Back to Journals » OncoTargets and Therapy » Volume 12
circANKS1B regulates FOXM1 expression and promotes cell migration and invasion by functioning as a sponge of the miR-149 in colorectal cancer
Authors Li D, Yang R, Yang L, Wang D, Zhou X, Sun Y
Received 12 January 2019
Accepted for publication 7 March 2019
Published 23 May 2019 Volume 2019:12 Pages 4065—4073
DOI https://doi.org/10.2147/OTT.S201310
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sanjay Singh
Dongzheng Li,1 Ruixue Yang,2 Liu Yang,1 Dongfeng Wang,1 Xin Zhou,1 Yan Sun1
1Department of General Surgery, Jiangsu Cancer Hospital, Jiangsu Institute of Cancer Research, Nanjing Medical University Affiliated Cancer Hospital, Nanjing, Jiangsu 210009, People’s Republic of China; 2Department of Paediatrics, Taikang Xianlin Drum Tower Hospital, Nanjing, Jiangsu 210046, People’s Republic of China
Background: Circular RNAs (circRNAs) and microRNAs (miRNAs) play key roles in the development of human cancers. CircANKS1B has been reported to be increased in breast cancer.
Methods: Real-time polymerase chain reaction (real-time PCR) assay was used to measure expressions of circANKS1B, ANKS1B, and FOXM1. Western blot assay was employed to examine the protein level of FOXM1 and Slug. The abilities of cell migration and invasion were measured by wound-healing and transwell assays. The interaction between circANKS1B and miR-149 was confirmed by site-directed mutagenesis and luciferase assays.
Results: The expression of circANKS1B was up-regulated in colorectal cancer tissues and cells. Additionally, circANKS1B increased the expression of FOXM1. Furthermore, the enhancement of CRC cell migration and invasion by circANKS1B was dependent on FOXM1. However, previous studies have shown that miR-149 can directly target FOXM1 and act as tumor suppressor in CRC. Consequently, our results showed that miR-149 could directly bind to circANKS1B and FOXM1. The inhibition of circANKS1B could reduce FOXM1 and Slug protein levels, thus suppressing CRC cell migration and invasion.
Conclusion: Taken together, circANKS1B promotes colorectal cancer cell migration and invasion by acting as a molecular sponge of miR-149 to modulate FOXM1 and Slug protein levels.
Keywords: circANKS1B, cell invasion, miR-149, FOXM1, colorectal cancer
Introduction
Colorectal cancer (CRC) is one of the most frequent malevolence with approximately 1.2 million new cases every year, which is the second leading cause of cancer-related death and the third in the cancer morbidity worldwide.1,2 In addition to the developed countries, CRC is also prevalent in developing countries recently, particularly in China.3 Colorectal cancer is a multi-step biological process, including imbalance of cell differentiation, proliferation, and apoptosis, and polygenic alteration.4 Even though the tremendous advances in traditional surgery, targeted treatment and radiochemotherapy, the exact molecular mechanisms of CRC are still not uncovered.5 Thus, it is needed to understand the molecular mechanisms in CRC progression emergently and find new therapeutic strategies as well as prognostic biomarkers.
Circular RNAs (circRNAs), which are different from the canonical splicing of linear RNAs, represent a novel class of conserved endogenous RNAs that can regulate gene expression in many species.6 So far, lots of research have shown that more than 1,00,000 circRNAs have been predicted in different spices.7 Some reports have shown that circRNAs can reserve high cell and tissue specificity in different mammalian cells. For functional studies, circRNAs are described to be involved in complex RNA–RNA interaction networks and play a role in post-transcriptional gene regulation.8 Additionally, circRNAs can bind to proteins directly or via RNA-mediated indirect interactions.7 More and more works of literature show that circRNAs can bind to miRNAs, and is rich in functional miRNA-binding sites used primarily to regulate cancer progression, and these circRNA-miRNA axes are involved in various disease pathways, such as cancer metastasis, apoptosis, and invasion.9–11 Recently, Rong et al have shown that circPSMC3 suppresses the proliferation and metastasis of gastric cancer by acting as a competitive endogenous RNA through sponging miR-296-5p.12 Moreover, circRNAs showed distinctive prognostic information from their linear counterparts, hinting at distinctive clinical roles.13,14
Our study aims to investigate the effects of circANKS1B (hsa_circ_0007294) on colorectal cancer. The expression of circANKS1B is increased and correlated with the metastasis in breast cancer.15 In our study, we detected that circANKS1B was significantly upregulated in colorectal cancer tissues and cells. Further functional analysis showed that circANKS1B could affect CRC cell migration and invasion via acting as a ceRNA to regulate FOXM1 expression by decoying the miR-149. This study relieves that circANKS1B could be a novel potential target of CRC.
