Back to Journals » Journal of Inflammation Research » Volume 19
Chronic Radiation Dermatitis After Breast Cancer Radiotherapy: Mechanistic Insights, Therapeutic Challenges, and Emerging Roles of Piezo1-Mediated Mechanotransduction
Received 18 March 2026
Accepted for publication 19 May 2026
Published 11 June 2026 Volume 2026:19 609340
DOI https://doi.org/10.2147/JIR.S609340
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Anish R. Maskey
Zhiyao Wang,* Feiyu Chen,* Zhi Li
Department of Plastic Surgery, Third Hospital of Shanxi Medical University, Taiyuan, Shanxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiyao Wang; Zhi Li, Department of Plastic Surgery, Third Hospital of Shanxi Medical University, Shanxi Bethune Hospital, Shanxi Academy of Medical Sciences, Tongji Shanxi Hospital, Taiyuan, Shanxi, People’s Republic of China, Email [email protected]; [email protected]
Background: Chronic radiation dermatitis (CRD) is a common late complication of postoperative radiotherapy for breast cancer. Its pathological process involves multiple interrelated factors, including persistent inflammation, microvascular injury, fibrosis, and tissue remodeling, and may substantially impair patients’ quality of life. Current clinical management remains largely symptomatic, including topical therapies, hyperbaric oxygen therapy, laser treatment, and fat grafting; however, overall therapeutic efficacy remains limited, suggesting that the underlying mechanisms of CRD have not yet been fully elucidated.
Objective: This review aims to integrate the major pathological mechanisms underlying CRD after breast cancer surgery and the limitations of current therapeutic strategies, and to propose a mechanistic explanatory framework from the perspective of mechanotransduction, with the goal of exploring potential reasons for the limited efficacy of existing treatments.
Methods: By reviewing the relevant literature, this article systematically integrates key pathological processes involved in CRD, including inflammatory responses, vascular dysfunction, and fibrotic remodeling, and further interprets these processes in light of recent advances in the mechanical microenvironment and mechanotransduction.
Results: Current evidence indicates that the development and progression of CRD are closely associated with persistent inflammation, microvascular injury, and progressive fibrosis. However, alterations in the mechanical microenvironment caused by extracellular matrix deposition and increased tissue stiffness after radiation injury may represent an important but underrecognized regulatory factor. Multiple mechanotransduction pathways may be involved in this process, including integrin-mediated adhesion signaling, the YAP/TAZ pathway, and transient receptor potential channels. As a representative mechanosensitive ion channel, Piezo1 has been implicated in the regulation of inflammation, vascular function, and fibrosis-related processes. Nevertheless, direct evidence regarding its role in CRD remains limited, and current assumptions are largely extrapolated from indirect findings in other tissues or disease models.
Conclusion: This review re-examines the pathological process of CRD from the perspective of the mechanical microenvironment and its associated mechanotransduction mechanisms, positioning Piezo1 as a potential mechanistic link rather than a proven central pathogenic driver. This perspective may help contextualize the development and progression of CRD within a broader mechanotransduction network and provide a hypothesis-generating basis for future mechanistic studies and potential targeted interventions.
Keywords: breast neoplasms, radiation injuries, radiodermatitis, mechanotransduction, PIEZO1, fibrosis, mechanosensitive ion channels
Introduction
Breast cancer is one of the most frequently diagnosed malignancies among women worldwide.1 With advances in breast surgery and oncoplastic principles, an increasing number of patients undergo breast-conserving surgery or breast reconstruction after tumor resection, with the aim of improving postoperative appearance and quality of life while maintaining oncological safety.2 For patients undergoing breast-conserving surgery and selected patients after modified radical mastectomy, postoperative radiotherapy (RT) is commonly used as an adjuvant treatment to eradicate residual microscopic disease, reduce the risk of local recurrence, and improve long-term outcomes.3 Evidence has shown that RT can significantly reduce local recurrence by eliminating microscopic tumor foci remaining in breast-conserving tissue and may also reduce breast cancer–related mortality to some extent.4 Despite continuous advances in radiotherapy techniques and substantial improvements in overall prognosis, radiation-related skin injury remains one of the most common adverse effects in clinical practice, with CRD representing an important form of late toxicity.5,6
