Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 18
Circulating Interleukin-22 Is a Biomarker for Newly Diagnosed Type 2 Diabetes Mellitus and Associated with Hypoglycemic Effect of Sitagliptin
Authors Sun P ![]() , Xiao Y, Dong Y, Feng Y, Zheng H
, Xiao Y, Dong Y, Feng Y, Zheng H ![]() , Liao X
, Liao X ![]()
Received 13 December 2024
Accepted for publication 25 February 2025
Published 3 March 2025 Volume 2025:18 Pages 703—713
DOI https://doi.org/10.2147/DMSO.S509866
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rebecca Baqiyyah Conway
Peiye Sun,* Yuxi Xiao,* Yuan Dong, Yixiang Feng, Hongting Zheng, Xiaoyu Liao
Department of Endocrinology, Translational Research of Diabetes Key Laboratory of Chongqing Education Commission of China, the second Affiliated Hospital of Army Medical University, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaoyu Liao; Hongting Zheng, Department of Endocrinology, Translational Research of Diabetes Key Laboratory of Chongqing Education Commission of China, the second Affiliated Hospital of Army Medical University, Chongqing, 400037, People’s Republic of China, Tel +8602368774669 ; +8602368755709, Fax +8602368755707, Email [email protected]; [email protected]
Purpose: Interleukin-22 (IL-22) has been demonstrated to be involved in the regulation of glucose metabolism, insulin resistance and inflammation response, which indicates that IL-22 might be associated with the occurrence and progression of diabetes. This study aimed to assess serum IL-22 levels in participants with type 2 diabetes mellitus (T2DM) and analyze the association between IL-22 levels and T2DM risk.
Methods: Serum IL-22 concentrations of recruited healthy participants (n=48), newly diagnosed T2DM participants (n=46), and T2DM participants receiving placebo (n=7) or dipeptidyl peptidase-4 inhibitors (DPP-4i) sitagliptin monotherapy (n=7) were measured using a commercial enzyme-linked immunosorbent assay (ELISA) kit. Mice fed a high-fat diet (HFD) were administered sitagliptin and evaluated for IL-22 and intestinal inflammation-related indicators.
Results: Serum IL-22 levels were higher in the T2DM group (127.16 ± 75.35) than in healthy controls (69.18 ± 32.83, p < 0.001), significantly negatively correlated with high-density lipoprotein cholesterol (HDL-C), and positively correlated with body mass index (BMI), glycosylated hemoglobin (HbA1c) and fasting plasma glucose (FPG), regardless of adjustment for sex and age. Multivariate logistic regression analysis showed that serum IL-22 levels were associated with the risk of T2DM (OR = 2.37, 95% CI = 1.27– 4.42, p = 0.007). Additionally, sitagliptin treatment decreased the levels of IL-22 in the serum and colon tissues of T2DM participants and HFD mice. Moreover, intestinal inflammation was improved, and retinoid acid-related orphan receptor γt (RORγt, a marker of Th17 cells)- positive cells in the colon of HFD mice were decreased after sitagliptin treatment, which might be related to the reduction of IL-22.
Conclusion: Serum IL-22 is a significant independent risk factor for T2DM, implying that circulating IL-22 may be a predictive biomarker and therapeutic target for T2DM.
Keywords: IL-22, type 2 diabetes, sitagliptin, intestinal inflammation
Introduction
Type 2 Diabetes (T2DM) is a major global epidemiological event, and more than 500 million adults around the world are suffering from the disease 1 (International Diabetes Federation Diabetes Atlas 2021. IDF Atlas 10th edition. Available at: https://www.diabetesatlas.org). Various factors, such as genetics, diet, and the environment, are involved in the occurrence and development of T2DM. However, the etiology and pathogenesis of T2DM are not fully understood. Accumulating evidence has demonstrated that T2DM is accompanied by a chronic state of low-grade inflammation and an imbalance in the composition and function of intestinal flora.2–4
The dysbiosis of intestinal flora in individuals with obesity and T2DM is mainly manifested by an increase in the ratio of Firmicutes to Bacteroidetes, which impairs the integrity of the intestinal barrier and leads to the infiltration of lipopolysaccharide (LPS).5 LPS, a cell wall component of gram-negative bacteria, can affect the ratio of Treg/Th17 cells to induce intestinal inflammation.6 Upon entrance into circulation through the impaired intestinal barrier, some bacterial products and Th17 cells may induce chronic inflammation and metabolic disorders in metabolic tissues such as visceral adipose tissue and the liver.5,7 A chronic inflammatory state in T2DM individuals has been reported to be associated with an increase in the percentage of circulating Th17 cells and a decrease in the percentage of Treg cells.8 Moreover, studies have shown that the Th17 cells produced interleukin-22 (IL-22) was involved in the regulation of insulin resistance, as well as glucose metabolism through affecting inflammatory response. IL-22 promoted the production of IL-1β by activating the c-Jun pathway to induce pro-IL-1β transcription in macrophages, subsequently inducing insulin resistance.9 Additionally, IL-22 treatment significantly inhibited the insulin-mediated glucose uptake in isolated rat soleus muscles.10 In primary hepatocytes, IL-22 treatment reduced the phosphorylation level of Akt, a key protein kinase in the insulin signaling pathway.10 Contrary to the aforementioned view, recent studies reported that IL-22 has protective roles in inflammation and diabetes. Zhang Peng et al found that administration of recombinant IL-22 improved liver steatosis, inflammation and insulin resistance in MASLD.11 Similarly, it has been reported that targeting IL-22 in the pancreas and liver reduced hyperglycemia by improving β-cell function and insulin secretion.12 These discrepancies may be attributed to differences in experimental models and the diversity of signaling pathways. These studies indicated that IL-22 was associated with insulin resistance, and might be involved in the pathogenesis of metabolic diseases such as obesity and diabetes.