Material and methods
Clinical ethics and human tissues
Forty paired human paratumor and tumor tissue samples were obtained from newly diagnosed colorectal cancer patients in Jiangsu Cancer Hospital (Nanjing, China). The tissues were stored in the fridge at −80°C. All experiments were approved by the research ethics committee of Nanjing Medical University Affiliated Cancer Hospital (Nanjing, China). All patients gave informed consent and written informed consent was obtained.
Cell culture
The normal human intestinal epithelial cells (HIEC) and human colorectal cancer cell lines SW480, LoVo, HT29, SW480, and Caco-2 were obtained from Cellbank of Chinese Academy of Sciences (Shanghai, China). The SW480, LoVo, and HT29 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM, high glucose) supplemented with 10% fetal bovine serum (BI, Israel). The SW480 and Caco-2 cell lines were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (BI, Israel). The cells were cultured at 5% CO2 and humidified 37°C chamber (Thermo Fisher, USA).
Cell transfection
miR-149 mimics, inhibitor, mimics negative control, and inhibitor negative control were purchased from GenePharma Biotechnology (Shanghai, China). A total of 5×105 cells were seeded in per well of six-well plate. After 24 hrs, mix 10 μL NC, miR-149 mimics or inhibitor, 185 μL DMEM, and 5 μL lipofectamine 2,000 reagent (Thermo Fisher, USA). siRNA-ANKS1B and siRNA-circANKS1B were purchased from GenePharma Biotechnology (Shanghai, China). A total of 5×105 cells were seeded in per well of six-well plate. After 24 hrs, mix 15 μL NC, siRNA-ANKS1B and siRNA-circANKS1B, 180 μL DMEM, and 5 μL lipofectamine 2,000 reagent (Thermo Fisher, USA). After 20 mins, pour the mix into a well of six-well plate and cultured in cell culture chamber. siRNA target sites of circANKS1B: 5ʹ-GAAGCCAGAGTGTAACAGA-3ʹ, siRNA target sites of ANKS1B: 5ʹ-CCAGTCGTGT GATTACAAA-3ʹ and FOXM1: 5ʹ- GCCAATCGTTCTCTGACAGAA-3ʹ.
Plasmids construction
The wild type and mutation of circANKS1B-3ʹUTR were cloned into the pmirGLO (Promega, USA), as pmirGLO-WT-circANKS1B or pmirGLO-mut-circANKS1B. The plasmids were cloned into pmirGLO by GENEWIZ (Suzhou, China). A total of 5×105 cells were seeded in per well of six-well plate. After 24 hrs, mix 10 μL miR-149 mimics and 4 μg constructed plasmids, 185 μL DMEM, and 5 μL lipofectamine 2,000 reagent (Thermo Fisher, USA). After 36 hrs, the cells were lysed and assayed with a Dual Luciferase Reporter Assay Kit (Promega, USA) according to the manufacturer’s instructions. The hRluc (Humanized Renilla luciferase) values were measured and normalized to luc2 (firefly luciferase) values.
Real-time PCR
Total RNA was extracted from tissue and cell samples using Trizol reagent (Thermo Fisher, USA) according to the manufacturer’s instructions. The cDNA was synthesized from total RNA using PrimeScript RT-PCR Kit (TAKARA, Japan). The mRNA levels for circANKS1B, ANKS1B, and GAPDH were measured by real-time PCR on a StepOne Plus real-time PCR instruments (Applied Biosystems, USA) by using SYBR Premix Ex Taq (Takara, Japan). The relative mRNA expression level of each gene was normalized against GAPDH. The microRNA measure kit for miR-149 was purchased from GenePharma (Shanghai, China) and the level of miR-149 was tested according to the manufacturer’s instructions. The level of miR-149 was normalized to U6.
Primers for real-time PCR: circANKS1B forward: 5ʹ-GAAACCGTCACTGGAGAATTATCA-3ʹ, circANKS1B reverse: 5ʹ-AAAGCTGCTTCATGAAGTGCAC-3ʹ;
ANKS1B forward: 5ʹ-CAACCTTCGGAAAAGCAGATG-3ʹ,
ANKS1B reverse: 5ʹ-TGGTGCCCGTAAGTGCAGAC-3ʹ;
GAPDH forward: 5ʹ-GAACGGGAAGCTCACTGG-3ʹ,
GAPDH reverse: 5ʹ-GCCTGCTTCACCACCTTCT-3ʹ.