CRD is generally defined as skin injury that develops or persists more than 90 days after the completion of radiotherapy, and its clinical manifestations may progress gradually over months to years.7 Epidemiological studies indicate that radiation-related skin injury is a common adverse event after breast cancer radiotherapy, while late radiation-induced tissue injury, including chronic skin changes and radiation-induced fibrosis, has been reported in approximately 5–30% of patients across different studies. This incidence is influenced by multiple factors, particularly radiation dose, irradiated volume, and fractionation schedule, with a markedly increased risk under conditions of high-dose or large-volume irradiation.8 In some patients, CRD may further progress to chronic ulceration, tissue necrosis, and an increased risk of secondary malignancies, among which radiation-induced fibrosis (RIF) is a major pathological feature of the late stage.9 Persistent chronic skin injury and non-healing wounds not only substantially impair patients’ quality of life but also impose a sustained burden on healthcare systems.10 At present, the clinical management of CRD mainly relies on symptomatic treatment, including basic skin care, topical medications, and selected physical or laser-based interventions; however, overall therapeutic efficacy remains limited.11–13 These challenges suggest that the underlying pathogenesis of CRD has not been fully elucidated and highlight the need for new biological perspectives. In recent years, the mechanical microenvironment and key mechanosensing molecules, such as Piezo1, have increasingly been recognized as potential contributors to chronic tissue injury. Traditionally, the development and progression of CRD have been explained mainly in terms of persistent inflammation, microvascular injury, and progressive fibrosis. However, emerging evidence suggests that radiation injury not only induces cellular damage and inflammatory responses but also alters the mechanical properties of local tissues by promoting extracellular matrix (ECM) deposition and tissue remodeling.14 Reduced tissue compliance and increased matrix stiffness may create an abnormal mechanical microenvironment, which can influence cellular functions, including proliferation, migration, and fibrosis-related processes, through mechanotransduction mechanisms.14 It should be noted that mechanotransduction involves multiple regulatory mechanisms, including integrin-mediated adhesion signaling, cytoskeletal remodeling, the YAP/TAZ pathway, and mechanosensitive ion channels, which may act in a coordinated manner. Although the role of the mechanical microenvironment in various forms of chronic tissue injury has received increasing attention, its specific regulatory mechanisms in CRD remain poorly understood.
Among various mechanosensitive molecules, Piezo1 is a mechanosensitive ion channel that detects membrane tension and external mechanical stimuli. By mediating Ca2⁺ influx, Piezo1 can regulate multiple downstream signaling pathways and may thereby contribute to inflammatory responses, vascular homeostasis, and fibrotic remodeling.15,16 Previous studies have implicated Piezo1 in tissue fibrosis and chronic injury across several disease contexts; however, its role in radiation-induced skin injury, particularly the specific mechanisms by which it may contribute to the development and progression of CRD after breast cancer surgery, remains insufficiently supported by direct evidence.
Against this background, this review integrates current evidence on inflammatory responses, vascular dysfunction, and fibrotic remodeling from the perspective of mechanotransduction, and proposes a potential mechanistic framework for understanding the development and progression of CRD. On this basis, evidence from related fields is further discussed to explore the possible role of Piezo1 in this process, while positioning it within the broader mechanotransduction network, with the aim of providing a hypothesis-generating foundation for future mechanistic studies.
Pathological Mechanisms and Therapeutic Challenges of Chronic Radiation Dermatitis After Breast Cancer Treatment
CRD is a common late complication after breast cancer radiotherapy and primarily occurs within the irradiated field, most often involving the breast or chest wall tissues in the setting of postoperative adjuvant radiotherapy.17 Ionizing radiation can induce skin cell injury through direct DNA damage and the generation of reactive oxygen species (ROS), thereby triggering acute radiation dermatitis (ARD).18 In some patients, acute radiation-induced injury does not fully resolve and gradually progresses into a chronic phase. In parallel, the combined effects of surgical trauma and radiation injury may further reshape the local tissue microenvironment, leading to a series of complex pathological processes, including persistent inflammation, microvascular injury, and progressive fibrosis. These changes are considered important contributors to the limited efficacy of current CRD treatments.19
A persistent inflammatory microenvironment is generally considered one of the key factors driving the development and progression of CRD, and is often related to incomplete resolution of the inflammatory response after acute radiation injury. In this process, radiation-induced DNA damage can impair the proliferative capacity of basal stem and progenitor cells, thereby reducing epidermal renewal.20 Meanwhile, ionizing radiation generates secondary electrons and ROS, which directly damage cell membranes and DNA and trigger an acute inflammatory response.21–23 During the acute phase, inflammatory cells such as neutrophils are recruited to the irradiated area and release various cytotoxic mediators, further aggravating local tissue injury.24 In some patients, this inflammatory response may persist and gradually evolve into a chronic inflammatory state, thereby contributing to the progression of CRD. At the same time, radiation can activate keratinocytes, fibroblasts, and vascular endothelial cells, promote immune-cell infiltration, and induce the expression of multiple inflammatory cytokines and chemokines.25,26 These signals not only promote matrix degradation but also upregulate the expression of vascular adhesion molecules and enhance inflammatory-cell recruitment, further disrupting tissue homeostasis and laying the groundwork for subsequent microvascular injury and fibrosis.27
