Back to Journals » Cancer Management and Research » Volume 13
Circ_0006174 Accelerates Colorectal Cancer Progression Through Regulating miR-138-5p/MACC1 Axis
Authors Wei J, Lin Y, Wang Z, Liu Y, Guo W
Received 4 December 2020
Accepted for publication 21 January 2021
Published 18 February 2021 Volume 2021:13 Pages 1673—1686
DOI https://doi.org/10.2147/CMAR.S295833
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Chien-Feng Li
Jianjun Wei,* Yuzhe Lin,* Zhiqiang Wang, Yeguang Liu, Wei Guo
Minimally Invasive Surgery, Linhe People’s Hospital of Bayannur City, Bayannur, Inner Mongolia, 015000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiqiang Wang Tel +86-04788253451
Email [email protected]
Background: Circular RNAs (circRNAs) were reported to be involved in the progression of a variety of cancers, including colorectal cancer (CRC). However, the precise functions and mechanism of circRNAs in CRC have not been elucidated. This study aimed to investigate the effect and mechanism underlying circ_0006174 in CRC.
Methods: The expression of circ_0006174, microRNA (miR-138-5p) and metastasis associated in colon cancer 1 (MACC1) mRNA was detected by quantitative real-time polymerase chain reaction (RT-qPCR) assay. Western blot was employed to measure MACC1 protein expression. The effects of circ_0006174 knockdown, MACC1 overexpression or miR-138-5p inhibition on cell proliferation, migration, invasion, and apoptosis were assessed by cell counting kit 8 (CCK-8) assay, clone formation assay, transwell assay and flow cytometry assay, respectively. The interaction between miR-138-5p and circ_0006174 or MACC1 was confirmed by RNA pull down assay or dual-luciferase reporter assay. Xenograft tumor model in nude mice was used to verify the function of circ_0006174 in vivo.
Results: Circ_0006174 and MACC1 expression was highly expressed, while miR-138-5p expression was downregulated in CRC cells and tissues. Meanwhile, circ_0006174 functioned as a sponge of miR-138-5p to upregulate MACC1 expression. Furthermore, circ_0006174 knock down-mediated suppression on cell proliferation, migration and invasion, and promotion on cell apoptosis could be alleviated by MACC1 overexpression or miR-138-5p inhibition in CRC cells. Besides, circ_0006174 knockdown also inhibited CRC procession in vivo.
Conclusion: Circ_0006174 advanced CRC progression via sponging miR-138-5p to upregulate MACC1 expression, which may provide a promising molecular target for CRC treatment.
Keywords: colorectal cancer, circ_0006174, miR-138-5p, MACC1
Introduction
According to cancer statistics in China in 2015, colorectal cancer (CRC) ranked sixth among all cancers in terms of incidence and mortality.1 Adenoma carcinoma sequence gives a detailed account of the CRC natural history, which hypothesized that CRC was originated from precancerous lesions and gradually acquired additional mutations that eventually lead to malignant carcinogenesis.2 It takes decades for precancerous lesions to develop into CRC,3 and genetic mutations are the decisive factors for driving this process.4 The relationship between development of adenoma to cancer and mutated genes in the process has been reported.5 In CRC, three basic pathways were involved in widespread genomic instability: 1) chromosomal instability, 2) microsatellite instability, and 3) CpG island methylate phenotype (CIMP) pathways.6 Thus, it is appealing to illustrate the underlying molecular pathogenesis of CRC to provide evidence for CRC treatment.
Circular RNAs (circRNAs) were considered as the byproducts of improper RNA splicing or transcriptional garbage for a long time, due to their low expression levels and the limitation of molecular biological technique.7 According to different cycling mechanisms, circRNAs can be divided into three types including exonic circRNAs, intronic circRNAs (ciRNAs) and retained-intron circRNAs (ElciRNAs).8 Exonic circRNAs were the most studied circRNAs that were primarily localized in cytoplasm and associated with microRNAs (miRNAs) or RNA-binding proteins (RBPs).9,10 As a kind of non-coding RNA, the multiple functions of circRNA have been illustrated in amount of studies. CircRNAs have been recognized as sponges of miRNAs and RBPs,11 which contributed to translating peptides12 and regulated post-transcription13 in cancers. The most studied feature of circRNAs was that circRNAs played a pivotal role in various cancer processes through acting as sponges of miRNAs to regulate mRNAs expression, finally raising up the effects on the proliferation and metastasis in cancer cells.14,15 In CRC, the dysregulation of circRNAs has been found to be associated with cancer progression.16 Zheng et al revealed that circHIPK3 was upregulated in CRC tissues contrasted with the corresponding normal tissues, and harbored miR-124 to modulate cell proliferation.15 Zhu et al uncovered that circ-BANP was markedly upregulated and circ-BANP silencing suppressed the proliferation in CRC cells.17 Xie et al proposed that circ_001569 was markedly upregulated in CRC tissues, which was positively associated with CRC malignant characteristics including the promotion in proliferation and metastasis. Besides, high-throughput microarray studies have uncovered that circ_0006174 was high expressed in CRC tissues in contrast with the normal tissues based on the data from Gene Expression Omnibus databases (GEO, NIH, GSE142837, GSE138589, and GSE126094). However, the role of circ_0006174 in CRC progression remained unknown.
In this study, we revealed the regulation mechanism underlying circ_0006174 in CRC. The expression of circ_0006174 was increased in CRC tissues and cells. Besides, circ_0006174 served as a molecular sponge for miR-138-5p, and miR-138-5p inhibition attenuated the decreasing proliferation, metastasis, and the elevated apoptosis in CRC cells that were induced by circ_0006174 knockdown. Meanwhile, metastasis associated in colon cancer 1 (MACC1) was predicted as the downstream target of miR-138-5p. Herein, our data displayed that circ_0006174 functioned as a tumor-promoter in CRC progression, which might provide a novel biomarker for CRC therapy.
Materials and Methods
Clinical Samples
This study was conducted in accordance with the Declaration of Helsinki and was approved by the Clinical Research Ethics Committee of Linhe people’s Hospital of Bayannur City. Written informed consent was obtained from all patients prior to this study. CRC cancer tissues and corresponding adjacent normal tissues were obtained from 40 patients (18 male, 22 female) during operation at Linhe people’s Hospital of Bayannur City and then frozen in liquid nitrogen immediately. The inclusion criteria were as follows: (1) Cancer was confirmed by pathologic diagnosis, and the patients volunteered to participate in the study. (2) None of these patients received preoperative chemotherapy or radiotherapy. The exclusion criteria were as follows: (1) Patients with other intestinal diseases, such as ulcerative colitis and Crohn’s disease. (2) Patients with another primary cancer. (3) Patients with known primary organ failure. The patient characteristics were as follows: Mean age of the patients at diagnosis was 54 ± 11.4 years (range 29–78 years). Forty patients including 28 patients with stage I–II CRC and 12 patients with stage III, and 23 had no local lymph node metastasis (N0) while 17 patients were found to have lymph node metastasis (N1-2).
Cell Lines and Culture
Human CRC cell lines (SW480, SW620, HT29 and HCT116) were obtained from American Type Culture Collection (ATCC; Manassas, VA, USA) and NCM460, the normal human colon mucosal epithelial cell line was obtained from INCELL Corporation LLC (San Antonio, TX, USA). Dulbecco’s modified Eagle’s medium (DMEM; HyClone Laboratories, Logan, UT, USA) containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA, USA), 100 IU/mL penicillin (Sigma-Aldrich, St. Louis, MO, USA) and 100 µg/mL streptomycin (Sigma-Aldrich) were applied to culture cells in an incubator containing 5% CO2 at 37°C.
RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
All tissues and cells were treated with TRIzol® reagent (Thermo Fisher Scientific) for total RNA isolation following the manufacturer’s instructions. RNA quality and concentration were determined by NanoDrop 2000 (Thermo Fisher Scientific, Rockford, IL, USA). Then, SuperScript IV VILO kit (Invitrogen, Carlsbad, CA, USA) was employed to synthesize cDNA, while for miRNA analysis, All-in-One™ First-Strand cDNA Synthesis Kit (GeneCopoeia, Guangzhou, China) was used. The ABI SYBR Green Master Mix (Invitrogen) was used for RT-qPCR. The 2−ΔΔCT method was applied to calculate relative expression of circ_0006174, MACC1 mRNA and miR-138-5p. The primers sequences were shown as circ_0006174, forward: 5ʹ-CATCCATCACTCCAGCATCAG-3ʹ, reverse: 5ʹ-GGTCACCATAACCACCACAAAG-3ʹ; miR-138-5p, forward: 5ʹ-GCGAGCTGGTGTTGTGAATC-3ʹ, reverse: 5ʹ-AGTGCAGGGTCCGAGGTATT-3ʹ; MACC1, forward: 5ʹ-TTCTTTTGATTCCTCCGGTGA-3ʹ, reverse: 5ʹ-ACTCTGATGGGCATGTGCTG-3ʹ; glyceraldehyde 3-phosphate dehydrogenase (GAPDH), forward: 5ʹ-GGAGGGAGATCCCTCCAAAAT-3ʹ, reverse: 5ʹ-GGCTGTTGTCATACTTCTCATGG-3ʹ; U6 small nuclear RNA (U6), forward: 5ʹ-CTCGCTTCGGCAGCACA-3ʹ, reverse: 5ʹ-AACGCTTCACGAATTTGCGT-3ʹ, 18S ribosomal ribonucleic acid (18S rRNA), forward: 5ʹ-GAGTGTAAGGACCCATCGGA-3ʹ, reverse: 5ʹ-CCTCCAATGGATCCTCGTTA-3ʹ. 18S rRNA and U6 were used as internal controls of circ_0006174. GAPDH and U6 were used as internal controls of MACC1 and miR-138-5p, respectively.
Transient Transfection
Short hairpin RNA (shRNA) stably against circ_0006174 (sh-circ #1, sh-circ #2, sh-circ #3), shRNA negative control (sh-NC), miR-138-5p mimic (miR-138-5p), miR-138-5p negative control (miR-NC), miR-138-5p inhibitor (anti-miR-138-5p), anti-miR-138-5p negative control (anti-NC), MACC1 overexpression (oe-MACC1), oe-MACC1 negative control (Vector) were obtained from RiboBio (Guangzhou, China). Transient transfection was performed by Lipofectamine 2000 (Invitrogen). All vectors or oligonucleotides were transfected into cells by Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). Transfected cells were cultured in an incubator at 37°C. Upon transfection for 24, 48, or 72 hours, the cells were collected for further research.
Cell Counting Kit 8 (CCK-8) Assay
CCK-8 assay was employed to access cell proliferation. HT29 and SW620 cells (5 × 103) upon transfection were cultured in 96-well plates at 37°C for 24 h, 48 h, or 72 h. Then, 10 μL of CCK-8 solution (Beyotime, Shanghai, China) was added into each well and incubated at 37°C for 4 h. The absorbance at 450 nm was measured by a microplate reader (Tecan Sunrise, Mannedorf, Switzerland) at the required cultivation period.
Clone Formation Assay
Cells were cultured in 6-well plates with an approximate quantity of 1×103 cells in each well. Ten days later, 4% paraformaldehyde was added into each well to incubate for 30 min, then cells were stained with 1% crystal violet solution for 10 min. The number of colonies was counted using a microplate reader (Tecan Sunrise).
Transwell Assay
Both cell migration and invasion were assessed by transwell assay. HT29 and SW620 cells cultured in serum-free medium were plated into upper chambers (Corning, Corning, NY, USA) that were coated with (invasion assay) or without (migration assay) 8 μM matrigel mix (BD Biosciences, San Jose, CA, USA) at a density of 2×105 cells/insert according to the manufacturer’s instructions. The bottom chambers were added with fresh medium together with 10% FBS (Thermo Fisher Scientific). After incubation for 24 h, the cells at the upper surfaces of the transwell chambers were scraped with cotton swab, and the cells at the lower surfaces of the transwell chambers were fixed with methanol for 10 min, followed by stained with 0.1% crystal violet (Corning) for 20 min. Then, the stained cells were photographed and counted in three randomly selected fields by a microscope with the magnification of 100.
Flow Cytometry Assay
Apoptosis was assessed by AnnexinV-fluorescein isothiocyanate (AnnexinV-FITC)/propidium iodide (PI) kit (BD Pharmingen, San Diego, CA, USA) according to the manufacturer’s instruction. In brief, 1 × 106 HT29 and SW620 cells upon transfection were collected and stained with 5 µL PI and FITC for 15 min in the darkness. After washing with phosphate buffer saline (PBS, Sigma-Aldrich) 3 times, the cell apoptosis rate was analyzed by flow cytometry (BD FACS Aria; BD Biosciences).
RNA Immunoprecipitation (RIP)
The correlation between miRNAs and circ_0006174 was estimated by Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore, Bedford, MA, USA). When miRNAs form the binding of RISC complex, it was mainly encapsulated by Ago2 protein. Thus, the binding miRNA and the RISC-bound circRNA will also be pulled down when incubated with Anti-Ago2 antibody.18 IgG antibody was used as a negative control. Firstly, lentivirus vectors containing sh-NC or sh-circ #1 were constructed and transfected into HT29 and SW620 cells. Then, 1× 107 cells were lysed in RIP Lysis Buffer (Thermo Fisher Scientific) and incubated with magnetic beads conjugated with anti-IgG (1:1, 000, ab48386, Abcam, Cambridge, UK) or anti-Ago2 (1:300, ab233727, Abcam). Following, protease K buffer (Invitrogen) was applied to isolate the RNAs. Finally, RT-qPCR was used to analyze the enrichment of circ_0006174.
Pull-Down Assay
Pierce Magnetic RNA-Protein Pull-Down Kit (Thermo Fisher Scientific) was used to determine the relationship between miRNAs and circ_0006174 according to manufacturer’s instruction. Firstly, HT29 and SW620 cells (1×107) were lysed in RIP Lysis Buffer (Thermo Fisher Scientific). The circ_0006174 probe or oligo probes (RiboBio) was incubated with C-1 magnetic beads to obtain probe-coated beads, then incubated with cell lysate at 4°C overnight, followed by washed and RT-qPCR.
Dual-Luciferase Reporter Assay
The circ_0006174 wild type (circ_0006174-WT) or MACC1 3ʹ untranslated sequences (UTR, MACC1 3ʹUTR-WT) sequences were amplified and inserted into the luciferase reporter vectors pmiR-GLO (Promega, Madison, WI, USA). The mutant sequences of circ_0006174 (circ_0006174-MUT) or MACC1 3ʹUTR (MACC1 3ʹUTR-MUT) were inserted into pmiR-GLO (Promega) vectors. Then, all reporter vectors together with miR-138-5p mimics or miR-NC were co-transfected into HT29 and SW620 cells using Lipofectamine 2000 (Thermo Fisher Scientific). Forty-eight hours upon transfection, dual-luciferase Reporter Assay System (Genomeditech, Shanghai, China) was applied to detect the luciferase activities. Relative luciferase activity is shown as the ratio between the firefly luciferase activity and the Renal luciferase activity.
Western Blot
CRC cells and tissues were lysed by RIPA lysis buffer (Pierce, Rockford, IL, USA) and the upper liquid was collected after centrifugation. Total protein concentration was determined by the BCA assay kit (Beyotime, Shanghai, China). Equal amount of proteins (30 μg) was separated by 10–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE, Beyotime), and electrotransferred into polyvinylidene difluoride membrane (EMD, Millipore). Then, the membranes were blocked by 5% non-fat dry milk for 1 h at 25°C. The primary antibodies against MACC1 (1:1000; cat. no. ab106579, Abcam) and GADPH (1:5,000; cat. no. 10494-1-AP; Proteintech Group, Inc., Chicago, IL, USA) were used to incubate with membranes at 4°C overnight. Then, the membranes were washed with 0.5% Tris Buffered Saline Tween (TBST, Sigma-Aldrich), then horseradish peroxidase-conjugated secondary antibody (1:5000; cat. no. sc-2054; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) was employed to incubate with the membranes for 2 h. The protein levels were detected using enhanced chemiluminescence (Thermo Fisher Scientific) by Bio-Rad Gel imaging system (Thermo Fisher Scientific). GADPH was used as an internal control of MACC1.