Western blot
Cells were lysed in RIPA lysis buffer with 1 mM PMSF and protease inhibitor cocktail for 30 mins on ice, protein quantificated with BCA protein assay kit (Bio-rad, USA). Equal amounts of protein were separated by SDS-PAGE and transferred onto polyvinylidene fluoride (PVDF) membrane (Millipore, USA). The membranes were blocked with 5% BSA (Beyotime, China) and incubated the following antibodies: FoxM1 (CST, #5436, 1:1000), Slug (CST, #9585, 1:1000), and GAPDH (CST, #5174, 1:1000) at 4°C overnight. After wash with three times, the membranes were incubated with HRP-conjugated secondary antibody (1:5000) at room temperature. The signals were visualized using ECL kit (Thermo Fisher, USA). GAPDH was used as an endogenous protein for normalization.
Wound-healing and transwell assay
When reaching 80–90% confluence, the cell layer was scratched. Cells were then continued cultured, and the average extent of wound closure was quantified. Transwell assays were performed using 24-well transwells (8-μm, Millipore, USA) precoated with Matrigel (BD Biosciences, USA). Cells were then harvested, and 105 cells were seeded in serum-free medium into the upper chamber, whereas medium supplemented with 20% FBS was applied to the lower chamber as a chemoattractant. After 24 hrs, the migrated cells at the bottom surface of the filter were fixed with pre-cooled methanol, stained with 0.2% crystal violet solution, and counted with microscope.
Statistical analysis
The statistical analyses were performed using Prism 5 software (GraphPad Software, USA). Data in the bar graphs are presented as mean±SD. The P-values calculated by Student’s t test or ANOVA (Analysis of Variance) and P<0.05 was considered to be statistically significant.
Results
CircANKS1B is significantly upregulated in CRC tissues and cells
circANKS1B (Hsa_circ_0007294) is derived from exons 5 to 8 of the ANKS1B gene (Figure 1A). Additionally, we used 40 pairs of CRC tumor tissues and paratumor tissues to examine the expression of circANKS1B. The real-time PCR analysis showed that the expression of circANKS1B was identified to be upregulated (approximately twofold) in CRC tumor tissues compared with paratumor tissues (Figure 1B). Then, we performed Chi-square tests to explore the association between the clinicopathological characteristics and circANKS1B expression. The findings are summarized in Table 1. Increased circANKS1B expression was significantly associated with regional lymph node