Back to Journals » Infection and Drug Resistance » Volume 14
Clonal Diversity, Low-Level and Heterogeneous Oxacillin Resistance of Oxacillin Sensitive MRSA
Authors Liu R, Zhang J, Du X, Lv Y, Gao X, Wang Y, Wang J
Received 30 October 2020
Accepted for publication 31 January 2021
Published 19 February 2021 Volume 2021:14 Pages 661—669
DOI https://doi.org/10.2147/IDR.S288991
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Roushan Liu,1,* Jian Zhang,2,* Xiaoli Du,3 Yingying Lv,1 Xiangyu Gao,1 Yanyan Wang,1 Junrui Wang1
1Department of Laboratory Medicine, Affiliated Hospital of Inner Mongolian Medical University, Hohhot, 010050, People’s Republic of China; 2Department of Laboratory Medicine, Bayannur People’s Hospital, Bayannur City, 015000, People’s Republic of China; 3National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention, Beijing, 102206, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Junrui Wang
Department of Laboratory Medicine, Affiliated Hospital of Inner Mongolian Medical University, Hohhot, 010050, People’s Republic of China
Tel +86 13347104892
Email [email protected]
Purpose: This study investigates the phenotypic and genotypic resistance features of OS-MRSA clinical isolates and their distinctive sensitivities to oxacillin.
Methods: 1200 clinical isolates of Staphylococcus aureus were enrolled in this study. Automated antibiotics susceptibility tests on VITEK-2 and BD Phoenix-100TM, cefoxitin disc diffusion method, oxacillin broth microdilution method, mecA, and mecC gene detection were performed to identify OS-MRSA. MLST, PFGE, SCCmec, and spa typing methods were employed to determine genotypes of OS-MRSA isolates. Heterogeneous resistance of OS-MRSA isolates was detected using the population analysis profiling method, and PBP2a latex agglutination assay was used to detect the expression of PBP2a protein for 14 OS-MRSA isolates and their highly resistant subpopulations.
Results: A total of 14 OS-MRSA isolates (1.17%, 14/1200) were identified, and all of the isolates were confirmed to be positive with the mecA gene and negative with the mecC gene. All of the 14 OS-MRSA isolates were identified as MSSA by VITEK-2, BD Phonenix-100, and oxacillin broth microdilution methods, while 21.43% (3/14) isolates were determined to be MRSA by the cefoxitin disk diffusion method. Genotypes of the 14 OS-MRSA isolates were diverse, and no dominant clones were identified. The prevalence of pvl gene among 14 OS-MRSA isolates was high up to 64.29% (9/14). All of the isolates showed heterogeneous resistance to oxacillin, while frequencies of the oxacillin-resistant subpopulations ranged from 10− 9 to 10− 5 and differed significantly among different isolates.
Conclusion: The overall prevalence of OS-MRSA was relatively lower, but lower oxacillin MICs, low testing sensitivity of routine antibiotics susceptibility testing methods and weak PBP2a protein expression were observed in this study. 14 OS-MRSA showed diverse genotypes and universal heterogeneous resistance, and inaccurate laboratory identification and improper antimicrobial usage may promote the induction of highly resistant subpopulations and lead to treatment failure.
Keywords: oxacillin sensitive MRSA, molecular typing, heterogeneous resistance
Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) is an important bacterial pathogen, leading to multiple kinds of invasive infections and increasing mortality of inpatients.1 The primary resistance mechanism of MRSA is mediated by the expression of a modified penicillin-binding protein 2a (PBP2a), which was encoded by the mecA gene.2,3 Recently, a new issue, rarely reported previously, was the misidentification of MRSA in clinical laboratories, which was mainly owing to oxacillin susceptible mecA positive Staphylococcus aureus (OS-MRSA) emerged in several areas around the globe.4–6
It is well known that the detection of resistance phenotypes with antibiotics susceptibility testing methods was routinely performed in most clinical laboratories. By contrast, widespread usage of genotypic methods was problematic. Since OS-MRSA isolates often showed lower oxacillin MICs (typically≤2 μg/mL), these isolates were often mistakenly identified as methicillin-susceptible Staphylococcus aureus (MSSA) with phenotypic methods, and even the isolates may become highly resistant ones when exposed to β-lactams antibiotics or mupirocin.7,8 Therefore, to discriminate OS-MRSA isolates, other alternative methods are required to minimize the error caused by traditional phenotypic techniques. The mecA gene detection or PBP2a protein agglutination assay was the most frequently suggested methods to complement conventional phenotypic methods. However, the discrepancies between the mecA gene and PBP2a protein expressions were observed, and a novel mecA homologue (mecC) was not detectable by PBP2a agglutination tests.9,10 Therefore, a combination of phenotypic and genotypic methods simultaneously is strongly suggested for the accurate identification of OS-MRSA.5
Moreover, heterogeneous resistance was reported to be a universal phenomenon among OS-MRSA isolates collected in Africa.8 It may pose a more knotty issue for clinicians when β-lactam antibiotics were prescribed to treat infections caused by OS-MRSA or even lead to treatment failure.11 However, characterization of heterogeneous resistance was not reported in other areas as far as I know. The actual existence of heterogeneous resistance among isolates from different groups of patients needs to be further investigated.
