Back to Journals » Cancer Management and Research » Volume 11
circPCNX and Pecanex Promote Hepatocellular Carcinoma Cell Viability by Inhibiting miR-506
Authors Sun P, Fan X, Hu X, Fu X, Wei Q, Zang Y
Received 30 September 2019
Accepted for publication 18 December 2019
Published 3 January 2020 Volume 2019:11 Pages 10957—10967
DOI https://doi.org/10.2147/CMAR.S232940
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Chien-Feng Li
Peng Sun,1,* Xinyi Fan,2,* Xiao Hu,3 Xiaoyue Fu,1 Qian Wei,3 Yunjin Zang1
1Department of Liver Transplantation, The Affiliated Hospital of Qingdao University, Qingdao 266003, People’s Republic of China; 2Intensive Care Unit, the Affiliated Hospital of Qingdao University, Qingdao 266003, People’s Republic of China; 3Department of Hepatobiliary and Pancreatic Surgery, The Affiliated Hospital of Qingdao University, Qingdao 266003, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yunjin Zang
Department of Liver Transplantation, The Affiliated Hospital of Qingdao University, 69 Haier Road, Qingdao 266003, People’s Republic of China
Tel/Fax +86 532 82915906
Email [email protected]
Introduction: Hepatocellular carcinoma (HCC) is one of the most common malignant cancers, while the molecular mechanism is not clear. Circular RNAs (circRNAs) are a class of naturally occurring endogenous noncoding RNAs that prove to play important roles in the occurrence, development and prognosis of HCC. In this study, we focused on an abnormally expressed circular RNA-circPCNX in HCC and study the function of circPCNX and Pecanex (PCNX) in HCC.
Methods: Circular RNA sequencing was used to find the abnormally expressed circRNAs and qRT-PCR was used to verify it. CCK8 assay, colony formation assay and cell apoptosis assay were used to study biological functions, and Luciferase reporter assay and Western blot analysis were used to study the mechanism.
Results: We observed that circPCNX and Pecanex were significantly upregulated in tumor tissues of patients with HCC and correlated with clinicopathological variables or prognosis of HCC patients. Functional investigations showed circPCNX and Pecanex could promote the viability of HCC cells. Mechanistic investigations suggested that both circPCNX and Pecanex 3ʹUTR could bind to miR-506 and subsequently inhibited the miR-506-induced anticarcinogenic effect in HCC.
Conclusion: Our study revealed the function of circPCNX and Pecanex in promoting HCC progression and acting as biomarkers in the clinical estimate and treatment of HCC.
Keywords: circPCNX, Pecanex, miR-506, hepatocellular carcinoma
Introduction
Hepatocellular carcinoma (HCC) is one of the most common primary malignancies of the liver and a major global health problem.1,2 Surgical treatments, such as liver resection and transplantation, remain the most frequently used curative therapy for patients with early-stage HCC. However, the prognosis after curative resection of HCC has remained unsatisfactory because of a high incidence of postoperative recurrence.3,4 Recently, many studies have explored the molecular mechanism of HCC tumorigenesis, and further investigations of new molecular targets affecting tumor progression are required for both the clinical diagnosis and treatment of patients with HCC.
Circular RNAs (circRNAs) represent a class of naturally occurring, endogenous, noncoding RNAs that have recently been recognized as important regulators of gene expression networks. Unlike linear RNAs that terminate with 5ʹ and 3ʹ tails, circRNAs are characterized by covalently closed-loop structures with neither 5ʹ caps nor 3ʹ polyadenylated tails.5,6 circRNAs are conserved and stable, and numerous circRNAs have been reported to be specifically expressed in a given cell type or developmental stage.7,8 circRNAs contain selectively conserved microRNA (miRNA) target sites and serve as efficient miRNA sponges, interacting with miRNAs to regulate gene expression.9 Therefore, circRNAs associate with cancer-related miRNAs and influence the progression of cancers by regulating the function of miRNAs. The circRNA-miRNA axes in cancer-related pathways have been a hot topic in recent studies. Abnormal circRNA expression has been observed in many human cancers and might play significant roles in the pathogenesis and diagnosis of these cancers.10–12
In our study, we analyzed the expression profile of circRNAs in HCC tissues and identified that hsa_circRNA_0102533, which we termed circPCNX because it is derived from the PCNX gene, was significantly up-regulated in HCC tissues. A recent study revealed a role for PCNX in non-small cell lung cancer (NSCLC), including promoting cell growth and proliferation, inducing cell cycle progression and repressing cell apoptosis.13 PCNX may act as a competitive endogenous RNA (ceRNA) of S-phase kinase-associated protein 2 (Skp2) to positively regulate Skp2 expression, and PCNX might exert its oncogenic effect on Skp2 by inducing Akt phosphorylation in a miRNA-dependent manner.13 However, no studies have examined PCNX and circPCNX expression in HCC, and the functions of PCNX and circPCNX in HCC remain unclear. In this study, we focused on PCNX and circPCNX to study their biological functions, mechanisms and abilities to serve as biomarkers of HCC.
Materials and Methods
Tissue Specimens from Patients
Seventy pairs of HCC tissues and adjacent nontumor liver tissues (ANLTs) were randomly selected from patients who underwent liver resection at The Affiliated Hospital of Qingdao University from January 2011 to December 2016. The notumor samples were removed from a site located more than 2 cm from the tumor site. The study protocol conformed to the ethical guidelines of the Declaration of Helsinki. All human materials were obtained with written informed consent and the procedure was approved by the Ethics Committee of The Affiliated Hospital of Qingdao University.
Cell Culture
HL-7702, SMMC-7721, HuH-7, Hep3B and HepG2 cells were obtained from the Cell Resource Center of Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences (Shanghai, China) and cultured in DMEM containing 10% fetal bovine serum in a 5% CO2 atmosphere at 37°C.