metastasis (P=0.004) and distance metastasis (P=0.018). No significant correlations were found between circANKS1B expression and age, gender, tumor size, and T stage. To focus on the biological function of circANKS1B, we detected the expression levels of circANKS1B in CRC cell lines and circANKS1B was significantly up-regulated in CRC cell lines compared with normal human intestinal epithelial cells (HIEC) (Figure 1C).
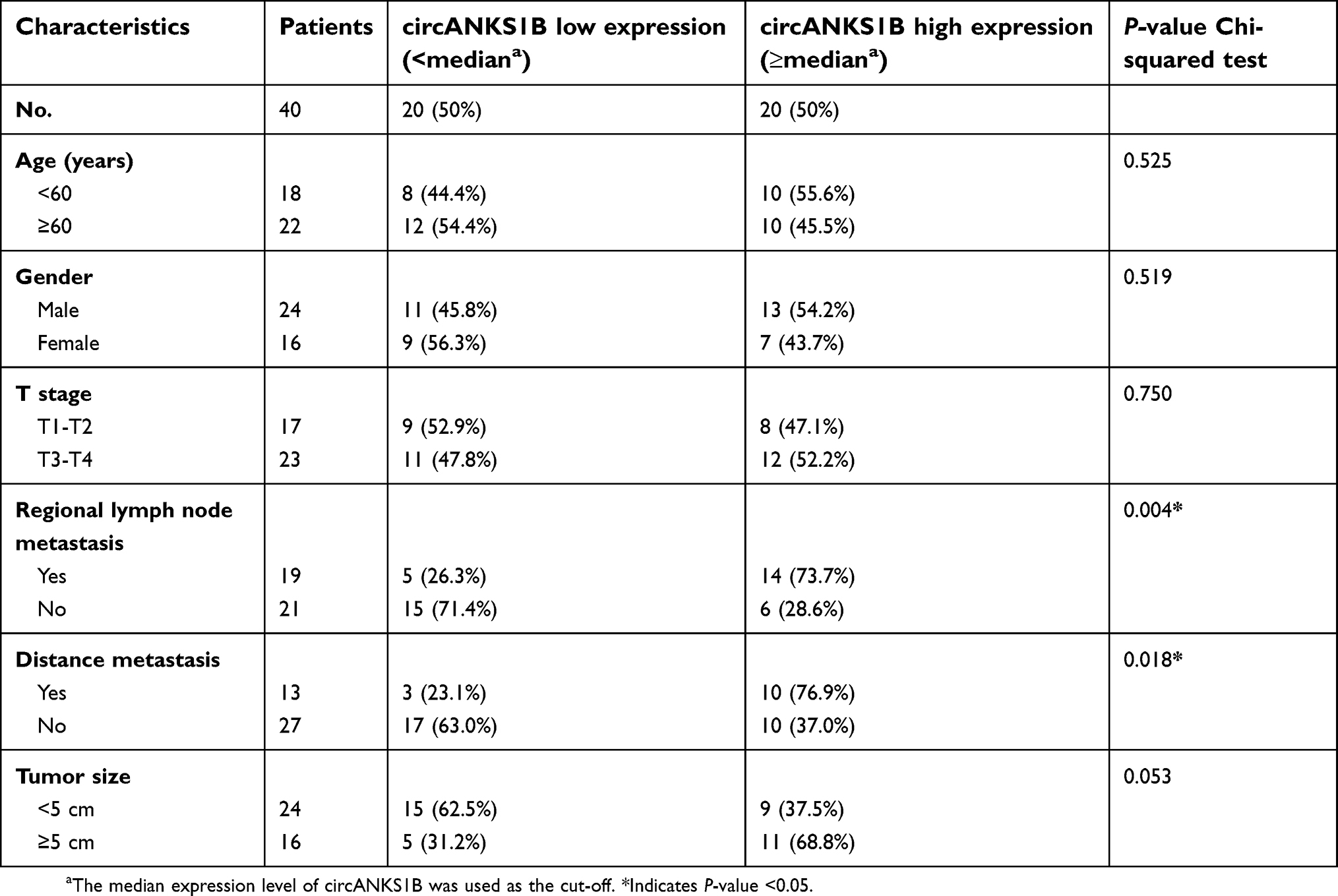 | Table 1 Correlation between circANKS1B expression and clinicopathological characteristics of colorectal cancer patients (n=40) |
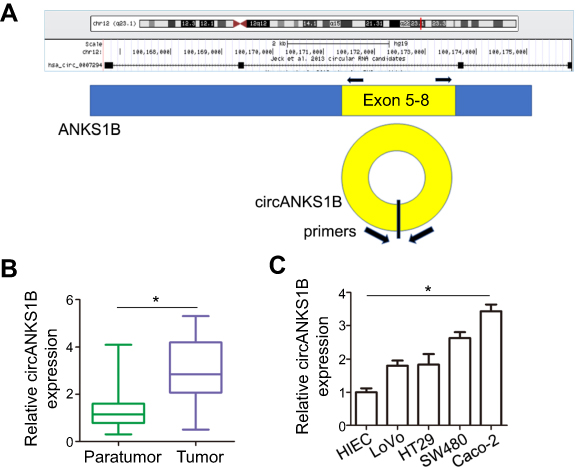 | Figure 1 Increased levels of circANKS1B expression in CRC tissues and cell lines. (A) Schematic model of the primers of circANKS1B. The primers target the back-splice junction of circANKS1B. (B) Real-time PCR analysis of circANKS1B expression in paratumor and tumor tissues of 40 CRC tissues. The expression of circANKS1B was normalized by GAPDH expression. Data are shown as mean±SD; *P<0.05 by Student’s t test. (C) Real-time PCR analysis of circANKS1B expression in normal human intestinal epithelial cells (HIEC) and various tumor cell lines. Data are shown as mean±SD; n=3, *P<0.05 by ANOVA.Abbreviations: CRC, colorectal cancer; PCR, polymerase chain reaction; SD, standard deviation; ANOVA, analysis of variance. |
CircANKS1B promotes cell migration and invasion in CRC
To analyze the role of circANKS1B in CRC, we used circANKS1B-specific small interfering RNAs (siRNAs) to down-regulate the expression levels of circANKS1B. Thus, a siRNA targeting the backsplice junction sequence of circANKS1B was designed (Figure 2A). We found that siRNA targeting the backsplice junction knocked down only the circular transcript and did not affect the expression of linear species. Contrarily, siRNA targeting linear transcript knocked down only the ANKS1B linear transcript but not the circular transcript (Figure 2B and C). To determine the biological function of circANKS1B, we transfected circANKS1B-specific siRNAs in CRC cell lines SW480 and Caco-2. The findings of wound-healing assay displayed that cell migration ability was decreased following the silencing of circANKS1B in CRC cells (Figure 2D and E). Similarity, the results of transwell assay showed that the invasive capability was dramatically reduced in siRNA-circANKS1B transfected CRC cells (Figure 2F and G), suggesting that circANKS1B is crucial for CRC migration and invasion.