Our previous study showed that the dipeptidyl peptidase-4 inhibitors (DPP-4i) sitagliptin, an oral antidiabetic agent, improved the dysbiosis of gut microbiota in T2DM individuals and high-fat diet (HFD) mice and induced a pattern shift of microbial metabolites, including short-chain fatty acids (SCFAs), branched-chain amino acids, and tryptophan derivatives, which might mediate glucose homeostasis.13,14 Among the significantly increased metabolites, indole-3-acetic acid (IAA, a tryptophan derivative) has been reported to reduce the proportion of Th17 cells and increase the proportion of Treg cells in the lamina propria of the ileum in a mouse model of ankylosing spondylitis. Consequently, IAA reduced the production of pro-inflammatory factors and improved the intestinal barrier.15 Likewise, sitagliptin has also been found to directly reduce the proportion of Th17 cells in the peripheral blood of individuals and mice with diabetes,16,17 suggesting that sitagliptin might play a role in improving inflammation while lowering blood glucose levels. In fact, we initially observed that sitagliptin treatment reduced the number of retinoid acid-related orphan receptor γt (RORγt) (a marker of Th17 cells) -positive cells in colon tissues of HFD mice. Therefore, we speculated that Th17 produced IL-22 might be associated with the pathogenesis of T2DM and improvement of intestinal inflammation mediated by sitagliptin.
In the current study, to investigate the association between IL-22 and the risk of T2DM, we recruited a subgroup of our previous study including healthy controls and newly diagnosed T2DM participants. We first analyzed the association between circulating IL-22 levels and the risk of T2DM, and the hypoglycemic effect of DPP-4i sitagliptin in newly diagnosed T2DM individuals and HFD mice. Moreover, we assessed the relationship between IL-22 levels and intestinal inflammation in the HFD mice.
Materials and Methods
Healthy and T2DM Participants
This study enrolled ninety-four individuals. Individuals with T2DM were newly diagnosed based on the 1998 World Health Organization diagnostic criteria:18 fasting plasma glucose (FPG) ≥ 7.0 mmol/L, 2-hour postprandial plasma glucose (2hPG) ≥ 11.1 mmol/L or both. The exclusion criteria were as follows: (i) presence of acute or chronic complications of diabetes; (ii) pregnancy; and (iii) history of chronic physical/mental diseases such as cancer, Alzheimer’s disease, or Parkinson’s disease. The experimental protocol was approved by the Ethics Committee of the Second Affiliated Hospital of the Army Medical University and was registered online (Clinical Trial Registry Number ChiCTR-ROC-17010719). Written informed consent was obtained from all the participants.
Sitagliptin treatment participants were recruited from our previous study.13 Briefly, one group of newly diagnosed T2DM participants was treated with sitagliptin (n = 7), and the other group was treated with placebo (n = 7). Sitagliptin (Merck Sharp & Dohme) was administered at 100 mg/d. The detailed inclusion and exclusion criteria for T2DM were described in our previous study. Written informed consent was obtained from all the participants. The study was approved by the Ethics Committee of the Second Affiliated Hospital of the Army Medical University and was registered online (ChiCTR-OPC-17010757).
The current study involves secondary analysis of serum samples obtained from 94 participants from our previous studies,14,19 and 14 participants from another clinical study.13 Enzyme-linked immunosorbent assay (ELISA) of serum IL-22 was performed for each set of samples.
In the study, power calculation was performed using PASS software (version 15.0, NCSS, Silver Spring, Md) to ensure sufficient statistical power for detecting meaningful differences in the subgroup analysis. The calculation was based on a significance level (α) of 0.01, and a statistical power of 0.9. Based on these parameters, the required sample size for the healthy group and the T2DM group was estimated to be 40 participants, to achieve adequate power to detect the differences in IL-22 levels between subgroups.
Animal Study
C57BL/6 male mice were purchased from GemPharmatech Co., Ltd., China, and fed a normal diet (ND) or a HFD (60% fat, 20% protein, 20% carbohydrate (kcal/100 g), Research Diets, #D12492) for 14 weeks. HFD mice were divided into two groups according to the method of stratified randomization, to match weight and fasting blood glucose (FBG), and treated with 4 g/kg of sitagliptin or vehicle for the last 4 weeks during the HFD period. The animal experiment was conducted in strict adherence to the Guide for the Care and Use of Laboratory Animals, and in accordance with protocols approved by the Laboratory Animal Welfare and Ethics Committee of the Army Medical University (Approval No: AMUWEC20234810).
Measurement of Serum IL-22 Levels
The levels of cytokines (such as IL-22) might vary over time, and be influenced by biological rhythms, such as diurnal cycles, and differences in sampling time may lead to fluctuations in IL-22 measurement values. Therefore, for both human and mouse samples, we ensured an 8–12 hour fasting period before blood collection in the morning. Then, the blood samples were centrifuged at 3000 rpm for 15 minutes at 4°C to separate the serum, and immediately stored at −80 °C.
Serum IL-22 concentrations in T2DM and healthy participants were measured using a commercial ELISA kit according to the manufacturer’s instructions (Human IL-22 ELISA kit, Proteintech Group, Inc., #KE00008). The detection sensitivity of the kit was 14.2 pg/mL. The intra- and inter-assay coefficients of variation (CV) were lower than 8.1%. All the samples were diluted 2 times before detection. No significant cross-reactivity or interference was observed during this assay.
Serum IL-22 concentrations in ND and HFD mice were measured using an ELISA kit (Mouse IL-22 ELISA kit, Proteintech Group, Inc., #KE10101). The detection sensitivity of this kit was 0.4 pg/mL. The intra-assay CV was lower than 2.7%, and the inter-assay CV was lower than 5.5%. No significant cross-reactivity or interference was observed during this assay.
Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted from the colon tissues using RNAiso Plus (Takara, #9109). One microgram of total RNA was reverse-transcribed into cDNA using the PrimeScript RT Reagent Kit with gDNA Eraser (Takara, # RR047A). qRT-PCR was performed using the TB Green Premix (Takara, # RR820A) on a CFX Connect Real-Time System (Bio-Rad). The relative mRNA expression level was calculated using the following equation: Relative expression = 2− (Ct (target gene) − Ct (reference gene)). GAPDH was used as an internal reference gene for normalization. Primer sequences used are listed in Supplementary Table 1.