In addition to inflammation, radiation injury can markedly affect the cutaneous microvasculature. Ionizing radiation may directly damage vascular endothelial cells, leading to capillary disruption and reduced microvascular density, thereby compromising local blood perfusion.19 As the microvascular network becomes progressively impaired, the irradiated area often develops chronic hypoxia, which is considered one of the key factors contributing to the sustained progression of radiation-induced tissue injury.19 Although a certain degree of compensatory angiogenic response may occur during the early phase of injury, persistent inflammation and tissue damage often lead to gradual vascular degeneration, characterized by decreased capillary density and abnormal vascular dilatation.28 These microcirculatory changes further impair tissue perfusion and are accompanied by increased ROS levels and sustained oxidative stress, thereby aggravating cellular damage and maintaining the pathological process.28
Progressive fibrosis is considered one of the major pathological features during the progression of CRD. Persistent inflammation and microvascular injury can activate multiple profibrotic signaling pathways, among which transforming growth factor-β (TGF-β) plays a central role in radiation-induced fibrosis.19 Sustained activation of TGF-β signaling promotes the differentiation of fibroblasts into myofibroblasts through Smad-dependent and related pathways, accompanied by upregulation of α-smooth muscle actin (α-SMA). Activated myofibroblasts produce large amounts of collagen and fibronectin, thereby promoting ECM deposition. In addition, TGF-β can induce the expression of connective tissue growth factor (CTGF/CCN2), further amplifying profibrotic signaling. As ECM continues to accumulate, tissue architecture is progressively remodeled, leading to fibrotic changes characterized by increased collagen deposition. This process constitutes an important histological basis for the chronic progression of CRD.28
With the progressive deposition of ECM in the dermis, tissue changes occur not only at the morphological level but also in terms of mechanical properties. Fibrotic tissue is typically characterized by increased matrix stiffness and collagen fiber reorganization, thereby creating an abnormal mechanical microenvironment. In this context, cells may be exposed to increased mechanical stress, which can be sensed and converted into intracellular signals through mechanotransduction, thereby influencing processes such as cell proliferation, migration, and matrix remodeling.28 Notably, existing studies related to breast cancer radiotherapy have suggested persistent remodeling of irradiated skin and subcutaneous tissues from both pathological and biophysical perspectives. Beyond inflammation, microvascular injury, and fibrotic changes, post-radiotherapy skin may exhibit reduced elasticity, altered tissue compliance, and localized induration. These changes indicate that CRD is not merely an epidermal injury, but rather a chronic process involving long-term structural remodeling of the dermis and subcutaneous tissues.19,23,27,28 Therefore, ECM deposition, collagen fiber reorganization, and increased tissue stiffness in CRD may collectively establish a persistently abnormal mechanical microenvironment, providing disease-specific rationale for further investigating the role of mechanotransduction in this condition.
In breast cancer radiotherapy–related CRD, alterations in the local mechanical microenvironment may be even more complex. Breast tissue is inherently compliant and is continuously exposed to dynamic mechanical stimuli during respiration, upper-limb movement, and postural changes. In addition, scar tissue formed after breast surgery, such as breast-conserving surgery or mastectomy, can alter the local distribution of tissue tension.29 Scar tissue is typically characterized by highly aligned collagen fibers and reduced tissue compliance, thereby increasing local mechanical loading. On this basis, radiotherapy-induced fibrosis may further promote ECM deposition and increase tissue stiffness, together shaping a persistently evolving mechanical environment.
In addition, axillary procedures, such as sentinel lymph node biopsy or axillary lymph node dissection, may impair lymphatic drainage to varying degrees, leading to fluid retention and increased interstitial pressure. These changes may further alter the local hydrodynamic environment and increase mechanical loading.30 The combined effects of these factors create a complex and dynamically changing mechanical microenvironment within the irradiated breast region, which may play an important role in the development and progression of CRD. The overall framework of these pathological processes is shown in Figure 1.