This study aims to investigate the phenotypic and genotypic characteristics of OS-MRSA strains isolated from patients admitted to two tertiary teaching hospitals in China. The heterogeneous resistance and its potential clinical significance were further explored.
Materials and Methods
Ethics Approval
This study is exempt from formal ethical approval and informed consent according to the local ethical guidelines, and the statement of exempt from ethical approval was provided by Ethics committee of Affiliated hospital of Inner Mongolian medical university (Reference number KY2020029), since no samples were specifically collected for this research, and the data analysis was analyzed anonymously.
Collection and Identification of Isolates
One thousand two hundred clinical isolates of S. aureus were collected at Affiliated hospital of Inner Mongolian Medical University and Bayannur people’s hospital from January 2011 to March 2019. All of the isolates were isolated from routine clinical samples and the isolates were frozen at −80°C. The isolates were retrospectively identified as MSSA with AST-GP67 card on VITEK-2 compact automation equipment (BioMérieux, France). The isolates were sub-cultured on blood agar plates during this study, and all of them were re-validated by MALDI-TOF MS equipment (micro-typer MS, Tianrui, China) as well as 16S rRNA gene sequencing.
mecA, mecC, and pvl Gene Detection
Generally, bacterial precipitate for each isolate was digested with lysostaphin (1 mg/mL) (Merck, USA) at 37°C for 30 min, and then the bacterial chromosomal DNA was extracted based on the TIANamp Bacterial DNA kit (Tiangen Biotech, China). 16S rRNA gene, mecA gene, mecC gene, and pvl gene for all 1200 isolates were amplified, and PCR conditions were set according to the procedure described previously.12–16 The primers used in this study are presented in Table 1. S. aureus ATCC25923 and ATCC43300 reference strains were used as control.
 |
Table 1 Primers for PCR Amplification in This Study |
Antibiotics Susceptibility Testing
Phenotypic methods were used to detect their resistance levels to oxacillin or cefoxitin in the isolates positive with the mecA or mecC gene expression. The methods comprise of AST-GP67 card on VITEK-2 fully automated equipment (BioMérieux, France), BD PhoenixTM PMIC/ID panel on BD Phonenix-100 equipment (BD, Sparks, MD, USA), cefoxitin disc diffusion method as well as oxacillin microdilution broth method based on the procedure recommended by CLSI M100 (29th edition).17
Molecular Typing
Pulsed-Field Gel Electrophoresis (PFGE)
The bacterial density of the tested isolates was adjusted to 3.5~4.0 Mcf and was digested with lysostaphin (1 mg/mL) (Merck, USA) at 37°C for 30 min. Bacterial chromosomal DNA of S. aureus isolates was prepared from all the isolates and cleaved with 40U SmaI (Takara, China). Meanwhile, the standard strain of S. Braenderup H9812 was prepared using 40U XbaI (Takara, China) and was used as the standard molecular mass. Electrophoresis was performed on 1% agarose gels (Lonza, Rockland, ME, USA) in 0.5 M Tris/borate/EDTA buffer on a CHEF-Mapper XA PFGE system (Bio-Rad, Hercules, CA, USA). The gel was stained with ethidium bromide (0.5 μg/mL) and photographed under the GelDoc XR system (Bio-Rad, Hercules, CA, USA). PFGE profiles were analyzed by Bionumerics v7.6 software.
Multi-Locus Sequence Typing (MLST) and Spa Typing
The experiment was performed as follows. Generally, the bacterial chromosomal DNA was extracted based on the instruction of the TIANamp Bacterial DNA kit (Tiangen Biotech, China). PCRs were carried out with 30-μL reaction volumes containing 0.3 μL of chromosomal DNA (approximately 0.3 μg), 1 μg of each primer as well as Taq DNA polymerase mixture supplied by Taq PCR MasterMix kit (Tiangen Biotech, China). The PCR reaction conditions were set as described by Enright et al.18 Seven respective PCR assays were conducted to amplify seven housekeeping genes for S.aureus, including arc, aroE, glpF, gmk, pta, tpi and yqil. The PCR products were sequenced with the same primers as those used in PCR amplification and Sanger sequencing was performed on ABI 3730XL DNA Analyzer (Applied Biosystems, USA). The sequences were compared with the known alleles in the MLST database (http://saureus.mlst.net). The variable repeat region of the spa was amplified with the following primers, Spa-1113f (TAAAGACGATCCTTCGGTGAGC) and Spa-1514r (CAGCAGTAGTGCCGTTTGCT),16 and the PCR products were sequenced on ABI 3730XL DNA Analyzer (Applied Biosystems, USA), and the sequences were analyzed using the Random web server (http://spaserver.ridom.de). The typing of chromosomal cassette mec (SCCmec) for S. aureus isolates was carried out based on the well-known protocol,19 and the unique types were designated. The standard SCCmec typing strains (NCTC10042, type I SCCmec; N315, type II SCCmec; 85/2080, type III SCCmec; JCSC4744, type IV SCCmec; HS663, type V SCCmec) were used as controls.
Population Analysis Profiling (PAP)
According to the procedure introduced previously,20 the PAP assay was performed as follows. The isolates were cultured with shaking in Tryptic Soy Broth (TSB) at 35°C, and overnight cultures were centrifuged and diluted to a suitable density. The diluted cultures were plated onto Tryptic Soy Agar (TSA) plates at a density of 109 CFU/plate. The TSA plates were prepared with serial concentrations of oxacillin (0.06–800 μg/mL) with or without 0.03 μg/mL mupirocin. The plates were then incubated at 35°C for 48 hours at aerobic condition, and the number of colonies on each plate was counted, and the frequency of highly resistant subpopulations was calculated.