Quantitative Real-Time PCR (qRT-PCR)
Total RNA was isolated using TRIzol reagent (Life Technologies, USA). The nuclear and cytoplasmic fractions were isolated using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific). Complementary DNAs were synthesized using the PrimeScript RT reagent kit (Takara Bio Inc., China), and RT-PCR was performed using the SYBR Premix Ex Taq (Takara Bio Inc.). The following primers were used: circPCNX, F: 3ʹ-ATCCAGAGTGAGCGATTACAG-5ʹ, R: 3ʹ-ACAAAACCCACAAAGCCAACC-5ʹ; PCNX, F: 5ʹ-GTGCTCAGTGCTAGTGCCTCCT-3ʹ, R: 5ʹGATGCTCTTCTTGCTGCTGGTTCA3ʹ; and β-actin, F: 5ʹ-CAGAAGGAGATTACTGCTCTGGCT-3ʹ, R: 5ʹ-TACTCCTGCTTGCTGATCCACATC-3ʹ.
Circular RNA Sequencing
Total RNA was extracted from five HCC tissue samples and five ANLT samples using Trizol (Invitrogen, Carlsbad, CA, USA). RNAs were digested with RNase R (Epicentre Technologies, Madison, WI, USA) to remove the linear RNAs and enrich the circular configuration. The sequencing analysis was performed using an Illumina HiSeq2000 Platform (Illumina Inc., San Diego, CA, USA). The expression profile was sorted using Cluster 3.0 software (University of Tokyo, Human Genome Center).
Luciferase Reporter Assay
Luciferase reporter vectors carrying the wild-type and mutant binding sites from circPCNX or the 3ʹUTR of PCNX were constructed. The circPCNX sequences containing predicted wild-type miR-506 binding sites were inserted into the region directly downstream of a cytomegalovirus (CMV) promoter-driven firefly luciferase cassette in a pCDNA3.1 vector. The PCNX 3ʹ UTR sequences containing the predicted wild-type miR-506 binding sites were inserted into the region directly downstream of a CMV promoter-driven firefly luciferase cassette in the pCDNA3.1 vector. Mutant reporter plasmids were prepared using a mutagenesis kit (Stratagene, La Jolla, CA, USA). All constructs were verified by sequencing. Cells were seeded into 96-well plates and were co-transfected with a mixture of 30 ng of firefly luciferase reporter, 5 ng of pRL-TK Renilla luciferase reporter, and miRNA mimics or inhibitors. Luciferase activity assay was conducted using the Dual Luciferase Assay System (Promega, Madison, WI, USA). The pRL-TK plasmid (Promega) was used to normalize the transfection efficiency.
Oligonucleotide and Plasmid Transfection
The miR-506 mimics and miR-506 inhibitors were purchased from GenePharma (Shanghai, China). Oligonucleotides were transfected into cells using the Hiperfect transfection reagent (Qiagen), and plasmids were transfected using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA). The circPCNX siRNA sequences were: sense, GAAAUCCAGAGUGAGCGAUUATT; antisense, UAAUCGCUCACUCUGGAUUUCTT. The mature sequence for circPCNX was synthesized and cloned into pCD-ciR vector to construct circPCNX overexpression plasmids. The overexpression of circPCNX was validated by Sanger sequencing.
CCK8 Assay
Cell proliferation was assessed using the Cell Counting Kit-8 assay (Dojindo, Kumamoto, Japan). Cells were seeded into 96-well plates and transfected with the corresponding oligonucleotides or plasmids. Then, 10 μL of the CCK-8 solution was added to each well at 24, 48, and 72 hrs. After a 30-min incubation at 37°C, the absorbance was measured at 450 nM.
Colony Formation Assays
After transfection, cells were counted and 500 cells per well were seeded. Then, the cells were cultured in a 5% CO2 atmosphere at 37°C for 2 weeks of culture. Methanol was used to fix the cells and 1% crystal violet was used to dye the cells.
Cell Apoptosis Assay
After transfection, cells were collected and stained with Annexin V-FITC and 5 μL propidium iodide (PI). The apoptosis rates were then analyzed by flow cytometry.
Western Blot Analysis
Levels of the Snail2 and YAP1 proteins were analyzed using Western blot analyses. Total proteins were extracted, separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane (Millipore, USA). Membranes were incubated with 5% skim milk powder at room temperature for 1 h to block nonspecific binding. The membrane was then incubated with a primary antibody, followed by a horseradish peroxidase-labeled secondary antibody and detected using chemiluminescence. Antibodies used in this assay included a Snail2 antibody (ab106077; Abcam, Cambridge, MA, USA), YAP1 antibody (ab205270; Abcam), and β-actin antibody (ab8227; Abcam) that was used as a protein loading control.
Statistical Analysis
Statistical analyses were performed using the SPSS program (version 18.0; SPSS, Chicago, IL, USA). Data are presented as means±SD Statistical significance was calculated using Student’s t-test, χ2 test, Fisher’s exact test or one-way ANOVA. Pearson’s or Spearman correlation coefficients were calculated for the correlation analyses. The Kaplan-Meier survival analysis (log-rank test) was used to analyze disease-free survival (DFS) and overall survival (OS); the factors with P values<0.05 were included in the Cox regression hazard model. P<0.05 was considered statistically significant.
Results
The Expressions of circPCNX and PCNX are Upregulated in HCC Tumor Tissues and Cell Lines
We analyzed circRNA expression patterns between HCC tissues and paired adjacent nontumor liver tissues (ANLTs) using circRNA sequencing. Increased hsa-circ-0102533 expression was detected in HCC tumor tissues compared to matched nontumor liver tissues, with fold change greater than 3, P < 0.05 (Figure 1A). We then measured hsa-circ-0102533 levels in 70 pairs of HCC tissues and ANLTs using quantitative reverse transcription-PCR (qRT-PCR) to confirm these results, and hsa-circ-0102533 was expressed at significantly higher levels in HCC tissues (Figure 1B and C). Because hsa-circ-0102533 was derived from the PCNX gene locus, we renamed it circPCNX. We then detected PCNX mRNA levels in 70 pairs of HCC tissues and ANLTs using qRT-PCR and observed significantly higher PCNX expression in HCC tissues (Figure 1D and E) that positively correlated with circRCNX expression (Figure 1F). Furthermore, we showed that both circPCNX and PCNX mRNA expressions were increased in HCC cell lines (Figure 1G and H). Thus, we focused on the expression and roles of circPCNX and PCNX in HCC progression in this study.
