Back to Journals » Cancer Management and Research » Volume 12
Circ_0084927 Facilitates Cervical Cancer Development via Sponging miR-142-3p and Upregulating ARL2
Authors Chen L, Zhang X, Wang S, Lin X, Xu L
Received 2 June 2020
Accepted for publication 15 August 2020
Published 28 September 2020 Volume 2020:12 Pages 9271—9283
DOI https://doi.org/10.2147/CMAR.S263596
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Seema Singh
Liquan Chen, Xiaowei Zhang, Su Wang, Xiaoting Lin, Lizhen Xu
Department of Obstetrics and Gynecology, The First Affiliated Hospital of Guangzhou Medical University, Guangzhou, Guangdong 510000, People’s Republic of China
Correspondence: Xiaowei Zhang Email [email protected]
Background: Cervical cancer is a fatal burden for women. Circular RNAs (circRNAs) are important regulators in cancer development. Our study aimed to investigate the function and action mechanism of a novel circRNA, circ_0084927, in cervical cancer.
Methods: The expression of circ_0084927, miR-142-3p and ADP-ribosylation factor-like protein 2 (ARL2) mRNA was measured by quantitative real-time polymerase chain reaction (qRT-PCR). For functional analyses, cell proliferation was assessed using cell counting kit-8 (CCK-8) assay. Cell cycle distribution was monitored by flow cytometry assay. Cell migration and cell invasion were evaluated by transwell assay. The interaction between miR-142-3p and circ_0084927 or ARL2 was predicted by the bioinformatics analysis and validated by dual-luciferase reporter assay and RNA immunoprecipitation assay (RIP) assay. The expression of ARL2 at the protein level was detected by Western blot. Animal tumor formation assay was performed to monitor the tumorigenicity of circ_0084927 in vivo.
Results: The expression of circ_0084927 and ARL2 was enhanced in cervical cancer tissues and cells, while the expression of miR-142-3p was opposite to them. Circ_0084927 knockdown significantly blocked cervical cancer cell proliferation, migration and invasion and induced cell cycle arrest. MiR-142-3p was targeted by circ_0084927, and miR-142-3p inhibition reversed the effects of circ_0084927 knockdown. Besides, miR-142-3p bound to ARL2, and the inhibitory effects of miR-142-3p restoration on cell proliferation, cycle, migration and invasion were counteracted by ARL2 overexpression. More importantly, circ_0084927 upregulated ARL2 expression by sponging miR-142-3p. Circ_0084927 knockdown retarded tumor growth in vivo by regulating miR-142-3p and ARL2.
Conclusion: Circ_0084927 accelerated the progression of cervical cancer partly by mediating the miR-142-3p/ARL2 axis.
Keywords: circ_0084927, miR-142-3p, ARL2, cervical cancer
Introduction
Cervical cancer, one of the most common gynecological tumors, has become the fourth leading cause of tumor-related deaths among women, second to breast cancer, colorectal cancer and lung cancer.1,2 According to statistical analysis, there were about 570,000 newly diagnosed cases of cervical cancer worldwide in 2018 and about 311,000 cases of death.2 Though significant improvements in screening, diagnosis, and treatment have made, the prognosis of patients with cervical cancer remains poor,3 and the survival rate for patients with cervical cancer is less than 50% within 5 years in certain countries.4 Therefore, exploring the molecular mechanisms of cervical cancer initiation, invasion and migration is essential to obtain better therapeutic effects for cervical cancer patients.
Recently, several circular RNAs (circRNAs) were identified to be dysregulated in cervical cancer tissues and involved in the pathogenesis of cervical cancer.5 CircRNAs are a type of non-coding RNA with unique circular-closed structures. CircRNAs are more stable than linear mRNA molecules because there are no 5ʹ cap and 3ʹ tail in structure.6,7 Hence, circRNAs are gradually regarded as more ideal biomarkers in the diagnosis and treatment of cancers.8 Numerous circRNAs were reported to regulate cervical cancer cell malignant activities, including cell proliferation, viability, cycle, autophagy, angiogenesis, migration and invasion, thereby affecting cancer development.9–11 Accumulating evidence suggests that circRNAs exert biological functions by ensnaring target microRNAs (miRNAs) in a “competing endogenous RNA (ceRNA)” mechanism, which broadens the insights to understand the role of circRNAs.12 Circ_0084927 is a novel circRNA, and its function is largely unknown in human cancers. It is necessary to investigate the role of circ_0084927 in cervical cancer.
The extensive disruption of miRNA expression in cancer tissues and cells is well studied, and the function of miRNAs has been widely determined in cancers.13 Some representative miRNAs are characterized as tumor biological regulators to modulate cancer progression, such as miR-143, miR-21 and let-7a in cervical cancer.14,15 MiR-142-3p was previously mentioned to suppress cell proliferation and invasion in cervical cancer.16 However, evidence supporting its function in cervical cancer is insufficient, and the potential mechanism of miR-142-3p action in cervical cancer is limited.
ADP-ribosylation factor-like protein 2 (ARL2) is a member of the ADP-ribosylation factor family, which belongs to small G-proteins.17 Small G-proteins are implicated in several cellular signaling pathways and functions, such as cell differentiation, vesicle transport and cytoskeletal construction.18 Besides, the involvement of ARL2 in cancer progression has been documented,17,19 hinting that ARL2 regulates tumor development. Whereas, the functional mechanism of ARL2 and the interplay among ARL2, miR-142-3p and circ_0084927 are unknown in cervical cancer.
Here, our study focused on the expression and implications of circ_0084927 in cervical cancer and unclosed the putative molecular mechanism associated with miR-142-3p and ARL2, aiming to further explore the pathogenesis of cervical cancer and provide a strategy for cancer treatment.
Materials and Methods
Tissues and Ethical Approval
All cervical cancer patients were pathologically diagnosed at the First Affiliated Hospital of Guangzhou Medical University. A total of 50 pairs of tumor tissues and adjacent normal tissues were obtained from these patients who had signed the written informed consent. The excised fresh tissues were placed into liquid nitrogen at once and then preserved at −80°C freezer. This study was conducted in line with the Ethics Committee of the First Affiliated Hospital of Guangzhou Medical University (No.2019GZ336). The correlation between circ_0084927 expression and clinicopathological parameters of patients is shown in Table 1.