Hematoxylin-Eosin (H&E) and Immuno-Histochemical (IHC) Staining
Colon tissues from ND and HFD mice were collected using the Swiss-rolling method.20 Briefly, the colon tissues were fixed in 4% paraformaldehyde, embedded in paraffin and sectioned. Tissue slides (4 μm) were deparaffinized in xylene, rehydrated with ethanol, and incubated for 10 min in 3% hydrogen peroxide to quench the endogenous peroxidase activity. For antigen retrieval, the slides were heated in citrate buffer. Then, the slides were incubated overnight at 4 °C with primary antibody. Nuclei were counterstained with hematoxylin. Lastly, DAB chromogen was detected using a commercial IHC staining kit (ZSGB-BIO, PV-6000). Anti-RORγ antibody (Abcam, #ab207082, 1:3000) was used as the primary antibody.
Statistical Analysis
All statistical analyses were conducted using SPSS software (IBM, Armonk, NY, version 26.0). The significance of differences between two groups was analyzed using Student’s t-test, except for gender difference, which was analyzed using the chi-square test. To compare multiple groups, differences were analyzed using one-way ANOVA. Correlations between clinical variables and serum IL-22 levels were estimated with Spearman correlation and partial correlation analysis. Binary logistic regression analysis was conducted to explore the association between serum IL-22 levels and the risk of T2DM. Receiver operating characteristic (ROC) curve analysis was performed to evaluate the diagnostic effectiveness of circulating IL-22 levels in T2DM. A p value < 0.05 was defined as statistically significant.
Results
The Serum Level of IL-22 Was Higher in T2DM Participants
To investigate the association between serum IL-22 levels and T2DM, 48 healthy controls and 46 T2DM participants were included in this study. The main clinical characteristics of the enrolled individuals were presented in Table 1. Sex and age were matched between healthy and T2DM groups. There were no significant differences in total cholesterol (TC), aspartate aminotransferase (AST), serum creatinine (Scr), blood urea nitrogen (BUN), or uric acid (UA) between the two groups. Compared to the healthy controls, the T2DM group had higher levels of hemoglobin A1c (HbA1c), FPG, triglyceride (TG), low-density