Xenograft Tumor Model
Animal welfare and experimental procedures were carried out strictly in accordance with the Guide for the Care and Use of Laboratory Animals (Ministry of Science and Technology of China) and were approved by Animal Ethics Committee of Linhe people’s Hospital of Bayannur City. For xenograft tumor model, 4-weeks old male BALB/c nude mice purchased from Slaccas Shanghai Laboratory Animal Co., Ltd. (Shanghai, China) were randomly divided into two groups: sh-NC group (n = 7), sh-circ#1 group (n = 7). SW620 cells (2×106/100 μL) stable transfected with sh-circ #1 or sh-NC were subcutaneously injected into the right flank of nude mice. Seven days after injection, tumor volume was measured every 4 days with digital calipers and calculated using the following formula: tumor volume = 1/2 (length × width2). Thirty-five days after injection, the mice were sacrificed and the weight of the tumors was measured. Tumor tissues were harvested for the detection of circ_0006174, miR-138-5p and MACC1 expression in tumor tissues of nude mice with colorectal cancer.
Statistical Analysis
SPSS 21.0 software (IBM, Somers, NY, USA) and GraphPad primer 7.0 (GraphPad, La Jolla, CA, USA) were used for statistical analysis. All data from at least three independent experiments were presented as mean ± standard deviation (SD). Student’s t-test and one way analysis of variance (ANOVA) were applied to assess the differences in two or multiple groups. Kaplan–Meier method was used to evaluate the survival rate and analyzed by Log rank test. Pearson correlation coefficient analysis was used to analyze the relationship between the expression of circ_0006174 and miR-138-5p or MACC1 in CRC tissues. P <0.05 was considered to be statistically significant.
Results
The Upregulation of Circ_0006174 Was Associated with CRC Progression
To find the potential circRNAs in CRC, circRNAs expression profiles in 5 human CRC tissues and corresponding non-cancerous tissues (GSE142837), circRNA expression profiles of 6 human colorectal adenocarcinoma and normal mucosa (GSE138589), and circRNA dataset of 10 CRC patients (GSE126094), that from GEO database were selected. The differential expression analysis for GSE142837, GSE138589 and GSE126094 was conducted by using GEO2R tool provided by GEO database. Then, the top 15 upregulated circRNAs in 3 datasets were shown by cluster analysis. As listed in Figure 1A–C, has_circ_0006174 was highly upregulated in all CRC tissues in contrast with the adjacent non-tumor tissues. Has_circ_0006174 was circularized by exons 3 and 4 of the RAD23B gene [NM_002874] with a length of 349 nt according to circBase database (Figure 1D). Besides, RT-qPCR analysis indicated that has_circ_0006174 was markedly up-regulated in 40 CRC tumor tissues compared to paired adjacent normal tissues (Figure 1E). And the expression of circ_0006174 was also elevated in CRC patients with TNM stage III cancer (Figure 1F) and lymph node metastasis (Figure 1G), compared with patients with TNM stage I and II cancer and no lymph node metastasis. In addition, CRC patients were divided into high or low circ_0006174 expression level groups defined by the median value of circ_0006174, and the overall survival rate of patients was monitored. As shown in Figure 1H, Kaplan-Meier analysis showed that patients with high expression of circ_0006174 had a shorter overall survival (P < 0.05) (Figure 1H). These findings demonstrated that circ_0006174 was upregulated in CRC, and was correlated with poor prognosis.
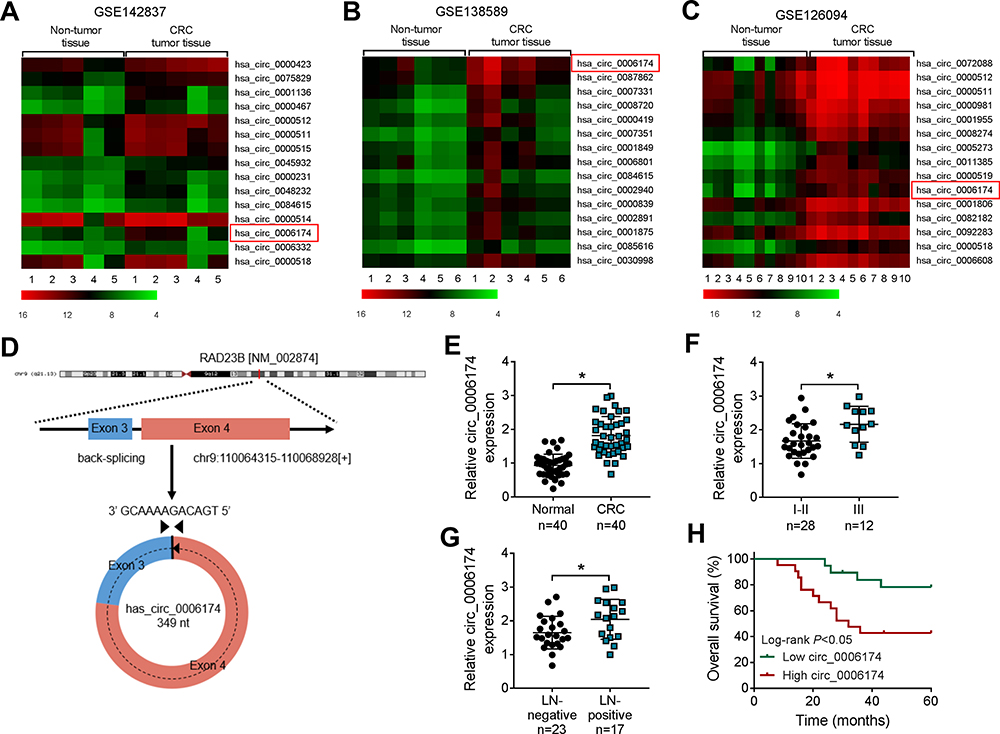 |
Figure 1 Circ_0006174 was upregulated in CRC tissues and was associated with CRC progression. (A–C) Heat map of top 15 upregulated circRNAs in CRC tissues and adjacent normal tissues from GSE142837, GSE138589, and GSE126094 datasets. (D) Circ_0006174 forms a circular structure through exons 3 and exons 4 in RAD23B gene. (E) Relative expression of circ_0006174 in CRC tissues (n=40) and adjacent normal tissues (n=40) was detected by RT-qPCR. (F) RT-qPCR was employed to detect circ_0006174 expression in patients with stage I–II CRC tissues (n=28) or stage III CRC tissues (n=12). (G) RT-qPCR was used to analyze circ_0006174 expression in patients with negative lymph node metastasis tissues (n=23) or positive lymph node metastasis tissues (n=17). (H) Kaplan-Meier survival analysis with a Log rank test was performed to compare the overall survival rate of CRC patients with higher (High circ_0006174) or lower (Low circ_0006174) level than median circ_0006174 expression level. Experiments were performed in triplicate, and data were presented as means ± SD. *P<0.05. |
Circ_0006174 Knockdown Inhibited the Proliferation, Metastasis and Facilitated the Apoptosis of CRC Cells
To explore the functional effect of circ_0006174 in CRC progression, loss-of-function experiments were performed to examine the biological behaviors of CRC cells. Firstly, the expression of circ_0006174 was detected by RT-qPCR in CRC cells and the normal colon epithelial cells. As shown in Figure 2A, circ_0006174 expression was significantly increased in CRC cells compared with normal colon epithelial cells NCM460. Then, three sh-circ (sh-circ #1, sh-circ #2, sh-circ #3) were employed to knock down circ_0006174 in CRC cells. As shown in Figure 2B and C, circ_0006174 expression was inhibited by sh-circ, especially sh-circ #1. And transfection of sh-circ nearly has no effect on RAD23B mRNA expression. Then, CCK-8 assay, clone formation assay, transwell assay and flow cytometry assay were conducted to explore the effect of circ_0006174 in CRC cells transfected with sh-circ #1. CCK-8 assay results indicated that circ_0006174 knockdown suppressed the viability of CRC cells (Figure 2D and E). Meanwhile, clone formation assay revealed that circ_0006174 knockdown restrained the proliferation in CRC cells (Figure 2F and G). Moreover, Transwell assay showed that circ_0006174 knockdown leads to the inhibition of cell migration and invasion (Figure 2H–K). Besides, flow cytometry assay disclosed that circ_0006174 knockdown promoted the apoptosis of CRC cells (Figure 2L and M). Taken together, these data suggested that circ_0006174 plays a carcinogenic role by affecting the proliferation, metastasis and apoptosis in CRC cells.