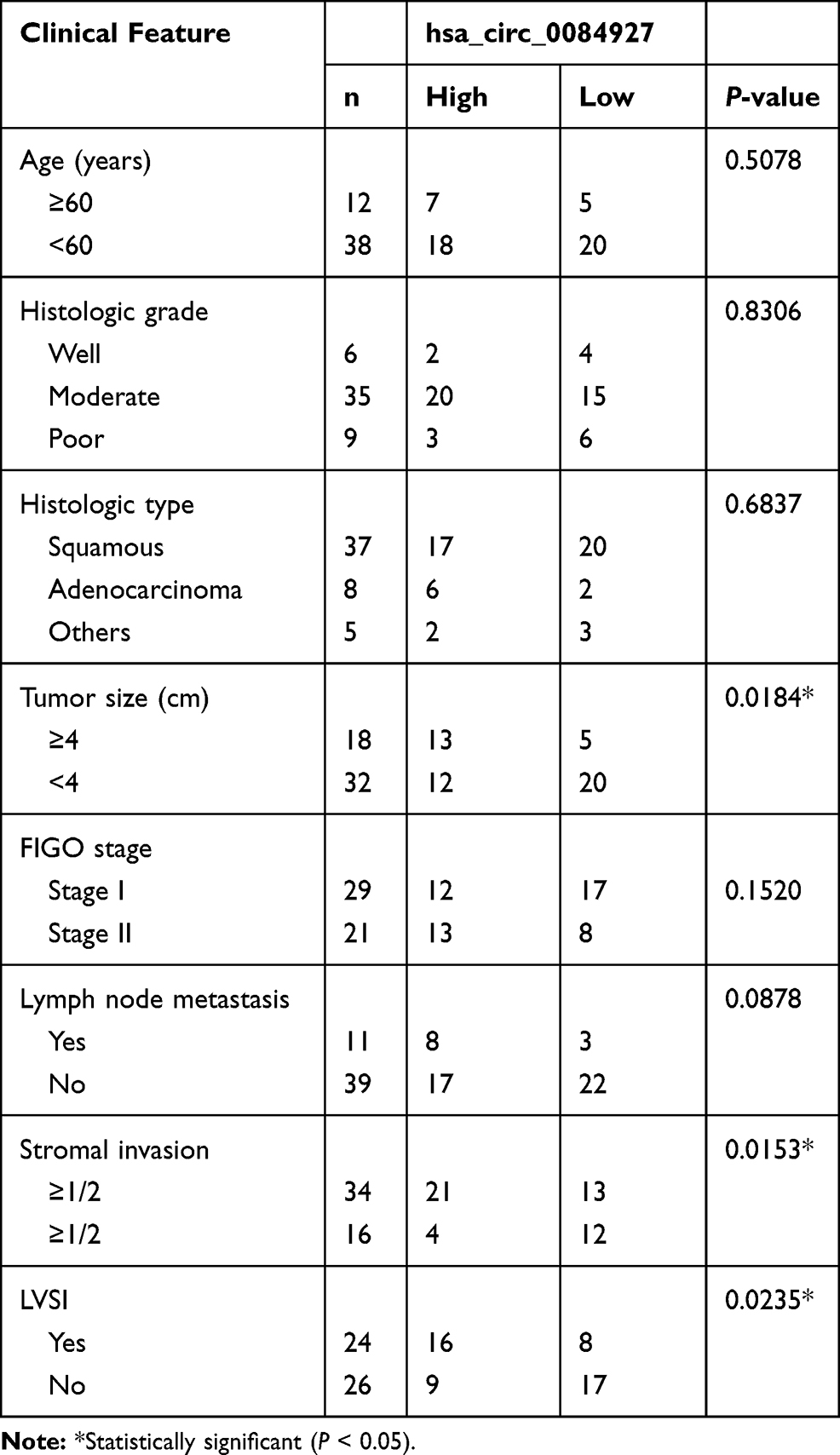 |
Table 1 Correlation Between hsa_circ_0084927 Expression and Clinicopathological Parameters of Patients (n = 50) |
Cell Lines and Cell Culture
Human cervical cancer cell lines (Hela and SiHa) and cervical epithelial cell line (H8) were purchased from Bena Culture Collection (Beijing, China). According to the culture methods, Hela and H8 cells were cultured in 90% Roswell Park Memorial Institute 1640 (RPMI 1640; Sigma-Aldrich, St. Louis, MO, USA) containing 10% fetal bovine serum (Sigma-Aldrich). SiHa cells were cultured in 90% Dulbecco’s Modified Eagle Medium (DMEM; Sigma-Aldrich) plus 10% FBS. All cells were cultured in a 37°C incubator supplemented with 5% CO2.
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
Cells or tissues were exposed to the TRIzol reagent (Sigma-Aldrich) to obtain total RNA. Then, reverse transcription and qPCR amplification for circ_0084927 and ARL2 were performed using the riboSCRIPT qRT-PCR Starter Kit (Ribobio, Guangzhou, China). Besides, reverse transcription and qPCR amplification for miR-142-3p were performed using the riboSCRIPT miRNA qRT-PCR Starter Kit (Ribobio). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was utilized as an endogenous control for circRNA and mRNA, and U6 was utilized as a control for miRNA.20 The relative expression was calculated using the 2−ΔΔCt method. All primer sequences were listed as below:
Circ_0084927, F: 5ʹ-CTGGTGCAGCAAGATGGAAC-3ʹ and R: 5ʹ-CTGCACCTCCCTTGGCAATA-3ʹ; miR-142-3p, F: 5ʹ-ACACTCCAGCTGGGTGTAGTGTTTCCTACTTTA-3ʹ and R: 5ʹ-CTCAACTGGTGTCGTGGA-3ʹ; ARL2, F: 5ʹ-GAGCACCGCGGATTCAA-3ʹ and R: 5ʹ-GCAAAGATGAGGAGGGTTCG-3ʹ; GAPDH, F: 5ʹ-GAAGAGAGAGACCCTCACGCTG-3ʹ and R: 5ʹ-ACTGTGAGGAGGGGAGATTCAGT-3ʹ; U6, F: 5ʹ-CTCGCTTCGGCAGCACA-3ʹ and R: 5ʹ-AACGCTTCACGAATTTGCGT-3ʹ;
RNase R Treatment
The isolated RNA was exposed to 3 U·mg−1 RNase R (Epicentre, Madison, WI, USA) at 37°C for 30 min, and non-treatment was used as blank control. Then, qRT-PCR was performed to examine the expression of circ_0084927.