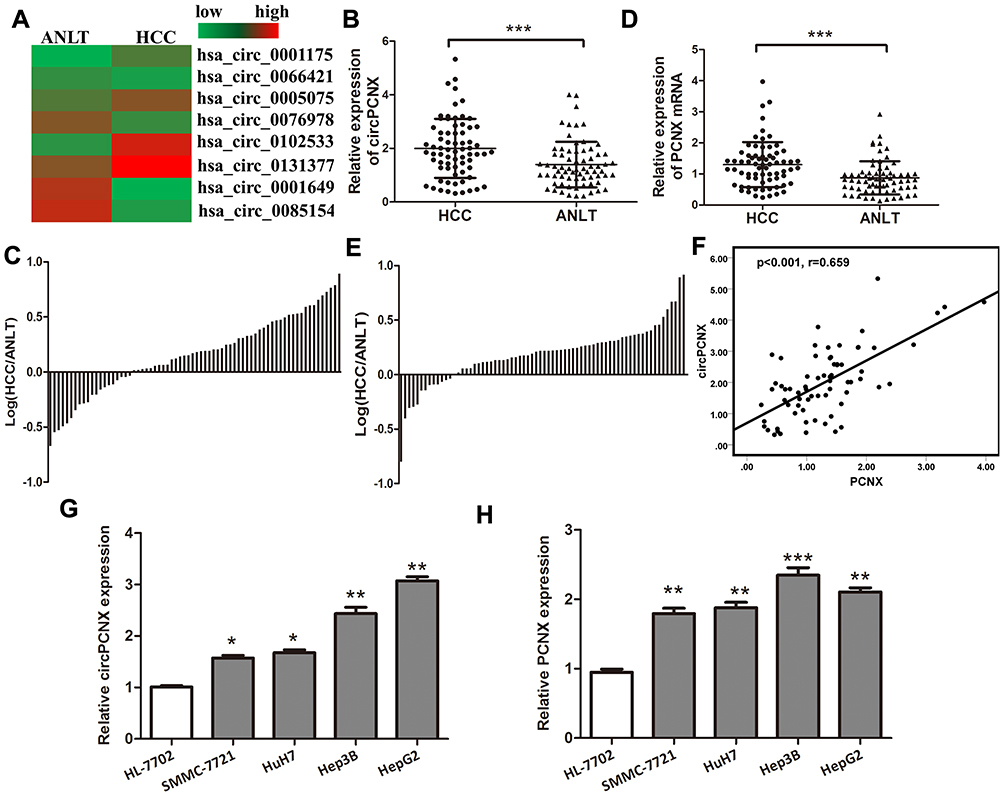 |
Figure 1 The expressions of circPCNX and PCNX are upregulated in HCC tumor tissues and cell lines. (A) Heatmap showing the circRNA expression profiles of HCC tissues and ANLTs. Red and green indicate high and low expression, respectively. (B) Relative expression levels of circPCNX in 70 pairs of HCC tissues and ANLTs were determined using qRT-PCR. The data were analyzed using the delta Ct method and compared using paired Student’s t-tests. (C) Relative circPCNX expression with the ratio of its level in HCC tissues versus ANLTs shown on the logarithmic scale. (D) Relative expression levels of the PCNX mRNA in 70 pairs of HCC tissues and ANLTs. (E) Relative PCNX mRNA levels with the ratio of its level in HCC tissues versus ANLTs shown on the logarithmic scale. (F) Spearman correlation analysis of the relative expression of circPCNX and PCNX. (G and H) Relative expression levels of circPCNX and PCNX mRNA in human normal liver cell line and HCC cell lines. Results are presented as means± SD *P<0.05, **P<0.01, and ***P<0.001. Abbreviation: NS, not significant. |
Correlation Between circPCNX or PCNX Expression and the Clinicopathological Variables or Prognosis of Patients with HCC
First, circPCNX or PCNX expression in HCC tissues was divided into two groups including the high expression group (higher than the median expression in HCC) and the low expression group (lower than the median expression in HCC) to investigate correlations between circPCNX or PCNX expression and the clinicopathological variables or prognosis of patients with HCC. The results revealed a relative increase in circPCNX expression in HCC tissues from patients whose tumor size was >5 cm and TNM stage was II-III, and a relative increase in PCNX expression in HCC tissues from patients whose tumor size was >5 cm (Table 1). Moreover, the Kaplan-Meier survival analysis also showed that patients with high circPCNX levels showed lower disease-free survival and overall survival rates (Figure 2A and B), and patients with high PCNX levels showed lower overall survival but not disease-free survival rates (Figure 2C and D). According to the Cox regression hazard model, PCNX was an independent risk factor for the overall survival rates of patients with HCC (Tables 2 and 3).