lipoprotein-cholesterol (LDL-C), alanine aminotransferase (ALT) and γ-glutamyl transferase (GGT), while the levels of high-density lipoprotein-cholesterol (HDL-C) were significantly lower in the T2DM group (p < 0.001). As expected, the T2DM group exhibited a significant increase in serum IL-22 levels (69.18±32.83 vs 127.16±75.35, p < 0.001, Table 1 and Figure 1A).
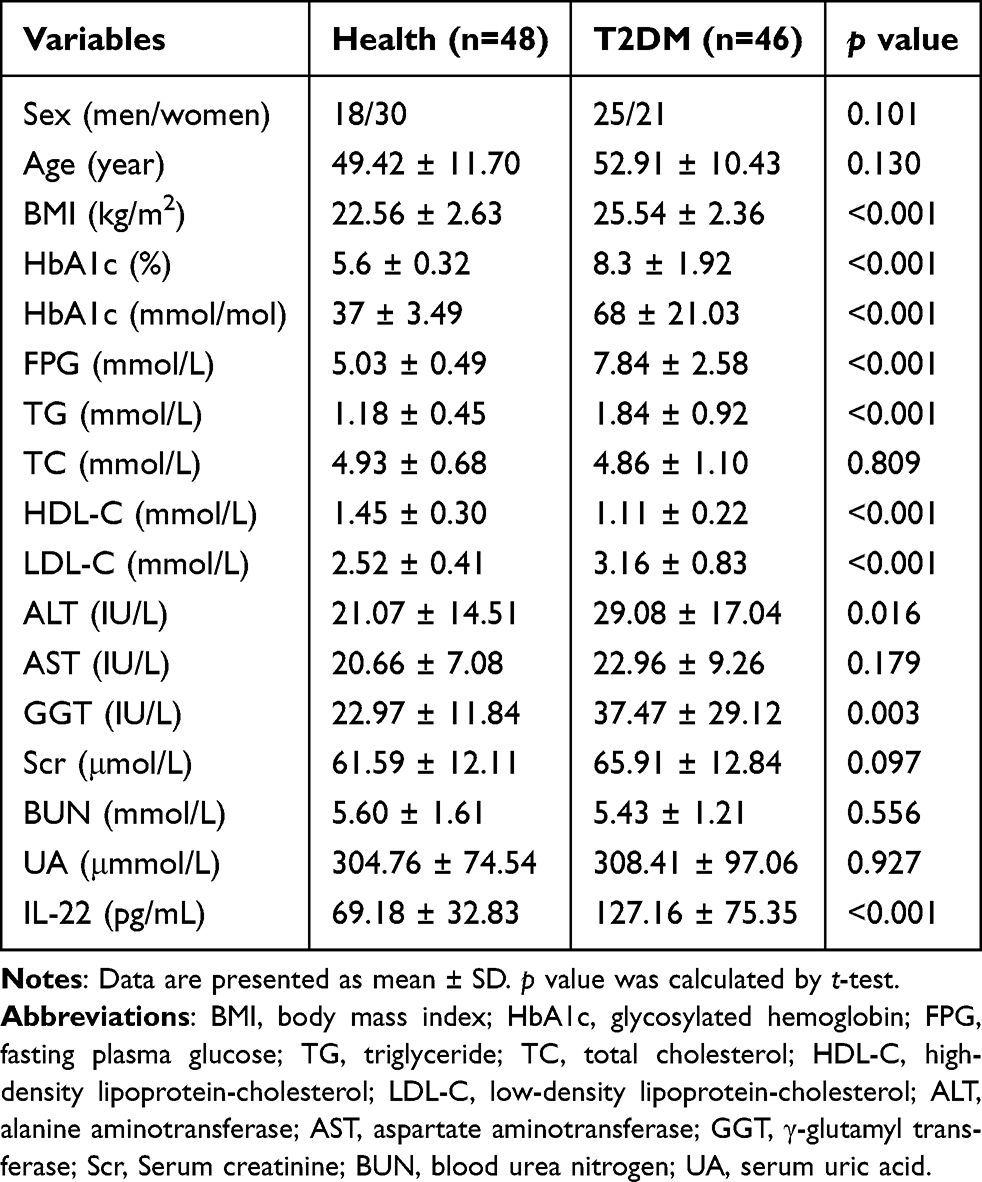 |
Table 1 Clinical Characteristics of Enrolled Healthy and T2DM Participants |
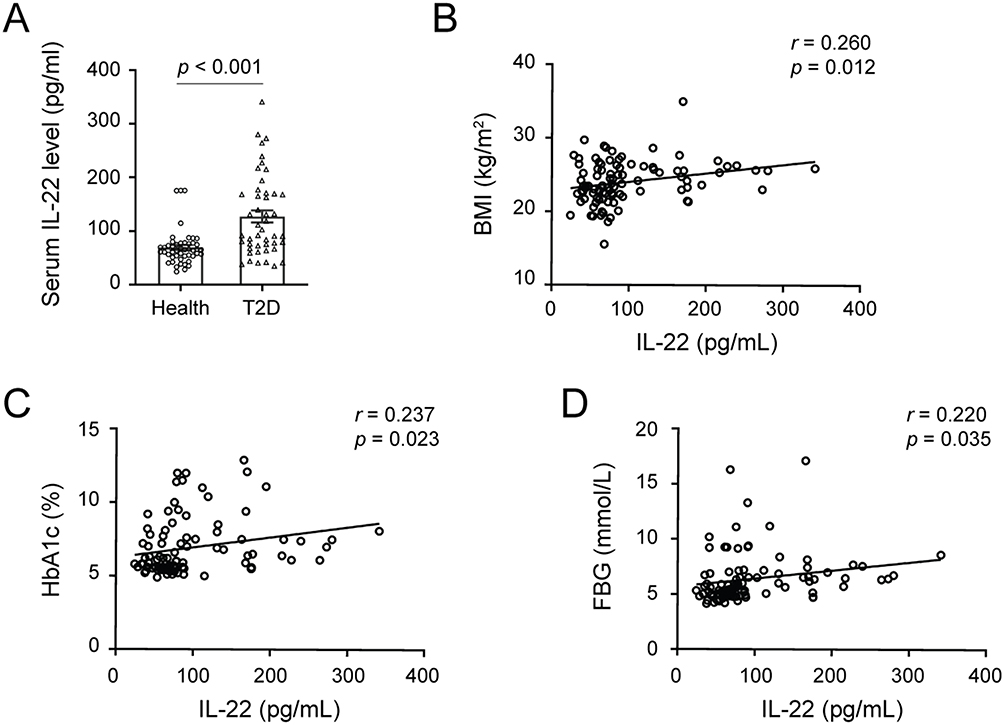 |
Figure 1 The serum IL-22 levels were increased in T2DM participants. (A) The levels of IL-22 in the serum of healthy controls and newly diagnosed T2DM participants. Data are presented as means ± S.E.M. p value was calculated by t test. (B-D). The correlations between serum IL-22 and BMI (B), HbA1c (C), and FBG (D). Data were analyzed by partial correlation test (sex and age adjusted), n=94. |
In addition, we analyzed the correlation between IL-22 levels and metabolic parameters. As shown in Table 2, serum IL-22 concentrations were positively correlated with body mass index (BMI), HbA1c, FPG, LDL-C and ALT, and negatively correlated with HDL-C (Table 2). After adjusting for sex and age, the negative correlation between IL-22 levels and HDL-C and the positive correlations between IL-22 and BMI, HbA1c and FPG remained significant (p < 0.05, Table 2 and Figure 1B–D).
 |
Table 2 Correlations Between Clinical Characteristics and Serum IL-22 Levels in the Participants |
Serum IL-22 Level Was Positively Associated With T2DM Risk
To further confirm the association between serum IL-22 levels and the risk of T2DM, the enrolled participants were divided into four groups according to the quartile of IL-22 levels (quartile 1, <58.00; quartile 2, 58.00–75.04; quartile 3, 75.04–121.85; quartile 4, >121.85pg/mL). Univariate regression analysis showed that serum IL-22 levels and several metabolic characteristics, including BMI, TG, HDL-C, LDL-C, ALT, and GGT levels, were significantly associated with T2DM risk (Table 3). After adjusting for these characteristics, the results of the multivariate analysis showed that serum IL-22 level was a significant independent risk factor for T2DM (OR = 2.37, 95% CI = 1.27–4.42, p = 0.007).