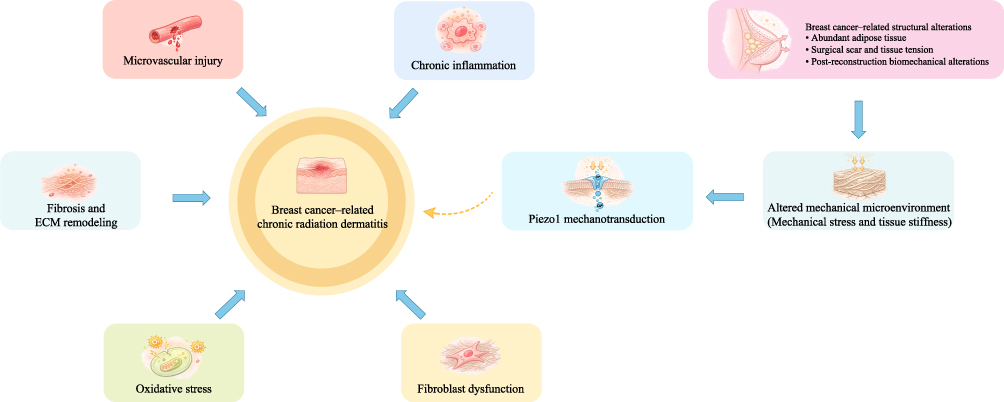 |
Figure 1 Proposed mechanisms involved in breast cancer–related chronic radiation dermatitis. CRD after breast cancer radiotherapy is associated with persistent inflammation, microvascular injury, fibrosis, and ECM remodeling. Structural alterations related to breast cancer surgery and reconstruction may further modify the local mechanical microenvironment, potentially increasing tissue stiffness and mechanical stress and contributing to disease progression. |
Current Therapeutic Strategies for Chronic Radiation Dermatitis and Their Limitations
Current therapeutic strategies for CRD remain limited, and clinical interventions are largely symptomatic, targeting known pathological processes. Existing approaches mainly focus on key pathological components, including persistent inflammation, microvascular injury, and progressive fibrosis, and include anti-inflammatory treatment, improvement of local blood supply, antifibrotic therapy, and surgical intervention for severe lesions. Their main mechanisms, advantages, and limitations are summarized in Table 1.
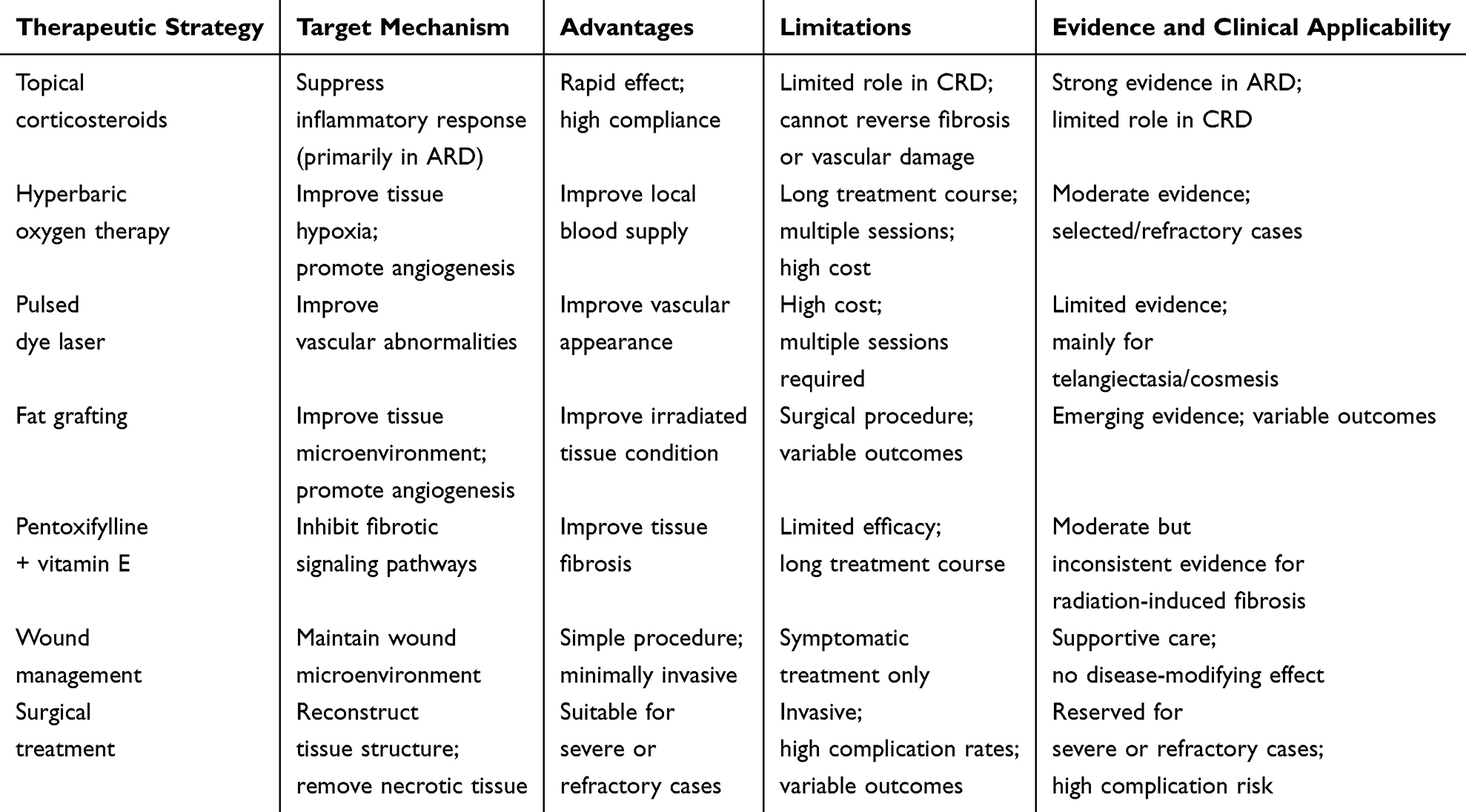 |
Table 1 Current Therapeutic Strategies for Chronic Radiation Dermatitis (CRD) and Their Limitations |
Glucocorticoids are commonly used anti-inflammatory agents for the prevention and alleviation of ARD in clinical practice. By suppressing inflammatory signaling pathways and reducing the expression of proinflammatory cytokines such as TNF-α, IL-1, and IL-6, glucocorticoids can attenuate radiation-related inflammatory responses.31 Several randomized controlled trials have shown that topical glucocorticoids, including mometasone furoate and betamethasone cream, can improve erythema, pruritus, and irritation.32,33 However, these treatments primarily act during the acute inflammatory phase and have limited effects on persistent fibrosis and microvascular injury once CRD has entered the chronic stage. Previous studies have suggested that radiation-induced inflammation can further activate profibrotic pathways, such as TGF-β signaling, thereby promoting fibroblast activation and ECM deposition.19 This may partly explain why anti-inflammatory therapy alone is insufficient to halt the long-term progression of CRD.