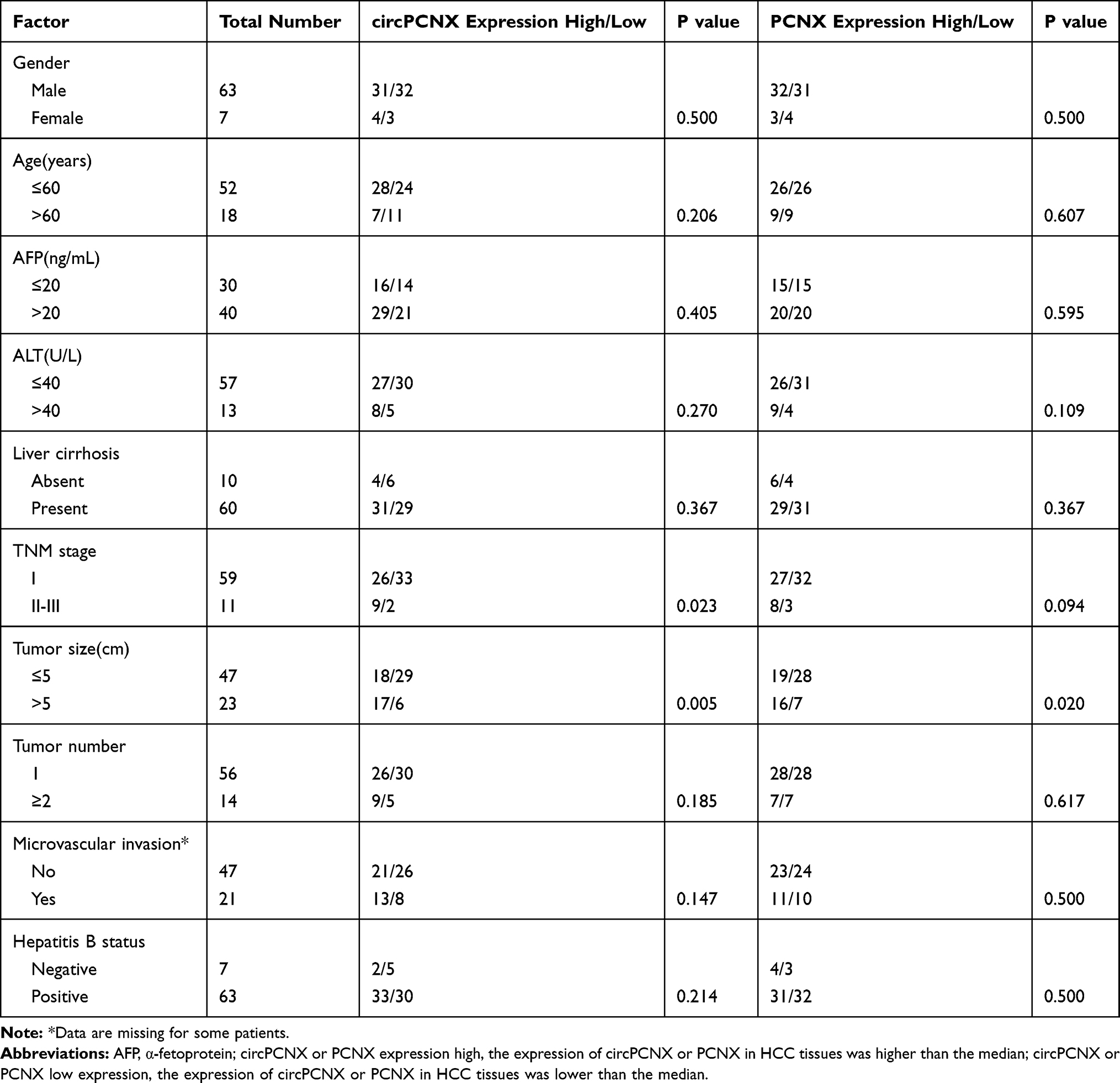 |
Table 1 Correlations Between the Clinicopathological Characteristics and circPCNX and PCNX Expression in HCC Tissues |
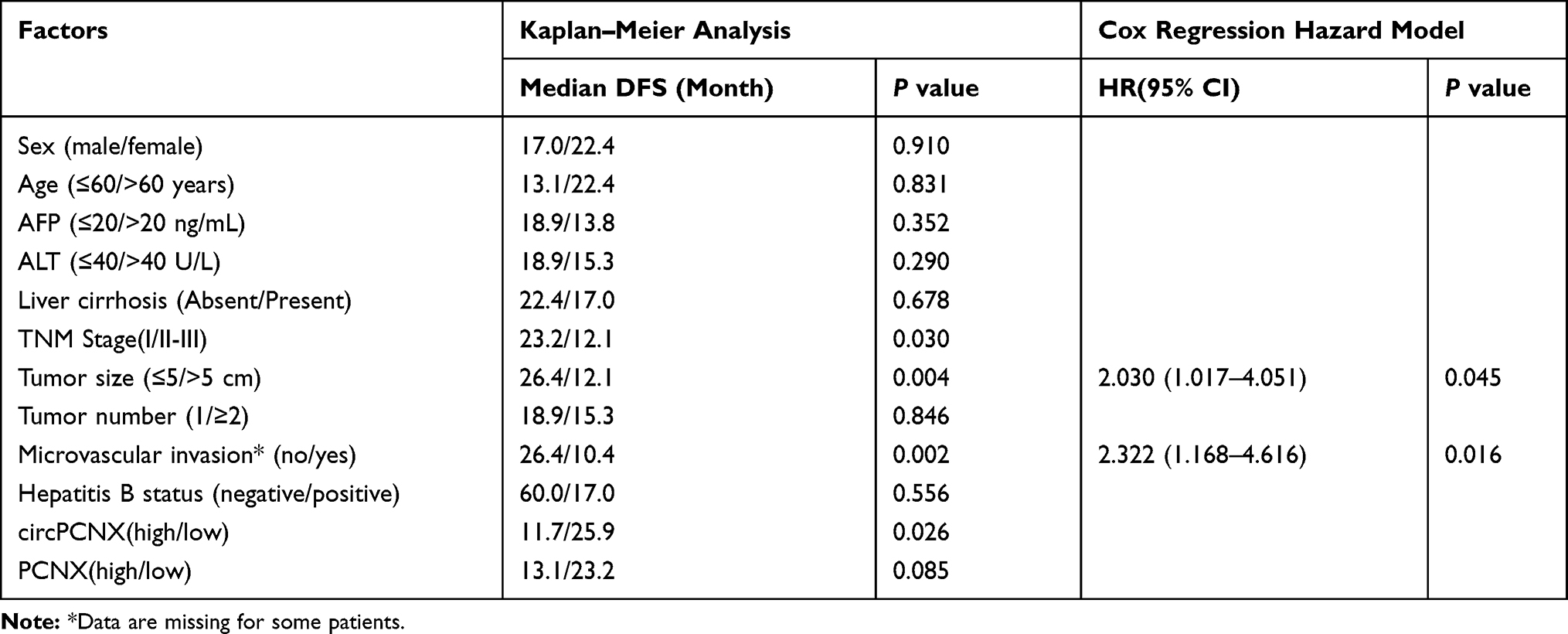 |
Table 2 Factors Influencing the Disease-Free Survival (DFS) of Patients with HCC |
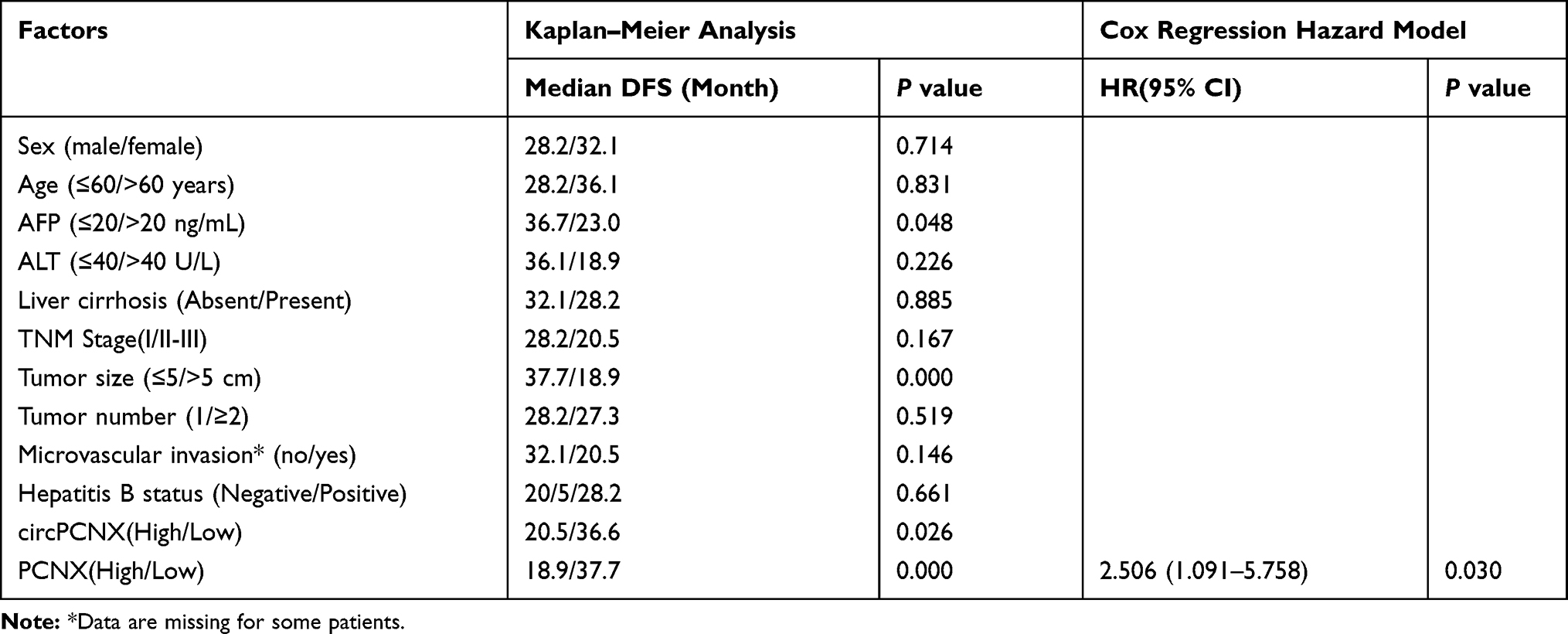 |
Table 3 Factors Influencing the Overall Survival (OS) of Patients with HCC |
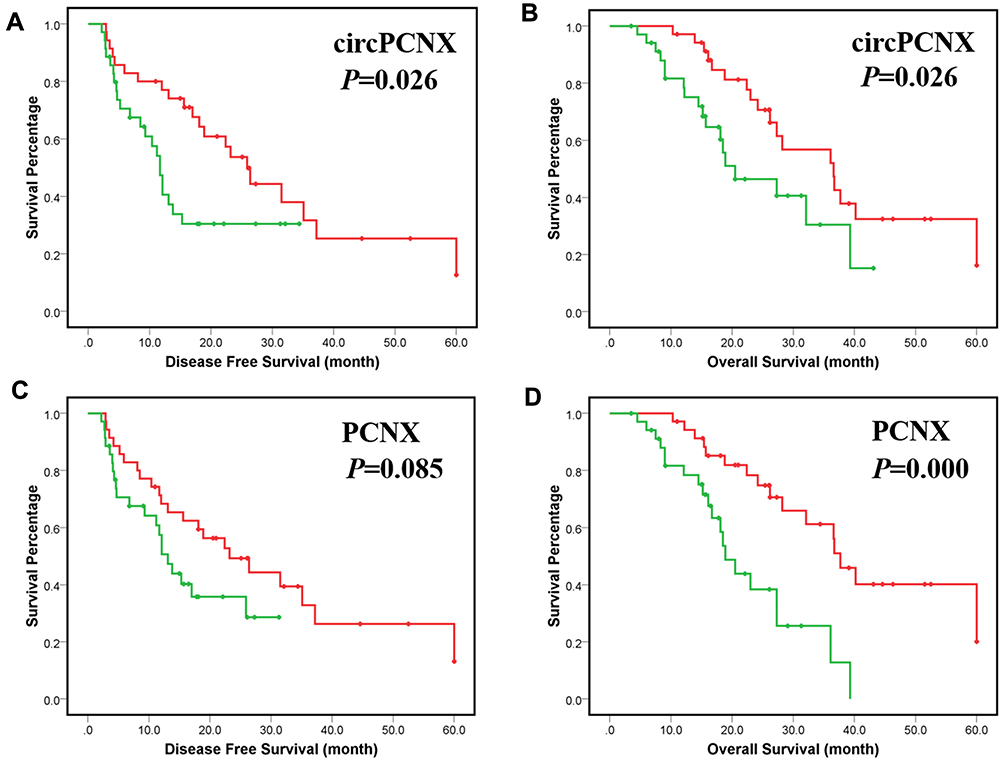 |
Figure 2 Correlations between circPCNX or PCNX expression and the clinicopathological variables or prognosis of patients with HCC. (A) Disease-free survival (DFS) of patients with a high-level circPCNX expression and low level of circPCNX expression. (B) Overall survival (OS) of patients with a high-level circPCNX expression and a low level of circPCNX expression. (C) Disease-free survival of patients with a high level of PCNX expression and a low level of PCNX expression. (D) Overall survival of patients with a high level of PCNX expression and a low level of PCNX expression. Red line, circPCNX or PCNX low expression group; green line, circPCNX or PCNX high expression group; +, censored points. |
circPCNX Serves as a Sponge for miR-506
Because circRNA has been shown to act as a miRNA sponge, we first detected the intracellular enrichment of circPCNX in SMMC-7721 cells using qRT-PCR, and circPCNX was mainly located in the cytoplasm (Figure 3A). Next, we explored whether circPCNX bound to miRNAs. Using a prediction tool based on TargetScan, potential miR-506 binding sites were presented in the circPCNX sequence (Figure 3B). Thus, we