")
Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 16
Clinical Associations of Bitter Taste Perception and Bitter Taste Receptor Variants and the Potential for Personalized Healthcare
Authors Mao Z, Cheng W , Li Z, Yao M, Sun K
Received 17 September 2022
Accepted for publication 7 February 2023
Published 12 February 2023 Volume 2023:16 Pages 121—132
DOI https://doi.org/10.2147/PGPM.S390201
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Ziwen Mao,1,2 Weyland Cheng,1,2 Zhenwei Li,2 Manye Yao,2 Keming Sun2
1Henan Provincial Key Laboratory of Children’s Genetics and Metabolic Diseases, Children’s Hospital Affiliated to Zhengzhou University, Henan Children’s Hospital, Zhengzhou Children’s Hospital, Zhengzhou, Henan, People’s Republic of China; 2Department of Orthopaedic Surgery, Children’s Hospital Affiliated to Zhengzhou University, Henan Children’s Hospital, Zhengzhou Children’s Hospital, Zhengzhou, Henan, People’s Republic of China
Correspondence: Weyland Cheng, Henan Provincial Key Laboratory of Children’s Genetics and Metabolic Diseases, Children’s Hospital Affiliated to Zhengzhou University, Henan Children’s Hospital, Zhengzhou Children’s Hospital, 33 Longhu Waihuan East Road, Zhengzhou, Henan, People’s Republic of China, Tel +86 18502758200, Email [email protected]
Abstract: Bitter taste receptors (T2Rs) consist of 25 functional receptors that can be found in various types of cells throughout the human body with responses ranging from detecting bitter taste to suppressing pathogen-induced inflammation upon activation. Numerous studies have observed clinical associations with genetic or phenotypic variants in bitter taste receptors, most notably that of the receptor isoform T2R38. With genetic variants playing a role in the response of the body to bacterial quorum-sensing molecules, bacterial metabolites, medicinal agonists and nutrients, we examine how T2R polymorphisms, expression levels and bitter taste perception can lead to varying clinical associations. From these genetic and phenotypic differences, healthcare management can potentially be individualized through appropriately administering drugs with bitter masking to increase compliance; optimizing nutritional strategies and diets; avoiding the use of T2R agonists if this pathway is already activated from bacterial infections; adjusting drug regimens based on differing prognoses; or adjusting drug regimens based on T2R expression levels in the target cell type and bodily region.
Keywords: bitter taste receptors, TAS2R38, chronic rhinosinusitis, obesity, oral health, cancer
Introduction
Bitter taste receptors (T2Rs) are G-protein coupled receptors (GPCRs) that are classified as Frizzled/Taste2 family based on the GRAFS classification system.1 The seven-transmembrane receptor is encoded by a gene family consisting of 25 functional genes, which leads to the expression of 25 functional receptor isoforms in the human body where they can be activated by various ligands. Activation of these receptors leads to the dissociation of the GPCR’s heterotrimeric G-proteins. The Gα subunit upregulates phosphodiesterase, causing the hydrolyzation of cAMP, whereas the Gβγ subunit initiates the phospholipase Cβ2/inositol-1,4,5-triphosphate signaling pathway, releasing stored calcium from the endoplasmic reticulum into the cytoplasm.2,3 In the gustatory system, bitter taste receptors are located in specialized epithelial taste receptor cells that reside in the taste bud. In the transduction pathway for bitter taste, the increase in intracellular calcium is followed by the release of ATP through the activation of calcium homeostasis modulator channels, thereby depolarizing the taste receptor cell. The released ATP subsequently activates ionotropic purinergic receptors on the sensory ganglion neurons that innervate the taste buds, thereby relaying bitter taste perception to the brain.4
In recent years, there has been interest in the role of extraoral T2Rs in various diseases such as asthma, chronic rhinosinusitis, COVID-19, systemic inflammation, cancer, eczema and osteomyelitis.5–10 Most studies have focused on extraoral T2Rs in the respiratory system where T2R activation can increase ciliary beat frequency, reduce inflammatory markers and emit anti-pathogenic substances, such as antimicrobial peptides and low levels of nitric oxide via endothelial nitric oxide synthase.11,12 In immune cells such as lymphocytes, monocytes, granulocytes, leukocytes and mast cells, T2R activation can also decrease or inhibit pro-inflammatory cytokines such as TNF-alpha, IL-1β, IL-2, IL-4, IL-5, G-CSF, GM-CSF, MCP-1, histamine, PGD2, etc.13,14 Other studies in literature have also found functional effects of T2Rs in the gastrointestinal, genitourinary, skin, cardiovascular and neurovascular system.6,15–19
Here, we examine the phenotypic and genetic variations of bitter taste perception and T2Rs, mainly focusing on the more well-researched variation of T2R38. We discuss their clinical associations and significance and offer our perspectives on applying this knowledge in health management and towards future studies on personalizing medical treatments.
Genetic Variants
T2R genes (TAS2R) have been noted to have genetic polymorphisms that affect its sensitivity and function upon activation. Some relevant isoforms and their single nucleotide polymorphisms (SNPs) are listed in Table 1, which include TAS2R3, -4, -5, -14, -16, -19, -38, and -46. Current studies in literature have predominantly focused on the TAS2R38 polymorphisms. Three SNPs consisting of rs713598, rs1726866 and rs10246939 reside in the TAS2R38 gene on chromosome 7, which forms two predominantly common haplotypes. These two haplotypes are denoted by the affected amino acids at positions 49, 262 and 296: alanine-valine-isoleucine (AVI) and proline-alanine-valine (PAV).8,20 Different phenotypic expressions have been noted based on these two haplotypes.
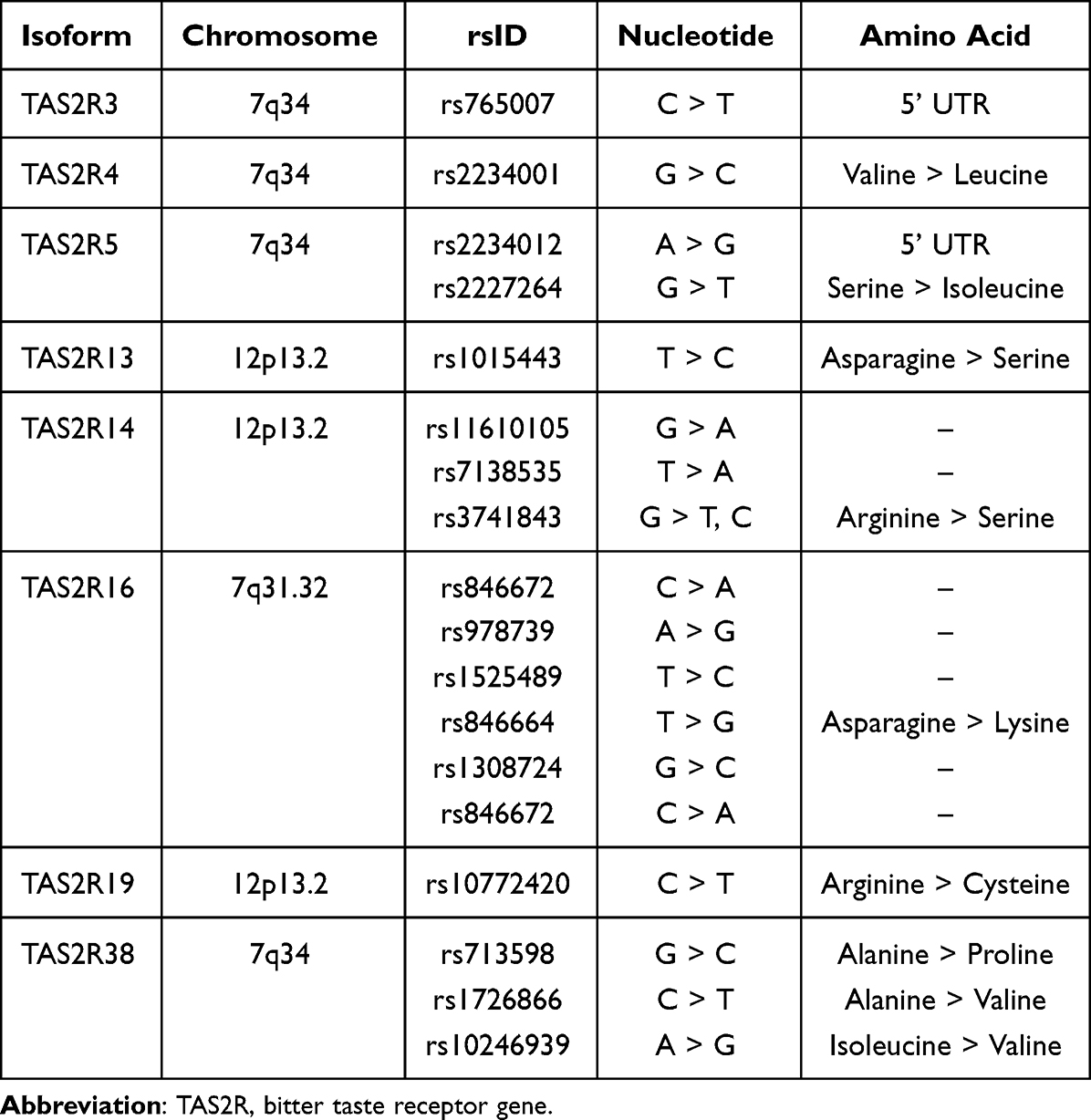 |
Table 1 Genetic Variants of Bitter Taste Receptor Isoforms That Have Been Found to Have Clinical Associations |
Two often used agonists for T2R38 activation are phenylthiocarbamide (PTC) and 6-n-propylthiouracil (PROP).21 Taste tests of these two compounds can be administered to determine phenotypic variations in bitter taste caused by TAS2R38 polymorphisms. Studies have shown that AVI/AVI diplotypes are insensitive to T2R38 activation (non-tasters), PAV/AVI diplotypes (tasters) have a moderate reaction to T2R38 activation and PAV/PAV diplotypes (supertasters) are ultrasensitive to T2R38 activation. This difference in sensitivity is also significant in bacterial infections as T2R38 can be activated by bacterial quorum-sensing molecules such as acyl-homoserine lactone and bacterial metabolites, which in turn induce antimicrobial or anti-inflammatory effects in immune cells, ciliated cells of lung epithelium and sinonasal epithelial cells.22,23
Clinical Studies
In addition to the underlying mechanisms, understanding clinical associations of bitter taste receptor polymorphisms is necessary to optimize healthcare plans and medical treatment. The clinical associations based on current literature are covered in this section and have been compiled in Table S-1.