Cell Transfection
Small interference RNA targeting circ_0084927 (si-circ_0084927), including si-circ_0084927#1, si-circ_0084927#2 and si-circ_0084927#3, and its negative control (si-NC) were synthesized by Ribobio and used for circ_0084927 knockdown. Special overexpression vector for circRNA containing circ_0084927 sequence and its negative control (circ-NC) were purchased from BersinBio (Guangzhou, China) and used for circ_0084927 overexpression. MiR-142-3p mimics (miR-142-3p), miR-142-3p inhibitions (anti-miR-142-3p) and their controls (NC or anti-NC) were obtained from Ribobio. For ARL2 overexpression, ARL2 sequence cloned into pcDNA vector (ARL2) and its control (vector) were synthesized by BersinBio. The oligonucleotides or fusion plasmids were introduced into experimental cells using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA).
Cell Counting Kit-8 (CCK-8) Assay
Cells were planted into a 96-well plate (5 × 103 cells/well) with different transfections. At the indicated time points after transfection (24, 48, 72 and 96 h), 10 μL CCK-8 solution (Beyotime, Shanghai, China) was pipetted into every well for another 2 h. Afterwards, the absorbance at 450 nm was measured using a microplate reader (Bio-Rad, Hercules, CA, USA).
Cell Cycle Distribution
At 96 h post-transfection, cells (1 × 106) were treated with 0.25% trypsin and collected by centrifugation without EDTA after washing by phosphate-buffered saline (PBS). Cells were then rinsed by 5% bovine serum albumin (BSA) followed by the treatment of 70% pre-cooled alcohol and incubated at −20°C overnight. Next, cells were collected and resuspended in PBS with the addition of RNase A. After reacting for 20 min, propidium iodide (PI; Beyotime) was used to stain cells for 15 min at 4°C. Then, cell cycle distribution was analyzed using Attune™ NxT Acoustic Focusing Cytometer (Invitrogen).
Transwell Assay
Cell migration and cell invasion were monitored using the transwell chambers (BD Bioscience, San Jose, CA, USA). The transfected cells were resuspended in serum-free medium and added into the top of chambers for migration analysis or chambers pre-coated with Matrigel (BD Bioscience) for invasion analysis. Meanwhile, the bottom of the chambers was filled with the corresponding culture medium with 10% FBS. Allowing migration or invasion for 24 h, cells in the upper chambers were discarded, and cells in the lower chambers were fixed and stained with 0.1% crystal violet (Beyotime). Subsequently, cells were observed using a light microscope (100 × magnification; Olympus).
Western Blot
The extracted proteins were quantified by the BCA Protein Assay Kit (Beyotime) and electrophoresed by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The separated proteins were then transferred onto polyvinylidene difluoride membranes (PVDF; Bio-Rad) followed by blockage in the blocking buffer. Next, the primary antibodies and secondary antibodies were introduced to probe membranes, including anti-cyclin D1 (ab16663; Abcam, Cambridge, MA, USA), anti-matrix metallopeptidase 2 (anti-MMP2; ab97779), anti-matrix metallopeptidase 9 (anti-MMP9; ab76003), anti-ARL2 (ab109742), anti-proliferating cell nuclear antigen (anti-PCNA; ab92552), anti-GAPDH (ab9485) and goat anti-Rabbit IgG (ab205718). The protein signals were detected using an enhanced chemiluminescence kit (Beyotime).
Bioinformatics Analysis
We predicted the potential targets of circRNA or miRNA using the bioinformatics tools, including Circular RNA Interactome (https://circinteractome.nia.nih.gov/) and starBase (http://starbase.sysu.edu.cn/).
Dual-Luciferase Reporter Assay
Luciferase reporter plasmids were constructed using PGL4 vector (Promega, Madison, WI, USA). Briefly, the wild-type and mutant-type sequences of circ_0084927 and RAL2 were amplified and inserted into PGL4 vector to generate fusion plasmids, including circ_0084927-wt, circ_0084927-mut, ARL2-wt and ARL2-mut. The fusion plasmid was transfected into Hela and SiHa cells. Then, these transfected cells were reintroduced with miR-142-3p or NC and incubated for 48 h. Later, the luciferase activity was examined using the Dual-Luciferase Assay System (Promega).
RNA Immunoprecipitation Assay (RIP) Assay
Hela and SiHa cells were used to undergo RIP assay using the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore Corp, Billerica, MA, USA) with Argonaute 2 antibody (anti-Ago2) or Immunoglobulin G antibody (anti-IgG; control). RNAs bound to beats were eluted and quantified by qRT-PCR as previously mentioned.
Tumor Formation Assay in Nude Mice
The animal procedures obtained the approval of the Animal Care and Use Committee of the First Affiliated Hospital of Guangzhou Medical University. Animal studies were performed in compliance with the ARRIVE guidelines and the Basel Declaration. All animals received humane care according to the National Institutes of Health (USA) guidelines. A total of 12 BALB/C mice (Female, 4–6 weeks, 20 ± 2 g) were purchased from Guangdong Medical Laboratory Animal Center (Guangzhou, China). The short hairpin RNA (shRNA) targeting circ_0084927 (sh-circ_0084927) was packed into the lentiviral vector for stable transfection by Genepharma Company (Shanghai, China), and shRNA negative control (sh-NC) was used as a comparison. Hela cells (4 × 105) were stably transfected with sh-circ_0084927 or sh-NC and subcutaneously injected into BALB/C mice, with 6 mice in each group. After injection, the length and width of neoplasm were recorded once a week to monitor volume. After 5 weeks, all mice were sacrificed, and tumor nodes were weighed and used for further detection.