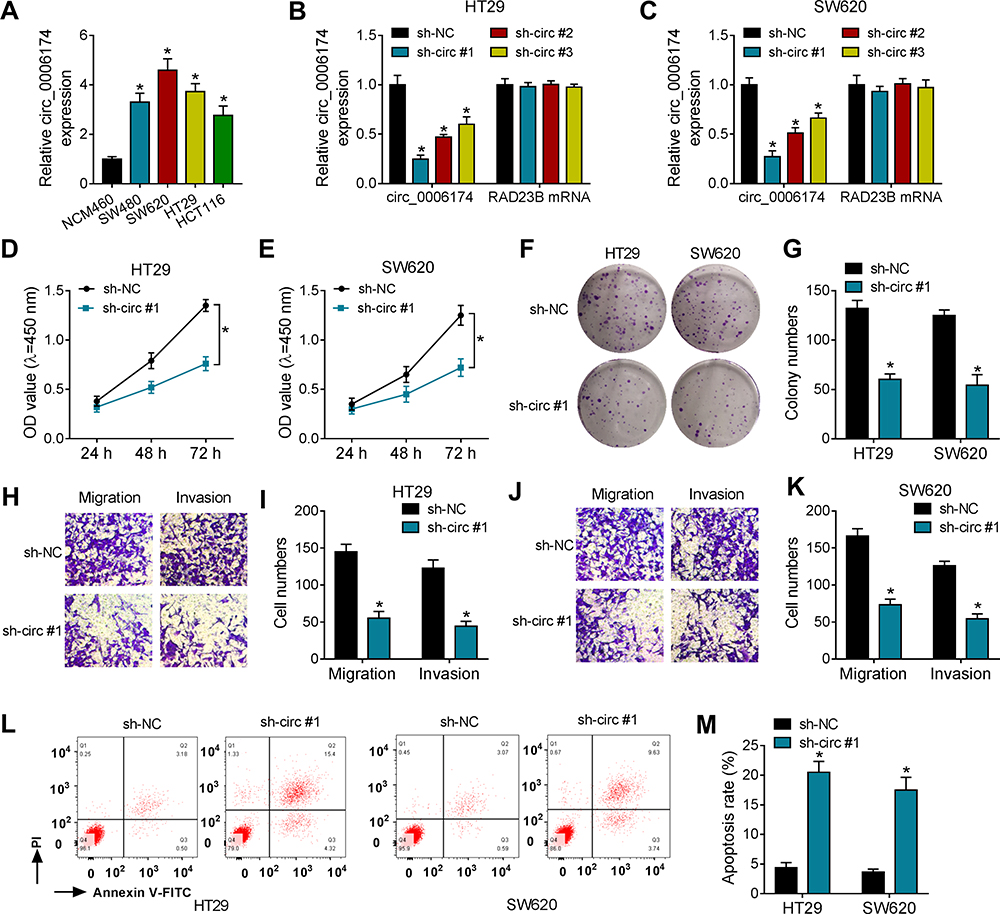 |
Figure 2 Circ_0006174 inhibition suppressed the proliferation, migration, invasion and promoted apoptosis in CRC cells. (A) Circ_0006174 expression in CRC cells and the normal colon epithelial cells was detected by RT-qPCR. (B and C) HT29 and SW620 cells were transfected with sh-NC, sh-circ #1, sh-circ #2 or sh-circ #3, respectively. And the expression of circ_0006174 and its linear transcript RAD23B were evaluated by RT-qPCR. (D and E) Cell viability of CRC cells transfected with sh-circ#1 or sh-NC was detected by CCK-8 assay. (F and G) Clone formation assay was used to test the proliferation of CRC cells transfected with sh-circ #1 or sh-NC. (H–K) Transwell assay was used to assess the migration and invasion abilities of CRC cells transfected with sh-circ #1 or sh-NC. (L and M) Flow cytometry assay was performed to evaluate the effect of circ_0006174 knockdown on the apoptosis of CRC cells. Three independent assays were performed, and data were presented as means ± SD. *P<0.05. |
Circ_0006174 Acted as a Molecular Sponge of miR-138-5p
The subcellular localization of circ_0006174 was confirmed by Nuclear and cytoplasmic separation experiment. The result indicated that circ_0006174 was mainly localized in the cytoplasm of CRC cells (Figure 3A and B). And Ago2 RIP assay revealed the existence of Ago2 (the main effector of RNA-induced silencing complex) binding sites in circ_0006174, as the enrichment of Ago2 protein was significantly increased in circ_0006174 group in contrast with GAPDH group, and the enrichment of Ago2 protein could be inhibited by sh-circ#1 (Figure 3C and D). These results indicating that circ_0006174 might influence CRC progression and functions by acting as a miRNA sponge that possess biological activity in the cytoplasm.
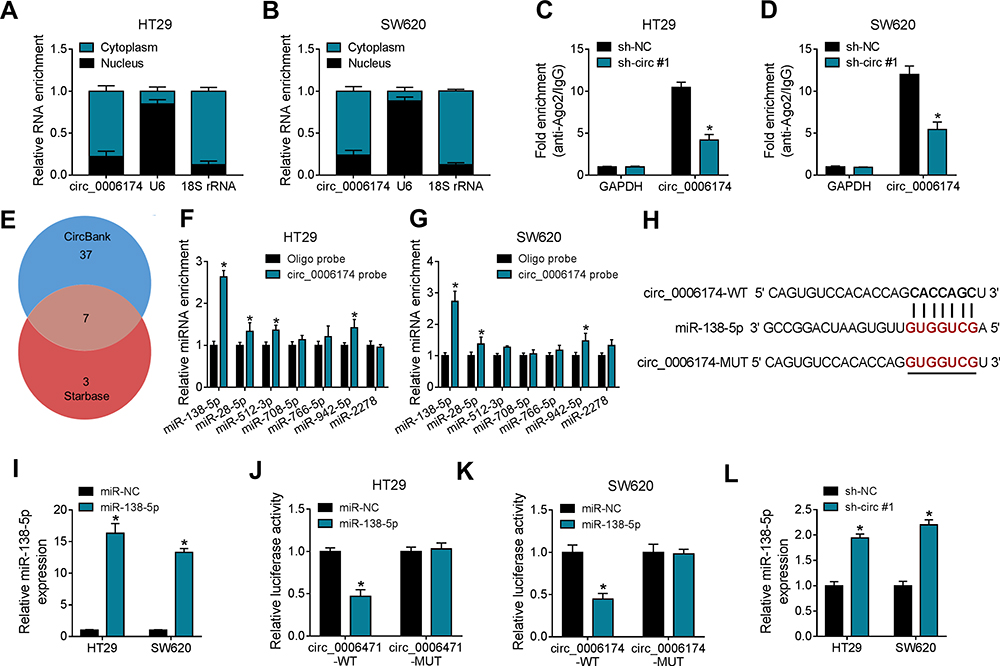 |
Figure 3 Circ_0006174 acted as a sponge of miR-138-5p. (A and B) Relative enrichments of circ_0006174, U6 and 18S rRNA in cytoplasm and nucleus were detected by RT-qPCR. (C and D) Ago2 RIP assay was employed to assess the combination of Ago2 and circ_0006174 in CRC cells. (E) The potential target miRNAs of circ_0006174 were predicted by Circbank and Starbase 2.0. (F and G) RNA pull down assay was performed to evaluate the target relationship between miRNAs and circ_0006174. (H) The online tool StarBase 2.0 was used to predict the binding sites between circ_0006174 and miR-138-5p. (I) RT-qPCR assay was performed to analyze miR-138-5p expression in CRC cells transfected with miR-138-5p or miR-NC. (J and K) Dual-luciferase reporter assay was performed to verify the interaction between miR-138-5p and circ_0006174. (L) Relative expression of miR-138-5p in CRC cells transfected with sh-circ #1 or sh-NC was detected by RT-qPCR. Experiments were performed in triplicate, and data were presented as means ± SD. *P<0.05. |
To investigate the potential miRNAs of circ_0006174, the online bioinformatics database CircBank and Starbase 2.0 were used, and 7 overlapping miRNAs including miR-138-5p, miR-28-5p, miR-512-3p, miR-708-5p, miR-766-5p, miR-942-5p and miR-2278 were recognized that were the most potentially miRNAs related to circ_0006174 (Figure 3E). RNA pull down assay indicated that the enrichment of miR-138-5p, miR-28-5p, miR-512-3p and miR-942-5p were significantly increased by circ_0006174 in HT29 and SW620 cells (Figure 3F and G). Among these predicted target miRNAs, we found that circ_0006174 was the most significant upregulated. Then, dual-luciferase reporter assay was performed. Circ_0006174 sequences contained wild type (circ_0006174-WT) or mutant (circ_0006174-MUT) miR-138-5p binding sites were used to construct luciferase reporter vector (Figure 3H). And miR-138-5p expression was 15-fold increased by miR-138-5p mimic (Figure 3I). Dual-luciferase reporter assay uncovered that transfection of miR-138-5p inhibited the luciferase activity of circ_0006174-WT group compared with miR-NC transfection, while it had little effect on the luciferase activity of circ_0006174-MUT group (Figure 3J and K). Moreover, miR-138-5p was upregulated in CRC cells transfected with sh-circ #1 (Figure 3L). Taken together, circ_0006174 served as a sponge for miR-138-5p.