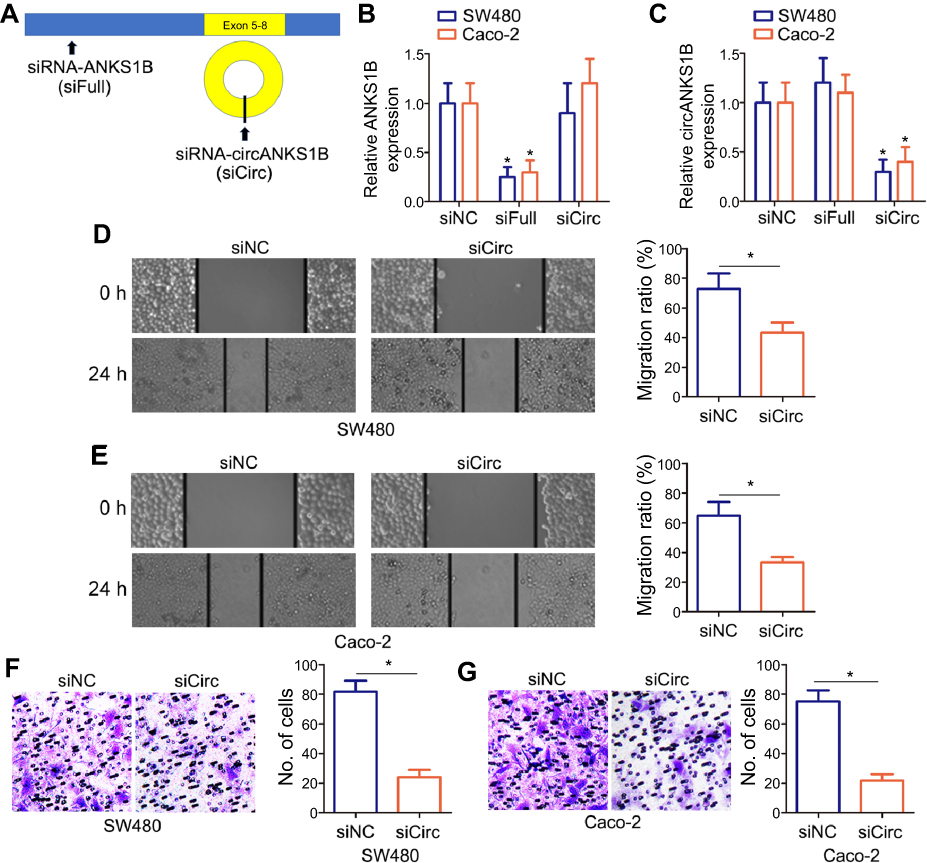 | Figure 2 Silencing of circANKS1B inhibited cell migration and invasion in CRC cells. (A) Schematic model of the siRNAs. siFull (siRNA of ANKS1B) targets the ANKS1B linear transcript, siCirc (siRNA of circANKS1B) targets the back-splice junction of circANKS1B. (B and C) Real-time PCR analysis of ANKS1B (B) and circANKS1B (C) expression in CRC tumor cell lines SW480 and Caco-2 with siFull or siCirc transfection. Data are shown as mean±SD; n=3, *P<0.05 by ANOVA. (D and E) The wound-healing assay was performed to determine the migration of CRC tumor cell lines SW480 (D) and Caco-2 (E) with siCirc transfection. The relative migration rate was quantified after scratch 24 hrs. (100X) . The data were presented as the mean±SD, n=3, *P<0.05 by Student’s t test. (F and G) The ability of cell invasion was measured by transwell assay in CRC tumor cell lines SW480 (F) and Caco-2 (G) with siCirc transfection. The representative magnified sections of transwell cell staining images are shown. Statistical results based on three independent experiments is shown in right. (200X). Data are shown as mean±SD; n=3, *P<0.05 by Student’s t test.Abbreviations: CRC, colorectal cancer; SD, standard deviation; ANOVA, analysis of variance. |
CircANKS1B is a direct target of miR-149 in CRC cells
We then used CircInteractome database (
 | Figure 3 circANKS1B served as a sponge for the microRNA miR-149. (A) A schematic model showing the putative binding sites for miRNAs and 3ʹUTR of circANKS1B. (B and C) miR-149 mimics or negative control (NC) was co-transfected with pmirGLO-WT-circANKS1B-3ʹUTR (B) or pmirGLO-mut-circANKS1B-3ʹUTR (C) in CRC cell line SW480. The hRluc values were measured and normalized to luc2 (firefly luciferase) values. Data are shown as mean±SD; n=3, *P<0.05 by ANOVA or Student’s t test.Abbreviations: CRC, colorectal cancer; UTR, untranslated region; hRluc, humanized renilla luciferase. |
CircANKS1B regulates FOXM1 expression by serving as a sponge for miR-149
Previously, several studies have shown that miR-149 suppresses CRC migration and invasion by directly targeting FOXM1.17 FOXM1 was found to endogenously bind to and stimulate the promoter of Slug that was crucial for cancer progression.18 So, we examined the mRNA and protein expression of FOXM1 in the group transfected with siRNA-circANKS1B and miRNA-149 inhibitor. The results showed that knockdown of circANKS1B inhibited the expression of FOXM1 and Slug, while the effects were abrogated by down-regulating miR-149 expression with transfection of miR-149 inhibitor in CRC cells SW480 and Caco-2 (Figure 4A and B), and invasive capability showed the same results (Figure 4C). To confirm the effects of FOXM1 on Slug expression, we examined the levels of Slug in CRC cells transfected with siRNA-FOXM1. The Western blot results showed that silencing of FOXM1 decreased the levels of Slug protein compared with siRNA negative control group (Figure 4D). These findings illustrated that circANKS1B took part in the invasion of CRC through the increasing expression of FOXM1 and Slug by competing for miR-149 (Figure 5).