performed luciferase reporter assays to determine whether circPCNX functioned as an miR-506 sponge. The luciferase activity was decreased when SMMC-7721 cells were co-transfected with luciferase reporters and miR-506 mimics (Figure 3C) but was increased when SMMC-7721 cells were co-transfected with luciferase reporters and the miR-506 inhibitor (Figure 3D). Then, the target sites of miR-506 in the luciferase reporter were mutated. Co-transfection of the mutated luciferase reporter and miR-506 mimics or miR-506 inhibitor had no significant effect on luciferase activity (Figure 3C and D). Thus, circPCNX functioned as a sponge for miR-506. Because miR-506 was shown to inhibit HCC cell proliferation in many studies, we then detected the expression of the oncogenes proven to be targets of miR-506 in cells overexpressing circPCNX. Snail2 and YAP1, which are targets of miR-506 and promote proliferation, were significantly upregulated by circPCNX (Figure 3E); We also observed that the Snail2 and YAP1 upregulation induced by circPCNX could be rescued by miR-506 (Figure 3E). Therefore, we postulate that circPCNX may promote the expression of miR-506 targeted oncogenes via miR-506. When SMMC-7721 cells were transfected with circPCNX plasmids or siRNAs, a significant change in miR-506 expression was not observed (p>0.05) (Figure 3F and G). When SMMC-7721 cells were transfected with miR-506 mimics or inhibitors, significant changes in cirxPCNX expression were not observed (p>0.05) (Figure 3H and I). Based on these results, miR-506 and circPCNX might not cause the degradation of each other.