For microvascular abnormalities, several approaches aimed at improving local perfusion have been explored in clinical practice. Hyperbaric oxygen therapy (HBOT) can increase tissue oxygen tension and alleviate hypoxia, thereby promoting tissue repair.34 Pulsed dye laser (PDL) is mainly used to improve post-radiotherapy telangiectasia and cosmetic skin changes.35 In addition, fat grafting has also been suggested to exert beneficial effects by improving the local tissue environment and promoting angiogenesis.36 Although these approaches may improve local blood supply, tissue hypoxia, or cosmetic appearance to some extent, their effects are largely confined to local perfusion and tissue repair. They have limited impact on established fibrosis, ECM deposition, and structural remodeling, and therefore remain insufficient for modifying the long-term course of CRD.34–36
For progressive fibrosis, several antifibrotic strategies have been explored. Among them, pentoxifylline combined with vitamin E is one of the more commonly used regimens and may reduce ECM deposition by inhibiting TGF-β–related pathways and improving microcirculation.37 Clinical studies have shown that this combination can improve the extent of fibrosis and tissue compliance to some extent.38 In addition, fat grafting may also participate in tissue remodeling through the secretion of various growth factors by adipose-derived stem cells.36 However, the overall body of evidence remains limited, and these treatments are generally unable to fully reverse established fibrotic changes. In particular, their effects on long-term tissue induration, reduced compliance, and the abnormal mechanical microenvironment remain unclear, and their efficacy in the long-term management of CRD requires further validation.
For CRD-related wounds, symptomatic measures such as moist dressings, moisturizers, and local wound care are commonly used in clinical practice to maintain an appropriate wound environment and promote repair.19 In patients with more severe disease progression, such as refractory ulceration, tissue necrosis, or pronounced fibrosis, surgical intervention may be required, including excision of affected tissues and flap reconstruction.39 However, previous studies have shown that such surgical approaches are often associated with a relatively high complication rate and uncertain therapeutic outcomes. Therefore, although these measures may improve local conditions to some extent, they remain essentially supportive or salvage treatments and are unlikely to fundamentally reverse the deep tissue remodeling caused by radiation injury.
Taken together, current therapeutic strategies for CRD mainly target different pathological components, including inflammation, microvascular injury, tissue fibrosis, and wound repair. However, most approaches intervene in a single component or downstream pathological process, and their overall efficacy remains limited. Notably, the chronic stage of CRD is characterized not only by persistent inflammation and progressive fibrosis but also by biophysical alterations such as ECM deposition, reduced tissue compliance, and increased local stiffness, which together create a persistently abnormal mechanical microenvironment. Nevertheless, most existing treatment strategies do not directly address remodeling of the mechanical microenvironment or the associated mechanotransduction processes, and relevant interventions remain limited. Therefore, further understanding the development and progression of CRD from the perspective of the mechanical microenvironment and its signaling mechanisms, and exploring potential molecular regulatory nodes, may provide additional insights into the limited efficacy of current treatments and the development of new therapeutic strategies.
Potential Roles and Mechanisms of Piezo1 in CRD
Structure and Basic Biological Functions of Piezo1
Piezo1 is a large mechanosensitive cation channel composed of approximately 2500 amino acids and organized as a trimeric propeller-like complex. Each subunit contains approximately 38 transmembrane helices, with peripheral blades forming a curved, dome-like configuration in the resting state.40,41 The central region of the channel consists of an ion-conducting pore and an extracellular cap, while intracellular beam-like structures may contribute to mechanically induced conformational changes through a lever-like mechanism, thereby facilitating channel opening.42,43 Upon activation, Piezo1 mediates the influx of cations such as Ca2⁺, Na⁺, K⁺, and Mg2⁺, and is associated with the regulation of multiple downstream signaling pathways, including MAPK and YAP/TAZ. In this way, Piezo1 may, to some extent, link mechanical stimuli to changes in cellular functions, including gene expression, inflammatory responses, and tissue remodeling.44 A schematic overview of Piezo1-mediated mechanotransduction and its potentially related signaling pathways is shown in Figure 2.