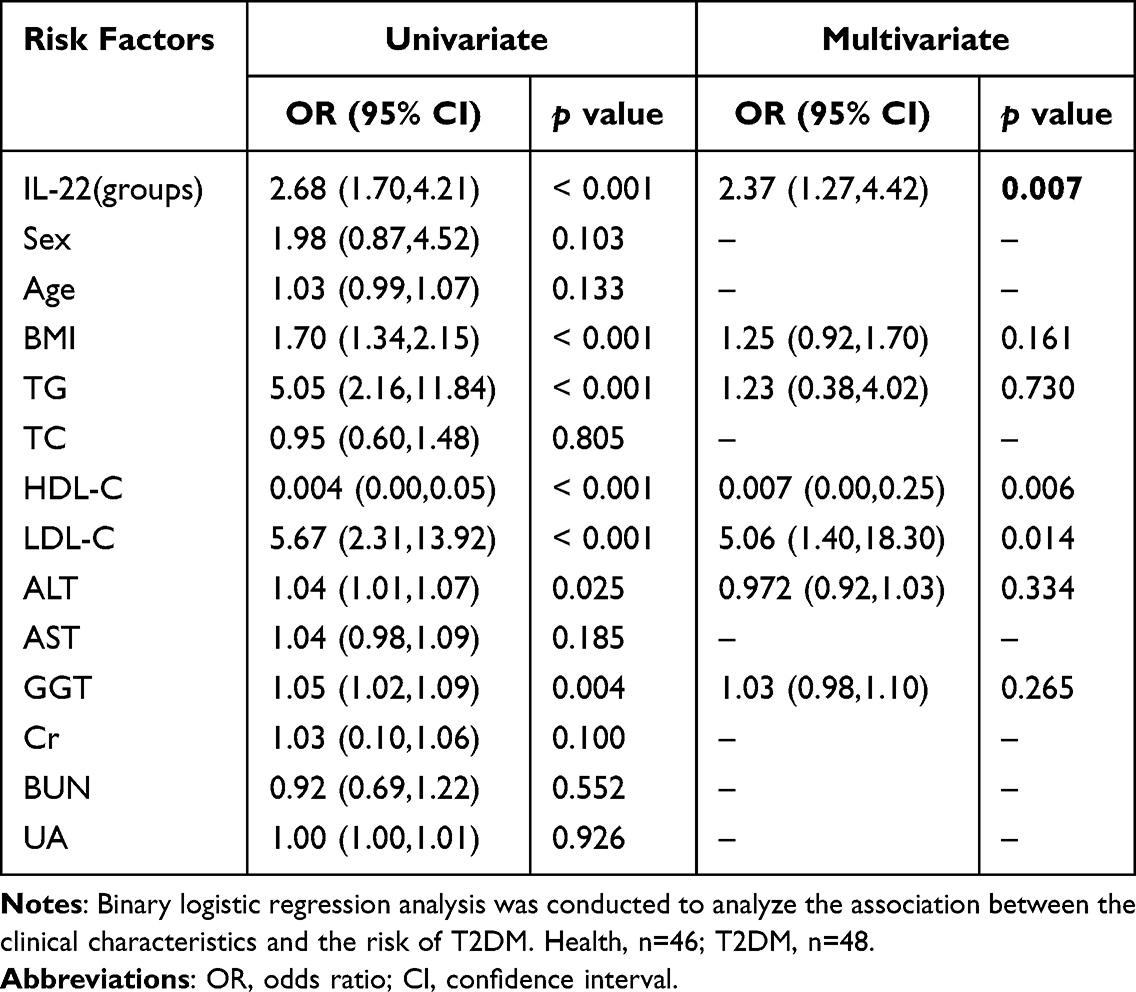 |
Table 3 The Association Between Serum IL-22 Levels and T2DM Risk |
Additionally, the ability of circulating IL-22 to predict T2DM was tested by ROC curve and AUC calculation. As shown in Supplementary Figure 1, circulating IL-22 levels exhibited a relatively good ability to differentiate T2DM from healthy individuals (AUC (95% CI) = 0.767 (0.667–0.866), p < 0.001). These results suggest that circulating IL-22 might be a potential diagnostic biomarker of T2DM.
Sitagliptin Treatment Decreased the Level of IL-22 and Improves Intestinal Inflammation
Next, to further investigate whether the hypoglycemic agent DPP-4i could change IL-22 levels in the clinical setting, we measured serum IL-22 concentrations in another group of newly diagnosed T2DM participants who either received sitagliptin or did not receive any antidiabetic agents. This subset of individuals was enrolled in our previous study. The detection showed that sitagliptin monotherapy induced a decreasing trend in IL-22 levels, although there was no statistically significant difference (130.1 vs 103.1, p = 0.070, Figure 2A). Consistently, serum levels of IL-22 were higher in HFD mice than in ND mice. After sitagliptin treatment, IL-22 levels in the serum of HFD mice (HFD-Sit) significantly decreased (Figure 2B). Moreover, the expression level of IL-22 in the colon tissues of HFD mice was also higher than that in ND mice, and was distinctly decreased by sitagliptin (Figure 2C). These results suggest that IL-22 levels were associated with the hypoglycemic effect of DPP-4i sitagliptin.
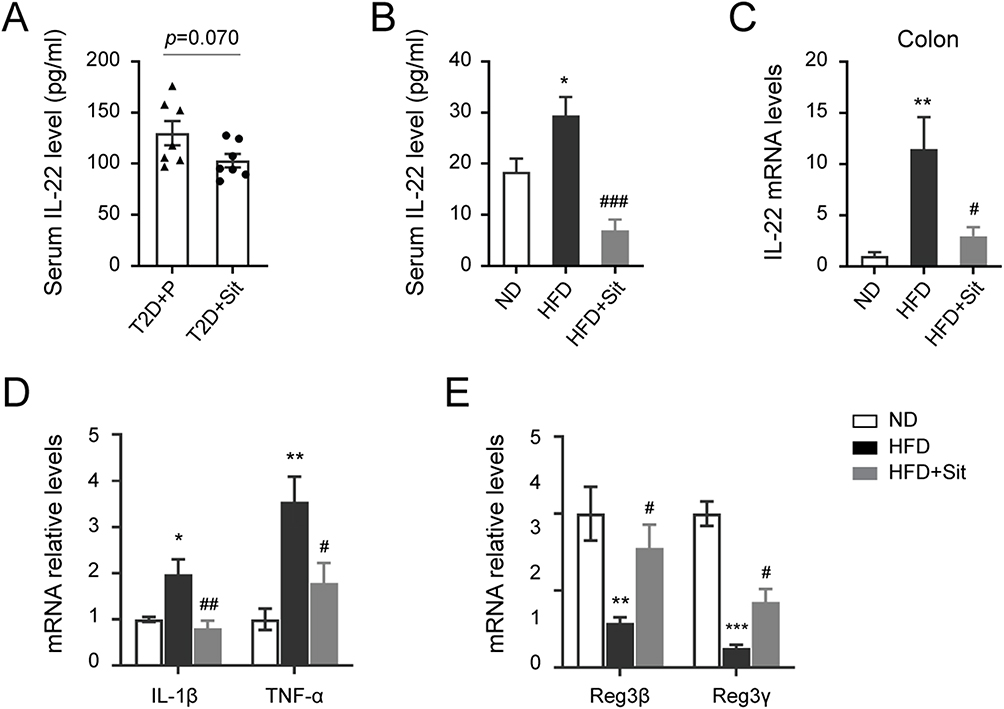 |
Figure 2 The levels of IL-22 were decreased in T2DM participants and HFD mice after sitagliptin treatment. (A) The serum levels of IL-22 in T2DM participants administrated with or without sitagliptin (Sit). n=7. (B) The serum levels of IL-22 in ND mice, and HFD mice treated with or without sitagliptin. (C-E) The mRNA expression levels of IL-22 (C), inflammatory factors (D), and antimicrobial peptides (E) in the colon tissues of ND, HFD and HFD+Sit mice. All data in the bar blot are expressed as the mean ± S.E.M. B-E, n=5 mice per group, *p < 0.05, **p < 0.01 and ***p < 0.001 versus the ND group, #p < 0.05, ##p < 0.01 and ###p < 0.001 versus the HFD group. |
Meanwhile, the expression levels of pro-inflammatory factors (IL-1β and TNF-α) were significantly reduced, and the expression levels of antimicrobial peptides (Reg3β and Reg3γ) were obviously increased in the colon tissues of sitagliptin-treated HFD mice (Figure 2D and E), indicating that sitagliptin treatment attenuated HFD-induced intestinal inflammation. In addition, compared to ND mice, H&E staining of the colon tissue sections of HFD mice revealed detached mucosal epithelial cells and atrophied crypts (Figure 3A). The number of lymphoid nodules in the lamina propria of the colon was increased, some of which were fused in the HFD group. Increased inflammatory cell infiltration in the lamina propria and submucosa was observed in the colon tissues of HFD mice. In contrast, sitagliptin treatment attenuated the inflammatory status induced by HFD. Moreover, IHC staining for RORγt (a marker of Th17 cells)21 was performed to assess the number of Th17 cells. As shown in Figure 3B, the number of RORγt-positive cells in the colon tissues of HFD mice was higher than that in ND mice, however, the number of specific cells decreased in sitagliptin (Sit) treated mice (HFD+Sit), indicating that sitagliptin may reduce IL-22 levels by decreasing the number of Th17 cells in the colon tissues of HFD mice. Taken together, these results suggest that IL-22 might be associated with improvement in intestinal inflammation mediated by the hypoglycemic agent sitagliptin.
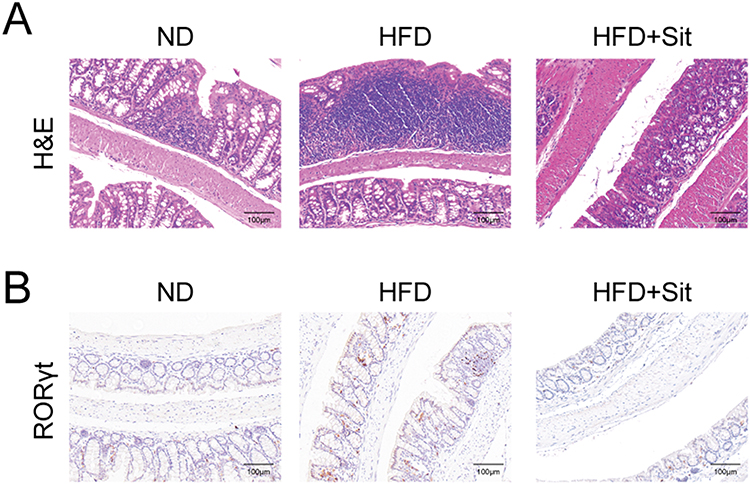 |
Figure 3 RORγT positive cells were decreased in the colon of HFD mice after sitagliptin treatment. (A) Representative images of H&E staining of colon tissue sections. (B) IHC staining of RORγt in the colon tissues of ND, HFD and HFD+Sit mice. Scale bar, 100μm. |
Discussion