Statistical Analysis
All data were collected from three independent experiments. The survival analysis was performed by Kaplan–Meier plot and Log-rank test. The expression correlation analysis was conducted using Spearman correlation analysis. Results were processed using the GraphPad Prism 7.0 Software (GraphPad Prism; San Diego, CA, USA) and exhibited as the mean ± standard deviation. Difference analysis was performed using Student’s t-tests between two groups or using analyses of variance with Tukey post hoc test for multiple groups. P < 0.05 was deemed as statistically significant.
Result
Circ_0084927 Was Upregulated in Cervical Cancer Tumor Tissues and Cells Lines
We collected 50 pairs of tumor tissues and matched normal tissues, and the data from qRT-PCR suggested that the expression of circ_0084927 was higher in tumor tissues compared with that in normal tissues (Figure 1A). Besides, further analysis indicated that high circ_0084927 expression was associated with a relatively lower survival rate compared to low circ_0084927 expression (Figure 1B). In cell lines, the expression of circ_0084927 was notably elevated in Hela and SiHa cells compared to H8 cells (Figure 1C). Moreover, compared with linear GAPDH mRNA, circ_0084927 was more resistant to RNase R digestion in Hela and SiHa cells (Figure 1D). We speculated that the abnormal expression of circ_0084927 might affect cervical cancer progression.
 |
Figure 1 Circ_0084927 was highly expressed in cervical cancer tissues and cells. (A) The expression of circ_0084927 in tumor tissues (n=50) and normal tissues (n=50) was detected by qRT-PCR. (B) The survival curve of patients associated with circ_0084927 expression was generated by Kaplan–Meier plot and Log-rank test. (C) The expression of circ_0084927 in Hela, SiHa and H8 cells was detected by qRT-PCR. (D) The stability of circ_0084927 was examined using RNase R. *P<0.05. |
Circ_0084927 Knockdown Inhibited Cell Proliferation, Colony Formation, Cell Cycle and Migration/Invasion of Cervical Cancer in vitro
A total of three siRNAs specially targeting circ_0084927 were obtained to knock down the expression of circ_0084927, and si-circ_0084927#1 was chosen for further experiments based on its higher knockdown efficiency relative to si-circ_0084927#2 and si-circ_0084927#3 (Figure 2A). Then, Hela and SiHa cells were transfected with si-circ_0084927#1 or si-NC to monitor the function of circ_0084927. CCK-8 assay suggested that circ_0084927 knockdown remarkably restrained cell proliferation (Figure 2B). In addition, circ_0086927 knockdown induced cell cycle arrest at the G1-S phase, thus blocking cell growth (Figure 2C). Moreover, transwell assay presented that circ_0086927 knockdown significantly suppressed cell migration and invasion (Figure 2D and E). Several representative proteins, including cyclin D1, MMP2 and MMP9, were quantified by Western blot, and the data showed that all of them were signally downregulated in Hela and SiHa cells with circ_0086927 knockdown (Figure 2F and G). These functional analyses exposed that circ_0086927 knockdown restricted the aggravation of cervical cancer cells.
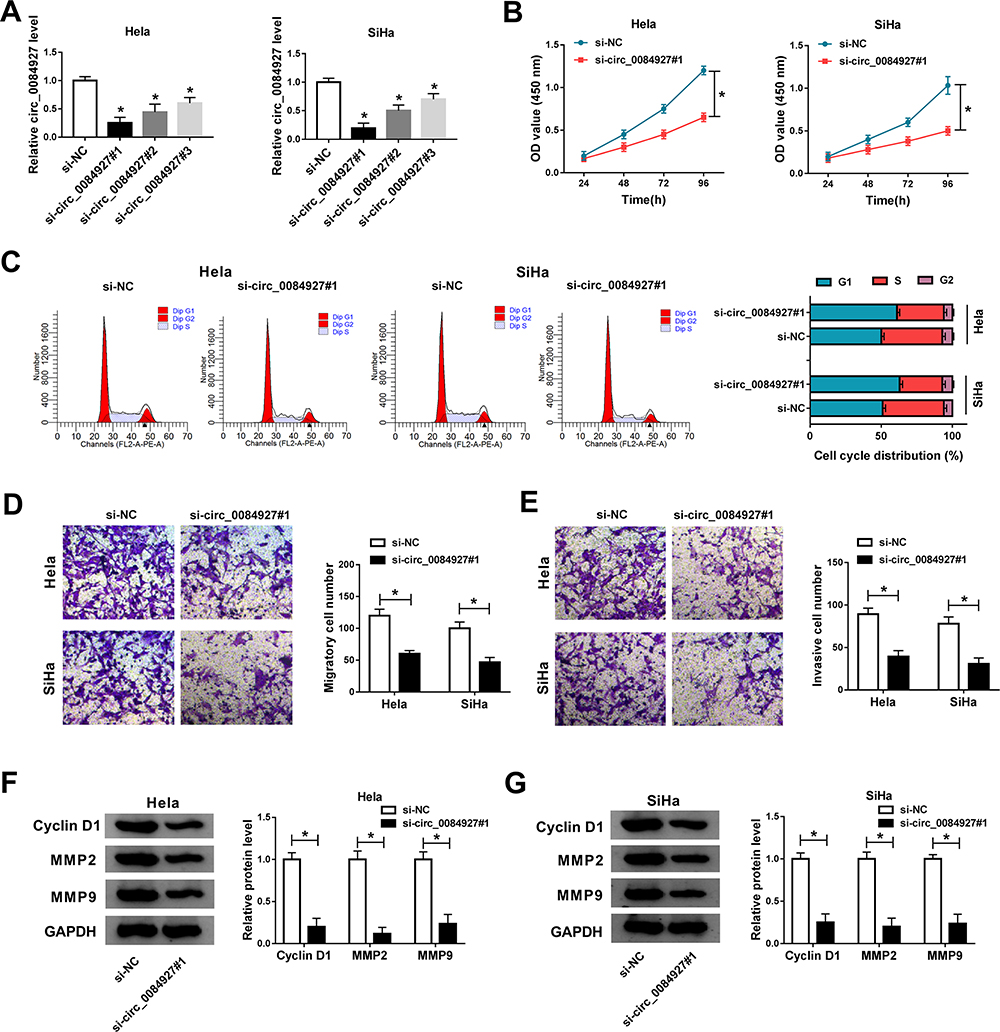 |
Figure 2 Circ_0084927 knockdown suppressed cervical cancer cell proliferation, cycle, migration and invasion. (A) The efficiency of circ_0084927 knockdown was examined by qRT-PCR. In si-circ_0084927#1-transfected Hela and SiHa cells, (B) cell proliferation was assessed by CCK-8 assay. (C) Cell cycle distribution was checked by flow cytometry assay. (D and E) Cell migration and invasion were monitored by transwell assay. (F and G) The expression of cyclin D1, MMP2 and MMP9 was examined by Western blot. *P<0.05. |