 |
Figure 3 circPCNX functions as a sponge for miR-506. (A) The intracellular enrichment of circPCNX in SMMC-7721 cells was detected using qRT-PCR. (B) Predicted miR-506 binding site and corresponding mutant sites in the circPCNX sequence (wt, wild-type; mt, mutant). (C and D) Luciferase activity in SMMC-7721 cells co-transfected with luciferase reporters and miR-506 mimics or miR-506 inhibitors. The luciferase activity of each group was normalized to the value obtained in the cells transfected with NC mimics. (E) Relative Snail2 and YAP1 expression in SMMC-7721 cells transfected with circPCNX expression plasmids and miR-506 mimics, as analyzed using Western blot assays. (F and G) qRT-PCR analysis of miR-506 expression in SMMC-7721 cells transfected with circPCNX expression plasmids or siRNAs. (H and I) qRT-PCR analysis of circPCNX expression in SMMC-7721 cells transfected with miR-506 mimics or inhibitors. Results are presented as means± SD *P<0.05, and **P<0.01.U6, RNU6-1. Abbreviations: NS, not significant; NC, negative control. |
circPCNX Promotes HCC Cell Viability via miR-506
Because circPCNX expression was higher in HepG2 cells than that in SMMC-7721 cells, we overexpressed circPCNX in SMMC-7721 cells using expression plasmids and validated it by Sanger sequencing (Figure 4A and B). We silenced circPCNX expression in HepG2 cells with an siRNA (Figure 4C); the circPCNX siRNA did not affect PCNX expression (Figure 4D). After transfection, we used CCK-8 assays, colony formation assays and cell apoptosis assays to assess proliferation and apoptosis. Overexpression of circPCNX in SMMC-7721 cells promoted cell proliferation and restrained cell apoptosis which could be attenuated by miR-506 mimics (Figure 4E, G, I and J). Accordingly, silencing circPCNX in HepG2 cells inhibited cell proliferation and accelerated cell apoptosis which could be reversed by miR-506 inhibitor (Figure 4F, H, K and L). Based on these data, circPCNX promoted HCC cell viability via miR-506.
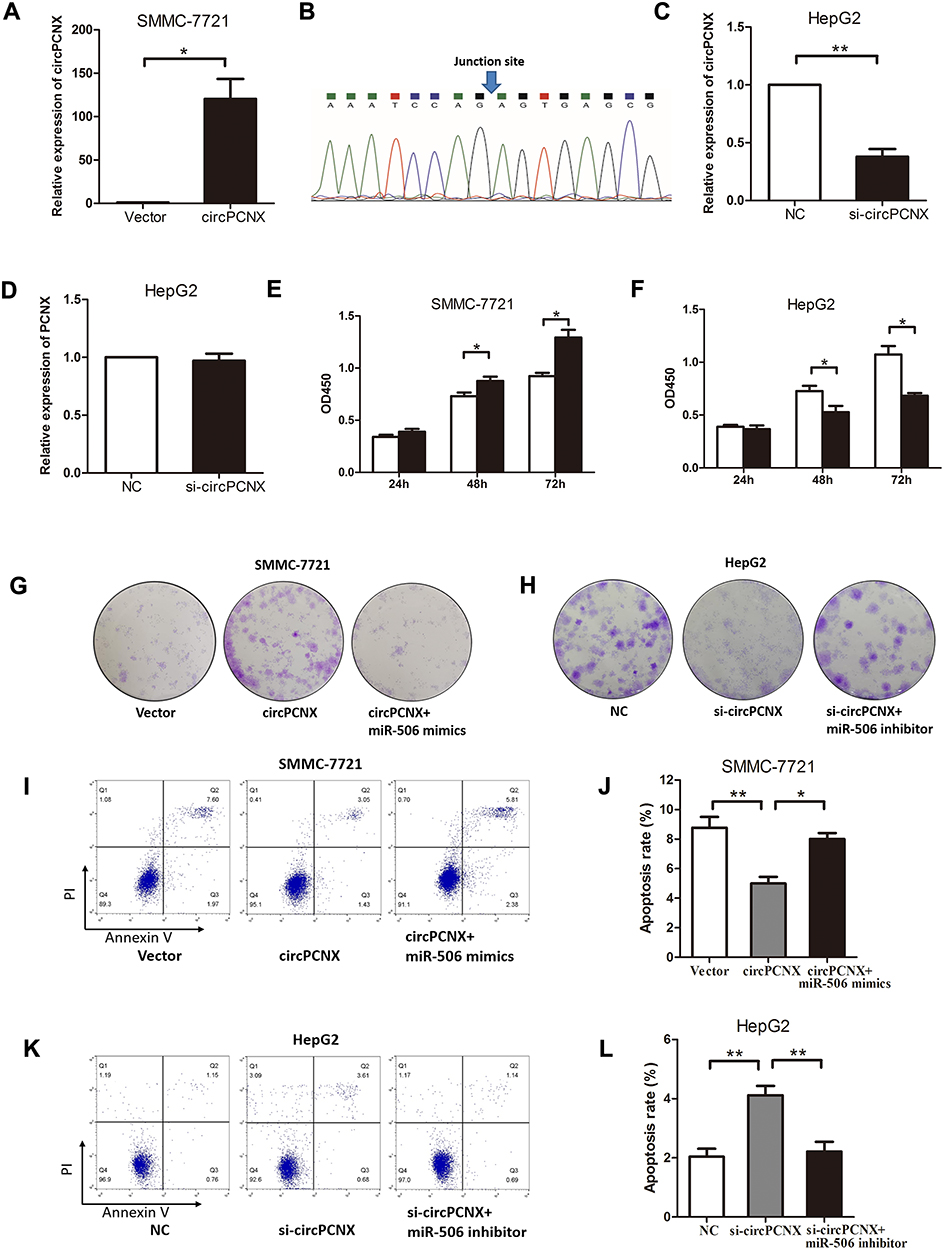 |
Figure 4 circPCNX promotes HCC cell viability via miR-506. (A and B) Relative circPCNX expression in SMMC-7721 cells transfected with circPCNX expression plasmids and validation by Sanger sequencing. (C and D) Relative circPCNX and PCNX expression in HepG2 cells transfected with circPCNX siRNA. (E and F) CCK8 assays, (G and H) colony formation assays, and (I-L) cell apoptosis assays in SMMC-7721 cells transfected with circPCNX expression plasmids and miR-506 mimics or HepG2 cells transfected with circPCNX siRNA and miR-506 inhibitor. Results are presented as means± SD *P<0.05 and **P<0.01. Abbreviations: NC, negative control; OD450, optical density at 450 nm. |
PCNX-3ʹUTR Promotes HCC Cell Viability via a miR-506-Dependent Manner
Because the PCNX-3ʹUTR promotes lung cancer cell proliferation via a PCNX-miRNA-Skp2 regulatory pattern,13 we then explored the function of the PCNX-3ʹUTR in HCC. Using microRNA.org, we identified a binding site for miR-506 in the PCNX-3ʹUTR (Figure 5A). Then, we performed luciferase reporter assays to examine this interaction. The miR-506 mimics decreased the expression of the reporter gene containing the wt PCNX-3ʹUTR but not mut PCNX-3ʹUTR (Figure 5B), and miR-506 inhibitors induced the expression of the reporter gene containing wt PCNX-3ʹUTR but not mut PCNX-3ʹUTR (Figure 5C). Thus, PCNX-3ʹUTR was a direct target of miR-506. We also observed that Snail2 and YAP1 were significantly upregulated by overexpression of PCNX-3ʹUTR which could be rescued by miR-506 (Figure 5D). Overexpression of PCNX-3ʹUTR in SMMC-7721 cells promoted cell proliferation and restrained cell apoptosis which could be attenuated by miR-506 mimics (Figure 5E–H). Thus, PCNX might also function as an oncogene in HCC in a miR-506-dependent manner.