Accumulating evidence supports that chronic inflammation contributes to the onset and progression of diabetes.22,23 In this study, we observed that circulating IL-22 levels were significantly increased in newly diagnosed T2DM participants. Moreover, the regression analysis indicated that serum level of IL-22 was an independent risk factor for T2DM. Additionally, ROC analysis suggested that circulating IL-22 levels exhibited a relatively good ability to differentiate T2DM patients from healthy individuals. These results suggested that circulating IL-22 might be a potential diagnostic biomarker for T2DM.
Th17/IL-22 axis has been implicated in the pathogenesis of intestinal inflammatory responses and metabolic diseases.24,25 It has been reported that the increase of Th17 cells in peripheral blood and adipose tissues is related to BMI, serum HDL levels and insulin resistance.8,26,27 Th17 cells that secrete IL-22 have been identified to be involved in the regulation of insulin resistance, as well as glucose metabolism by affecting the inflammatory response.10 Additionally, high serum levels of IL‑22 were positively associated with several cardiometabolic risk factors.28 However, a mouse model of obesity revealed the beneficial effects of IL-22 treatment on inflammation and insulin sensitivity.29 Several clinical studies have found that IL-22 levels in peripheral blood might be associated with diabetes, but the conclusions are inconsistent.30,31 These contradictory results might be attributed to differences in the course of diabetes, clinical treatment, sample numbers of enrolled participants, and so on. In this study, we recruited participants with newly diagnosed T2DM participants, and excluded the influence of different treatments. Nevertheless, prospective studies and fundamental research are needed to determine the role of IL-22 in the pathogenesis of T2DM.
In a previous study, we demonstrated that treatment with the hypoglycemic agent DPP-4i improved glucose homeostasis, accompanied by alterations in the composition of the gut microbiota and metabolites in HFD mice.13 HFD and gut microbiota, as well as their interactions, could induce chronic low-grade inflammation in the intestine.32,33 Dysbiosis of gut microbiota induced an increase in the levels of endotoxins and pathogens in the intestine. These toxic substances promote intestinal macrophages to release pro-inflammatory factors such as TNF-α and histamine, resulting in an inflammatory response. Chronic inflammation exacerbated intestinal permeability, allowing bacterial LPS to pass through the intestinal barrier and affecting the secretion spectrum of intestinal hormones.34,35 These changes are crucial to the occurrence and development of obesity and diabetes. On the other hand, Lactobacillus produced indole-3-lactic acid (ILA) was demonstrated to protect against intestinal inflammation and promoted the expression of key bacterial enzymes implicated in tryptophan metabolism, leading to the synthesis of other indole derivatives, including IAA and indole-3-propionic acid (IPA).36 Indole, acting as a ligand of aryl hydrocarbon receptor (AhR), has been reported to inhibit inflammation to improve hepatic steatosis and insulin resistance.37,38 Our previous metabolomic data showed that the relative content of IAA was significantly increased in the feces of sitagliptin-treated HFD mice.13 IAA has been reported to reduce the proportion of Th17 cells,15 and inhibit LPS-stimulated production of pro-inflammatory cytokines in macrophages.39 That is to say, gut microbiota contributes to inflammation and metabolic dysfunction might be associated with the bacterial metabolites or produced enzymes. Additionally, in the current study, sitagliptin treatment decreased the number of RORγt-positive cells, and reduced IL-22 levels in the colon tissues of HFD mice. These results suggest that sitagliptin might improve inflammation status by increasing IAA to reduce the production of IL-22, which might be an additional benefit of DPP-4i like sitagliptin, suggesting a broader therapeutic utility. Further fundamental studies are required to clarify this issue.