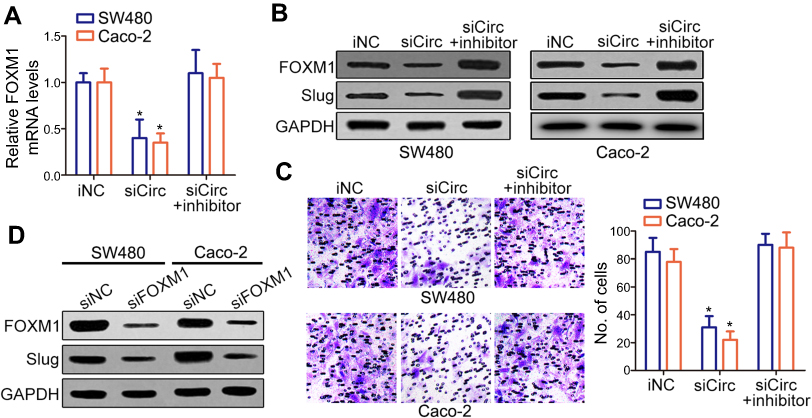 | Figure 4 Silencing of circANKS1B repressed FOXM1 expression. (A–C) siCirc (siRNA of circANKS1B) or miR-149 iNC was transfected or co-transfected with miR-149 inhibitor in CRC cell lines SW480 and Caco-2. (A) Real-time PCR analysis of FOXM1 mRNA expression in above CRC cell lines SW480 and Caco-2. Data are shown as mean±SD; n=3, *P<0.05 by ANOVA. (B) Western blot analysis of FOXM1 and Slug protein levels in CRC cell lines SW480 and Caco-2. The representative blots were shown from three independent experiments. (C) The ability of cell invasion was measured by transwell assay in CRC tumor cell lines SW480 and Caco-2. The representative magnified sections of transwell cell staining images are shown. Statistical results based on three independent experiments is shown in right. (200×) Data are shown as mean±SD; n=3, *P<0.05 by ANOVA. (D) Western blot analysis of FOXM1 and Slug protein levels in CRC cell lines SW480 and Caco-2 with siFOXM1 (siRNA of FOXM1) or siRNA negative control (siNC). The representative blots were shown from three independent experiments.Abbreviations: iNC, inhibitor negative control; CRC, colorectal cancer; SD, standard deviation; ANOVA, analysis of variance. |
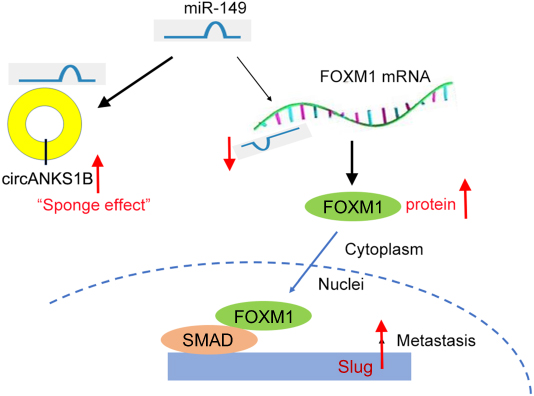 | Figure 5 The schematic model demonstrates that circANKS1B regulates FOXM1/Slug pathway by serving as a sponge for the miR-149 in colorectal cancer metastasis. |
Discussion
Hsu and Coca-Prado used electron microscopy to observe circRNAs first time, and then circRNAs were found in yeast.19 Over decades ago, circRNAs, constructed by exons of a handful transcribed genes, were also discovered in human cells. circRNAs are thought to be produced by gene rearrangement, abnormal splicing, or non-linear reverse splicing.8 Moreover, circRNAs have become promising biomarkers, therapy targets, and may exert potential functions in the regulation of gene expression.7 Actually, most circRNAs have no function and only a few circRNAs have been definite as efficient miRNA sponges.20 For example, ciRS-7, circRNA-100782, and circRNA-Sry were reported to function as a sponge of miR-7, miR-124, and miR-138, respectively,.20–22 In a recent study, circANKS1B is derived from exons 5 to 8 of ANKS1B gene and is observed to act as a sponge for miR-148a and miR-152-3p in breast cancer.15
In this study, circANKS1B was thought to be functioned as a decoy of the miR-149 regulating FOXM1 expression. There are multiple miRNA-binding sites predicated in circANSKS1B through MREs analysis. In fact, some circRNAs have multiple binding sites for a particular miRNA, most of which contain only one or two miRNA binding sites.23 As we all know, miRNAs play important roles in many biological functions of cancer, including tumor development, tumorigenesis, metastasis, and drug resistance.24 Among them, miR-149 serves as an onco-microRNA and is significantly dysregulated in several cancers, such as breast cancer, lung cancer, and prostate cancer.25 Our results showed that circANKS1B was also increased in CRC tumor tissues and served as a direct target of miR-149 in CRC