Taste Preference and Perception
Compliance to Medication
Taste preference and perception can play a large role in adherence for liquid oral medication and dietary habits in the pediatric population.24–28 Bitterness perception based on the TAS2R38 genotype can vary depending on age where children with the AVI haplotype have higher sensitivity to bitter agonists compared to adults.29 Genotyping studies have shown that children with the more bitter-sensitive PAV/AVI and PAV/PAV diplotypes may be more inclined to take solid pills over liquid medication and are more likely to reject medication due to bitterness.27,28 Thus, masking bitter taste or adjusting the administration method for bitter-sensitive children have been regarded as viable strategies in improving compliance.30
Dietary Habits and Nutrition
In addition to bitter medications, genetic variation can also play a role in food consumption and dietary habits, which can be a factor in increasing the risk of cancer, obesity, oral diseases and life span. Several SNPs of various T2R isoforms, including TAS2R13, -16, -19, -38 and a TAS2R3/4/5 haploblock, have been related to bitter taste perception of various beverages, including caffeine and alcohol.31–37 Greater perception of PROP bitterness was found to be a significant predictor for alcohol intake and greater perception of ethanol intensity.33 SNPs in TAS2R4 and TAS2R14 have been, respectively, associated with the regulation and predictability of bitter taste sensitivity towards stevioside solution.38 Differences in food preferences and dietary habits based on TAS2R38 have also been noted in several studies where PAV/PAV supertasters were generally more averse to bitter foods.34,35,39–43 In infants, TAS2R38 genotype was associated with first complementary food intake in infants, where a higher percentage of non-tasters completed their meal on the first attempt.26 TAS2R38 polymorphisms have also been found to affect smoking habits, where menthol cigarette smokers were found to have a higher frequency of the supertaster PAV haplotype compared to non-menthol cigarette smokers.44,45 In one study, smokers were associated with the phenotype of higher bitter taste sensitivity of PROP compared to non-smokers.46
Obesity
Given that nutrition plays an important role in bodily health, researchers have observed the consequential effects of TAS2R variation. Obese patients were found to perceive bitter taste more intensely or have higher detection thresholds compared to lean or normal-weighted patients based on the PROP taste test.47,48 Likewise, body mass index (BMI) was also inversely associated PTC taste sensitivity.49 Increased sensitivity to certain bitter substances could indicate a lower quantity of vegetable consumption.40,41 Contrarily, one study found the AVI/AVI non-taster diplotype to be associated with higher risk of obesity in European Americans and Asians,50 possibly due to differences in nutrient sensing, energy metabolism, immune responses or reduced sensitivity to sweet taste recognition.43
Oral Health
Dietary factors consisting of sugary and fatty foods, in turn, contribute to incidents of dental caries and oral disease.51 As the non-taster phenotype and genotype are also associated with reduced sensitivity to sweet taste and liking sweets,43,52 numerous phenotype taste tests have also been conducted, relating the non-taster status of PROP to dental decay and dental caries.53–66 Generally, studies have found that PROP non-tasters were at higher risk of decayed, missing, and filled surfaces and caries experience compared to tasters. Medium tasters were also at higher risk compared to supertasters. This association was also found in a PTC taste sensitivity study.56 Discrepancies in dietary habits caused by phenotypic variants can also have an effect on the offspring of non-taster mothers, where the children had a higher prevalence of dental caries.61,66
The relationship between the non-taster status and tooth decay has also been confirmed in a genetic study relating TAS2R38 variation to oral health where the PAV haplotype was associated with protection from dental caries and the AVI haplotype associated with caries risk.67 Contrarily, in another study, the AVI/AVI genotype was associated with decreased prevalence of periodontal disease in Thai patients.68 Discrepancies can possibly be explained through additional factors. In obese children, AVI non-taster haplotypes were associated with increased decayed, missing and filled permanent/primary teeth scores for obese patients but had the opposite effect in a control non-obese population where non-tasters were associated with decreased scores.69 In another study, it was suggested that PROP supertasters may express wider anhedonia leading to a higher proportion of people who disliked sweets.70
Longevity
Since nutrition, obesity, oral health, and cancer also factor into aging and longevity, studies have also noticed associations with TAS2R38 and TAS2R16 polymorphisms to human life span, although there has been some heterogeneity for both of these genes.71–74 Whether or not associations with longevity are due to differences in dietary habits or if the variance in responses to extra-oral T2R activation plays a significant role in aging remains to be determined. Further data is required to examine the influence of additional factors such as demographics, nutrition, culture, environment and other genetic factors.