While the current study has some limitations that need to be pointed out. First, our study used a cross-sectional design, so the causality between IL-22 levels and T2DM could not be clarified. Second, due to the small sample size of the T2DM group that received sitagliptin monotherapy, the correlation between IL-22 levels and clinical treatment with sitagliptin could not be determined, which highlights the need for studies with larger sample sizes or longitudinal studies to confirm our findings in the future. On the other hand, mouse experiments were performed and observed sitagliptin treatment reduced IL-22 levels in the serum and colon tissues, which supports the findings of clinical trials, to some extent. While, further molecular experiments should be performed to explore the role and molecular mechanism of DPP-4i/IAA/IL-22 in regulating chronic inflammation.
In conclusion, our study demonstrated that serum levels of IL-22 were significantly increased in newly diagnosed T2DM participants when compared to healthy participants and were positively correlated with the risk of T2DM. Moreover, DPP-4i sitagliptin treatment obviously decreased IL-22 levels accompanied by improving intestinal inflammation. Thus, circulating IL-22 may be a biomarker for T2DM diagnosis and a potential therapeutic target for T2DM.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Ethics Approval and Consent to Participate
The study was approved by the Ethics Committee of the Second Affiliated Hospital of Army Medical University and registered online (Clinical trial registry number ChiCTR-ROC-17010719 and ChiCTR-OPC-17010757). Informed written consent was obtained from all the participants, and the research was conducted in accordance with the principles outlined in the Declaration of Helsinki.
Consent for Publication
Consent for publication was obtained from all study participants.
Author Contributions
All authors contributed to data acquisition, data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 82230025, No. 82370815 and No. 81925007), the “Talent Project” of Army Medical University (No. 2019R055), the Special Program for Basic Research Frontier of Military Medicine of the Second Affiliated Hospital of Army Medical University (No. 2019YQYLY002), and the Talent Project for Young Doctor of the Second Affiliated Hospital of Army Medical University (No. 2022YQB057).
Disclosure
All authors declare no conflicts of interest.
References
1. IDF diabetes atlas reports. 2021, Internatiional Diabetes Federation.
2. Lontchi-Yimagou E, Sobngwi E, Matsha TE, et al. Diabetes mellitus and inflammation. Curr Diab Rep. 2013;13(3):435–444. doi:10.1007/s11892-013-0375-y
3. Gutiérrez-Salmerón M, Lucena SR, Chocarro-Calvo A, et al. Remodelling of colorectal cancer cell signalling by microbiota and immunity in diabetes. Endocr Relat Cancer. 2021;28(6):R173–r190. doi:10.1530/ERC-20-0315
4. Scheithauer TPM, Rampanelli E, Nieuwdorp M, et al. Gut microbiota as a trigger for metabolic inflammation in obesity and type 2 diabetes. Front Immunol. 2020;11:571731. doi:10.3389/fimmu.2020.571731
5. Sittipo P, Lobionda S, Lee YK, et al. Intestinal microbiota and the immune system in metabolic diseases. J Microbiol. 2018;56(3):154–162. doi:10.1007/s12275-018-7548-y
6. Candelli M, Franza L. Interaction between lipopolysaccharide and gut microbiota in inflammatory bowel diseases. Int J Mol Sci. 2021;22(12).
7. Zi C, Wang D, Gao Y, et al. The role of Th17 cells in endocrine organs: involvement of the gut, adipose tissue, liver and bone. Front Immunol. 2022;13:1104943. doi:10.3389/fimmu.2022.1104943
8. Jagannathan-Bogdan M, McDonnell ME, Shin H, et al. Elevated proinflammatory cytokine production by a skewed T cell compartment requires monocytes and promotes inflammation in type 2 diabetes. J Immunol. 2011;186(2):1162–1172. doi:10.4049/jimmunol.1002615
9. Dalmas E, Venteclef N, Caer C, et al. T cell-derived IL-22 amplifies IL-1β-driven inflammation in human adipose tissue: relevance to obesity and type 2 diabetes. Diabetes. 2014;63(6):1966–1977. doi:10.2337/db13-1511
10. Fabbrini E, Cella M, Mccartney SA, et al. Association between specific adipose tissue CD4+ T-cell populations and insulin resistance in obese individuals. Gastroenterology. 2013;145(2):
11. Zhang P, Liu J, Lee A, et al. IL-22 resolves MASLD via enterocyte STAT3 restoration of diet-perturbed intestinal homeostasis. Cell Metab. 2024;36(10):2341–2354e6. doi:10.1016/j.cmet.2024.08.012
12. Sajiir H, Keshvari S, Wong KY, et al. Liver and pancreatic-targeted interleukin-22 as a therapeutic for metabolic dysfunction-associated steatohepatitis. Nat Commun. 2024;15(1):4528. doi:10.1038/s41467-024-48317-x
13. Liao X, Song L, Zeng B, et al. Alteration of gut microbiota induced by DPP-4i treatment improves glucose homeostasis. EBioMedicine. 2019;44:665–674. doi:10.1016/j.ebiom.2019.03.057
14. Liao X, Liu B, Qu H, et al. A high level of circulating valine is a biomarker for type 2 diabetes and associated with the hypoglycemic effect of sitagliptin. Mediators Inflamm. 2019;2019:8247019. doi:10.1155/2019/8247019
