increasing or decreasing the gene expression levels, as previously described by Carlini et al.48 Correspondence analysis (COA) of RSCU values was conducted to find major trends in codon usage variation among psoriasis genes. We observed that the codon usage pattern of psoriasis genes was not uniform, and both natural selection and mutation may have affected the codon usage bias. Herein, Axis 1 contributed to total codon usage variation by 23.76% and Axis 2 by 12.55%, suggesting Axis 1 as the major effector of the CUB. Additionally, the codons were closer to axes, representing a concentrated distribution. Comparable findings were stated by Uddin,49 where CoA showed 50.33% contribution to axis 1 and 9.50% of axis 2, and codons were close to axes, hence indicating that mutation bias might influence the codon bias.
The overall relative abundance of dinucleotide odds ratio values indicated that four dinucleotides—ApG, CpA, CpT, and TpG—were over-represented, while two dinucleotides, CpG and TpA, were under-represented across the coding regions of psoriasis genes. Similarly, Khandia et al13 reported that four dinucleotides (ApG, GpA, CpT, and TpG) were over-represented in genes related to human depression, while three dinucleotides (CpG, TpA, and GpT) were under-represented.13 The over-represented dinucleotides found across different organisms may be attributed to the spontaneous mutation of methylated cytosine (C) in the CpG dinucleotide, which results in thymine (T) and leads to the formation of TpG dinucleotides on the existing strand, as well as CpA dinucleotides on the reverse strand during DNA replication.50
The Parity Rule 2 (PR2) plot was also plotted by taking the GC bias values along the x‐axis and AT bias along the y‐axis. An equal (0.5) overall GC bias and AT bias value suggests no bias between mutation and selection pressure. However, disproportionate distribution towards the centre confirms the role of mutation pressure along with natural selection in the CUB of the psoriasis genes. These results are in line with those of Li et al51 who reported parallel Parity rule 2 analysis outcomes in mitochondrial genes of Japanagallia species. Moving forward, earlier studies have divulged that codon usage patterns of human genes are influenced by mutation pressure and that codon bias exhibits a strong relation with the GC contents.52 Therefore, to further explore the relationship between codon bias and mutation pressure, a neutrality plot was drawn between GC12 and GC3 values. The linear regression analysis resulted in a coefficient value of 0.290, representing that the mutation pressure accounted for 29% of the selection force and natural selection 71%. This shows that natural selection dominates and affects the codon usage bias of the psoriasis genes. Also, the plot points were diagonally dispersed, exhibiting a narrow range of GC3 distribution, showing that both mutation pressure and natural selection may play a major role in shaping the patterns of codon usage, similar to a study by Chakraborty et al50 where CUB analysis of chloroplast genes in Oryza species was conducted. The regression coefficient value of the neutrality plot was observed to be < 0.5, and points on the plot were also diagonally dispersed, indicating a narrow GC3 distribution. In the same way, neutrality plot results were observed by Uddin et al,32 while doing CUB analysis of ATP6 and ATP8 genes. Lastly, biochemical properties of the psoriasis susceptibility genes were also assessed through the Gravy score, as they may also influence the CUB of genes.53 Gravy particularly determines the hydrophobic (positive value) or hydrophilic (negative value) nature of the protein.54 The current analysis showed that GRAVY was positively correlated with GC and GC3s values, while negatively correlated with ENC values of psoriasis genes. The regression coefficient value of ENC and GRAVY was −35.05, GC% and GRAVY were 40.76, while GC3s and GRAVY were 60.42. This declares that gravy is closely related to the patterns of codon usage of genes, as formerly observed by Mazumder et al10 in human oral cancer genes.
The pathway enrichment analysis provides critical insights into the functional roles of the candidate genes, revealing their involvement in key psoriasis-related pathways such as IL-17 signaling, JAK-STAT signaling, and TNF signaling. These pathways are central to the immunological mechanisms of psoriasis and are targeted by current biologic therapies, such as IL-17 inhibitors and JAK inhibitors.55 The observed codon usage patterns, especially the preference for GC-ending codons and low ENC values in genes like IL23R and LCE3C, may enhance the expression efficiency of these pathway components, thereby influencing disease severity and therapeutic response. Furthermore, the comparison of candidate genes with the whole genome demonstrates that these genes exhibit distinct codon usage characteristics, including higher GC content and lower ENC values, and suggest that natural selection has shaped the codon usage of psoriasis-related genes to optimize their function in the disease context, potentially enhancing mRNA stability and translation efficiency in the skin environment where psoriasis manifests.56 These findings validate the role of codon usage in psoriasis and open avenues for exploring how genetic variations in codon usage might influence disease susceptibility and progression. While a direct correlation analysis with clinical outcomes was beyond the scope of this study, prior research using the GSE13355 dataset has shown that differentially expressed genes in psoriasis, including those with high codon usage bias, are associated with clinical outcomes such as disease severity (eg, PASI scores) and treatment response.57 For instance, genes with lower ENC values may exhibit higher expression in lesional skin, correlating with increased disease severity. These findings suggest that codon usage patterns could serve as potential biomarkers for predicting clinical outcomes in psoriasis patients, warranting further investigation in larger cohorts.