Circ_0086927 Directly Targeted miR-142-3p That Was Downregulated in Cervical Cancer Tissues and Cells
Based on the hypothesis that circ_0086927 functioned in cervical cancer through the classical ceRNA mechanism, the target miRNAs of circ_0086927 were analyzed. By the prediction of the database “Circular RNA Interactome”, we found circ_0086927 could bind to miR-142-3p in a special binding site, and the sequence in binding site was mutated to generate a mutant sequence of circ_0086927 for further analysis (Figure 3A). As verifications, miR-142-3p reintroduction strikingly lessened the luciferase activity of Hela and SiHa cells transfected with circ_0084927-wt but not circ_0084927-mut by the dual-luciferase reporter assay (Figure 3B). Besides, RIP assay proved that both circ_0084927 and miR-142-3p were abundantly captured in the Anti-Ago2 group relative to that in the Anti-IgG group (Figure 3C). Moreover, the expression of miR-142-3p was negatively regulated by circ_0084927 in Hela and SiHa cells (Figure 3D). These results verified that miR-142-3p was a target of circ_0084927, and circ_0084927 suppressed miR-142-3p expression. In addition, the data from qRT-PCR indicated that the expression of miR-142-3p was markedly declined in cervical cancer tissues (n=50) and cell lines (Figure 3E and F), and circ_0084927 expression was negatively correlated with miR-142-3p expression in tumor tissues (Figure 3G). The data suggested that miR-142-3p was lowly expressed in cervical cancer.
 |
Figure 3 MiR-142-3p was a target of circ_0084927. (A) MiR-142-3p was predicted as a target of circ_0084927 by the bioinformatics tool “Circular RNA Interactome”. (B and C) MiR-142-3p was validated as a target of circ_0084927 by dual-luciferase reporter assay and RIP assay. (D) The effect of circ_0084927 overexpression or knockdown on the expression of miR-142-3p was detected by qRT-PCR. (E and F) The expression of miR-142-3p in cervical cancer tissues and cells was measured by qRT-PCR. (G) The correlation between circ_0084927 expression and miR-142-3p expression was analyzed by Spearman correlation analysis. *P<0.05. |
miR-142-3p Inhibition Blocked the Role of circ_0084927 Knockdown
Subsequently, we examined the interaction between circ_0084927 and miR-142-3p in function. The expression of miR-142-3p promoted in Hela and SiHa cells transfected with si-circ_0084927 was repressed in cells transfected with si-circ_0084927#1+anti-miR-142-3p, suggesting that miR-142-3p was downregulated (Figure 4A). The capacities of cell proliferation blocked by circ_0084927 knockdown were recovered by the reintroduction of anti-miR-412-3p (Figure 4B). Besides, circ_0084927 knockdown-induced cell cycle arrest at the G1-S phase was relieved by the reintroduction of anti-miR-412-3p (Figure 4C). The activities of cell migration and invasion were inhibited in cells transfected with si-circ_0084927#1 but largely regained in cells transfected with si-circ_0084927#1+anti-miR-142-3p (Figure 4D and E). Additionally, the expression levels of cyclin D1, MMP2 and MMP9 were reduced in cells transfected with si-circ_0084927#1 but restored in cells with si-circ_0084927#1+anti-miR-142-3p transfection (Figure 4F and G). It could be concluded that circ_0084927 knockdown restricted the aggravation of cervical cancer cells by upregulating miR-142-3p.