15. Shen J, Yang L, You K, et al. Indole-3-acetic acid alters intestinal microbiota and alleviates ankylosing spondylitis in mice. Front Immunol. 2022;13:762580. doi:10.3389/fimmu.2022.762580
16. Pinheiro MM, Stoppa CL, Valduga CJ, et al. Sitagliptin inhibit human lymphocytes proliferation and Th1/Th17 differentiation in vitro. Eur J Pharm Sci. 2017;100:17–24. doi:10.1016/j.ejps.2016.12.040
17. Aso Y, Fukushima M, Sagara M, et al. Sitagliptin, a DPP-4 inhibitor, alters the subsets of circulating CD4+ T cells in patients with type 2 diabetes. Diabet Res Clin Pract. 2015;110(3):250–256. doi:10.1016/j.diabres.2015.10.012
18. Alberti KG, Zimmet PZ. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet Med. 1998;15(7):539–553. doi:10.1002/(SICI)1096-9136(199807)15:7<539::AID-DIA668>3.0.CO;2-S
19. Li X, Liao M, Shen R, et al. Plasma asprosin levels are associated with glucose metabolism, lipid, and sex hormone profiles in females with metabolic-related diseases. Mediators Inflamm. 2018;2018:7375294. doi:10.1155/2018/7375294
20. Bialkowska AB, Ghaleb AM, Nandan MO, et al. Improved Swiss-rolling technique for intestinal tissue preparation for immunohistochemical and immunofluorescent analyses. J Vis Exp. 2016; 113. doi:10.3791/54161
21. Ivanov II, McKenzie BS, Zhou L, et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126(6):1121–1133. doi:10.1016/j.cell.2006.07.035
22. Rohm TV, Meier DT, Olefsky JM, et al. Inflammation in obesity, diabetes, and related disorders. Immunity. 2022;55(1):31–55. doi:10.1016/j.immuni.2021.12.013
23. Luck H, Tsai S, Chung J, et al. Regulation of obesity-related insulin resistance with gut anti-inflammatory agents. Cell Metab. 2015;21(4):527–542. doi:10.1016/j.cmet.2015.03.001
24. Zhang S, Gang X, Yang S, et al. The alterations in and the role of the Th17/Treg balance in metabolic diseases. Front Immunol. 2021;12:678355. doi:10.3389/fimmu.2021.678355
25. Ip B, Cilfone NA, Belkina AC, et al. Th17 cytokines differentiate obesity from obesity-associated type 2 diabetes and promote TNF α production. Obesity. 2016;24(1):102–112. doi:10.1002/oby.21243
26. Zeng C, Shi X, Zhang B, et al. The imbalance of Th17/Th1/Tregs in patients with type 2 diabetes: relationship with metabolic factors and complications. J Mol Med. 2012;90(2):175–186. doi:10.1007/s00109-011-0816-5
27. Gomes AL, Teijeiro A, Burén S, et al. Metabolic inflammation-associated IL-17A causes non-alcoholic steatohepatitis and hepatocellular carcinoma. Cancer Cell. 2016;30(1):161–175. doi:10.1016/j.ccell.2016.05.020
28. Herder C, Kannenberg JM, Carstensen-Kirberg M, et al. Serum levels of interleukin-22, cardiometabolic risk factors and incident type 2 diabetes: KORA F4/FF4 study. Cardiovasc Diabetol. 2017;16(1):17. doi:10.1186/s12933-017-0498-6
29. Wang X, Ota N, Manzanillo P, et al. Interleukin-22 alleviates metabolic disorders and restores mucosal immunity in diabetes. Nature. 2014;514(7521):237–241. doi:10.1038/nature13564
30. Shen J, Fang Y, Zhu H, et al. Plasma interleukin-22 levels are associated with prediabetes and type 2 diabetes in the Han Chinese population. J Diabetes Investig. 2018;9(1):33–38. doi:10.1111/jdi.12640
31. Gong F, Wu J, Zhou P, et al. Interleukin-22 might act as a double-edged sword in Type 2 diabetes and coronary artery disease. Mediators Inflamm. 2016;2016:8254797. doi:10.1155/2016/8254797
32. Li X, Huang G, Zhang Y, et al. Succinate signaling attenuates high-fat diet-induced metabolic disturbance and intestinal barrier dysfunction. Pharmacol Res. 2023;194:106865. doi:10.1016/j.phrs.2023.106865
33. Kang Y, Kang X, Yang H, et al. Lactobacillus acidophilus ameliorates obesity in mice through modulation of gut microbiota dysbiosis and intestinal permeability. Pharmacol Res. 2022;175:106020. doi:10.1016/j.phrs.2021.106020
34. Yang G, Wei J, Liu P, et al. Role of the gut microbiota in type 2 diabetes and related diseases. Metabolism. 2021;117:154712. doi:10.1016/j.metabol.2021.154712
35. Gao D, Jiao J, Wang Z, et al. The roles of cell-cell and organ-organ crosstalk in the type 2 diabetes mellitus associated inflammatory microenvironment. Cytokine Growth Factor Rev. 2022;66:15–25. doi:10.1016/j.cytogfr.2022.04.002
36. Wang G, Fan Y, Zhang G, et al. Microbiota-derived indoles alleviate intestinal inflammation and modulate microbiome by microbial cross-feeding. Microbiome. 2024;12(1):59. doi:10.1186/s40168-024-01750-y
37. Ma L, Li H, Hu J, et al. Indole alleviates diet-induced hepatic steatosis and inflammation in a manner involving myeloid cell 6-Phosphofructo-2-Kinase/Fructose-2,6-Biphosphatase 3. Hepatology. 2020;72(4):1191–1203. doi:10.1002/hep.31115
38. Krishnan S, Ding Y, Saedi N, et al. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2018;23(4):1099–1111. doi:10.1016/j.celrep.2018.03.109
39. Krishnan S, Ding Y, Saeidi N, et al. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2019;28(12):3285. doi:10.1016/j.celrep.2019.08.080
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.