miR-138-5p Inhibition Attenuated the Suppression on Proliferation, Metastasis and the Promotion on Apoptosis Mediated by Circ_0006174 Knockdown in CRC Cells
RT-qPCR assay indicated that miR-138-5p expression was dramatically decreased in 40 CRC tissues (Figure 4A) and was negatively correlated (r = −0.37, P < 0.05) with circ-PRRX1 expression (Figure 4B). Besides, miR-138-5p expression was declined in CRC cells compared with the normal human colon mucosal epithelial cell line NCM460 (Figure 4C). And the expression of miR-138-5p was inhibited by half in cells with miR-138-5p transfection (Figure 4D). To illuminate the interaction between circ_0006174 and miR-138-5p in CRC cells, HT29 and SW620 cells were transfected with sh-NC + anti-NC, sh-circ #1 + anti-NC, or sh-circ #1 + anti-miR-138-5p, respectively. CCK-8 assay analysis displayed that sh-circ #1-mediated the decrease in viability was reversed by anti-miR-138-5p in CRC cells (Figure 4E and F). Meanwhile, clone formation assay showed that anti-miR-138-5p could attenuate the suppression on proliferation that induced by sh-circ #1 in CRC cells (Figure 4G). Moreover, anti-miR-138-5p could relieve sh-circ #1-mediated decrease in migration and invasion in CRC cells (Figure 4H and I). Besides, the increasing apoptosis rate mediated by sh-circ #1 was alleviated by anti-miR-138-5p in CRC cells (Figure 4J). These data demonstrated that miR-138-5p performs the anti-tumor function by inhibiting cell proliferation and metastasis, but promoting apoptosis in CRC cells.
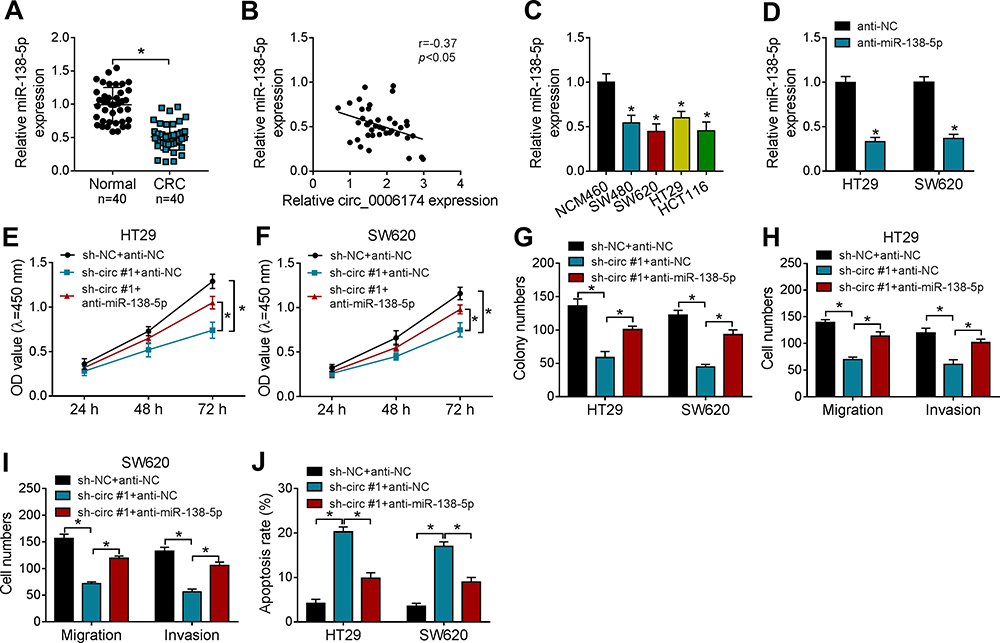 |
Figure 4 Downregulation of miR-138-5p reversed the effect of circ_0006174 on proliferation, migration, invasion and apoptosis of CRC cells. (A) The expression of miR-138-5p in 40 CRC tissues and adjacent normal tissues was detected by RT-qPCR. (B) The relationship between miR-138-5p expression and circ_0006174 expression was assessed by Pearson’s correlation analysis. (C) MiR-138-5p expression in CRC cells and the normal colon epithelial cells was detected by RT-qPCR. (D) The effect of miR-138-5p inhibitor on miR-138-5p expression in CRC cells was assessed by RT-qPCR. (E and F) CCK-8 assay was conducted to assess the viability of CRC cells transfected with sh-circ #1 + anti-miR-138-5p, sh-circ#1 + anti-NC or sh-NC + anti-NC. (G) The proliferation of CRC cells transfected with sh-circ #1 + anti-miR-138-5p or negative controls was measured by clone formation assay. (H and I) The migration and invasion abilities of CRC cells transfected with sh-circ #1 + anti-miR-138-5p or negative controls were estimated by transwell assay. (J) The apoptosis of CRC cells transfected with sh-circ #1 + anti-miR-138-5p or negative controls was detected by flow cytometry assay. Three independent assays were performed, and data were presented as means ± SD. *P<0.05. |
miR-138-5p Was Interacted with MACC1 in CRC Cells
Then, we further investigated the downstream target mRNAs of miR-138-5p using the bioinformatics tool Starbase 2.0. And we also screened the top 100 most upregulated mRNAs in 10 human CRC tissues based on GSE126092 datasets. As shown in Fig. 5A and 4 overlapping mRNAs were eventually screened out. A heat map represents the expression level of 4 overlapping mRNAs in CRC tissues in contrast with adjacent normal tissues. The results manifested that RNF43, KRT80, MACC1, and ONECUT2 were upregulated in CRC tissues (Figure 5A). Besides, the GEPIA database (http://gepia.cancer-pku.cn/detail.php) revealed the expression levels of 4 potential mRNAs in CRC tissues and adjacent normal tissues. As displayed in Figure 5B–E, RNF43, KRT80 and MACC1 levels were obviously upregulated in CRC tissues in contrast with adjacent normal tissues, however, ONECUT2 level showed no significant change. Further investigation indicated that MACC1 expression was the most significantly decreased in both HT29 and SW620 cells transfected with miR-138-5p mimic, compared to cells with miR-NC transfection (Figure 5F and G). To confirm the target relationship between miR-138-5p and MACC1, dual-luciferase reporter assay system was conducted. As shown in Figure 5H, the 3ʹUTR of MACC1 sequences contained wild or mutant type miR-138-5p binding sites were inserted into luciferase reporter vector. Dual-luciferase reporter assay illustrated that the luciferase activity of MACC1 3ʹUTR-WT group, not MACC1 3ʹUTR-MUT group, was inhibited by miR-138-5p transfection (Figure 5I and J), indicating that MACC1 was a target of miR-138-5p. Besides, the protein level of MACC1 was upregulated by anti-miR-138-5p, but was inhibited by miR-138-5p (Figure 5K). Furthermore, circ_0006174 knockdown suppressed the protein level of MACC1, whereas this suppression effect was reversed by co-transfection of anti-miR-138-5p (Figure 5L). These results indicated that circ_0006174 upregulated MACC1 protein level by sponging miR-138-5p in CRC cells.