PBP2a Protein Detection
The PBP2a protein expression for 14 OS-MRSA isolates and their corresponding high-level resistant subpopulations were detected with the following procedure. Generally, approximately 109 CFU of the tested isolates were collected in an eppendorf tube and centrifuged at 10,000 rpm for 10 min, and the bacterial precipitate was collected. The precipitate was then incubated for 5 min at 100°C and the mixture of bacterial protein was extracted. An agglutination test was then carried out based on the instructions of the PBP2a agglutination test kit (Oxoid Limited, US). S. aureus ATCC 25923 and ATCC43300 reference strains were used as negative and positive controls, respectively.
Results
Among 1200 S. aureus isolates tested in this study, 14 OS-MRSA isolates (1.17%) were identified, and all of them were the mecA gene positive and negative with the mecC gene. Most of the OS-MRSA isolates (64.29%, n=9) were isolated from wound samples, followed by pus samples (14.29%, n=2), whole blood sample (7.14%, n=1), BALF sample (7.14%, n=1) and sputum sample (7.14%, n=1). All of the isolates were isolated from inpatients, and the average age of the patients were 36.7 years old (8~57 years). Two isolates (OS-200 and OS-1068) of OS-MRSA were isolated from pediatric patients (14.29%, 2/14).
The resistance rates of the 14 OS-MRSA isolates to penicillin, erythromycin, tetracycline, clindamycin, gentamicin, levofloxacin and trimethoprim-sulfamethoxazole (SXT) were 100%, 78.57%, 64.29%, 35.71%, 21.43%, 7.14%, and 7.14%, respectively. All 14 OS-MRSA isolates were sensitive to nitrofurantoin, vancomycin, linezolid, rifampicin, and daptomycin, as shown in Table 2.
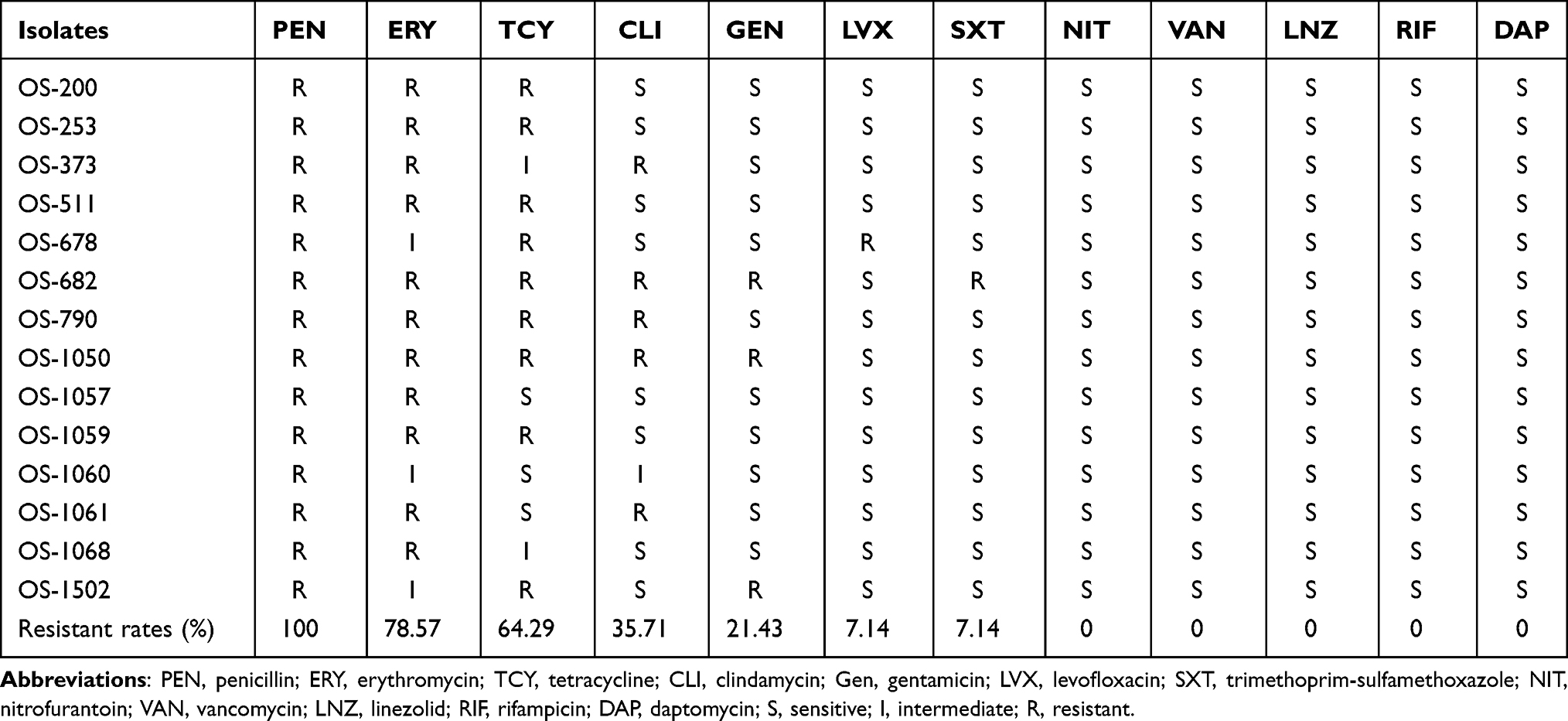 |
Table 2 Antimicrobial Susceptibility Features of 14 OS-MRSA Isolates |
Furthermore, different phenotypic methods were assessed in differentiating OS-MRSA isolates. Oxacillin MICs detected by VITEK-2 oxacillin and BD phoenix-100 showed good agreement with those detected by the oxacillin broth microdilution method. Unfortunately, all of the 14-OS MRSA isolates were mistakenly identified to be MSSA by VITEK-2, BD phoenix-100, and oxacillin broth microdilution methods, as shown in Table 3. Three out of 14 OS-MRSA isolates were determined to be MRSA by cefoxitin disk diffusion method, the oxacillin MICs of which were 1 μg/mL (OS-1068 and OS-1502) and 0.5 μg/mL (OS-200), respectively. Moreover, lower expression of PBP2a protein was observed in these 14 OS-MRSA isolates and only 42.86% (6/14) showed a positive reaction with agglutination assay (Table 3). The correlation between levels of oxacillin MIC and expression level of PBP2a protein was not observed.