The primary limitation of this study is its focus on a curated panel of 79 psoriasis-susceptibility genes, which restricts conclusions to the gene-set level and may not fully apply to individual loci. The computational analyses conducted do not directly establish causal effects on mRNA stability or translation. Additionally, clinical phenotypes, such as PASI scores or treatment responses, were not incorporated, leaving the potential connections between codon-usage patterns and disease burden or therapeutic outcomes unexplored. These findings are primarily hypothesis-generating and require validation through experimental systems and clinically annotated cohorts. While the gene-set analysis highlights codon-usage patterns indicative of differential expression potential, experimental approaches, such as reporter assays and ribosome profiling in inflammatory skin models, are necessary to confirm causal effects on mRNA stability and translation. Furthermore, prospective studies integrating codon-bias metrics with clinical phenotypes, including PASI severity and standardized treatment-response measures, will be critical to determine whether codon-usage bias is linked to disease burden or therapeutic outcomes. In conclusion, our study demonstrates that psoriasis susceptibility genes exhibit a low overall codon bias (ENC = 46.2 ± 9.9), with a preference for GC-ending codons (eg, GCC, CTG, GTG), which likely influence gene expression patterns. Specific genes, such as LCE3C and IL23R, show high codon bias, suggesting tightly regulated expression that may contribute to skin barrier dysfunction or immune dysregulation in psoriasis. Both mutation pressure and natural selection shape CUB, with natural selection playing a dominant role (71%). These findings provide an initial understanding of codon usage in psoriasis-related genes, which could serve as a basis for future research into how genetic variations at the codon level influence gene expression and disease phenotype. This work lays the groundwork for future translational studies exploring CUB as a tool for advancing therapeutic strategies in psoriasis.
Limitations
This analysis focuses on a curated set of 79 psoriasis-susceptibility genes, which is modest in size and may introduce selection bias because inclusion is conditioned on prior locus and gene curation. Accordingly, all inferences are intended at the gene-set level and should not be over-interpreted as evidence for optimization or selection acting on individual genes. To mitigate bias and provide context, we benchmarked codon-usage measures for the psoriasis gene set against genome-wide backgrounds. We quantified codon usage with the effective number of codons (ENC) and relative synonymous codon usage (RSCU), summarized multivariate structure via correspondence analysis (COA) on RSCU matrices, and examined GC3-based neutrality plots to consider the relative contributions of mutational pressure versus putative selection. Given the panel size, effect estimates and distributional differences should be interpreted cautiously, and conclusions are framed as patterns consistent with codon-usage shifts rather than definitive causal claims.
Data Sharing Statement
This study exclusively utilized publicly available data, and the sources and handling of these data are thoroughly described in the Materials and Methods section. The coding sequences (CDS) of psoriasis-susceptible candidate genes having perfect start and stop codons were retrieved from the National Centre for Biotechnology Information, USA (http://www.ncbi.nlm.nih.gov/). A comprehensive list of these psoriasis susceptibility loci, candidate genes, and their GenBank accession numbers is provided in Table 1. Additional information and details can be obtained from the two corresponding author upon request.
Ethics Declarations
This study used de-identified, publicly available human data and involved no interaction with individuals. Under the Measures for Ethical Review of Life Sciences and Medical Research Involving Humans (National Health Commission, Ministry of Education, Ministry of Science and Technology, and National Administration of Traditional Chinese Medicine; 18 Feb 2023), Article 32, items (1) and (2) permit exemption from ethics review for research using lawfully obtained public data or anonymized information, provided no harm, sensitive personal information, or commercial interests are involved. Our study meets these conditions; therefore, it qualifies for ethics-review exemption.