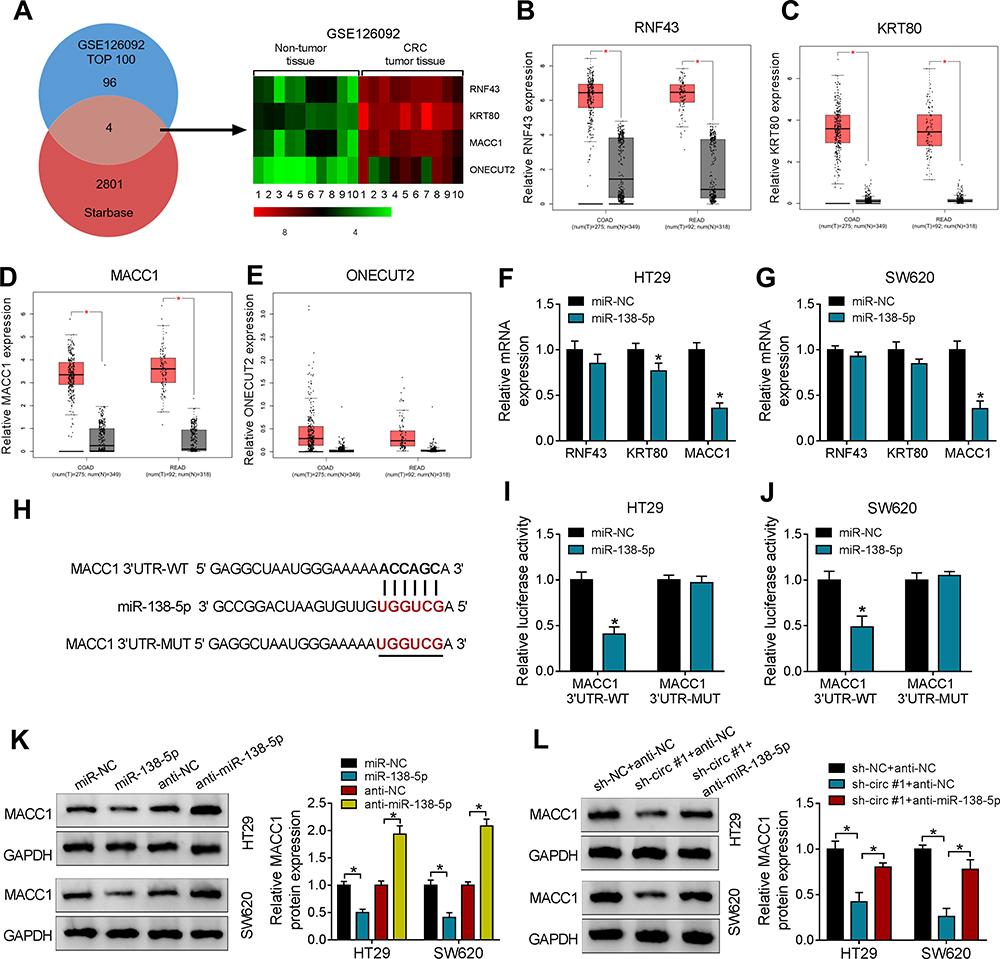 |
Figure 5 MiR-138-5p directly targeted MACC1 3ʹUTR in CRC cells. (A) Target mRNAs of miR-138-5p was predicted by Starbase software, and the intersection genes (RNF43, KRT80, MACC1 and ONECUT2) of the predicting results and the top 100 upregulated genes in CRC tissues compared to adjacent normal tissue based on the GEO dataset (GSE12902) were shown by heatmap. (B–E) The GEPIA database was used to analyze the expression of RNF43, KRT80, MACC1 and ONECUT2 in CRC tissues (COAD stands for colon adenocarcinoma and READ for rectal adenocarcinoma). (F and G) Relative mRNA expression of RNF43, KRT80 and MACC1 in CRC cells transfected with miR-138-5p mimic or miR-NC were measured by RT-qPCR. (H) The online tool StarBase 2.0 was employed to predict the binding sites of miR-138-5p in MACC1 3ʹUTR. (I and J) Dual-luciferase reporter assay was used to analyze the interaction between MACC1 and miR-138-5p in CRC cells. (K) The protein level of MACC1 in CRC cells transfected with miR-138-5p, anti-miR-138-5p or negative controls was detected by Western blot. (L) Relative MACC1 protein level in CRC cells transfected with sh-NC + anti-NC, sh-circ #1 + anti-NC, or sh-circ #1 + anti-miR-138-5p was measured by Western blot. Three independent assays were performed, and data were presented as means ± SD. *P<0.05. |
Circ_0006174 Knockdown Inhibited CRC Progression by miR-138-5p/MACC1 Axis in vitro
Then, we further investigate whether MACC1 participates in the effect of circ_0006174/miR-138-5p axis in CRC. MACC1 mRNA and protein levels were upregulated in 40 CRC tissues (Figure 6A and B). Besides, a strong positive correlation (r = 0.55, P < 0.05) was found in circ_0006174 expression and MACC1 mRNA level (Figure 6C). And the elevated level of MACC1 protein was also detected in CRC cells compared to normal human colon mucosal epithelial cell line NCM460 (Figure 6D). Then, loss-of-function assay was performed to investigate the function role of MACC1 in CRC cells. As shown in Figure 6E, the protein level of MACC1 was triple increased in cells with MACC1 overexpression. Besides, MACC1 overexpression partly reversed the suppression effect on proliferation, migration and invasion (Figure 6F–J) and promotion effect on apoptosis (Figure 6K) that mediated by sh-circ #1 in CRC cells. These results suggested that circ_0006174 advanced CRC progression through upregulating MACC1 level in CRC cells.