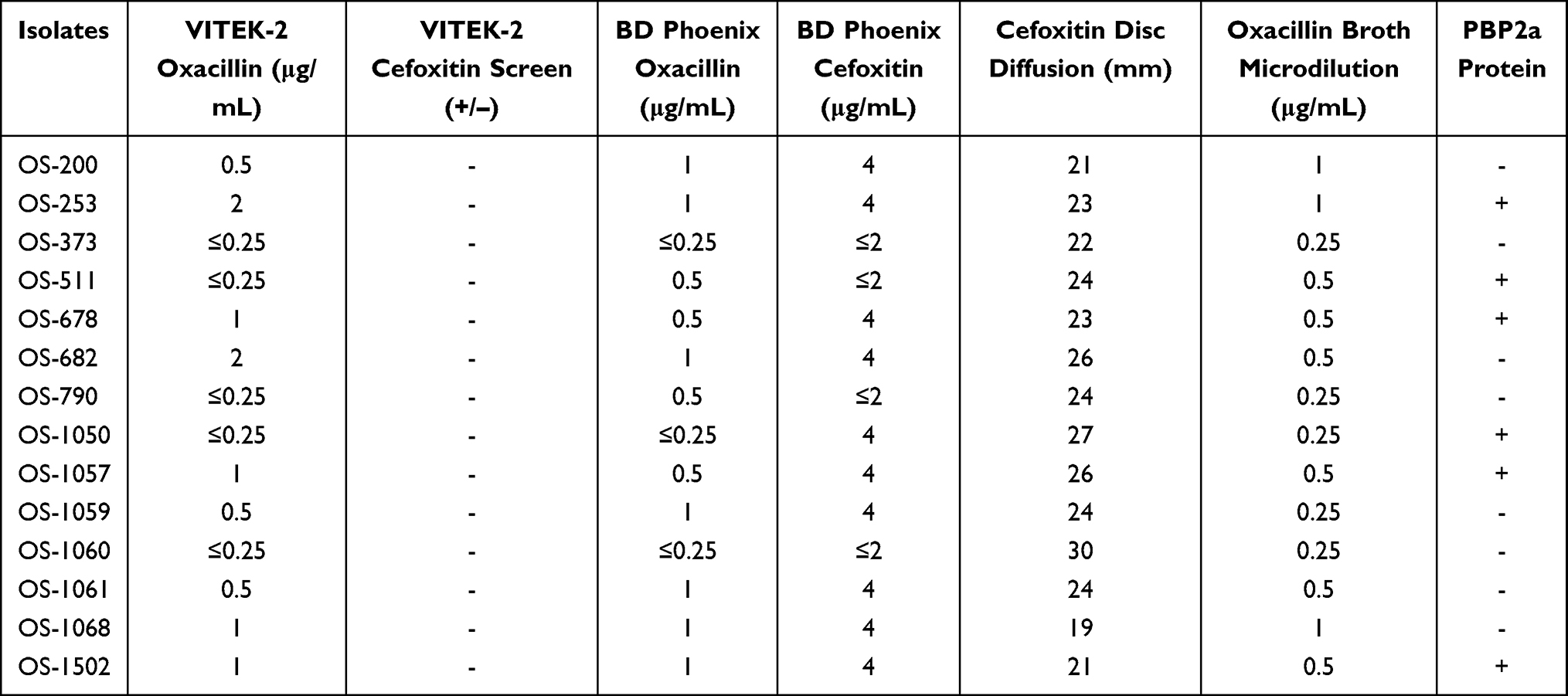 |
Table 3 Phenotypic Resistance Features of 14 OS-MRSA Isolates with Different Methods |
MLST and spa types of the 14 OS-MRSA isolates showed a diverse tendency, only ST22-t309 types accounted for 14.29% (2/14) and both of them were not typeable with the SCCmec typing method. Besides, ST398, ST59, ST25 accounted for 42.86% (6/14) and 14.29% (2/14) for each ST type, respectively. Except for OS-200, OS-790, and OS-373 isolates, ten out of 14 isolates belonged to the SCCmec IV type, while only one isolate (OS-678) was categorized as SCCmec I type (Figure 1). Furthermore, the prevalence of the pvl gene among 14 OS-MRSA isolates was high up to 64.29% (9/14), while that of 1200 S. aureus isolates was 25.5% (306/1200).