Acknowledgments
Thanks to Shaanxi Provincial Hospital of Traditional Chinese Medicine, Xi’an, and Shaanxi University of Chinese Medicine, Xianyang, China, for providing the necessary facilities in carrying out this research work.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
Funding: Supported by the National Natural Science Foundation of China (No. 82174386), the Key Industry Innovation Chain Project of Shaanxi Province (No. 2021ZDLSF04-12), Xi’an Science and Technology Project (No. 24YXYJ0002).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Beera AM, Nori LP. Psoriasis: a non-communicable chronic autoimmune inflammatory skin disorder, affecting the quality of life. Scopus Indexed. 2023;16(6):7081–7092. doi:10.37285/ijpsn.2023.16.6.8
2. Chen ML, Kao WM, Huang JY, Hung YM, Wei JCC. Human papillomavirus infection associated with increased risk of new-onset psoriasis: a nationwide population-based cohort study. Int J Epidemiol. 2020;49(3):786–797. doi:10.1093/ije/dyaa027
3. Zhao F, Zhao J, Wei K, et al. Targeted siRNA therapy for psoriasis: translating preclinical potential into clinical treatments. Immunotargets Ther. 2024;13:259–271. doi:10.2147/ITT.S458800
4. Wang Y, Cui Y, Zheng Z. Adalimumab-induced erythrodermic psoriasis associated with alopecia in a patient with palmoplantar pustulosis. Indian J Dermatol. 2024;69(2):178–180. doi:10.4103/ijd.ijd_155_23
5. Dand N, Mahil SK, Capon F, Smith CH, Simpson MA, Barker JN. Psoriasis and genetics. Acta Derm Venereol. 100(3):adv00030.
6. Sun LD. research progress of genomic variation in psoriasis. Int J Dermatol Venereol. 2022;5(4):207. doi:10.1097/JD9.0000000000000276
7. Patel HA, Revankar RR, Pedroza ST, Graham S, Feldman SR. the genetic susceptibility to psoriasis and the relationship of linked genes to our treatment options. Int J Mol Sci. 2023;24(15):12310. doi:10.3390/ijms241512310
8. Rossi S, Richards EL, Orozco G, Eyre S. functional genomics in psoriasis. Int J Mol Sci. 2024;25(13):7349. doi:10.3390/ijms25137349
9. Liu Y, Yang Q, Zhao F. Synonymous but not silent: the codon usage code for gene expression and protein folding. Annu Rev Biochem. 2021;90:375–401. doi:10.1146/annurev-biochem-071320-112701
10. Mazumder TH, Uddin A. Understanding the nucleotide composition and patterns of codon usage in the expression of human oral cancer genes. Mutat Res. 2024;829:111880. doi:10.1016/j.mrfmmm.2024.111880
11. Uddin A, Paul N, Chakraborty S. The codon usage pattern of genes involved in ovarian cancer. Ann N Y Acad Sci. 2019;1440(1):67–78. doi:10.1111/nyas.14019
12. Liu XY, Li Y, Ji KK, et al. Genome-wide codon usage pattern analysis reveals the correlation between codon usage bias and gene expression in Cuscuta australis. Genomics. 2020;112(4):2695–2702. doi:10.1016/j.ygeno.2020.03.002
13. Khandia R, Gurjar P, Kamal MA, Greig NH. Relative synonymous codon usage and codon pair analysis of depression associated genes. Sci Rep. 2024;14(1):3502. doi:10.1038/s41598-024-51909-8
14. Xing B, Chen X, Wang Y, Sun R, Zhang Z, Lin H. The complete mitochondrial genome of capitulum mitella with characterization and phylogenetic implications. Russ J Genet. 2023;59(10):1032–1043. doi:10.1134/S1022795423100149
15. Comeron JM. Selective and mutational patterns associated with gene expression in humans: influences on synonymous composition and intron presence. Genetics. 2004;167(3):1293–1304. doi:10.1534/genetics.104.026351
16. Wright F. The “effective number of codons” used in a gene. Gene. 1990;87(1):23–29. doi:10.1016/0378-1119(90)90491-9
17. Moutinho AF, Eyre-Walker A. No evidence that selection on synonymous codon usage affects patterns of protein evolution in bacteria. Genome Biol Evol. 2024;16(2):evad232. doi:10.1093/gbe/evad232
18. Bahiri-Elitzur S, Tuller T. Codon-based indices for modeling gene expression and transcript evolution. Comput Struct Biotechnol J. 2021;19:2646–2663. doi:10.1016/j.csbj.2021.04.042