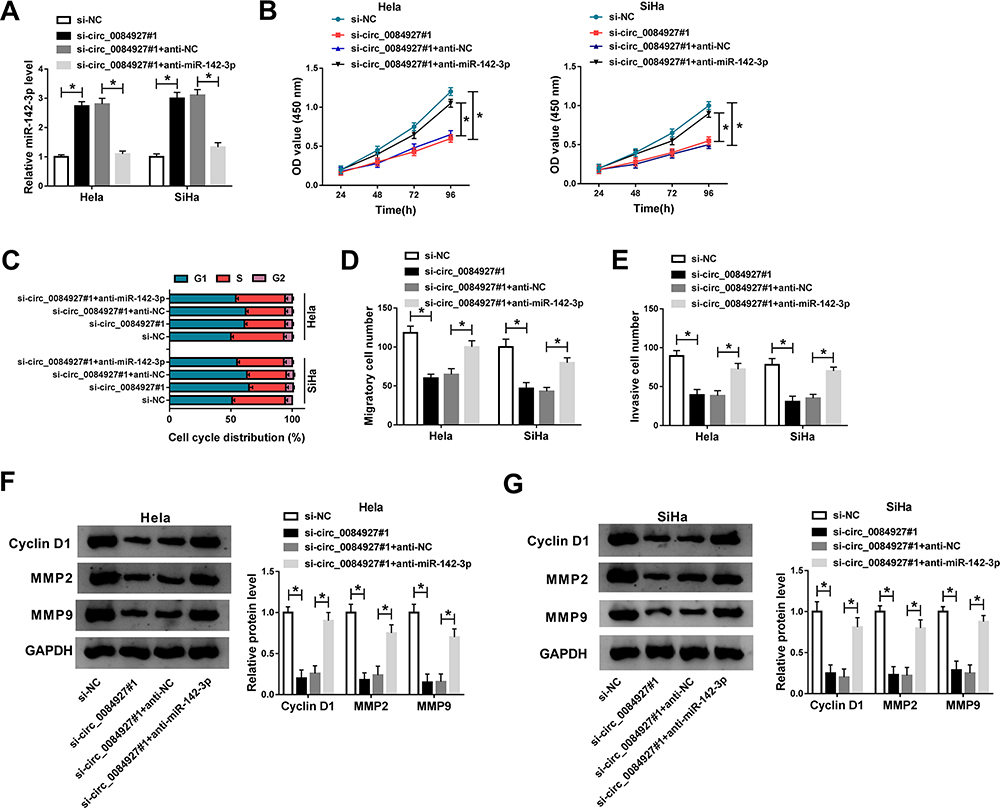 |
Figure 4 MiR-142-3p inhibition reversed the effects of circ_0084927 knockdown in cervical cancer cells. Hela and SiHa cells were introduced with si-circ_0084927#1, si-NC, circ_0084927#1+anti-miR-142-3p or si-circ_0084927#1+anti-NC. (A) The expression of miR-142-3p was examined using qRT-PCR. (B) Cell proliferation was evaluated by CCK-8 assay. (C) Cell cycle distribution was monitored by flow cytometry assay. (D and E) Cell migration and cell invasion were investigated by transwell assay. (F and G) The expression of cyclin D1, MMP2 and MMP9 was detected by Western blot. *P<0.05. |
miR-142-3p Bound to ARL2, and ARL2 Was Upregulated in Cervical Cancer Tissues and Cells
Further study analyzed the downstream genes targeted by miR-142-3p, and ARL2 was a putative target mRNA of miR-142-3p with a special binding site between its 3ʹ-untranslated region (3ʹUTR) and miR-142-3p by the analysis of starBase (Figure 5A). Moreover, miR-142-3p reintroduction significantly impaired the luciferase activity of Hela and SiHa cells transfected with ARL2-wt but not ARL2-mut (Figure 5B), and miR-142-3p and ARL2 could be enriched in the Anti-Ago2 RIP group compared to the Anti-IgG RIP group (Figure 5C). More crucially, ARL2 expression was inhibited by miR-142-3p restoration, while circ_0084927 reintroduction recovered the expression of ARL2 (Figure 5D), suggesting that ARL2 expression was regulated by circ_0084927 through miR-142-3p. In addition, ARL2 was prominently upregulated in cervical cancer tissues (n=50) compared to normal tissues (n=50) detected by qRT-PCR and Western blot (Figure 5E and F). ARL2 expression was also elevated in Hela and SiHa cells relative to that in H8 cells (Figure 5G). Besides, ARL2 expression was negatively correlated with miR-142-3p expression, and circ_00084927 expression was positively correlated with ARL2 expression in tumor tissues (Figure 5H and I). These data summarized that ARL2 was a target of miR-142-3p, and ARL2 was aberrantly upregulated in cervical cancer.
 |
Figure 5 ARL2 was a target of miR-142-3p. (A) ARL2 was predicted to be a target of miR-142-3p by the bioinformatics tool “starBase”. (B and C) ARL2 was verified to be a target of miR-142-3p by dual-luciferase reporter assay and RIP assay. (D) The expression of ARL2 in Hela and SiHa cells transfected with miR-142-3p, NC, circ_0084927+miR-142-3p or circ-NC+miR-142-3p was detected by Western blot. (E and F) The expression of ARL2 in cervical tissues (n=50) and normal tissues (n=50) was detected by qRT-PCR or Western blot. (G) The expression of ARL2 in H8, Hela and SiHa cells was detected by Western blot. (H and I) The correlation between ARL2 expression and miR-142-3p expression or circ_0084927 expression was determined by Spearman correlation analysis. *P<0.05. |
miR-142-3p Restoration-Suppressed Cell Proliferation, Cycle, Migration and Invasion Were Recovered by ARL2 Reintroduction
Hela and SiHa cells were transfected with miR-142-3p, NC, miR-142-3p+ARL2 or miR-142-3p+vector to monitor the interaction between miR-142-3p and ARL2 in function. The expression of ARL2 was lessened in cells transfected with miR-142-3p but substantially promoted in cells transfected with miR-142-3p+ARL2 (Figure 6A). Cell proliferation ability was sequestered in cells transfected with miR-142-3p but stimulated in cells transfected with miR-142-3p+ARL2 (Figure 6B). Besides, miR-142-3p restoration induced cell cycle arrest, while ARL2 reintroduction partly ameliorated cell cycle arrest (Figure 6C). Moreover, miR-142-3p enrichment strikingly blocked cell migration and cell invasion, while ARL2 reintroduction enhanced migration and invasion (Figure 6D and E). Additionally, the expression levels of cyclin D1, MMP2 and MMP9 were weakened by miR-142-3p transfection but restored by miR-142-3p+ARL2 transfection in Hela and SiHa cells (Figure 6F and G). It could be deduced that miR-142-3p blocked the malignant development of cervical cancer by downregulating ARL2.