Parkinson’s Disease and Gut Microbiota
A higher frequency of the PROP non-taster and PTC non-taster status has been found in patients with Parkinson’s disease compared to healthy control groups.75,76 Testing for the TAS2R38 variant also showed an association between Parkinson’s disease and the non-tasting haplotype where there was a significantly higher frequency of the AVI haplotype and lower frequency of the PAV/PAV diplotype compared to the control group.75 The study also found no differences in genotype results of the gustin gene, indicating that the differences in taste status were not due to the non-functional form of gustin nor due to a low number of taste buds. A subsequent study showed that patients with Parkinson’s disease who were insensitive to PROP and had at least one AVI haplotype showed consistent differences in gut microbiota compared to other patients.77 Patients with at least one AVI haplotype had lower bacterial diversity with significantly lower genus Clostridium. Based on the author’s discussion, there are beneficial effects of microbiota from Clostridium genus in the gastrointestinal tract where the microbiota contributes towards the metabolism of short-chain fatty acids, bile acids and tryptophan. These key metabolites are known to have positive health contributions including controlling bacterial overgrowth, modulating microbial composition, inducing gut hormone secretion, stimulating gastrointestinal motility, and generating anti-inflammatory or anti-oxidative effects.78 In turn, gut microbiota composition and inflammation are associated with the pathogenesis of Parkinson’s disease.79,80
Cancer
Activation of T2Rs in various cancerous cells has been noted to produce several anti-cancerous effects, including decreasing cell proliferation, migration, invasion, motility, angiogenesis and metastasis along with increasing apoptosis and cell cycle arrest.10,81–86 Known T2R agonists and bitter flavonoids, such as quercetin, chrysin, coumarin, noscapine and naringin, have also been found to have similar beneficiary effects in vitro and in animal studies along with clinical studies using the flavonoids a supplementary treatment or adjuvant.87–91
In a systematic review, Zehentner et al noted numerous TAS2R isoforms were either downregulated or upregulated in several cancerous tissues and cell lines compared to their non-cancerous counterparts.10 It has been discovered that T2R4 is comparatively downregulated in breast cancer cells, whereas T2R14 is upregulated.81,92 Multiple T2R isoforms have also been found to be differentially expressed among ovarian cancer cells, tissue of fallopian tube origin, prostate cancer cells and benign prostatic hyperplasia.82 Variability in gene expression can have consequences in terms of the survival rate or prognosis of cancer patients. Based on the Cancer Genome Atlas, increased expression of T2Rs in head and neck squamous cell carcinomas was associated with improved overall survival.86 Zehentner et al determined that the upregulation of TAS2Rs were associated with an improved prognosis, whereas lower expression levels of certain TAS2Rs were associated with a poor prognosis.10 Consequently, T2R agonist drugs that are used to supplement cancer treatments would also likely vary in efficacy based on TAS2R expression. Studies and drug administration targeting T2Rs for cancer thus must consider testing for TAS2R expression along with possible functional variants prior to application.
Several clinical studies have also been conducted on T2R polymorphisms in relation to cancer risk. Results for cancer risk associations with the three main TAS2R38 diplotypes have been heterogeneous. The PAV/AVI diplotype was associated with increased gastric cancer risk among Korean participants despite the variant not being associated with dietary intake.93 Among Japanese gastrointestinal cancer patients, there was a higher frequency of the AVI/AVI diplotype and a lower frequency of the PAV/PAV diplotype compared to controls, although no associations were found with TAS2R46 genotypes.94 Similarly, a study on the Czech and German population found a, respectively, borderline significant and statistically significant relationship between AVI/AVI diplotypes and increased colorectal cancer risk.95 In contrast, AVI/AVI was associated with reduced risk of colorectal cancer in Korean patients despite no positive associations found between AVI/AVI diplotypes and cancer-inducing dietary habits.96 A study among Latin Americans found that the PAV/PAV diplotype showed an increase risk in gastric cancer compared to the PAV/AVI diplotype.97 In terms of other TAS2R isoforms, only one out of six SNPs of TAS2R16 (rs1525489) was found to be associated with increased risk of rectal cancer in European patients.98 No associations were found between TAS2R14 polymorphisms and colon cancer.99
Respiratory Illnesses
Due to antipathogenic effects of T2R activation in the respiratory system, several studies have found relationships between phenotypic or genetic variants of TAS2R38 and chronic rhinosinusitis (CRS). The non-taster phenotype has been associated with increased severity in Caucasian patients with CRS without polyps, and the supertaster phenotype was associated with less incidences of sinus infections and better nasal quality of life scores.100,101 Concurrently, genotype studies have shown that 1) CRS patients with the nonfunctional AVI/AVI variant were significant more likely to undergo surgical intervention due medical management being ineffective; 2) patients with PAV/PAV genotypes had significantly lower computed tomography (CT) scores compared to AVI/AVI genotypes; 3) presence of culturable bacteria was more likely in nasal swabs from patients with the AVI/AVI genotype; 4) in vivo biofilm formation was more likely in sinonasal mucosa samples of AVI/AVI patients; 5) there was a higher frequency of CRS patients with both the minor allele (A) at SNP rs10772420 for the TAS2R19 gene and the non-taster allele (A) of the TAS2R38 gene; 6) CRS patients had higher levels of TAS2R38 expression compared to controls and higher TAS2R38 expression was associated with increased severity of inflammation; and 7) TAS2R38 expression in the inferior turbinate mucosa was higher in CRS patients with more severe symptoms and in patients with both asthma and nasal polyps.102–109
These trends held true for disorders that caused similar respiratory manifestations, such as cystic fibrosis and primary ciliary dyskinesia. There was a lower frequency of the PAV allele in cystic fibrosis patients with nasal polyposis who required surgery and in cystic fibrosis patients with chronic pulmonary colonization by Pseudomonas aeruginosa.110 Similarly, in patients with primary ciliary dyskinesia, one study found that there was a lower percentage of PAV/PAV patients with frequent respiratory exacerbations and no PAV/PAV patients with chronic colonization by Pseudomonas aeruginosa.111 Concurrently, AVI/AVI primary ciliary dyskinesia patients with CRS tended to have nasal polyposis and displayed increased severity of disease.112
Aside from TAS2R38, TAS2R14 polymorphisms were associated with Korean asthmatic patients, where increased bronchodilator response and lower mean asthma control test scores were significantly associated with SNPs 815T>C, 1267A>G, 1897T>C and 815T>C, respectively.113 In line with respiratory illnesses, a couple of studies have examined the effect of the T2R38 phenotype on coronavirus disease 2019 (COVID-19) symptoms. Based on phenotypic expression, non-tasters were more likely to test positive for COVID-19, more likely to be hospitalized and more likely to display longer duration of symptoms, whereas moderate tasters only displayed mild-to-moderate symptoms.114,115 Contrarily, a more recent study by Risso et al found no relationship between the TAS2R38 SNPs and severity of COVID-19 symptoms.116
Expression Levels on Skin
The presence of T2R isoforms and the level of TAS2R mRNA expression were found to be variable in the skin, where the expression levels of some TAS2Rs were possibly related to sun exposure (TAS2R14, −30, −42, −60), sex (TAS2R3, -4, -8, -9, -14, -60) and age (TAS2R5).117 A prior study also found that TAS2R1 and TAS2R38 were expressed and functional in human epidermal keratinocytes.17 The activation of HaCaT keratinocytes using diphenidol and amarogentin induced intracellular calcium release and stimulated the expression of differentiation markers.
Osteomyelitis
Previously, we surmised that T2Rs had a role in bone inflammation.5 In our own preliminary clinical observation of 34 pediatric patients with acute osteomyelitis, the frequency of PAV/PAV, PAV/AVI and AVI/AVI patients were found to be 44.1% (15/34), 41.2% (14/34) and 14.7% (5/34), respectively. Blood parameters showed that PAV/PAV patients had significantly higher levels of peak platelet counts (P = 0.0096) and longer hospital stays (P = 0.0266) compared to PAV/AVI patients. Interestingly, the canonical T2R pathway is known upregulate PLCβ2 production and decrease cAMP levels,11 both of which can lead to increases in platelet aggregation.118
In our preliminary results, only 2 out of 29 of the PAV/PAV and PAV/AVI patients were infected with methicillin-resistant Staphylococcus aureus (MRSA), compared to 3 out of 5 AVI/AVI patients being infected with MRSA. A theoretical reasoning for the low incidences of MRSA infections in PAV/PAV and PAV/AVI patients could be due to the differences in the volatile organic compounds produced by MRSA and methicillin-sensitive Staphylococcus aureus (MSSA).119 Volatile metabolites from S aureus that can activate the T2R38 pathway include 2-pentanone, 2-methylpropanal, dimethyl disulfide and methyl mercaptan.120–122 Metabolites from MRSA may be able to activate the T2R38 pathway in PAV/PAV and PAV/AVI patients, inducing antipathogenic mechanisms in immune cells.
Clinical Application and Future Perspectives
Several considerations arise when applying knowledge of T2R SNPs and taste variation to health management, personalizing drug regimens and future studies. Masking the bitter taste of medication has been a long-known strategy for improving compliance, especially for the pediatric population. Knowledge of a patient’s variant can help improve treatment regimens, administration methods or patient compliance by choosing medications, delivery methods or brands that are more palatable to the patient. For instance, a study on the taste perception of two commonly used antibiotics, chloramphenicol and ofloxacin, was found to be associated with the TAS2R38 diplotype and a TAS2R9 SNP, respectively.123 Considering that TAS2R variants play a factor in medication compliance,27 patients susceptible to increased bitterness of these antibiotics may be administered solid oral medications over liquid medications or brands that particularly block the activation of the associated T2R isoform. Infants or mentally disabled patients who are unable to provide feedback on their reason for noncompliance can also undergo SNP testing where considerations regarding their bitter taste perception can be worked into a medication compliance protocol.