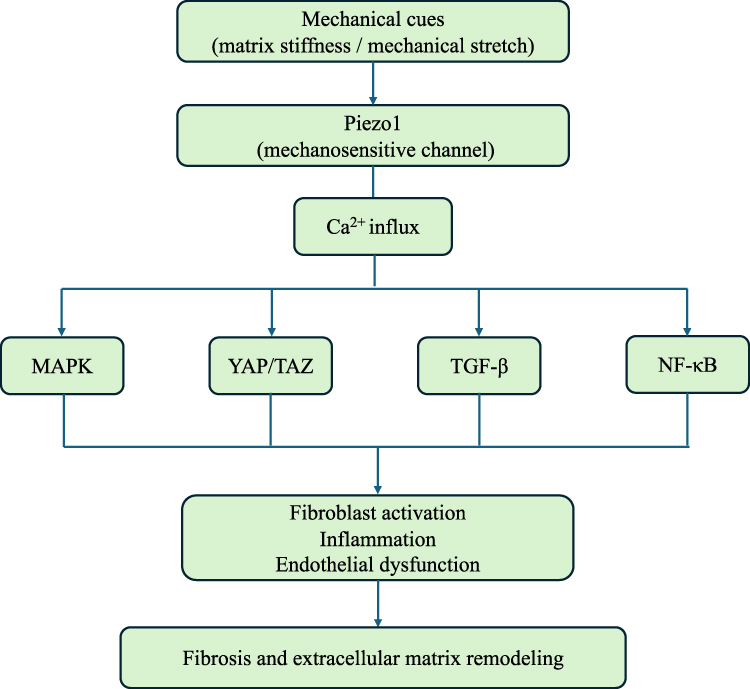 |
Figure 2 Potential Piezo1-mediated mechanotransduction pathways involved in tissue remodeling and fibrosis. Mechanical stimuli such as matrix stiffness and tissue stretch may activate the mechanosensitive ion channel Piezo1, leading to Ca2⁺ influx and activation of downstream signaling pathways including MAPK, YAP/TAZ, TGF-β, and NF-κB, which are associated with inflammation, vascular regulation, and fibrotic remodeling. |
Functionally, Piezo1 is predominantly expressed in various non-sensory tissues and has been implicated in vascular development,45 erythrocyte volume regulation,46 maintenance of epithelial homeostasis,47 and the development of the nervous and urinary systems.48,49 Recent studies suggest that Piezo1 may contribute to tissue fibrosis and chronic injury in multiple contexts, with its activation associated with fibroblast contraction and increased ECM deposition. For example, in models of skin fibrosis, Piezo1 has been reported to participate in the regulation of Wnt2/Wnt11–CCL24-related signaling.50 Similar findings have also been observed in studies of fibrosis in organs such as the kidney, lung, and heart.51 In these processes, Piezo1 can sense membrane tension and changes in the ECM, mediate Ca2⁺ influx, and may interact with several canonical signaling pathways, including TGF-β and YAP/TAZ, thereby contributing to inflammatory responses and tissue remodeling.15,52
It should be noted that current evidence regarding Piezo1 function is largely derived from in vitro experiments and animal models, and its role may vary across different tissues. In addition, mechanotransduction itself involves the coordinated regulation of multiple pathways. Piezo1 is therefore more likely to act as one regulatory node within this network rather than as the sole upstream driver. Thus, although its role in fibrosis and chronic injury has attracted increasing attention, its specific mechanisms and functions in particular diseases, such as CRD, remain to be further clarified.
Potential Molecular Mechanisms of Piezo1 in CRD Following Breast Cancer Treatment
In the context of an abnormal mechanical microenvironment, cells rely on multiple mechanosensing mechanisms to detect and respond to external stimuli. As a mechanosensitive cation channel, Piezo1 can be activated by membrane tension or mechanical stretch and is associated with the influx of cations such as Ca2⁺, thereby contributing to the regulation of multiple mechanotransduction-related signaling pathways, including MAPK and YAP/TAZ.44 Recent studies suggest that Piezo1 may participate in the sensing of and adaptation to changes in the mechanical environment; however, its specific role is likely to depend on the cell type and pathological context rather than following a single linear regulatory pathway.