 |
Figure 5 PCNX-3ʹUTR promotes HCC cell viability via a miR-506-dependent manner. (A) Predicted miR-506-binding site and corresponding mutant sites in the 3ʹUTR of the PCNX mRNA (wt, wild-type; mt, mutant). (B and C) Luciferase activity in SMMC-7721 cells co-transfected with luciferase reporters and miR-506 mimics or miR-506 inhibitors. The luciferase activity of each group was normalized to the value obtained in the cells transfected with NC mimics. (D) Relative Snail2 and YAP1 expression in SMMC-7721 cells transfected with PCNX 3ʹUTR expression plasmids and miR-506 mimics, as analyzed using Western blot assays. (E-H) CCK8 assays, colony formation assays and cell apoptosis assays using SMMC-7721 cells transfected with PCNX 3ʹUTR expression plasmids and miR-506 mimics. Results are presented as means± SD *P<0.05, **P<0.01, and ***p<0.001. Abbreviations: NC, negative control; OD450, optical density at 450 nm. |
Discussion
Based on accumulating evidence, circRNAs might be involved in the progression and serve as biomarkers of many cancers, including HCC.14–16 In this study, we identified a large number of circular RNAs expressed in HCC tissues. Some circRNAs were also differentially expressed in HCC tissues compared with ANLTs, suggesting that these RNAs may be regulated and may exert a particular function. The expression of circPCNX was upregulated and positively correlated with the TNM stage and tumor size of HCC. The expression of circPCNX also correlated with poor clinical outcomes of patients with HCC. Moreover, circPCNX promoted the viability of HCC cells in vitro. Thus, circPCNX plays a vital role in HCC progression and may be a potential prognostic biomarker and therapeutic target in HCC.
PCNX was originally identified as a maternal neurogenic gene in Drosophila that potentially plays a critical role in maintaining the normal development of the nervous system of the fly.17 PCNX is a component of the N-signaling pathway, and its activity is essential for endoplasmic reticulum-regulated function in N signaling.18 A recent study revealed a role for PCNX in NSCLC, including promoting cell growth and proliferation, inducing cell cycle progression and repressing cell apoptosis. PCNX may function as a ceRNA of Skp2 to positively regulate Skp2 expression by inducing Akt phosphorylation in a miRNA-dependent manner.13 Our study also explored the function of PCNX in HCC and showed that the PCNX-3ʹUTR promoted HCC cell viability by binding to miR-506. Therefore, PCNX might also function as an oncogene in HCC in a miRNA-dependent manner.
Other researchers have shown that miR-506 suppresses the proliferation of HCC cells.19–21 The function of miR-506 has mainly been examined in cancer. Notably, miR-506 inhibits the proliferation, migration and invasion of various types of cancer.22–25 In this study, we predicted that circPCNX may function as a sponge for miR-506 and verified this hypothesis using luciferase reporter assays. We presented that circPCNX promoted HCC cell viability and increased the expression of the miR-506 target genes Snail226 and YAP127 through regulating miR-506. Since Snail2 and YAP1 are well-known oncogenes in cancers, these results further revealed the mechanism by which the circPCNX-miR-506-Snail2/YAP axis promotes the progression of HCC.
However, in this study, the expressions and functions of circPCNX and PCNX were only investigated in human tissues and in vitro. Therefore, the lack of an in vivo study is a limitation of this study.