 |
Figure 6 ARL2 overexpression partly abolished the antitumor role of miR-142-3p restoration in cervical cancer cells. Hela and SiHa cells transfected with miR-142-3p, NC, miR-142-3p+ARL2 or miR-142-3p+vector were used for further analyses. (A) The expression of ARL2 in these transfected cells was measured by Western blot. (B) Cell proliferation was assessed by CCK-8 assay. (C) Cell cycle distribution was observed by flow cytometry assay. (D and E) Cell migration and invasion were determined by transwell assay. (F and G) The expression of cyclin D1, MMP2 and MMP9 was quantified by Western blot. *P<0.05. |
Circ_0084927 Affected Tumor Growth in vivo by Regulating miR-142-3p and ARL2
The subsequent study explored the role of circ_0084927 on tumor growth in nude mice. Hela cells transfected with sh-circ_0084927 or sh-NC were injected into the subcutaneous part of nude mice. After injection, we found that both tumor volume and tumor weight were significantly lower in the sh-circ_0084927 group compared with that in the sh-NC group (Figure 7A and B). Using the removed tumor tissues, we examined the expression of circ_0084927, miR-142-3p and ARL2. The expression of circ_0084927 was decreased while the expression of miR-142-3p was increased in tumor tissues from the sh-circ_0084927 group relative to the sh-NC group (Figure 7C). Not surprisingly, the expression of ARL2 was declined in the sh-circ_0084927 group but not in the sh-NC group (Figure 7D). Moreover, we checked the expression of a proliferation-related marker (PCNA) and found that PCNA was also downregulated in the sh-circ_0084927 group compared to that in the sh-NC group (Figure 7D), suggesting that tumor growth was suppressed, and circ_0084927 also affected tumor growth in vivo.
 |
Figure 7 Circ_0084927 regulated tumor growth in vivo by miR-142-3p and ARL2. Hela cells with sh-circ_0084927 or sh-NC transfection were injected into nude mice. (A) Tumor volume was recorded once a week after injection. (B) Tumor was weighted after removing from body after 5 weeks. (C) The expression of circ_0084927 and miR-142-3p in excised tissues was detected by qRT-PCR. (D) The expression of ARL2 and PCNA in excised tissues was monitored by Western blot. *P<0.05. |
Discussion
Statistics on the incidence and mortality of cervical cancer suggest that new strategies of diagnosis and treatment for cervical cancer are necessary. Identifying effective biomarkers and targets should help improve the specificity and sensitivity of diagnostic/therapeutic tools. Although gene expression profiling has made immense progress at the molecular level of cervical cancer, there are currently not enough screening and treatment strategies.21 In our study, we concerned a poorly investigated circRNA, circ_0084927, which was abnormally upregulated in cervical cancer tissues from the GEO database. Our functional experiments demonstrated that circ_0084927 knockdown attenuated the malignant development of cervical cancer, hinting that circ_0084927 might be an indicator of cancer diagnosis and treatment.
Presently, the function of numerous circRNAs remains unknown. Some specially expressed circRNAs have been functionally investigated in cervical cancer. For example, circRNA8924 expression was elevated in cervical cancer tissues and linked to tumor size, staging and invasion, and circRNA8924 knockdown restrained cell proliferation and metastasis in vitro.22 Similarly, both circ_000515 and circCLK3 were upregulated in cervical cancer tissues, and their silencing contributed to the inhibition of cell growth, migration and invasion.10,23 Consistent with these consequences, we validated that the expression level of circ_0084927 was strikingly enhanced in clinical cervical tissues. Besides, in vitro and in vivo assays demonstrated that circ_0084927 downregulation blocked cell proliferation, cycle, migration and invasion as well as tumor formation and growth. A previous study mentioned that the expression of circ_0084927 was increased in lung cancer-related malignant pleural effusion compared to tuberculous pleural effusion by the analysis of microarray expression profile and qRT-PCR,24 suggesting that circ_0084927 participated in lung cancer-associated complication. We speculated that circ_0084927 was a wide regulator in various cancers, and our data for the first time explored its function in cervical cancer.
Mechanically, we found that circ_0084927 competitively sponged miR-142-3p. The deficiency of miR-142-3p reversed the effects of circ_0084927 knockdown, leading to malignant activities. In addition, miR-142-3p restoration alone exerted the antitumor role in cervical cancer cell development. Throughout the previous research, miR-142-3p was recorded to be downregulated in cervical cancer tissues, and low miR-142-3p expression was related to lymph node metastasis and poor survival rate of patients with cervical cancer.25 Besides, miR-142-3p played anti-proliferation and anti-invasion properties in cervical cancer cells.16 Metformin could promote the abundance of miR-142-3p to block cervical cancer cell migration and invasion.26 Our view was consistent with these findings, supporting that miR-142-3p was a tumor suppressor in cervical cancer.
MiRNAs are well known to bind to mRNA at the 3ʹ UTR, leading to the expression downregulation and function degradation of protein-coding genes.13 In this study, miR-142-3p directly bound to ARL2 3ʹ UTR and negatively regulated the expression of ARL2. The inhibitory role of miR-142-3p on cell proliferation, migration and invasion was abolished by ARL2 overexpression. ARL2 was documented to be targeted by certain miRNAs and involved in cervical cancer development. For instance, ARL2 was a target of miR-214 and miR-195-5p, and miR-214 and miR-195-5p repressed the malignant cellular behaviors in cervical cancer by downregulating ARL2,27 implying that ARL2 was an oncogene in cervical cancer. Moreover, we determined that circ_0084927 positively modulated the expression of ARL2 by suppressing miR-142-3p. Hence, the circ_0084927/miR-142-3p/ARL2 regulatory axis was generated.
Overall, circ_0084927 knockdown blocked cervical cancer cell growth and metastasis in vitro as well as tumor growth in vivo. As a potential mechanism, circ_0084927 played its functions in cervical cancer progression by mediating the miR-142-3p/ARL2 axis. The circ_0084927/miR-142-3p/ARL2 regulatory axis might be a promising strategy to antagonize cervical cancer through the targeted inhibition of circ_0084927 and ARL2 or the promotion of miR-142-3p.
Disclosure of Interest
The authors report no conflicts of interest in this work.