19. Sharp PM, Li WH. An evolutionary perspective on synonymous codon usage in unicellular organisms. J Mol Evol. 1986;24(1–2):28–38. doi:10.1007/BF02099948
20. Qi X, Wei C, Li Y, et al. The characteristic of the synonymous codon usage and phylogenetic analysis of hepatitis B virus. Genes Genomics. 2020;42(7):805–815. doi:10.1007/s13258-020-00932-w
21. Grantham R, Gautier C, Gouy M. Codon frequencies in 119 individual genes confirm consistent choices of degenerate bases according to genome type. Nucleic Acids Res. 1980;8(9):1893–1912. doi:10.1093/nar/8.9.1893
22. Shields DC, Sharp PM. Synonymous codon usage in Bacillus subtilis reflects both translational selection and mutational biases. Nucl Acids Res. 1987;15(19):8023–8040. doi:10.1093/nar/15.19.8023
23. Xia X. DAMBE6: new tools for microbial genomics, phylogenetics, and molecular evolution. J Heredity. 2017;108(4):431–437. doi:10.1093/jhered/esx033
24. Xu Q, Cao J, Rai KR, Zhu B, Liu D, Wan C. Codon usage bias of goose circovirus and its adaptation to host. Poult Sci. 2024;103(7):103775. doi:10.1016/j.psj.2024.103775
25. Sueoka N. Directional mutation pressure and neutral molecular evolution. Proc Natl Acad Sci USA. 1988;85(8):2653–2657. doi:10.1073/pnas.85.8.2653
26. Kyte J, Doolittle RF. A simple method for displaying the hydropathic character of a protein. J Mol Biol. 1982;157(1):105–132. doi:10.1016/0022-2836(82)90515-0
27. Wang H, Zhong H, Gao C, Zang J, Yang D. The distinct properties of the consecutive disordered regions inside or outside protein domains and their functional significance. Int J Mol Sci. 2021;22(19):10677. doi:10.3390/ijms221910677
28. Mazumder TH, Uddin A, Chakraborty S. Insights into the nucleotide composition and codon usage pattern of human tumor suppressor genes. Mol, Carcinog. 2020;59(1):15–23. doi:10.1002/mc.23124
29. Kokate PP, Techtmann SM, Werner T. Codon usage bias and dinucleotide preference in 29 Drosophila species. G3. G3. 2021;11(8):jkab191. doi:10.1093/g3journal/jkab191
30. Kawabe A, Miyashita NT. Patterns of codon usage bias in three dicot and four monocot plant species. Genes Genet Syst. 2003;78(5):343–352. doi:10.1266/ggs.78.343
31. Sueoka N. Intrastrand parity rules of DNA base composition and usage biases of synonymous codons. J Mol Evol. 1995;40(3):318–325. doi:10.1007/BF00163236
32. Uddin A, Mazumder TH, Barbhuiya PA, Chakraborty S. Similarities and dissimilarities of codon usage in mitochondrial ATP genes among fishes, aves, and mammals. IUBMB Life. 2020;72(5):899–914. doi:10.1002/iub.2231
33. Yiwen T, Zhang M, Yu F, Wu Z. Compositional dynamics and codon usage pattern of myogenin (MyoG) gene across different mammalian species. Biol Bull Russ Acad Sci. 2023;50(S4):S577–88. doi:10.1134/S1062359023604068
34. Victor MP, Acharya D, Chakraborty S, Ghosh TC. The combined influence of codon composition and tRNA copy number regulates translational efficiency by influencing synonymous nucleotide substitution. Gene. 2020;745:144640. doi:10.1016/j.gene.2020.144640
35. Yang X, Luo X, Cai X. Analysis of codon usage pattern in Taenia saginata based on a transcriptome dataset. Parasites Vectors. 2014;7(1):527. doi:10.1186/s13071-014-0527-1
36. Berg OG. Selection intensity for codon bias and the effective population size of escherichia coli. Genetics. 1996;142(4):1379–1382. doi:10.1093/genetics/142.4.1379
37. Orešič M, Dehn MHH, Korenblum DHH, Shalloway DHH. Tracing specific synonymous codon-secondary structure correlations through evolution. J Mol Evol. 2003;56(4):473–484. doi:10.1007/s00239-002-2418-x
38. Prat Y, Fromer M, Linial N, Linial M. Codon usage is associated with the evolutionary age of genes in metazoan genomes. BMC Evol Biol. 2009;9(1):285. doi:10.1186/1471-2148-9-285
39. Jenkins GM, Holmes EC. The extent of codon usage bias in human RNA viruses and its evolutionary origin. Virus Res. 2003;92(1):1–7. doi:10.1016/S0168-1702(02)00309-X