Several factors can influence the relationship between taste sensitivity, food preference and obesity, such as genetic factors, ethnicity, age, gender, estrogenic phase, endocannabinoid system, and cognitive factors.124 Consequently, studies on the relationship between bitter taste perception and obesity can potentially yield heterogeneous results. Although most studies in literature have found an association between obesity or BMI and TAS2R38 variation,42,43,47,48,50 a study on an Indian population, for instance, found that both the TAS2R38 genotype and PROP taster status were not significantly correlated to BMI.125 Age may be one factor that influences taste status and BMI among Indians. A study among female Indian adolescents found that a relationship between PTC taste and a higher BMI did exist among 11-year-olds, but this association was lost among older aged groups.126 The same study also found that the PTC tasters among 14- to 16-year-olds had a higher mean percentage body fat, fat mass index and fat-free mass index. Thus, a large-scale multivariate analysis would be beneficial in determining key influential variables that affect food intake. Nutritionists can then use these key determinants to optimize dietary courses in order to help prevent or treat diseases such as obesity, dental caries and Parkinson’s disease, as well as improve patient adherence to dietary plans.
The strategy of developing personalized nutritional plans through TAS2R38 genotype status has also been previously outlined by Bray et al.127 The authors note that childhood eating habits can carry over into adulthood and that repeated dietary exposure to a food can lower intake reluctance and increase the tendency of consuming that food. The authors further remark that masking the bitter taste of certain vegetables through sauces, spices or healthy fats can be an additional strategy in increasing dietary intake. PROP tasters among children, for example, were found to consume 80% more raw broccoli when the vegetable was accompanied by a sauce.128 On the other hand, broccoli consumption with the addition of a sauce did not change among PROP non-tasters. Thus, adopting food intake strategies to improve dietary behavior can be adopted based on the PROP/PTC taste status or TAS2R38 genotype.
Given the extensive research conducted on bacterial quorum-sensing molecules activating T2R38 in respiratory and sinonasal infections, further investigation should be conducted on examining the efficacy of T2R agonists among patients with differing TAS2R38 variants. Theoretically, drugs that function through the activation of T2R may be less effective for bacterial infections in patients carrying the PAV haplotype, as this pathway would already be activated through T2R38. Several anti-inflammatory medicinal herbs are also known for containing bitter flavonoids that activate T2R1, -10, -14 and -46.129 Future clinical research studies may want to consider comparing differences in the efficacy of bitter among patients with varying TAS2R SNPs, as inconsistencies and heterogeneity in therapeutic efficacy of flavonoids for asthma have been noted across several studies.130 Similarly, MP-AzeFlu (Dymista®; spray of azelastine/fluticasone propionate) is a drug used for allergic rhinitis and its dilatory effects on pre-contracted airways are thought to be enhanced by T2R activation rather than through blocking histamine receptors.131 Investigations can be conducted on the specific receptor isoforms that MP-AzeFlu activates and whether its efficacy is affected by T2R polymorphisms of the host. Moreover, T2R38 may also be naturally triggered in allergic rhinitis patients due to their altered and potentially more pathogenic bacterial microbiome in the nasal passage.132–134
Since the prognosis of gram-negative bacteria-infected patients can also differ based on TAS2R38 variation, adopting early aggressive treatment regimens for patients with the AVI diplotype can be contemplated. For example, in CRS patients, the non-taster phenotype and AVI diplotype are associated with increased disease severity, more frequent sinus infections, lower quality of life, higher likelihood of undergoing surgical invention, higher CT scores of the paranasal sinuses, higher likelihood of cultural bacteria in nasal swab samples, higher likelihood of in vivo biofilm formation, more frequent respiratory exacerbations, and higher likelihood of having nasal polyposis.100–105,112 This body of evidences suggests a significantly worse prognosis for AVI/AVI or non-taster CRS patients. The future development of more aggressive, but safe treatment regimens for this patient group would be beneficial for improving prognosis.
Aside from phenotype and genotype variation, TAS2R expression levels must also be taken into consideration. A systematic review outlined numerous in vitro studies showing the anti-cancerous effects of agonist-mediated T2R activation in cancer cell lines and its potential usefulness in enhancing chemotherapeutic effects.10 Attempting to activate the T2R pathway would theoretically be ineffective in tumor types with downregulated TAS2R expression. Thus, determining increased TAS2R expression profiles in tumor biopsies may be a viable strategy for improving complementary or adjuvant therapies, given the multi-faceted role T2R agonists can have in cancer therapy. Similarly, differences in TAS2R expression levels in the skin signify that T2R agonist drugs can possibly have heterogeneous effects based on the T2R isoform being targeted. The use of T2R agonist cocktails to treat atopic eczema, as seen in a 2013 patent,135 should take into consideration individual skin types. Different combinations of T2R agonists can be provided to appropriately activate the receptor isoforms that have higher expression levels specific to the patient and skin region.
In summary, improving compliance to medication and nutritional dietary strategies can be personalized based on variations in bitter taste genotype and phenotype. The efficacy of T2R agonists in treating bacterial infections should be investigated across different TAS2R38 diplotypes. Better or more aggressive treatment strategies can be developed for CRS patients who have a poor prognosis due to the TAS2R38 AVI/AVI diplotype. Lastly, T2R agonists used to treat cancer or skin conditions must take into consideration individual TAS2R expression levels.
Abbreviations
AHL, acyl-homoserine lactone; AVI, alanine-valine-isoleucine; COVID-19, coronavirus disease 2019; CRS, chronic rhinosinusitis; CT, computed tomography; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-sensitive Staphylococcus aureus; PAV, proline-alanine-valine; PROP, 6-n-propylthiouracil; PTC, phenylthiocarbamide; SNP, single nucleotide polymorphism; T2R, bitter taste receptor; TAS2R, bitter taste receptor gene.
Funding
This work was supported by the Henan Provincial Medical Science and Technology Research Plan Joint Construction Project (LHGJ20210667).
Disclosure
Dr Weyland Cheng reports grants from Henan Provincial Medical Science and Technology Research Plan Joint Construction Project, during the conduct of the study. The authors report no conflicts of interest in this work.