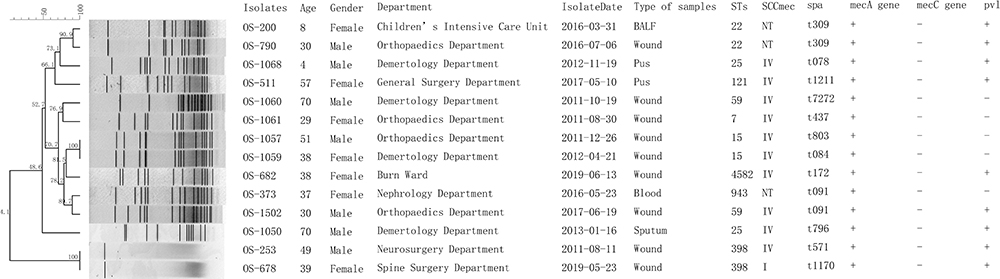 |
Figure 1 Molecular typing results of 14 OS-MRSA isolates. Abbreviations: BALF, bronchoalveolar lavage fluid; NT, not typeable. |
All OS-MRSA isolates presented with heterogeneous resistance, while frequencies of highly resistant subpopulations varied from 10−9 to 10−5 among different isolates. For each isolate, the MICs of the highly resistant subpopulations to oxacillin increased at least 16 folds compared with that of the primary isolate, and those of the four isolates (OS-682, OS-1059, OS-1060, OS-1502) increased up to 1024 folds. The effects in the induction of resistance by 0.03 μg/mL mupirocin were observed in 71.43% (10/14) isolates, while one out of them showed a significant inhibition effect (OS-1502). With PAP assay, the frequencies and resistance levels of the subpopulations with the highest resistant levels to oxacillin for each isolate differed significantly, and the oxacillin MICs of the most highly resistant subpopulations originated from 57.14% (8/14) isolates exceeded 100 μg/mL (Figure 2). It is noteworthy that all of the highly resistant subpopulations were positive with PBP2a protein expression, as shown in Table 4.
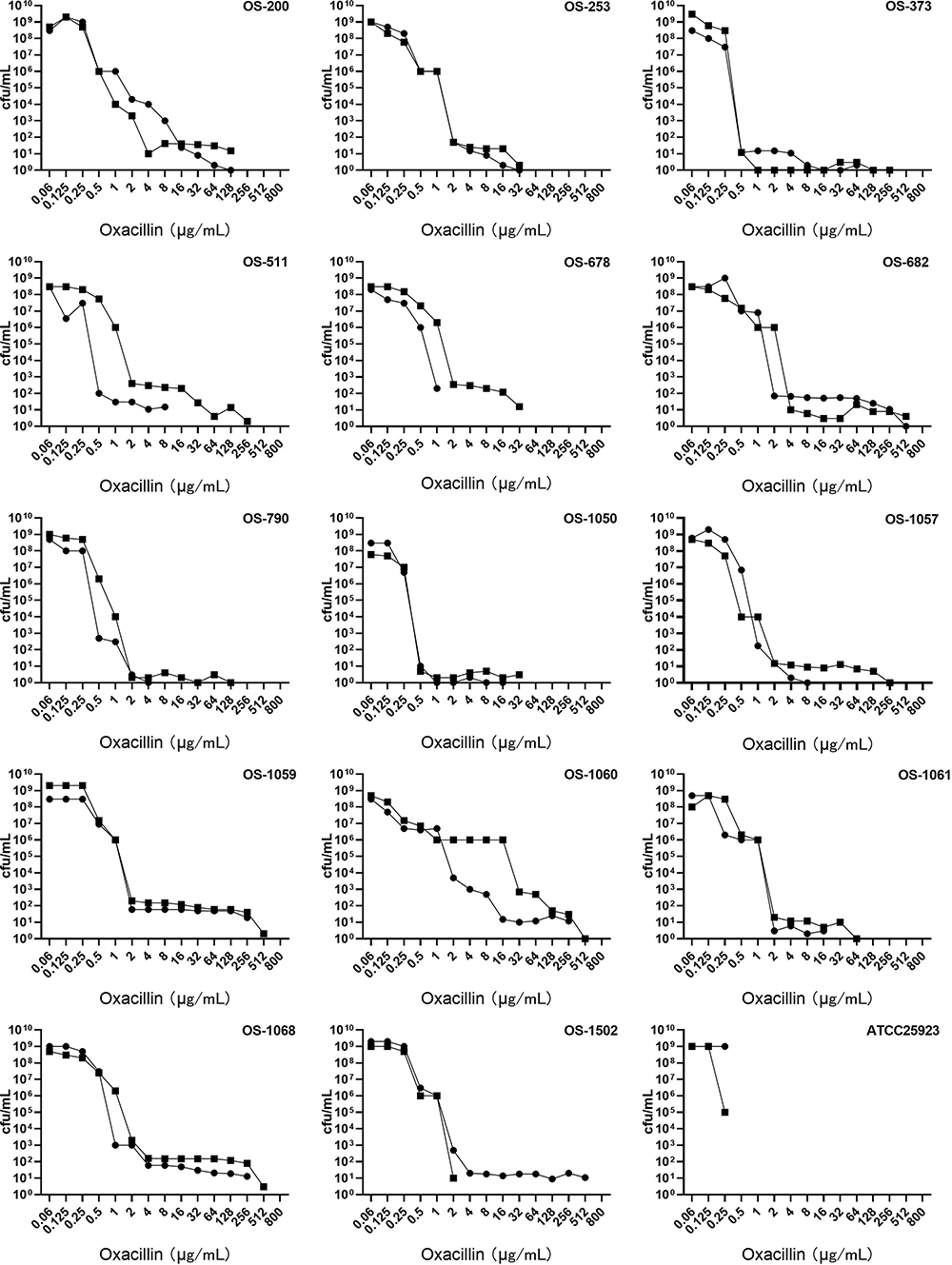 |
Figure 2 Population analysis profiles of 14 OS-MRSA, 1 MRSA isolate and 1 MSSA isolate. ●, oxacillin; ■, oxacillin+mupirocin. |
 |
Table 4 Frequency of Heterogeneous Resistance and Expression of PBP2a Protein of 14 OS-MRSA Isolates |
Discussion
As a particular subset of MRSA, the clonal distribution and prevalence rates of OS-MRSA varied greatly from 1.25% to 52% among different reports.6,7,21 The prevalence rate found in this study is 1.17% in this study, which is similar to those reported in two previous studies in China.7,22 The number of S. aureus isolates investigated in different studies may be a primary reason for significant variation of prevalence rates among different reports. Moreover, the 14 OS-MRSA isolates were all sensitive to both oxacillin and cefoxitin, no oxacillin susceptible but cefoxitin-resistant isolate was enrolled in this study. More multi-center investigations need to be further carried out to get knowledge of the actual prevalence of OS-MRSA. In consistent with some previous studies,7,8 most of the OS-MRSA isolates investigated in this study belonged to SCCmec IV type, which was closely correlated with prevalent CA-MRSA strains, including ST59, ST15 and ST25 genotypes.7,23 In contrast to a recent report from China, no dominant clone was found among 14 OS-MRSA isolates in this study and only ST22-t309 clone accounted for 14.29% (2/14). At the same time, ST22 was also reported to be a predominant clone among community-acquired infections.24 Notably, the prevalence of pvl gene among 14 OS-MRSA isolates was high up to 64.29% (9/14) and significantly higher than those reported in two studies from China,7,22 which was deemed to be closely correlated with enhanced virulence potential of CA-MRSA.25 Also, the majority of the isolates were isolated from the wound, pus, and blood samples, and only one isolate from sputum. The virulence potential of OS-MRSA should be paid more attention to.