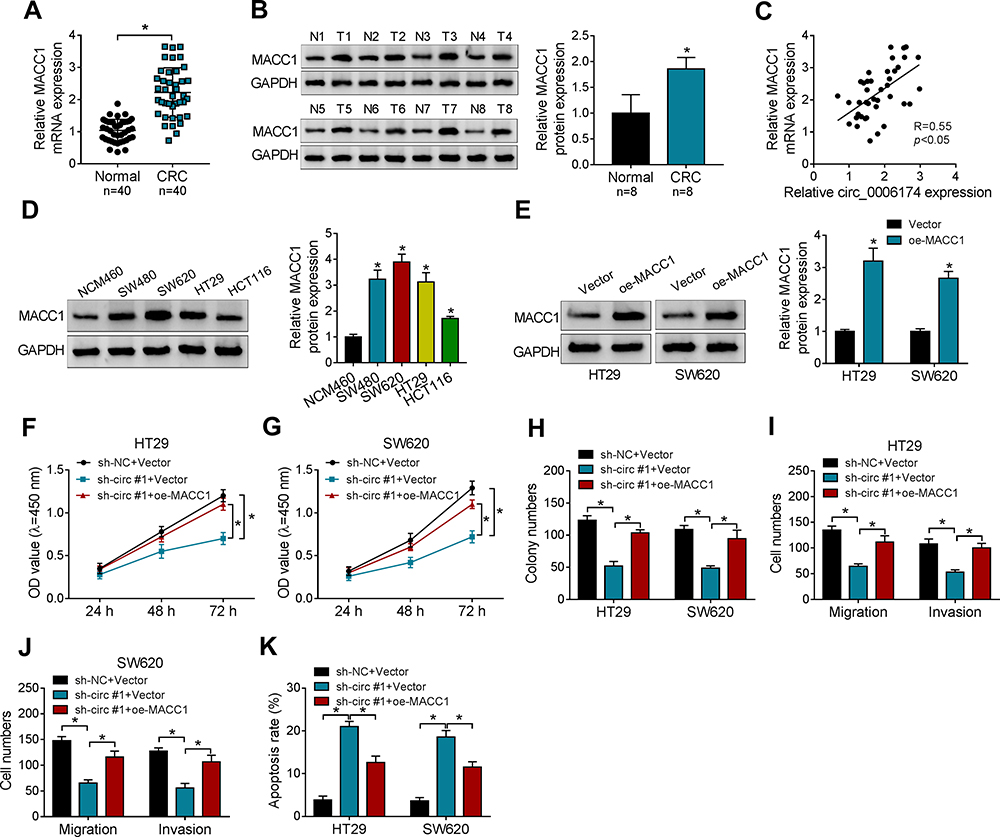 |
Figure 6 Overexpression of reversed the effect of circ_0006174 in CRC cells. (A) Relative MACC1 mRNA level in CRC tissues (n=40) and adjacent normal tissues (n=40) was detected by RT-qPCR. (B) Relative MACC1 protein level in CRC tissues (n=8) and adjacent normal tissues (n=8) was detected by Western blot. (C) The relationship between MACC1 mRNA level and circ_0006174 expression was evaluated by Pearson’s correlation analysis. (D) Western blot was used to analyze MACC1 protein level in CRC cells and the normal colon epithelial cells. (E) Western blot assay was conducted to evaluate the transfection efficiency of oe-MACC1. (F and G) The viability of CRC cells transfected with sh-circ #1 + oe-MACC1 or negative controls was evaluated by CCK-8 assay. (H) The proliferation of CRC cells transfected with sh-circ #1 + oe-MACC1 or negative controls was detected by clone formation assay. (I and J) The migration and invasion abilities of CRC cells was assessed by transwell assay. (K) The apoptosis of CRC cells transfected with sh-circ #1 + oe-MACC1 or negative controls was assessed by flow cytometry assay. Three independent assays were performed, and data were presented as means ± SD. *P<0.05. |
Circ_0006174 Knockdown Inhibited CRC Tumor Growth in vivo
Given the carcinogenic role of circ_0006174 in vitro, we further investigate the biological function of circ_0006174 in vivo. SW620 cells (2×106/100 μL) stable transfected with sh-circ #1 or sh-NC were subcutaneously injected into the right flank of nude mice (n = 7) to establish xenograft tumor model in nude mice. As expected, CRC tumor growth was restrained by circ_0006174 knockdown, as the tumor volume and weight in mice with circ_0006174 knockdown were dramatically declined compared to sh-NC group (Figure 7A and B). Besides, circ_0006174 expression was decreased while miR-138-5p expression was increased in xenograft tumor tissues with circ_0006174 knockdown in xenograft tumor model (Figure 7C and D). Meanwhile, circ_0006174 knockdown also downregulated the protein level of MACC1 (Figure 7E). Taken together, circ_0006174 promoted CRC progression in vivo through regulating miR-138-5p/MACC1 axis, which might be a promising therapeutic target for colorectal cancer.
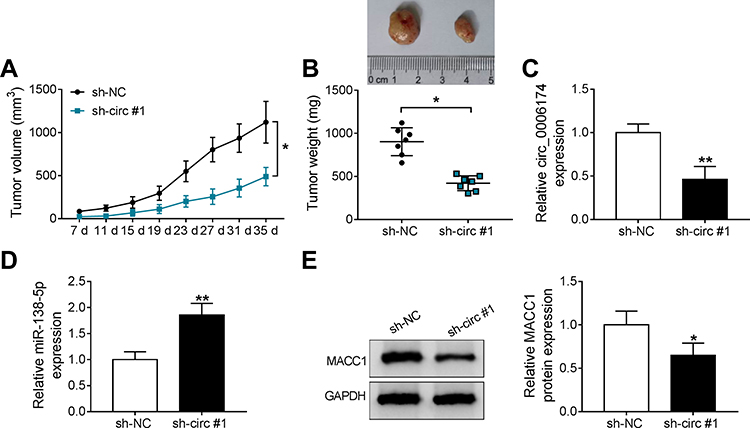 |
Figure 7 Knockdown of circ_0006174 inhibited tumor growth in vivo. (A) After nude mice were injected with SW620 cells transfected with sh-circ #1 or sh-NC for 7 days, tumor volumes were recorded every 4 days for 35 days. (B) Tumor weight was measured after the nude mice was euthanized. (C and D) The expression of circ_0006174 and miR-138-5p in transplanted tumor tissues with circ_0006174 knockdown. (E) Relative MACC1 protein level in nude mice that injected with SW620 cells stable transfected with sh-circ #1 or sh-NC. Three independent assays were performed, and data were presented as means ± SD. *P<0.05, **P<0.01. |
Discussion
Accumulating evidence has revealed that circRNAs played a pivotal role in cancer formation and progression, and served as regulators of cell biological behaviors including proliferation, cell cycle, metastasis, apoptosis.19 Moreover, the aberrant expression of circRNAs could be used as diagnostic or therapeutic biomarkers in various cancers.20 For instance, circN4BP2L2 and circEXOC6B served as bio-diagnostic markers in epithelial ovarian cancer.21 Circ_HIPK3 accelerated cell proliferation and metastasis via sponging miR-7, which could be used as a biomarker in CRC.22 In our research, the top 10 upregulated circRNAs in circRNAs expression profiles of CRC patients from GSE142837, GSE138589, and GSE126094 datasets were screened out. And circ_0006174 was chosen as the research object. The formation of circ_0006174 which was circularized by the exons 3 and 4 in RAD23B indicated that circ_0006174 belonged to ecircRNAs. A previous study illustrated that the dysregulation of circ_0006174 was contributed to chemo- or radio-resistant in CRC.23 However, the effects and mechanism underlying circ_0006174 in CRC have not yet been elucidated.
In our research, circ_0006174 was upregulated in CRC tissues and cells, and the elevated expression of circ_0006174 was closely related to the poor prognosis. Downregulation of circ_0006174 repressed cell proliferation, migration and invasion, but promoted the apoptosis in CRC cells. Besides, downregulation of circ_0006174 also curbed the tumor growth of CRC in vivo. These results indicated that circ_0006174 served as a tumor promoter in CRC progression. These results were consistent with previous research that circ_0006174 was upregulated in CRC cells.23
Some circRNAs affected the expression of microRNAs and parental genes, further affected the biological behaviors in cancer cells.13 EcircRNAs predominately situated in cytoplasm and acted as a molecular sponge for miRNAs.24 We also found that circ_0006174 mainly localized in cytoplasm, and contained the binding sites of Ago2, indicating that circ_0006174 may function as a miRNA sponge in CRC cells. We ultimately selected miR-138-5p for further research from 7 candidate miRNAs. A number of studies have found that miR-138-5p exerted a tumor-suppression role in CRC.25–27 Xu et al indicated that miR-138-5p inhibited cell migration and enhanced chemoresistance in CRC cells by NFIB-Snail1 axis.26 In our research, miR-138-5p was downregulated in CRC tissues and cells and was negatively correlated with circ_0006174 expression. Downregulation of miR-138-5p ameliorated circ_0006174 depletion-mediated depression in proliferation and metastasis, and promotion in apoptosis of CRC cells. These results uncovered that circ_0006174 acted as a molecular sponge for miR-138-5p and regulated the biological behaviors of CRC cells.