References
1. Schioth HB, Fredriksson R. The GRAFS classification system of G-protein coupled receptors in comparative perspective. Gen Comp Endocrinol. 2005;142(1–2):94–101. doi:10.1016/j.ygcen.2004.12.018
2. Wooding SP, Ramirez VA, Behrens M. Bitter taste receptors: genes, evolution and health. Evol Med Public Health. 2021;9(1):431–447. doi:10.1093/emph/eoab031
3. Margolskee RF. Molecular mechanisms of bitter and sweet taste transduction. J Biol Chem. 2002;277(1):1–4. doi:10.1074/jbc.R100054200
4. Kinnamon SC, Finger TE. A taste for ATP: neurotransmission in taste buds. Front Cell Neurosci. 2013;7:264. doi:10.3389/fncel.2013.00264
5. Cheng W, Yao M, Liu F. Bitter taste receptor as a therapeutic target in orthopaedic disorders. Drug Des Devel Ther. 2021;15:895–903. doi:10.2147/DDDT.S289614
6. Bloxham CJ, Foster SR, Thomas WG. A bitter taste in your heart. Front Physiol. 2020;11:431. doi:10.3389/fphys.2020.00431
7. Parsa S, Mogharab V, Ebrahimi M, et al. COVID-19 as a worldwide selective event and bitter taste receptor polymorphisms: an ecological correlational study. Int J Biol Macromol. 2021;177:204–210. doi:10.1016/j.ijbiomac.2021.02.070
8. Lee RJ, Cohen NA. The emerging role of the bitter taste receptor T2R38 in upper respiratory infection and chronic rhinosinusitis. Am J Rhinol Allergy. 2013;27(4):283–286. doi:10.2500/ajra.2013.27.3911
9. Sakakibara M, Sumida H, Yanagida K, Miyasato S, Nakamura M, Sato S. Bitter taste receptor T2R38 is expressed on skin-infiltrating lymphocytes and regulates lymphocyte migration. Sci Rep. 2022;12(1):11790. doi:10.1038/s41598-022-15999-6
10. Zehentner S, Reiner AT, Grimm C, Somoza V. The role of bitter taste receptors in cancer: a systematic review. Cancers. 2021;13(23):5891. doi:10.3390/cancers13235891
11. Gopallawa I, Freund JR, Lee RJ. Bitter taste receptors stimulate phagocytosis in human macrophages through calcium, nitric oxide, and cyclic-GMP signaling. Cell Mol Life Sci. 2021;78(1):271–286. doi:10.1007/s00018-020-03494-y
12. Lee RJ, Cohen NA. Taste receptors in innate immunity. Cell Mol Life Sci. 2015;72(2):217–236. doi:10.1007/s00018-014-1736-7
13. Orsmark-Pietras C, James A, Konradsen JR, et al. Transcriptome analysis reveals upregulation of bitter taste receptors in severe asthmatics. Eur Respir J. 2013;42(1):65–78. doi:10.1183/09031936.00077712
14. Tran HTT, Herz C, Ruf P, Stetter R, Lamy E. Human T2R38 bitter taste receptor expression in resting and activated lymphocytes. Front Immunol. 2018;9:2949. doi:10.3389/fimmu.2018.02949
15. Kumar SA, Cheng W. A hypothesis: bitter taste receptors as a therapeutic target for the clinical symptoms of SARS-CoV-2. Pharmazie. 2021;76(2):43–54. doi:10.1691/ph.2021.0840
16. Singh N, Vrontakis M, Parkinson F, Chelikani P. Functional bitter taste receptors are expressed in brain cells. Biochem Biophys Res Commun. 2011;406(1):146–151. doi:10.1016/j.bbrc.2011.02.016
17. Wolfle U, Elsholz FA, Kersten A, Haarhaus B, Muller WE, Schempp CM. Expression and functional activity of the bitter taste receptors TAS2R1 and TAS2R38 in human keratinocytes. Skin Pharmacol Physiol. 2015;28(3):137–146. doi:10.1159/000367631
18. Wu SV, Rozengurt N, Yang M, Young SH, Sinnett-Smith J, Rozengurt E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc Natl Acad Sci U S A. 2002;99(4):2392–2397. doi:10.1073/pnas.042617699
19. Tarragon E, Moreno JJ. Polyphenols and taste 2 receptors. Physiological, pathophysiological and pharmacological implications. Biochem Pharmacol. 2020;178:114086. doi:10.1016/j.bcp.2020.114086
20. Kim UK, Jorgenson E, Coon H, Leppert M, Risch N, Drayna D. Positional cloning of the human quantitative trait locus underlying taste sensitivity to phenylthiocarbamide. Science. 2003;299(5610):1221–1225. doi:10.1126/science.1080190
21. Bufe B, Breslin PA, Kuhn C, et al. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr Biol. 2005;15(4):322–327. doi:10.1016/j.cub.2005.01.047
22. Adappa ND, Howland TJ, Palmer JN, et al. Genetics of the taste receptor T2R38 correlates with chronic rhinosinusitis necessitating surgical intervention. Int Forum Allergy Rhinol. 2013;3(3):184–187. doi:10.1002/alr.21140
23. Verbeurgt C, Veithen A, Carlot S, et al. The human bitter taste receptor T2R38 is broadly tuned for bacterial compounds. PLoS One. 2017;12(9):e0181302. doi:10.1371/journal.pone.0181302
24. Pawellek I, Grote V, Rzehak P, et al. Association of TAS2R38 variants with sweet food intake in children aged 1–6 years. Appetite. 2016;107:126–134. doi:10.1016/j.appet.2016.07.034
25. Riccio MP, Franco C, Negri R, et al. Is food refusal in autistic children related to TAS2R38 genotype? Autism Res. 2018;11(3):531–538. doi:10.1002/aur.1912
26. Cont G, Paviotti G, Montico M, et al. TAS2R38 bitter taste genotype is associated with complementary feeding behavior in infants. Genes Nutr. 2019;14:13. doi:10.1186/s12263-019-0640-z
27. Mennella JA, Roberts KM, Mathew PS, Reed DR. Children’s perceptions about medicines: individual differences and taste. BMC Pediatr. 2015;15:130. doi:10.1186/s12887-015-0447-z
28. Lipchock SV, Reed DR, Mennella JA. Relationship between bitter-taste receptor genotype and solid medication formulation usage among young children: a retrospective analysis. Clin Ther. 2012;34(3):728–733. doi:10.1016/j.clinthera.2012.02.006
29. Mennella JA, Pepino MY, Duke FF, Reed DR. Age modifies the genotype-phenotype relationship for the bitter receptor TAS2R38. BMC Genet. 2010;11:60. doi:10.1186/1471-2156-11-60
30. Andrews D, Salunke S, Cram A, et al. Bitter-blockers as a taste masking strategy: a systematic review towards their utility in pharmaceuticals. Eur J Pharm Biopharm. 2021;158:35–51. doi:10.1016/j.ejpb.2020.10.017
31. Hayes JE, Wallace MR, Knopik VS, Herbstman DM, Bartoshuk LM, Duffy VB. Allelic variation in TAS2R bitter receptor genes associates with variation in sensations from and ingestive behaviors toward common bitter beverages in adults. Chem Senses. 2011;36(3):311–319. doi:10.1093/chemse/bjq132
32. Dotson CD, Wallace MR, Bartoshuk LM, Logan HL. Variation in the gene TAS2R13 is associated with differences in alcohol consumption in patients with head and neck cancer. Chem Senses. 2012;37(8):737–744. doi:10.1093/chemse/bjs063
33. Duffy VB, Davidson AC, Kidd JR, et al. Bitter receptor gene (TAS2R38), 6-n-propylthiouracil (PROP) bitterness and alcohol intake. Alcohol Clin Exp Res. 2004;28(11):1629–1637. doi:10.1097/01.ALC.0000145789.55183.D4
34. Mikolajczyk-Stecyna J, Malinowska AM, Chmurzynska A. TAS2R38 and CA6 genetic polymorphisms, frequency of bitter food intake, and blood biomarkers among elderly woman. Appetite. 2017;116:57–64. doi:10.1016/j.appet.2017.04.029
35. Perna S, Riva A, Nicosanti G, et al. Association of the bitter taste receptor gene TAS2R38 (polymorphism RS713598) with sensory responsiveness, food preferences, biochemical parameters and body-composition markers. A cross-sectional study in Italy. Int J Food Sci Nutr. 2018;69(2):245–252. doi:10.1080/09637486.2017.1353954
36. Hinrichs AL, Wang JC, Bufe B, et al. Functional variant in a bitter-taste receptor (hTAS2R16) influences risk of alcohol dependence. Am J Hum Genet. 2006;78(1):103–111. doi:10.1086/499253
37. Wang JC, Hinrichs AL, Bertelsen S, et al. Functional variants in TAS2R38 and TAS2R16 influence alcohol consumption in high-risk families of African-American origin. Alcohol Clin Exp Res. 2007;31(2):209–215. doi:10.1111/j.1530-0277.2006.00297.x
38. Risso D, Morini G, Pagani L, et al. Genetic signature of differential sensitivity to stevioside in the Italian population. Genes Nutr. 2014;9(3):401. doi:10.1007/s12263-014-0401-y
39. Ooi SX, Lee PL, Law HY, Say YH. Bitter receptor gene (TAS2R38) P49A genotypes and their associations with aversion to vegetables and sweet/fat foods in Malaysian subjects. Asia Pac J Clin Nutr. 2010;19(4):491–498.