40. Bourret J, Borvető F, Bravo IG. Subfunctionalisation of paralogous genes and evolution of differential codon usage preferences: the showcase of polypyrimidine tract binding proteins. J Evol Biol. 2023;36(10):1375–1392. doi:10.1111/jeb.14212
41. Zhang DH, Fujimoto T, Saxena S, Yu HQ, Miyoshi D, Sugimoto N. Monomorphic RNA G-quadruplex and polymorphic DNA G-quadruplex structures responding to cellular environmental factors. Biochemistry. 2010;49(21):4554–4563. doi:10.1021/bi1002822
42. Liu Z, Zhao S, Jin X, Wen X, Ran X. Host and structure-specific codon usage of G genotype (VP7) among group A rotaviruses. Front Vet Sci. 2024;11:1438243. doi:10.3389/fvets.2024.1438243
43. Tan X, Bao S, Lu X, Lu B, Shen W, Jiang C. Comprehensive analysis of codon usage bias in human papillomavirus type 51. Pol J Microbiol. 2024;73(4):455–465. doi:10.33073/pjm-2024-036
44. Deb B, Uddin A, Chakraborty S. Composition, codon usage pattern, protein properties, and influencing factors in the genomes of members of the family Anelloviridae. Arch Virol. 2021;166(2):461–474. doi:10.1007/s00705-020-04890-2
45. De Mandal S, Mazumder TH, Panda AK, Kumar NS, Jin F. Analysis of synonymous codon usage patterns of HPRT1 gene across twelve mammalian species. Genomics. 2020;112(1):304–311. doi:10.1016/j.ygeno.2019.02.010
46. Yu X, Liu J, Li H, Liu B, Zhao B, Ning Z. Comprehensive analysis of synonymous codon usage patterns and influencing factors of porcine epidemic diarrhea virus. Arch Virol. 2021;166(1):157–165. doi:10.1007/s00705-020-04857-3
47. Kumar U, Khandia R, Singhal S, et al. Insight into codon utilization pattern of tumor suppressor gene EPB41L3 from different mammalian species indicates dominant role of selection force. Cancers. 2021;13(11):2739. doi:10.3390/cancers13112739
48. Carlini DB, Chen Y, Stephan W. The relationship between third-codon position nucleotide content, codon bias, mRNA secondary structure and gene expression in the drosophilid alcohol dehydrogenase genes adh and adhr. Genetics. 159(2):623–633.
49. Uddin A. Compositional features and codon usage pattern of genes associated with anxiety in human. Mol Neurobiol. 2020;57(12):4911–4920. doi:10.1007/s12035-020-02068-0
50. Chakraborty S, Yengkhom S, Uddin A. Analysis of codon usage bias of chloroplast genes in Oryza species: codon usage of chloroplast genes in Oryza species. Planta. 2020;252(4):67. doi:10.1007/s00425-020-03470-7
51. Li M, Wang J, Dai R, Smagghe G, Wang X, You S. Comparative analysis of codon usage patterns and phylogenetic implications of five mitochondrial genomes of the genus Japanagallia Ishihara, 1955 (Hemiptera, Cicadellidae, Megophthalminae). PeerJ. 2023;11:e16058. doi:10.7717/peerj.16058
52. Shackelton LA, Parrish CR, Holmes EC. Evolutionary basis of codon usage and nucleotide composition bias in vertebrate DNA viruses. J Mol Evol. 2006;62(5):551–563. doi:10.1007/s00239-005-0221-1
53. Jamil Z, Uddin A, Alam SSM, et al. Analysis of the compositional features and codon usage pattern of genes involved in human autophagy. Cells. 2022;11(20):3203. doi:10.3390/cells11203203
54. Huang HJ, Chen WY, Wu JH. Total protein extraction for metaproteomics analysis of methane producing biofilm: the effects of detergents. Int J Mol Sci. 2014;15(6):10169–10184. doi:10.3390/ijms150610169
55. Nair RP, Duffin KC, Helms C, et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-kappaB pathways. Nat Genet. 2009;41(2):199–204. doi:10.1038/ng.311
56. Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28(1):27–30. doi:10.1093/nar/28.1.27
57. Swindell WR, Johnston A, Voorhees JJ, Elder JT, Gudjonsson JE. Dissecting the psoriasis transcriptome: inflammatory- and cytokine-driven gene expression in lesions from 163 patients. BMC Genomics. 2013;14(1):527. doi:10.1186/1471-2164-14-527
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.