It is widely known that circRNAs could act as a competitive endogenous RNA (ceRNA) for miRNAs to regulate the downstream mRNA expression.11 Our data revealed that miR-138-5p targeted MACC1 and negatively regulated MACC1 protein expression in CRC cells. Previous research suggested that MACC1 was regulated by miR-218 at the post-transcriptional level.28 MiR-944 directly targeted MACC1 to inhibit the metastasis of CRC cells.29 MiR-338-3p acted as an inhibitor in malignant melanoma via targeting MACC1.30 In our research, MACC1 overexpression relieved the suppression effect on proliferation, migration and invasion, and the promotion effect on apoptosis that induced by circ_0006174 depletion in CRC cells.
In this study, we uncovered that circ_0006174 mediated CRC progression via elevating MACC1 expression through sponging miR-138-5p (Figure 8). Our results revealed the role and potential mechanism of circ_0006174 and provided evidence that circ_0006174 served as a therapeutic target for CRC therapy. Lack of clinical data, this study still needs further investigation to elucidate the possibility of using circ_0006174 as a diagnostic target in CRC.
 |
Figure 8 Schematic illustrated the circ_0006174/miR-138-5p/MACC1 axis in CRC cells. Circ_0006174 promoted cell proliferation, metastasis, and inhibited cell apoptosis through miR-138-5p/MACC1 axis in CRC cells. |
Highlights
- We first confirmed the interaction between miR-138-5p and circ_0006174 or MACC1.
- Circ_0006174 knockdown suppressed the proliferation and metastasis, and activated the apoptosis in colorectal cancer cells.
- Circ_0006174 advanced colorectal cancer progression by regulating miR-138-5p/MACC1 axis.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
2. Cummings OW. Pathology of the adenoma-carcinoma sequence: from aberrant crypt focus to invasive carcinoma. Semin Gastrointest Dis. 2000;11(4):229–237.
3. Jass JR. Classification of colorectal cancer based on correlation of clinical, morphological and molecular features. Histopathology. 2007;50(1):113–130. doi:10.1111/j.1365-2559.2006.02549.x
4. Lee RM, Cardona K, Russell MC. Historical perspective: two decades of progress in treating metastatic colorectal cancer. J Surg Oncol. 2019;119(5):549–563. doi:10.1002/jso.25431
5. Goyette MC, Cho K, Fasching CL, et al. Progression of colorectal cancer is associated with multiple tumor suppressor gene defects but inhibition of tumorigenicity is accomplished by correction of any single defect via chromosome transfer. Mol Cell Biol. 1992;12(3):1387–1395. doi:10.1128/MCB.12.3.1387
6. Nearchou A, Pentheroudakis G. The significance of tumor-associated immune response in molecular taxonomy, prognosis and therapy of colorectal cancer patients. Ann Trad Med. 2016;4(14):271. doi:10.21037/atm.2016.05.54
7. Wang P, He X. Current research on circular RNAs associated with colorectal cancer. Scand J Gastroenterol. 2017;52(11):1203–1210. doi:10.1080/00365521.2017.1365168
8. Vicens Q, Westhof E. Biogenesis of Circular RNAs. Cell. 2014;159(1):13–14. doi:10.1016/j.cell.2014.09.005
9. Guo JU, Agarwal V, Guo H, Bartel DP. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014;15(7):409. doi:10.1186/s13059-014-0409-z
10. Jeck WR, Sorrentino JA, Wang K, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA (New York, NY). 2013;19(2):141–157. doi:10.1261/rna.035667.112
11. Li Z, Huang C, Bao C, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol. 2015;22(3):256–264. doi:10.1038/nsmb.2959
12. Granados-Riveron JT, Aquino-Jarquin G. The complexity of the translation ability of circRNAs. Biochim Biophys Acta. 2016;1859(10):1245–1251. doi:10.1016/j.bbagrm.2016.07.009
13. Chen I, Chen C-Y, Chuang T-J. Biogenesis, identification, and function of exonic circular RNAs. Wiley Interdiscip Rev RNA. 2015;6(5):563–579. doi:10.1002/wrna.1294
14. Gao Y, Wang J, Zhao F. CIRI: an efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015;16(1):4. doi:10.1186/s13059-014-0571-3
15. Zheng Q, Bao C, Guo W, et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016;7(1):11215. doi:10.1038/ncomms11215
16. Bachmayr-Heyda A, Reiner AT, Auer K, et al. Correlation of circular RNA abundance with proliferation–exemplified with colorectal and ovarian cancer, idiopathic lung fibrosis, and normal human tissues. Sci Rep. 2015;5(1):8057. doi:10.1038/srep08057
17. Zhu M, Xu Y, Chen Y, Yan F. Circular BANP, an upregulated circular RNA that modulates cell proliferation in colorectal cancer. Biomed Pharmacother. 2017;88:138–144. doi:10.1016/j.biopha.2016.12.097
18. Turchinovich A, Weiz L, Langheinz A, Burwinkel B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011;39(16):7223–7233. doi:10.1093/nar/gkr254
19. Sun Z, Yang S, Zhou Q, et al. Emerging role of exosome-derived long non-coding RNAs in tumor microenvironment. Mol Cancer. 2018;17(1). doi:10.1186/s12943-018-0831-z.
20. Meng S, Zhou H, Feng Z, et al. CircRNA: functions and properties of a novel potential biomarker for cancer. Mol Cancer. 2017;16(1). doi:10.1186/s12943-017-0663-2.
21. Ning L, Long B, Zhang W, et al. Circular RNA profiling reveals circEXOC6B and circN4BP2L2 as novel prognostic biomarkers in epithelial ovarian cancer. Int J Oncol. 2018. doi:10.3892/ijo.2018.4566
22. Zeng K, Chen X, Xu M, et al. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018;9(4):417. doi:10.1038/s41419-018-0454-8
23. Xiong W, Ai YQ, Li YF, et al. Microarray analysis of circular RNA expression profile associated with 5-fluorouracil-based chemoradiation resistance in colorectal cancer cells. Biomed Res Int. 2017;2017:8421614. doi:10.1155/2017/8421614
24. Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–338. doi:10.1038/nature11928
25. Chen LY, Wang L, Ren YX, et al. The circular RNA circ-ERBIN promotes growth and metastasis of colorectal cancer by miR-125a-5p and miR-138-5p/4EBP-1 mediated cap-independent HIF-1α translation. Mol Cancer. 2020;19(1):164. doi:10.1186/s12943-020-01272-9
26. Xu W, Chen B, Ke D, Chen X. MicroRNA-138-5p targets the NFIB-Snail1 axis to inhibit colorectal cancer cell migration and chemoresistance. Cancer Cell Int. 2020;20:475.
27. Yan Z, Bi M, Zhang Q, Song Y, Hong S. LncRNA TUG1 promotes the progression of colorectal cancer via the miR-138-5p/ZEB2 axis. Biosci Rep. 2020;40(6). doi:10.1042/BSR20201025
28. Ilm K, Fuchs S, Mudduluru G, Stein U. MACC1 is post-transcriptionally regulated by miR-218 in colorectal cancer. Oncotarget. 2016;7(33):53443–53458. doi:10.18632/oncotarget.10803
29. Wen L, Li Y, Jiang Z, Zhang Y, Yang B, Han F. miR-944 inhibits cell migration and invasion by targeting MACC1 in colorectal cancer. Oncol Rep. 2017;37(6):3415–3422. doi:10.3892/or.2017.5611
30. Yang G, Zhang C, Wang N, Chen J. miR-425-5p decreases LncRNA MALAT1 and TUG1 expressions and suppresses tumorigenesis in osteosarcoma via Wnt/beta-catenin signaling pathway. Int J Biochem Cell Biol. 2019;111:42–51. doi:10.1016/j.biocel.2019.04.004
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.