cells. Forkhead box transcription factor (FOXM1) is defined as owning a common DNA-binding domain and winged-helix domain. It functions as a classical transcription factor, regulating tumorigenesis and metastasis.26,27 FOXM1 is also an important oncogenic transcription factor and has been implicated in cell migration, invasion, angiogenesis, and metastasis.26,28 For example, FOXM1 promotes the epithelial to mesenchymal transition by stimulating the transcription of Slug in human breast cancer.18 Our findings revealed that miR-149 could target both FXOM1 and circANKS1B to modulate CRC cell migration and invasion.
In our study, several pieces of evidence implicated that circANKS1B could function as a ceRNA to FOXM1 in colorectal cancer by functioning as a sponge of miR-149. First, bioinformatic analyses indicated that the 3ʹUTR of circANKS1B contained binding sites for miR-149 and luciferase reporter assay proved this prediction. Second, knockdown of circANKS1B reduced expression of FOXM1. Third, inhibition of miR-149 reversed the effects of circANKS1B silencing on CRC cell migration and invasion. All of the above results proposed that circANKS1B and FOXM1 might be a couple of ceRNAs that harbor miR-149 competitively. FOXM1 was found to endogenously bind to and stimulate the promoter of Slug that was crucial for cancer progression.18 Taken together, the accumulating evidence indicates that circRNAs may also functional in CRC progression and exert regulatory functions by sponging miR-149.29,30 Moreover, further specific work should be conducted to uncover whether circANKS1B plays a role in other cancer processes. The roles of circRNAs will be critical for understanding CRC pathological process and proposing an innovative insight into the identification of new potential therapeutic targets and new biomarkers of CRC.
Conclusion
In summary, our findings reveal that circANKS1B expression is significantly upregulated in CRC patients. Functionally and mechanistically, circANKS1B enhances CRC migration and invasion by the sponge activity on miR-149 and upregulation of FOXM1/Slug expression, indicating its tumor-promoting role in CRC development. Our data suggest that circANKS1B may have considerable potential as a prognosis predictor and therapeutic target for CRC.
Acknowledgments
This work is supported by the Research Center of Nanjing Medical University. Dongzheng Li and Ruixue Yang are co-first authors for this study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet. 2014;383(9927):1490–1502. doi:10.1016/S0140-6736(13)61649-9
2. Siegel R, Desantis C, Jemal A. Colorectal cancer statistics, 2014. CA Cancer J Clin. 2014;64(2):104–117. doi:10.3322/caac.21220
3. Zhu J, Tan Z, Hollis-Hansen K, Zhang Y, Yu C, Li Y. Epidemiological trends in colorectal cancer in China: an ecological study. Dig Dis Sci. 2017;62(1):235–243. doi:10.1007/s10620-016-4362-4
4. Lugli A, Kirsch R, Ajioka Y, et al. Recommendations for reporting tumor budding in colorectal cancer based on the International Tumor Budding Consensus Conference (ITBCC) 2016. Mod Pathol. 2017;30(9):1299–1311. doi:10.1038/modpathol.2017.46