40. Calancie L, Keyserling TC, Taillie LS, et al. TAS2R38 predisposition to bitter taste associated with differential changes in vegetable intake in response to a community-based dietary intervention. G3 (Bethesda). 2018;8(6):2107–2119. doi:10.1534/g3.118.300547
41. Smith JL, Estus S, Lennie TA, Moser DK, Chung ML, Mudd-Martin G. TAS2R38 PAV haplotype predicts vegetable consumption in community-dwelling caucasian adults at risk for cardiovascular disease. Biol Res Nurs. 2020;22(3):326–333. doi:10.1177/1099800420913935
42. Choi JH. Variation in the TAS2R38 bitterness receptor gene was associated with food consumption and obesity risk in Koreans. Nutrients. 2019;11(9):1973. doi:10.3390/nu11091973
43. Cecati M, Vignini A, Borroni F, et al. TAS1R3 and TAS2R38 polymorphisms affect sweet taste perception: an observational study on healthy and obese subjects. Nutrients. 2022;14(9):1711. doi:10.3390/nu14091711
44. Oncken C, Feinn R, Covault J, et al. Genetic vulnerability to menthol cigarette preference in women. Nicotine Tob Res. 2015;17(12):1416–1420. doi:10.1093/ntr/ntv042
45. Risso D, Sainz E, Gutierrez J, Kirchner T, Niaura R, Drayna D. Association of TAS2R38 haplotypes and menthol cigarette preference in an African American cohort. Nicotine Tob Res. 2017;19(4):493–494. doi:10.1093/ntr/ntw275
46. Baker AN, Miranda AM, Garneau NL, Hayes JE. Self-reported smoking status, TAS2R38 variants, and propylthiouracil phenotype: an exploratory crowdsourced cohort study. Chem Senses. 2018;43(8):617–625. doi:10.1093/chemse/bjy053
47. Robino A, Rosso N, Guerra M, et al. Taste perception and expression in stomach of bitter taste receptor tas2r38 in obese and lean subjects. Appetite. 2021;166:105595. doi:10.1016/j.appet.2021.105595
48. Karmous I, Plesnik J, Khan AS, et al. Orosensory detection of bitter in fat-taster healthy and obese participants: genetic polymorphism of CD36 and TAS2R38. Clin Nutr. 2018;37(1):313–320. doi:10.1016/j.clnu.2017.06.004
49. Khan AM, Al-Jandan B, Bugshan A, et al. Correlation of PTC taste status with fungiform papillae count and body mass index in smokers and non-smokers of eastern province, Saudi Arabia. Int J Environ Res Public Health. 2020;17(16):5792. doi:10.3390/ijerph17165792
50. Chupeerach C, Tapanee P, On-Nom N, et al. The influence of TAS2R38 bitter taste gene polymorphisms on obesity risk in three racially diverse groups. Biomedicine. 2021;11(3):43–49. doi:10.37796/2211-8039.1175
51. Mobley C, Marshall TA, Milgrom P, Coldwell SE. The contribution of dietary factors to dental caries and disparities in caries. Acad Pediatr. 2009;9(6):410–414. doi:10.1016/j.acap.2009.09.008
52. Dioszegi J, Llanaj E, Adany R. Genetic background of taste perception, taste preferences, and its nutritional implications: a systematic review. Front Genet. 2019;10:1272. doi:10.3389/fgene.2019.01272
53. Lin BP. Caries experience in children with various genetic sensitivity levels to the bitter taste of 6-n-propylthiouracil (PROP): a pilot study. Pediatr Dent. 2003;25(1):37–42.
54. Rupesh S, Nayak UA. Genetic sensitivity to the bitter taste of 6-n propylthiouracil: a new risk determinant for dental caries in children. J Indian Soc Pedod Prev Dent. 2006;24(2):63–68. doi:10.4103/0970-4388.26018
55. Verma P, Shetty V, Hegde AM. Propylthiouracil (PROP)--a tool to determine taster status in relation to caries experience, streptococcus mutans levels and dietary preferences in children. J Clin Pediatr Dent. 2006;31(2):113–117. doi:10.17796/jcpd.31.2.34302r2857511268
56. Furquim TR, Poli-Frederico RC, Maciel SM, Gonini-Junior A, Walter LR. Sensitivity to bitter and sweet taste perception in schoolchildren and their relation to dental caries. Oral Health Prev Dent. 2010;8(3):253–259.
57. Jyothirmai J, Naganandini S, Aradhya S. Caries experience in 15-year-old school children in Bangalore with inherited taste sensitivity levels to 6-n-propylthiouracil: an observational study. J Investig Clin Dent. 2011;2(1):51–56. doi:10.1111/j.2041-1626.2010.00029.x
58. Oter B, Ulukapi I, Ulukapi H, Topcuoglu N, Cildir S. The relation between 6-n-propylthiouracil sensitivity and caries activity in schoolchildren. Caries Res. 2011;45(6):556–560. doi:10.1159/000332432
59. Pidamale R, Sowmya B, Thomas A, Jose T. Genetic sensitivity to bitter taste of 6-n Propylthiouracil: a useful diagnostic aid to detect early childhood caries in pre-school children. Indian J Hum Genet. 2012;18(1):101–105. doi:10.4103/0971-6866.96672
60. Pidamale R, Sowmya B, Thomas A, Jose T, Madhusudan KK, Prasad G. Association between early childhood caries, streptococcus mutans level and genetic sensitivity levels to the bitter taste of, 6-N propylthiouracil among the children below 71 months of age. Dent Res J. 2012;9(6):730–734.
61. Alanzi A, Minah G, Romberg E, Catalanotto F, Bartoshuk L, Tinanoff N. Mothers’ taste perceptions and their preschool children’s dental caries experiences. Pediatr Dent. 2013;35(7):510–514.
62. Shetty V, Hegde AM, Hegde AM. PROP test: prediction of caries risk by genetic taste perception among the visually impaired children. Spec Care Dentist. 2014;34(1):34–40. doi:10.1111/j.1754-4505.2012.00307.x
63. Lakshmi CR, Radhika D, Prabhat M, Bhavana SM, Sai Madhavi N. Association between genetic taste sensitivity, 2D:4D ratio, dental caries prevalence, and salivary flow rate in 6–14-year-old children: a cross-sectional study. J Dent Res Dent Clin Dent Prospects. 2016;10(3):142–147. doi:10.15171/joddd.2016.023
64. Vandal VB, Noorani H, Shivaprakash PK, Walikar B. Genetic specificity to 6-n-propylthiouracil and its association to dental caries: a comparative study. J Indian Soc Pedod Prev Dent. 2017;35(1):83–85. doi:10.4103/0970-4388.199233
65. Dusseja SH, Rao D, Panwar S, Ameen S. Determining caries risk susceptibility in children of Udaipur city with genetic taste sensitivity and hormonal fingerprint. J Indian Soc Pedod Prev Dent. 2021;39(1):36–41. doi:10.4103/jisppd.jisppd_527_20
66. Nellamakkada K, Patil SS, Kakanur M, Kumar RS, Thakur R. Association of mothers’ genetic taste perception to eating habits and its influence on early childhood caries in preschool children: an analytical study. Int J Clin Pediatr Dent. 2022;15(Suppl 2):S135–S9. doi:10.5005/jp-journals-10005-2139