During the development and progression of CRD, radiation injury can induce DNA damage, impaired epithelial renewal, and epithelial–mesenchymal transition (EMT), accompanied by fibroblast activation and increased ECM deposition, ultimately leading to tissue remodeling and increased local stiffness.20,28,53 These changes may collectively create an abnormal mechanical microenvironment and provide sustained stimulation for mechanotransduction signaling. Clinical and skin biophysical studies after breast cancer radiotherapy have also shown alterations in the biophysical properties of irradiated breast or chest wall tissues, reduced tissue elasticity, and RIF. These disease-specific changes support the possibility of long-term mechanical microenvironment abnormalities in CRD.19,23 In this context, Piezo1-related signaling may be involved in these processes; however, current assumptions are mainly extrapolated from other tissues or disease models, and direct evidence in CRD remains lacking.
In terms of inflammatory regulation, existing studies have shown that Piezo1 can participate in the modulation of mechanically induced inflammatory signaling in immune cells. In macrophages, Piezo1 activation is associated with Ca2⁺ influx, activation of the NF-κB pathway, and increased expression of proinflammatory cytokines.54,55 Under inflammatory stimuli such as IFN-γ or LPS, Piezo1 activation is linked to Ca2⁺ influx and the regulation of inflammation-related signaling pathways, including NF-κB, thereby upregulating proinflammatory mediators such as TNF-α and IL-6, enhancing the proinflammatory macrophage phenotype, and suppressing IL-4- and IL-13-mediated reparative responses.54,55 In addition, Piezo1 activation can promote adhesion between monocytes and endothelial cells and upregulate the expression of the adhesion molecule ICAM-1 and the transcriptional regulator YAP1. Studies have shown that YAP1 inhibitors can significantly attenuate Yoda1-induced proinflammatory responses, suggesting that Piezo1 may be involved in YAP1/ICAM-1–related signaling regulation.56,57 Meanwhile, Piezo1-mediated activation of NF-κB signaling can also increase the secretion of multiple chemokines, such as CCL20 and CXCL1, thereby recruiting and activating Th17 cells and further affecting epidermal cell proliferation, migration, and apoptosis.58 In the chronic inflammatory microenvironment of CRD, these mechanisms suggest that Piezo1 may contribute to the persistence of inflammation; however, this inference remains largely based on evidence from other research contexts.
In terms of vascular regulation, Piezo1 in vascular endothelial cells is considered to participate in the sensing of blood-flow shear stress and to be involved in the regulation of vascular-related signaling.59–61 Under conditions of altered shear stress or local mechanical stimulation, Piezo1 can be activated and mediate Ca2⁺ influx, thereby regulating the expression of multiple molecules involved in vascular remodeling. For example, Piezo1 activation is associated with ATP release and purinergic receptor–related signaling, and may influence the PI3K/AKT pathway, thereby promoting eNOS phosphorylation and activation and increasing NO production, which contributes to vasodilation and blood-flow regulation.62 In addition, Piezo1-mediated Ca2⁺ signaling can upregulate the expression of MMP-2 and MT1-MMP, promoting endothelial cell migration, alignment, and capillary-like structure formation, thereby contributing to angiogenic sprouting and vascular remodeling.63,64 In CRD, the combined effects of radiation injury and altered mechanical conditions may reshape the mechanical stimuli experienced by endothelial cells. Piezo1-related signaling may therefore influence vascular adaptive responses. However, these mechanisms are still mainly derived from vascular biology studies, and their specific role in CRD remains to be further validated.
In terms of fibrotic remodeling, Piezo1 may participate in fibroblast responses to mechanical stimulation. Previous studies have shown that, under conditions of increased matrix stiffness or mechanical stretch, Piezo1-related Ca2⁺ signaling can interact with profibrotic pathways such as TGF-β, thereby influencing fibroblast differentiation and ECM synthesis.22 Meanwhile, Piezo1-related signaling may also cooperate with the YAP/TAZ pathway in cellular responses to matrix stiffening, promoting cell proliferation and the expression of genes associated with ECM synthesis.65,66 These findings suggest that Piezo1 may have a regulatory role in fibrotic processes, although the available evidence is still mainly derived from other fibrosis models.
Notably, the mechanical microenvironment in breast cancer radiotherapy–related CRD is characterized by marked complexity and dynamic changes. The aforementioned factors, including surgical scar formation, radiotherapy-induced fibrosis, and impaired lymphatic drainage, may act together to disrupt local tissue tension distribution and increase interstitial pressure, thereby creating a persistently changing mechanical stimulus environment.29,30 In this context, cellular mechanisms that sense and respond to mechanical signals may play an important role in disease progression. As a potential regulatory node within the mechanotransduction network, Piezo1 may participate in converting these abnormal mechanical stimuli into intracellular signals and influence related cellular responses.
Taken together, Piezo1-related mechanotransduction signaling may be involved in inflammatory responses, vascular alterations, and fibrotic remodeling in CRD. However, current evidence is primarily derived from other disease models, and its specific role in CRD remains to be further validated. Therefore, understanding CRD from the perspective of the mechanical microenvironment and its signaling network may provide a new framework for future mechanistic studies, although its clinical significance should still be interpreted with caution.