In this study, a significant feature of OS-MRSA is its lower level of oxacillin resistance, when oxacillin MICs of the isolates are between 0.25μg/mL to 1μg/mL. Consistent with the result reported by Liu et al,7 cefoxitin disc diffusion method showed better sensitivity in the identification of OS-MRSA in this study and 21.43% (3/14) were classified into MRSA, while both VITEK-2 and BD phoenix-100TM misidentified all of the 14 OS-MRSA isolates as MSSA. Moreover, another significant feature of the 14 OS-MRSA isolates was the low-level expression of PBP2a protein, and only six isolates were positive with PBP2a protein using agglutination assay. In contrast, all of the tested OS-MRSA isolates were positive with PBP2a protein in two previous studies.7,8 On the contrary, all of the 14 OS-MRSA isolates were positive with the mecA gene. Based on the results of this study and some previous investigations, to accurately identify OS-MRSA and better treat its infections, the following suggestions were recommended. Firstly, for the patients with severe S.aureus infection and receiving β-lactam therapy for assumed MSSA with the phenotypic method, the mecA and mecC gene detection should be taken into account when the outcome of treatment is not ideal or even worsen. It is strongly recommended to communicate with laboratories about this issue, especially for the cases with oxacillin MICs ≥0.5 μg/mL, since oxacillin MICs of most of the OS-MRSA isolates ranged from 0.5 to 2.0 μg/mL based on the results of this study as well as some previous studies.7,8,22 Secondly, the total resistance rates of 14 OS-MRSA isolates to several commonly tested antimicrobials were lower than 50% except for penicillin, erythromycin, and tetracycline, which was consistent with a recent report by Liu et al.7 The clinician should consider non-β-lactam antimicrobials for severe S. aureus infection when the phenotypic method cannot be performed promptly. Finally, cefoxitin disk method is recommended to be carried out routinely to provide more accurate antibiotic susceptibility results for severe S. aureus infection, since this phenotypic method was shown to be with more detection sensitivity in discrimination of OS-MRSA.7 Alternatively, the laboratories should communicate on this issue with clinicians and seek help from other laboratories to perform genetic detection.