67. Wendell S, Wang X, Brown M, et al. Taste genes associated with dental caries. J Dent Res. 2010;89(11):1198–1202. doi:10.1177/0022034510381502
68. Khimsuksri S, Paphangkorakit J, Pitiphat W, Coldwell SE. TAS2R38 polymorphisms and oral diseases in Thais: a cross-sectional study. BMC Oral Health. 2022;22(1):21. doi:10.1186/s12903-022-02043-2
69. Kilic M, Gurbuz T, Kahraman CY, Cayir A, Bilgic A, Kurt Y. Relationship between the TAS2R38 and TAS1R2 polymorphisms and the dental status in obese children. Dent Med Probl. 2022;59(2):233–240. doi:10.17219/dmp/143252
70. Yeomans MR, Vi C, Mohammed N, Armitage RM. Re-evaluating how sweet-liking and PROP-tasting are related. Physiol Behav. 2022;246:113702. doi:10.1016/j.physbeh.2022.113702
71. Campa D, De Rango F, Carrai M, et al. Bitter taste receptor polymorphisms and human aging. PLoS One. 2012;7(11):e45232. doi:10.1371/journal.pone.0045232
72. Malovini A, Accardi G, Aiello A, et al. Taste receptors, innate immunity and longevity: the case of TAS2R16 gene. Immun Ageing. 2019;16:5. doi:10.1186/s12979-019-0146-y
73. Melis M, Errigo A, Crnjar R, Pes GM, Tomassini Barbarossa I. TAS2R38 bitter taste receptor and attainment of exceptional longevity. Sci Rep. 2019;9(1):18047. doi:10.1038/s41598-019-54604-1
74. Di Bona D, Malovini A, Accardi G, et al. Taste receptor polymorphisms and longevity: a systematic review and meta-analysis. Aging Clin Exp Res. 2021;33(9):2369–2377. doi:10.1007/s40520-020-01745-3
75. Cossu G, Melis M, Sarchioto M, et al. 6-n-propylthiouracil taste disruption and TAS2R38 nontasting form in Parkinson’s disease. Mov Disord. 2018;33(8):1331–1339. doi:10.1002/mds.27391
76. Moberg PJ, Balderston CC, Rick JH, et al. Phenylthiocarbamide (PTC) perception in Parkinson disease. Cogn Behav Neurol. 2007;20(3):145–148. doi:10.1097/WNN.0b013e31812570c3
77. Vascellari S, Melis M, Cossu G, et al. Genetic variants of TAS2R38 bitter taste receptor associate with distinct gut microbiota traits in Parkinson’s disease: a pilot study. Int J Biol Macromol. 2020;165(Pt A):665–674. doi:10.1016/j.ijbiomac.2020.09.056
78. Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9(1):3294. doi:10.1038/s41467-018-05470-4
79. Chen QQ, Haikal C, Li W, Li JY. Gut inflammation in association with pathogenesis of parkinson’s disease. Front Mol Neurosci. 2019;12:218. doi:10.3389/fnmol.2019.00218
80. Alexopoulos A. Role of the gut microbiome and bile acids. Dtsch Arztebl Int. 2022;119(35–36):609. doi:10.3238/arztebl.m2022.0177
81. Singh N, Shaik FA, Myal Y, Chelikani P. Chemosensory bitter taste receptors T2R4 and T2R14 activation attenuates proliferation and migration of breast cancer cells. Mol Cell Biochem. 2020;465(1–2):199–214. doi:10.1007/s11010-019-03679-5
82. Martin LTP, Nachtigal MW, Selman T, et al. Bitter taste receptors are expressed in human epithelial ovarian and prostate cancers cells and noscapine stimulation impacts cell survival. Mol Cell Biochem. 2019;454(1–2):203–214. doi:10.1007/s11010-018-3464-z
83. Seo Y, Kim YS, Lee KE, Park TH, Kim Y. Anti-cancer stemness and anti-invasive activity of bitter taste receptors, TAS2R8 and TAS2R10, in human neuroblastoma cells. PLoS One. 2017;12(5):e0176851. doi:10.1371/journal.pone.0176851
84. Stern L, Giese N, Hackert T, et al. Overcoming chemoresistance in pancreatic cancer cells: role of the bitter taste receptor T2R10. J Cancer. 2018;9(4):711–725. doi:10.7150/jca.21803
85. Salvestrini V, Ciciarello M, Pensato V, et al. Denatonium as a bitter taste receptor agonist modifies transcriptomic profile and functions of acute myeloid leukemia cells. Front Oncol. 2020;10:1225. doi:10.3389/fonc.2020.01225
86. Carey RM, McMahon DB, Miller ZA, et al. T2R bitter taste receptors regulate apoptosis and may be associated with survival in head and neck squamous cell carcinoma. Mol Oncol. 2022;16(7):1474–1492. doi:10.1002/1878-0261.13131
87. Reyes-Farias M, Carrasco-Pozo C. The anti-cancer effect of quercetin: molecular implications in cancer metabolism. Int J Mol Sci. 2019;20(13):3177. doi:10.3390/ijms20133177
88. Wu Y, Xu J, Liu Y, Zeng Y, Wu G. A review on anti-tumor mechanisms of coumarins. Front Oncol. 2020;10:592853. doi:10.3389/fonc.2020.592853
89. Mahmoudian M, Rahimi-Moghaddam P. The anti-cancer activity of noscapine: a review. Recent Pat Anticancer Drug Discov. 2009;4(1):92–97. doi:10.2174/157489209787002524
90. Xu Y, Tong Y, Ying J, et al. Chrysin induces cell growth arrest, apoptosis, and ER stress and inhibits the activation of STAT3 through the generation of ROS in bladder cancer cells. Oncol Lett. 2018;15(6):9117–9125. doi:10.3892/ol.2018.8522
91. Rauf A, Shariati MA, Imran M, et al. Comprehensive review on naringenin and naringin polyphenols as a potent anticancer agent. Environ Sci Pollut Res Int. 2022;29(21):31025–31041. doi:10.1007/s11356-022-18754-6
92. Singh N, Chakraborty R, Bhullar RP, Chelikani P. Differential expression of bitter taste receptors in non-cancerous breast epithelial and breast cancer cells. Biochem Biophys Res Commun. 2014;446(2):499–503. doi:10.1016/j.bbrc.2014.02.140
93. Choi JH, Lee J, Choi IJ, Kim YW, Ryu KW, Kim J. Genetic variation in the TAS2R38 bitter taste receptor and gastric cancer risk in Koreans. Sci Rep. 2016;6:26904. doi:10.1038/srep26904
94. Yamaki M, Saito H, Isono K, et al. Genotyping analysis of bitter-taste receptor genes TAS2R38 and TAS2R46 in Japanese patients with gastrointestinal cancers. J Nutr Sci Vitaminol (Tokyo). 2017;63(2):148–154. doi:10.3177/jnsv.63.148
95. Carrai M, Steinke V, Vodicka P, et al. Association between TAS2R38 gene polymorphisms and colorectal cancer risk: a case-control study in two independent populations of Caucasian origin. PLoS One. 2011;6(6):e20464. doi:10.1371/journal.pone.0020464
96. Choi JH, Lee J, Oh JH, et al. Variations in the bitterness perception-related genes TAS2R38 and CA6 modify the risk for colorectal cancer in Koreans. Oncotarget. 2017;8(13):21253–21265. doi:10.18632/oncotarget.15512
97. Giaccherini M, Rizzato C, Gentiluomo M, et al. TAS2R38 polymorphisms, Helicobacter pylori infection and susceptibility to gastric cancer and premalignant gastric lesions. Eur J Cancer Prev. 2021;31:401–407.