Conclusions
In summary, our study characterized a differentially expressed circRNA derived from the PCNX gene. We propose that circPCNX and PCNX are new prognostic markers for HCC. Based on our findings, circPCNX and PCNX may promote HCC progression by regulating miR-506.
Abbreviations
HCC, hepatocellular carcinoma; circRNA, circular RNAs; PCNX, Pecanex; miRNA, microRNA; NSCLC, non-small cell lung cancer; ceRNA, competitive endogenous RNA; Skp2, S-phase kinase associated protein 2; ANLTs, adjacent nontumor liver tissues; qRT-PCR, quantitative real-time PCR; DFS, disease-free survival; OS, overall survival; CCK-8, Cell Counting Kit-8.
Acknowledgment
This study was supported by a grant from the Qingdao Public Field Science and Technology Program (2014QPS01-68).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer J Clin. 2012;62:10–29. doi:10.3322/caac.20138
2. Llovet JM, Zucman-Rossi J, Pikarsky E, et al. Hepatocellular carcinoma. Nat Rev Dis Primers. 2016;2:16018. doi:10.1038/nrdp.2016.18
3. Poon RT. Differentiating early and late recurrences after resection of HCC in cirrhotic patients: implications on surveillance, prevention, and treatment strategies. Ann Surg Oncol. 2009;16:792–794. doi:10.1245/s10434-009-0330-y
4. Zimmerman MA, Ghobrial RM, Tong MJ, et al. Recurrence of hepatocellular carcinoma following liver transplantation: a review of preoperative and postoperative prognostic indicators. Arch Surg. 2008;143:
5. Chen LL, Yang L. Regulation of circRNA biogenesis. RNA Biol. 2015;12:381–388. doi:10.1080/15476286.2015.1020271
6. Hentze MW, Preiss T. Circular RNAs: splicing’s enigma variations. EMBO J. 2013;32:923–925. doi:10.1038/emboj.2013.53
7. Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495:333–338. doi:10.1038/nature11928
8. Jeck WR, Sorrentino JA, Wang K, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013;19:141–157. doi:10.1261/rna.035667.112
9. Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495:384–388. doi:10.1038/nature11993
10. Chen J, Li Y, Zheng Q, et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. 2017;388:208–219. doi:10.1016/j.canlet.2016.12.006
11. He R, Liu P, Xie X, et al. circGFRA1 and GFRA1 act as ceRNAs in triple negative breast cancer by regulating miR-34a. J Exp Clin Cancer Res. 2017;36:145. doi:10.1186/s13046-017-0614-1
12. Zhang J, Liu H, Hou L, et al. Circular RNA_LARP4 inhibits cell proliferation and invasion of gastric cancer by sponging miR-424-5p and regulating LATS1 expression. Mol Cancer. 2017;16:151. doi:10.1186/s12943-017-0719-3
13. Li J, Tian H, Pan J, et al. Pecanex functions as a competitive endogenous RNA of S-phase kinase associated protein 2 in lung cancer. Cancer Lett. 2017;406:36–46. doi:10.1016/j.canlet.2017.07.030
14. Han D, Li J, Wang H, et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology. 2017;66:1151–1164. doi:10.1002/hep.29270
15. Qin M, Liu G, Huo X, et al. Hsa_circ_0001649: a circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark. 2016;16:161–169. doi:10.3233/CBM-150552
16. Xu L, Zhang M, Zheng X, Yi P, Lan C, Xu M. The circular RNA ciRS-7 (Cdr1as) acts as a risk factor of hepatic microvascular invasion in hepatocellular carcinoma. J Cancer Res Clin Oncol. 2017;143:17–27. doi:10.1007/s00432-016-2256-7
17. Gilbert TL, Haldeman BA, Mulvihill E, O’Hara PJ. A mammalian homologue of a transcript from the Drosophila pecanex locus. J Neurogenet. 1992;8:181–187. doi:10.3109/01677069209083447
18. Yamakawa T, Yamada K, Sasamura T, et al. Deficient Notch signaling associated with neurogenic pecanex is compensated for by the unfolded protein response in Drosophila. Development. 2012;139:558–567. doi:10.1242/dev.073858
19. Wang Y, Cui M, Sun BD, Liu FB, Zhang XD, Ye LH. MiR-506 suppresses proliferation of hepatoma cells through targeting YAP mRNA 3ʹUTR. Acta Pharmacol Sin. 2014;35:1207–1214. doi:10.1038/aps.2014.59
20. Dai W, Huang HL, Hu M, et al. microRNA-506 regulates proliferation, migration and invasion in hepatocellular carcinoma by targeting F-spondin 1 (SPON1). Am J Cancer Res. 2015;5:2697–2707.
21. Deng Q, Xie L, Li H. MiR-506 suppresses cell proliferation and tumor growth by targeting Rho-associated protein kinase 1 in hepatocellular carcinoma. Biochem Biophys Res Commun. 2015;467:921–927. doi:10.1016/j.bbrc.2015.10.043
22. Deng L, Liu H. MicroRNA-506 suppresses growth and metastasis of oral squamous cell carcinoma via targeting GATA6. Int J Clin Exp Med. 2015;8(2):1862–1870.
23. Wen SY, Lin Y, Yu YQ, et al. miR-506 acts as a tumor suppressor by directly targeting the hedgehog pathway transcription factor Gli3 in human cervical cancer. Oncogene. 2014;34(6):717–725. doi:10.1038/onc.2014.9
24. Liu G, Sun Y, Ji P, et al. MiR-506 suppresses proliferation and induces senescence by directly targeting the CDK4/6-FOXM1 axis in ovarian cancer. J Pathol. 2014;233(3):308–318. doi:10.1002/path.4348
25. Zhang Z, Ma J, Luan G, et al. MiR-506 suppresses tumor proliferation and invasion by targeting FOXQ1 in nasopharyngeal carcinoma. PLoS One. 2015;10(4):e0122851. doi:10.1371/journal.pone.0122851
26. Yu Z, Zhang Y, Gao N, et al. Overexpression of miR-506 inhibits growth of osteosarcoma through Snail2. Am J Transl Res. 2015;7(12):2716.
27. Hua K, Yang W, Song H, et al. Up-regulation of miR-506 inhibits cell growth and disrupt the cell cycle by targeting YAP in breast cancer cells. Int J Clin Exp Med. 2015;8(8):12018.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.