Inspired by a previous study,8 we further investigated the heterogeneous resistance feature of the 14 OS-MRSA isolates. We found it is universal among OS-MRSA isolates identified in this study. The frequency of oxacillin-resistant subpopulations among different isolates varied considerably from 10−9 to 10−5 and lower than those reported by Chung et al.8 However, highly resistant subpopulations with oxacillin MICs exceeding 100 μg/mL existed in 57.14% (8/14) OS-MRSA isolates by the PAP method. It is revealed that sub-inhibitory concentrations of β-lactam antibiotics could promote biofilm formation of OS-MRSA26 and exposure to 0.5 MIC of oxacillin, facilitating the conversion of OS-MRSA into highly resistant ones.7 When OS-MRSA was misinterpreted as MSSA using routine phenotypic methods and the patients were treated with β-lactam antibiotics, the highly resistant subpopulations are most likely to be selected and may lead to failure of treatment. How to rapidly and accurately identify OS-MRSA isolates with heterogeneous resistance feature seems to be more critical. Some novel techniques on a single-cell level may provide more choices to rapidly detect heteroresistance, such as droplet digital PCR27 and plasmonic colloidosomes coupled MALDI-TOF MS technique.28 Moreover, heterogeneously resistant clones could convert into homogeneously and highly resistant ones when exposed to mupirocin, which is well recognized as an effective inducer of the stringent stress response.29 In this study, 0.03μg/mL mupirocin could significantly increase resistance levels of 10 OS-MRSA isolates to oxacillin at least two folds. In contrast, that of one isolate decreased (OS-1502). A similar phenomenon was reported by Chung et al8 and the mechanism of diverse responses of OS-MRSA to mupirocin needs to be further investigated.
Although this investigation revealed some new features of OS-MRSA, there are still some limitations in this study. Only 14 OS-MRSA isolates were detected from two hospitals, and a larger number of isolates should be collected from more hospitals. More detailed information about OS-MRSA may be more apparent to us. Owing to the limitation of expenditure, only the molecular typing features of 14 OS-MRSA isolates were investigated in this study, while those of the remaining 1186 non-OS-MRSA isolates were not further explored, which deserves to be further investigated. Furthermore, some molecular mechanisms were reported to be correlated with the occurrence and heteroresistance of OS-MRSA, such as point mutations within the mecA promoter,7 et al It was not investigated in this study, which is still under investigation in our laboratory.
In conclusion, 14 OS-MRSA isolates showed a significant clonal and resistance diversity. All of them were wrongly identified as MSSA with several phenotypic resistance detection methods that were widely applied in many clinical laboratories. The universal heterogeneous resistance of OS-MRSA observed in this study poses a more knotty issue for accurate identification and treatment of infections caused by OS-MRSA.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No.81260244, 81660352), Natural Science Foundation of Inner Mongolia Autonomous Region of China (Grant no. 2020MS08110), Affiliated hospital of Inner Mongolia Medical University (No, NYFY ZD 012), Graduate initiative program of Inner Mongolian Medical University (No, FYQMJH2020006) and Program for Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region (No. NJYT-17-B06).
Roushan Liu and Jian Zhang contributed equally to this manuscript.
Disclosure
The authors declare that they have no conflicts of interest for this work.
References
1. Woodford N, Livermore DM. Infections caused by Gram-positive bacteria: a review of the global challenge. J Infect. 2009;59(Suppl 1):S4–16. doi:10.1016/S0163-4453(09)60003-7
2. Schito GC. The importance of the development of antibiotic resistance in Staphylococcus aureus. Clin Microbiol Infect. 2006;12(Suppl 1):3–8. doi:10.1111/j.1469-0691.2006.01343.x
3. Peacock SJ, Paterson GK. Mechanisms of methicillin resistance in Staphylococcus aureus. Annu Rev Biochem. 2015;84(1):577–601. doi:10.1146/annurev-biochem-060614-034516
4. Bignardi GE, Woodford N, Chapman A, Johnson AP, Speller DC. Detection of the mec-A gene and phenotypic detection of resistance in Staphylococcus aureus isolates with borderline or low-level methicillin resistance. J Antimicrob Chemother. 1996;37(1):53–63. doi:10.1093/jac/37.1.53
5. Sharff KA, Monecke S, Slaughter S, et al. Genotypic resistance testing creates new treatment challenges: two cases of oxacillin-susceptible methicillin-resistant Staphylococcus aureus. J Clin Microbiol. 2012;50(12):4151–4153. doi:10.1128/JCM.01564-12
6. Hososaka Y, Hanaki H, Endo H, et al. Characterization of oxacillin-susceptible mecA positive Staphylococcus aureus: a new type of MRSA. J Infect Chemother. 2007;13(2):79–86. doi:10.1007/s10156-006-0502-7
7. Liu JL, Li TM, Zhong N, et al. Current status of oxacillin-susceptible mecA positive Staphylococcus aureus infection in Shanghai, China: a multi-center study. J Microbiol Immunol Infect. 2020. doi:10.1016/j.jmii.2020.07.021