98. Barontini J, Antinucci M, Tofanelli S, et al. Association between polymorphisms of TAS2R16 and susceptibility to colorectal cancer. BMC Gastroenterol. 2017;17(1):104. doi:10.1186/s12876-017-0659-9
99. Campa D, Vodicka P, Pardini B, et al. A gene-wide investigation on polymorphisms in the taste receptor 2R14 (TAS2R14) and susceptibility to colorectal cancer. BMC Med Genet. 2010;11:88. doi:10.1186/1471-2350-11-88
100. Rowan NR, Soler ZM, Othieno F, Storck KA, Smith TL, Schlosser RJ. Impact of bitter taste receptor phenotype upon clinical presentation in chronic rhinosinusitis. Int Forum Allergy Rhinol. 2018;8(9):1013–1020. doi:10.1002/alr.22138
101. Farquhar DR, Kovatch KJ, Palmer JN, Shofer FS, Adappa ND, Cohen NA. Phenylthiocarbamide taste sensitivity is associated with sinonasal symptoms in healthy adults. Int Forum Allergy Rhinol. 2015;5(2):111–118. doi:10.1002/alr.21437
102. Adappa ND, Zhang Z, Palmer JN, et al. The bitter taste receptor T2R38 is an independent risk factor for chronic rhinosinusitis requiring sinus surgery. Int Forum Allergy Rhinol. 2014;4(1):3–7. doi:10.1002/alr.21253
103. Dzaman K, Zagor M, Stachowiak M, et al. The correlation of TAS2R38 gene variants with higher risk for chronic rhinosinusitis in Polish patients. Otolaryngol Pol. 2016;70(5):13–18. doi:10.5604/00306657.1209438
104. Rom DI, Christensen JM, Alvarado R, Sacks R, Harvey RJ. The impact of bitter taste receptor genetics on culturable bacteria in chronic rhinosinusitis. Rhinology. 2017;55(1):90–94. doi:10.4193/Rhin16.181
105. Cantone E, Negri R, Roscetto E, et al. In vivo biofilm formation, gram-negative infections and TAS2R38 polymorphisms in CRSw NP patients. Laryngoscope. 2018;128(10):E339–E45. doi:10.1002/lary.27175
106. Purnell PR, Addicks BL, Zalzal HG, et al. Single nucleotide polymorphisms in chemosensory pathway genes GNB3, TAS2R19, and TAS2R38 are associated with chronic rhinosinusitis. Int Arch Allergy Immunol. 2019;180(1):72–78. doi:10.1159/000499875
107. Zborowska-Piskadlo K, Stachowiak M, Rusetska N, Sarnowska E, Siedlecki J, Dzaman K. The expression of bitter taste receptor TAS2R38 in patients with chronic rhinosinusitis. Arch Immunol Ther Exp (Warsz). 2020;68(5):26. doi:10.1007/s00005-020-00593-3
108. Piskadlo-Zborowska K, Stachowiak M, Sarnowska E, Jowik R, Dzaman K. Assessment of the effect of inflammatory changes and allergic reaction on TAS2R38 receptor expression in patients with chronic sinusitis (CRS). Otolaryngol Pol. 2020;74(5):17–23. doi:10.5604/01.3001.0014.1474
109. Jeruzal-Swiatecka J, Borkowska E, Laszczych M, Nowicka Z, Pietruszewska W. TAS2R38 bitter taste receptor expression in chronic rhinosinusitis with nasal polyps: new data on polypoid tissue. Int J Mol Sci. 2022;23(13):7345. doi:10.3390/ijms23137345
110. Castaldo A, Cernera G, Iacotucci P, et al. TAS2R38 is a novel modifier gene in patients with cystic fibrosis. Sci Rep. 2020;10(1):5806. doi:10.1038/s41598-020-62747-9
111. Piatti G, Ambrosetti U, Robino A, Girotto G, Gasparini P. Primary ciliary dyskinesia: the impact of taste receptor (TAS2R38) gene polymorphisms on disease outcome and severity. Int Arch Allergy Immunol. 2020;181(9):727–731. doi:10.1159/000508938
112. Piatti G, Ambrosetti U, Alde M, Girotto G, Concas MP, Torretta S. Chronic rhinosinusitis: T2r38 genotyping and nasal cytology in primary ciliary dyskinesia. Laryngoscope. 2022;133:248–254. doi:10.1002/lary.30112
113. Yoon SY, Shin ES, Park SY, et al. Association between polymorphisms in bitter taste receptor genes and clinical features in Korean asthmatics. Respiration. 2016;91(2):141–150. doi:10.1159/000443796
114. Barham HP, Taha MA, Hall CA. Does phenotypic expression of bitter taste receptor T2R38 show association with COVID-19 severity? Int Forum Allergy Rhinol. 2020;10(11):1255–1257. doi:10.1002/alr.22692
115. Barham HP, Taha MA, Broyles ST, Stevenson MM, Zito BA, Hall CA. Association between bitter taste receptor phenotype and clinical outcomes among patients with COVID-19. JAMA Netw Open. 2021;4(5):e2111410. doi:10.1001/jamanetworkopen.2021.11410
116. Risso D, Carmagnola D, Morini G, et al. Distribution of TAS2R38 bitter taste receptor phenotype and haplotypes among COVID-19 patients. Sci Rep. 2022;12(1):7381. doi:10.1038/s41598-022-10747-2
117. Shaw L, Mansfield C, Colquitt L, et al. Personalized expression of bitter ‘taste’ receptors in human skin. PLoS One. 2018;13(10):e0205322. doi:10.1371/journal.pone.0205322
118. Raslan Z, Naseem KM. The control of blood platelets by cAMP signalling. Biochem Soc Trans. 2014;42(2):289–294. doi:10.1042/BST20130278
119. Carey JR, Suslick KS, Hulkower KI, et al. Rapid identification of bacteria with a disposable colorimetric sensing array. J Am Chem Soc. 2011;133(19):7571–7576. doi:10.1021/ja201634d
120. Bos LD, Sterk PJ, Schultz MJ. Volatile metabolites of pathogens: a systematic review. PLoS Pathog. 2013;9(5):e1003311. doi:10.1371/journal.ppat.1003311
121. Filipiak W, Sponring A, Baur MM, et al. Molecular analysis of volatile metabolites released specifically by Staphylococcus aureus and Pseudomonas aeruginosa. BMC Microbiol. 2012;12:113. doi:10.1186/1471-2180-12-113
122. Filipiak W, Sponring A, Filipiak A, et al. Volatile Organic Compounds (VOCs) released by pathogenic microorganisms in vitro: potential breath biomarkers for early-stage diagnosis of disease. In Amann A, Smith D, eds. Volatile Biomarkers Non-Invasive Diagnosis in Physiology and Medicine. Elsevier; 2013:463–512.
123. Nolden AA, Hayes JE, Feeney EL. Variation in TAS2R receptor genes explains differential bitterness of two common antibiotics. Front Genet. 2022;13:960154. doi:10.3389/fgene.2022.960154
124. Tepper BJ, Banni S, Melis M, Crnjar R, Tomassini Barbarossa I. Genetic sensitivity to the bitter taste of 6-n-propylthiouracil (PROP) and its association with physiological mechanisms controlling body mass index (BMI). Nutrients. 2014;6(9):3363–3381. doi:10.3390/nu6093363
125. Deshaware S, Singhal R. Genetic variation in bitter taste receptor gene TAS2R38, PROP taster status and their association with body mass index and food preferences in Indian population. Gene. 2017;627:363–368. doi:10.1016/j.gene.2017.06.047
126. Sharma K, Kaur GK. PTC bitter taste genetic polymorphism, food choices, physical growth in body height and body fat related traits among adolescent girls from Kangra Valley, Himachal Pradesh (India). Ann Hum Biol. 2014;41(1):29–39. doi:10.3109/03014460.2013.822929
127. Bray SC, Carek PJ. How bitter taste influences nutrition and health in primary care. J Family Med Prim Care. 2020;9(7):3205–3208. doi:10.4103/jfmpc.jfmpc_305_20
128. Fisher JO, Mennella JA, Hughes SO, Liu Y, Mendoza PM, Patrick H. Offering “dip” promotes intake of a moderately-liked raw vegetable among preschoolers with genetic sensitivity to bitterness. J Acad Nutr Diet. 2012;112(2):235–245. doi:10.1016/j.jada.2011.08.032
129. Zhang Y-X, Wang X, Wanga S-F, Zhang Y-L, Qiao Y-J. Traditional Chinese Bitter Flavor theory: is there any relation with taste type II receptors? Eur J Integr Med. 2016;8(6):980–990. doi:10.1016/j.eujim.2016.04.011
130. de Almeida Brasiel PG, Guimaraes FV, Rodrigues PM, Bou-Habib DC, Carvalho VF. Therapeutic efficacy of flavonoids in allergies: a systematic review of randomized controlled trials. J Immunol Res. 2022;2022:8191253. doi:10.1155/2022/8191253
131. Ekstedt S, Kumlien Georen S, Cardell LO. Effects of MP-AzeFlu enhanced by activation of bitter taste receptor TAS2R. Allergy Asthma Clin Immunol. 2020;16:45. doi:10.1186/s13223-020-00438-w
132. Gluck U, Gebbers JO. Local pathogenic bacteria in allergic rhinitis: a novel concept of its pathogenesis. ORL J Otorhinolaryngol Relat Spec. 2003;65(4):202–205. doi:10.1159/000073115
133. Bender ME, Read TD, Edwards TS, et al. A comparison of the bacterial nasal microbiome in allergic rhinitis patients before and after immunotherapy. Laryngoscope. 2020;130(12):E882–E8. doi:10.1002/lary.28599
134. Gan W, Yang F, Meng J, Liu F, Liu S, Xian J. Comparing the nasal bacterial microbiome diversity of allergic rhinitis, chronic rhinosinusitis and control subjects. Eur Arch Otorhinolaryngol. 2021;278(3):711–718. doi:10.1007/s00405-020-06311-1
135. Schempp C, Woelfle U. Bitter taste receptor agonists for topical use patent WO2013112865A1. 2013.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.