Funding
There is no funding to report.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34. doi:10.3322/caac.21551
2. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
3. Biewenga P, van der Velden J, Mol BW, et al. Prognostic model for survival in patients with early stage cervical cancer. Cancer. 2011;117(4):768–776. doi:10.1002/cncr.25658
4. Sankaranarayanan R, Swaminathan R, Brenner H, et al. Cancer survival in Africa, Asia, and Central America: a population-based study. Lancet Oncol. 2010;11(2):165–173. doi:10.1016/S1470-2045(09)70335-3
5. Gao YL, Zhang MY, Xu B, et al. Circular RNA expression profiles reveal that hsa_circ_0018289 is up-regulated in cervical cancer and promotes the tumorigenesis. Oncotarget. 2017;8(49):86625–86633. doi:10.18632/oncotarget.21257
6. Quan G, Li J. Circular RNAs: biogenesis, expression and their potential roles in reproduction. J Ovarian Res. 2018;11(1):9. doi:10.1186/s13048-018-0381-4
7. Suzuki H, Tsukahara T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int J Mol Sci. 2014;15(6):9331–9342. doi:10.3390/ijms15069331
8. Qu S, Yang X, Li X, et al. Circular RNA: a new star of noncoding RNAs. Cancer Lett. 2015;365(2):141–148. doi:10.1016/j.canlet.2015.06.003
9. Ma HB, Yao YN, Yu JJ, Chen XX, Li HF. Extensive profiling of circular RNAs and the potential regulatory role of circRNA-000284 in cell proliferation and invasion of cervical cancer via sponging miR-506. Am J Transl Res. 2018;10(2):592–604.
10. Hong H, Zhu H, Zhao S, et al. The novel circCLK3/miR-320a/FoxM1 axis promotes cervical cancer progression. Cell Death Dis. 2019;10(12):950. doi:10.1038/s41419-019-2183-z
11. Cai H, Zhang P, Xu M, Yan L, Liu N, Wu X. Circular RNA hsa_circ_0000263 participates in cervical cancer development by regulating target gene of miR-150-5p. J Cell Physiol. 2019;234(7):11391–11400. doi:10.1002/jcp.27796
12. Yang C, Wu D, Gao L, et al. Competing endogenous RNA networks in human cancer: hypothesis, validation, and perspectives. Oncotarget. 2016;7(12):13479–13490. doi:10.18632/oncotarget.7266
13. Berindan-Neagoe I, Monroig Pdel C, Pasculli B, Calin GA. MicroRNAome genome: a treasure for cancer diagnosis and therapy. CA Cancer J Clin. 2014;64(5):311–336. doi:10.3322/caac.21244
14. Deftereos G, Corrie SR, Feng Q, et al. Expression of mir-21 and mir-143 in cervical specimens ranging from histologically normal through to invasive cervical cancer. PLoS One. 2011;6(12):e28423. doi:10.1371/journal.pone.0028423
15. Shishodia G, Shukla S, Srivastava Y, et al. Alterations in microRNAs miR-21 and let-7a correlate with aberrant STAT3 signaling and downstream effects during cervical carcinogenesis. Mol Cancer. 2015;14(1):116. doi:10.1186/s12943-015-0385-2
16. Deng B, Zhang Y, Zhang S, Wen F, Miao Y, Guo K. MicroRNA-142-3p inhibits cell proliferation and invasion of cervical cancer cells by targeting FZD7. Tumour Biol. 2015;36(10):8065–8073. doi:10.1007/s13277-015-3483-2
17. Wang Y, Guan G, Cheng W, et al. ARL2 overexpression inhibits glioma proliferation and tumorigenicity via down-regulating AXL. BMC Cancer. 2018;18(1):599. doi:10.1186/s12885-018-4517-0
18. Matozaki T, Nakanishi H, Takai Y. Small G-protein networks: their crosstalk and signal cascades. Cell Signal. 2000;12(8):515–524. doi:10.1016/s0898-6568(00)00102-9
19. Beghin A, Belin S, Hage-Sleiman R, et al. ADP ribosylation factor like 2 (Arl2) regulates breast tumor aggressivity in immunodeficient mice. PLoS One. 2009;4(10):e7478. doi:10.1371/journal.pone.0007478
20. Kori M, Yalcin Arga K. Potential biomarkers and therapeutic targets in cervical cancer: insights from the meta-analysis of transcriptomics data within network biomedicine perspective. PLoS One. 2018;13(7):e0200717. doi:10.1371/journal.pone.0200717
21. Liu J, Wang D, Long Z, Liu J, Li W. CircRNA8924 promotescervical cancer cell proliferation, migration and invasion by competively binding to MiR-518d-5p/519-5p family and modulating theexpression of CBX8. Cell Physiol Biochem. 2018;48(1):173–184. doi:10.1159/000491716
22. Tang Q, Chen Z, Zhao L, Xu H. Circular RNA hsa_circ_0000515 acts as a miR-326 sponge to promote cervical cancer progression through up-regulation of ELK1. Aging (Albany NY). 2019;11(22):9982–9999. doi:10.18632/aging.10235623
23. Wen Y, Wang Y, Xing Z, Liu Z, Hou Z. Microarray expression profile and analysis of circular RNA regulatory network in malignant pleural effusion. Cell Cycle. 2018;17(24):2819–2832. doi:10.1080/153841\01.2018.1558860
24. Li M, Li BY, Xia H, Jiang LL. Expression of microRNA-142-3p in cervical cancer and its correlation with prognosis. Eur Rev Med Pharmacol Sci. 2017;21(10):2346–2350.
25. Xia C, Liang S, He Z, Zhu X, Chen R, Chen J. Metformin, a first-line drug for type 2 diabetes mellitus, disrupts the MALAT1/miR-142-3p sponge to decrease invasion and migration in cervical cancer cells. Eur J Pharmacol. 2018;830:59–67. doi:10.1016/j.ejphar.2018.04.027
26. Peng R, Men J, Ma R, et al. miR-214 down-regulates ARL2 and suppresses growth and invasion of cervical cancer cells. Biochem Biophys Res Commun. 2017;484(3):623–630. doi:10.1016/j.bbrc.2017.01.152
27. Pan SS, Zhou HE, Yu HY, Xu LH. MiR-195-5p inhibits the cell migration and invasion of cervical carcinoma through suppressing ARL2. Eur Rev Med Pharmacol Sci. 2019;23(24):10664–10671. doi:10.26355/eurrev_201912_19764
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.