5. Kuipers EJ, Grady WM, Lieberman D, et al. Colorectal cancer. Nat Rev Dis Primers. 2015;1(1):5065.
6. Pamudurti NR, Bartok O, Jens M, et al. Translation of CircRNAs. Mol Cell. 2017;66(1):9–21.e7. doi:10.1016/j.molcel.2017.02.021
7. Conn SJ, Pillman KA, Toubia J, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160(6):1125–1134. doi:10.1016/j.cell.2015.02.014
8. Haque S, Harries LW. Circular RNAs (circRNAs) in health and disease. Genes (Basel). 2017;8(12):E353. doi:10.3390/genes8120353
9. Rong J, Wang Q, Zhang Y, et al. Circ-DLG1 promotes the proliferation of esophageal squamous cell carcinoma. Onco Targets Ther. 2018;11(6):723–730. doi:10.2147/OTT.S175826
10. Han D, Li J, Wang H, et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology. 2017;66(4):1151–1164. doi:10.1002/hep.29270
11. Kristensen LS, Hansen TB, Veno MT, Kjems J. Circular RNAs in cancer: opportunities and challenges in the field. Oncogene. 2018;37(5):555–565. doi:10.1038/onc.2017.361
12. Rong D, Lu C, Zhang B, et al. CircPSMC3 suppresses the proliferation and metastasis of gastric cancer by acting as a competitive endogenous RNA through sponging miR-296-5p. Mol Cancer. 2019;18(1):25. doi:10.1186/s12943-019-0958-6
13. Vo JN, Cieslik M, Zhang Y, et al. The landscape of circular RNA in cancer. Cell. 2019;176(4):869–881.e13. Doi:10.1016/j.cell.2018.12.021
14. Chen S, Huang V, Xu X, et al. Widespread and functional RNA circularization in localized prostate cancer. Cell. 2019;176(4):831–843.e22. doi:10.1016/j.cell.2019.01.025
15. Zeng K, He B, Yang BB, et al. The pro-metastasis effect of circANKS1B in breast cancer. Mol Cancer. 2018;17(1):160. doi:10.1186/s12943-018-0914-x
16. Dudekula DB, Panda AC, Grammatikakis I, De S, Abdelmohsen K, Gorospe M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016;13(1):34–42. doi:10.1080/15476286.2015.1128065
17. Xu K, Liu X, Mao X, et al. MicroRNA-149 suppresses colorectal cancer cell migration and invasion by directly targeting forkhead box transcription factor FOXM1. Cell Physiol Biochem. 2015;35(2):499–515. doi:10.1159/000369715
18. Yang C, Chen H, Tan G, et al. FOXM1 promotes the epithelial to mesenchymal transition by stimulating the transcription of Slug in human breast cancer. Cancer Lett. 2013;340(1):104–112. doi:10.1016/j.canlet.2013.07.004
19. Hsu MT, Coca-Prados M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature. 1979;280(5720):339–340.
20. Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–388. doi:10.1038/nature11993
21. Xu L, Zhang M, Zheng X, Yi P, Lan C, Xu M. The circular RNA ciRS-7 (Cdr1as) acts as a risk factor of hepatic microvascular invasion in hepatocellular carcinoma. J Cancer Res Clin Oncol. 2017;143(1):17–27. doi:10.1007/s00432-016-2256-7
22. Chen G, Shi Y, Zhang Y, Sun J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. Onco Targets Ther. 2017;10(5):783–794. doi:10.2147/OTT
23. Kang BW, Jeon HS, Chae YS, et al. Impact of genetic variation in MicroRNA-binding site on susceptibility to colorectal cancer. Anticancer Res. 2016;36(7):3353–3361.
24. Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov. 2017;16(3):203–222. doi:10.1038/nrd.2016.246
25. Zhi Y, Zhou H, Mubalake A, et al. Regulation and functions of MicroRNA-149 in human cancers. Cell Prolif. 2018;51(5):e12465. doi:10.1111/cpr.12465
26. Gartel AL. FOXM1 in cancer: interactions and vulnerabilities. Cancer Res. 2017;77(12):3135–3139. doi:10.1158/0008-5472.CAN-16-3566
27. Wierstra I. The transcription factor FOXM1 (Forkhead box M1): proliferation-specific expression, transcription factor function, target genes, mouse models, and normal biological roles. Adv Cancer Res. 2013;118:97–398. doi:10.1016/B978-0-12-407173-5.00004-2
28. Halasi M, Gartel AL. Targeting FOXM1 in cancer. Biochem Pharmacol. 2013;85(5):644–652. doi:10.1016/j.bcp.2012.10.013
29. Qu S, Yang X, Li X, et al. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015;365(2):141–148. doi:10.1016/j.canlet.2015.06.003
30. Tang W, Ji M, He G, et al. Silencing CDR1as inhibits colorectal cancer progression through regulating microRNA-7. Onco Targets Ther. 2017;10:2045–2056. doi:10.2147/OTT.S131597
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.