8. Chung M, Kim CK, Conceição T, et al. Heterogeneous oxacillin-resistant phenotypes and production of PBP2A by oxacillin-susceptible/mecA-positive MRSAA strains from Africa. J Antimicrob Chemother. 2016;71(10):2804–2809. doi:10.1093/jac/dkw209
9. Garci’a-A’lvarez L, Holden MT, Lindsay H, et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: a descriptive study. Lancet Infect Dis. 2011;11(8):595–603. doi:10.1016/S1473-3099(11)70126-8
10. Paterson GK, Morgan FJE, Harrison EM, et al. Prevalence and characterization of human mecC methicillin-resistant Staphylococcus aureus isolates in England. J Antimicrob Chemother. 2014;69(4):907–910. doi:10.1093/jac/dkt462
11. Duarte FC, Danelli T, Tavares ER, et al. Fatal sepsis caused by mecA-positive oxacillin-susceptible Staphylococcus aureus: first report in a tertiary hospital of southern Brazil. J Infect Chemother. 2019;25(4):293–297. doi:10.1016/j.jiac.2018.09.010
12. Monday SR, Bohach GA. Use of multiplex PCR to detect classical and newly described pyrogenic toxin genes in Staphylococcal isolates. J Clin Microbiol. 1999;37(10):3411–3414. doi:10.1128/JCM.37.10.3411-3414.1999
13. Cikman A, Aydin M, Gulhan B, et al. Absence of the mecC gene in methicillin-resistant Staphylococcus aureus isolated from various clinical samples: the first multi-centered study in Turkey. J Infect Public Health. 2019;12(4):528–533. doi:10.1016/j.jiph.2019.01.063
14. Stegger M, Andersen PS, Kearns A, et al. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecALGA251. Clin Microbiol Infect. 2012;18(4):395–400. doi:10.1111/j.1469-0691.2011.03715.x
15. McClure JA, Conly JM, Lau V, et al. Novel multiplex PCR assay for detection of the staphylococcal virulence marker panton-valentine leukocidin genes and simultaneous discrimination of methicillin-susceptible from -resistant staphylococci. J Clin Microbiol. 2006;44(3):1141–1144. doi:10.1128/JCM.44.3.1141-1144.2006
16. Mellmann A, Friedrich AW, Rosenkötter N, et al. Automated DNA sequence-based early warning system for the detection of methicillin-resistant staphylococcus aureus outbreaks. Plos Med. 2006;3(3):e33. doi:10.1371/journal.pmed.0030033
17. Wayne P. Performance Standards for Antimicrobial Susceptibility Testing.
18. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38(3):1008–1015. doi:10.1128/JCM.38.3.1008-1015.2000
19. Zhang K, McClure JA, Elsayed S, Louie T, Conly JM. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J Clin Microbiol. 2005;43(10):5026–5033. doi:10.1128/JCM.43.10.5026-5033.2005
20. Tomasz A, Nachman S, Leaf H. Stable classes of phenotypic expression in methicillin-resistant clinical isolates of staphylococci. Antimicrob Agents Chemother. 1991;35(1):124–129. doi:10.1128/aac.35.1.124
21. Andrade-Figueiredo M, Leal-Balbino TC. Clonal diversity and epidemiological characteristics of Staphylococcus aureus: high prevalence of oxacillin-susceptible mecA-positive Staphylococcus aureus (OS-MRSA) associated with clinical isolates in Brazil. BMC Microbiol. 2016;16(1):115. doi:10.1186/s12866-016-0733-4
22. Song YJ, Cui LQ, Lv Y, Li Y, Xue F. Characterisation of clinical isolates of oxacillin-susceptible mecA positive Staphylococcus aureus in China from 2009 to 2014. J Glob Antimicrob Resist. 2017;11:1–3. doi:10.1016/j.jgar.2017.05.009
23. Wang X, Shen YY, Huang WC, Zhou Y. Characterisation of community-acquired Staphylococcus aureus causing skin and soft tissue infections in a children’s hospital in Shanghai, China. Epidemiol Infect. 2019;147:e323. doi:10.1017/S0950268819002127
24. Xiao N, Yang JH, Ning Duan N, Lu BH, Wang LJ. Community-associated Staphylococcus aureus PVL+ ST22 predominates in skin and soft tissue infections in Beijing, China. Infect Drug Resist. 2019;12:2495–2503. doi:10.2147/IDR.S212358
25. Trista A, Bes M, Meugnier H, et al. Global distribution of panton-valentine leukocidin–positive methicillin-resistant Staphylococcus aureus, 2006. Emerg Infect Dis. 2007;13:594e600. doi:10.3201/eid1304.061316
26. He X, Li S, Yin Y, et al. Membrane vesicles are the dominant structural components of ceftazidime-induced biofilm formation in an oxacillin-sensitive MRSA. Front Microbiol. 2019;10:571. doi:10.3389/fmicb.2019.00571
27. Sun L, Talarico S, Yao L, et al. Droplet digital PCR detection of Helicobacter pylori clarithromycin resistance reveals frequent heteroresistance. J Clin Microbiol. 2018;56(9):e00019–18. doi:10.1128/JCM.00019-18
28. Dai Y, Li C, Yi J, Qin Q, Liu B, Qiao L. Plasmonic colloidosomes coupled with MALDI-TOF MS technique. Plasmonic colloidosome-coupled MALDI-TOF MS for bacterial heteroresistance study at single-cell level. Anal Chem. 2020;92(12):8051–8057. doi:10.1021/acs.analchem.0c00494
29. Kim C, Mwangi M, Chung M, Milheiriço C, de Lencastre H, Tomasz A. The mechanism of heterogeneous beta-lactam resistance in MRSA: key role of the stringent stress response. PLoS One. 2013;8(12):e82814. doi:10.1371/journal.pone.0082814
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.