Preclinical Evidence Supporting Piezo1 as a Potential Mechanistic Therapeutic Target in CRD Following Breast Cancer RT
Direct evidence supporting an interventional role for Piezo1 in breast cancer–related CRD after surgery is currently lacking. Nevertheless, preclinical studies in skin fibrosis, scar formation, and inflammation-related diseases suggest that modulation of Piezo1 activity is often associated with reduced fibrosis and attenuated inflammation, providing indirect support for its potential role as a mechanistic therapeutic target. From the perspective of CRD itself, persistent fibrosis, microvascular injury, tissue induration, and reduced compliance represent key features of the chronic stage. These pathological changes show a degree of biological overlap with the mechanical stimulation, matrix stiffness, and tissue remodeling processes investigated in Piezo1-related studies.27,28
Progressive fibrosis and ECM remodeling are among the major features of CRD-related pathology, and similar processes are also commonly observed in hypertrophic scarring and various models of skin fibrosis. Previous studies have shown that Piezo1 can participate in fibroblast responses to mechanical stimulation and is associated with myofibroblast differentiation and ECM deposition. In in vitro and animal models, mechanical stretch is accompanied by changes in Piezo1-related Ca2⁺ signaling and fibroblast phenotypic transition, whereas Piezo1 inhibition with GsMTx4 or genetic knockdown is associated with reduced secretory activity and attenuation of the scar phenotype.67 In addition, He et al found that increased matrix stiffness can activate Piezo1 and is associated with activation of the Wnt2/Wnt11–CCL24 axis and an enhanced fibrotic phenotype After Piezo1 knockout in HDFs cultured under high-stiffness conditions, CCR3, the receptor for CCL24, was downregulated, and si-Piezo1 treatment also reduced the rate of cell proliferation, suggesting that Piezo1 inhibition can markedly attenuate fibrosis.50 Xu et al reported that the mechanosensitive ion channel Piezo1 is highly expressed in periostin⁺ myofibroblasts and promotes myofibroblast activation and ECM deposition through Ca2⁺ influx and YAP/TAZ signaling activation In a bleomycin-induced mouse model of pulmonary fibrosis, gadolinium chloride (GdCl3)-mediated inhibition of Piezo1 significantly suppressed stretch-activated currents, whereas siRNA-mediated Piezo1 knockdown almost completely abolished this effect.65 These findings suggest that Piezo1 may participate in the regulatory link between the mechanical microenvironment and fibrotic responses, although its specific role in CRD remains largely inferred from cross-model evidence.
In addition to fibrosis, persistent inflammation is also an important driver of CRD progression. Similar inflammatory amplification processes have been observed in various immune- and inflammation-related diseases. Recent studies have shown that Piezo1 can act as a mechanosensor in immune cells, including macrophages, and participate in the regulation of inflammatory signaling. In several inflammatory or fibrotic models, inhibition of Piezo1 activity has been shown to reduce proinflammatory cytokine expression and attenuate tissue injury. Atcha et al reported that the mechanosensitive ion channel Piezo1 mediates macrophage sensing of changes in matrix stiffness and promotes NF-κB activation and M1 macrophage polarization through Ca2⁺-dependent signaling In BMDMs, treatment with Piezo1 siRNA, GdCl3, or GsMTx4 suppressed IFNγ/LPS-induced inflammatory responses while enhancing IL-4/IL-13-mediated healing responses, suggesting an important role for Piezo1 in mechanical microenvironment–mediated regulation of inflammation and tissue remodeling.55 He et al further reported that, in a CCl4-induced mouse model of liver fibrosis, macrophage-specific deletion of Piezo1 significantly reduced inflammation and TGF-β1 expression, thereby inhibiting hepatic stellate cell activation, decreasing ECM deposition, and ultimately attenuating the progression of liver fibrosis68 In these models, myeloid/macrophage-specific Piezo1 deficiency not only alleviated inflammation and reduced M1 polarization but also indirectly mitigated fibrosis.55,68 Taken together, these findings suggest that, in breast cancer radiotherapy–related CRD, Piezo1 may be associated not only with fibrosis but also with inflammatory amplification, and may therefore represent a potential interventional target. However, its specific role in the chronic inflammatory microenvironment of CRD remains unsupported by direct evidence.
Microvascular injury and hemodynamic abnormalities are also important pathological features of breast cancer radiotherapy–related CRD, and similar vascular dysfunction is widely observed in various ischemic, inflammatory, and vascular remodeling–related diseases. Existing studies have shown that Piezo1 is a key mechanosensor through which endothelial cells detect blood-flow shear stress, and that it plays an important role in vasodilation, endothelial inflammation, and vascular remodeling. Li et al found that disturbed flow can activate endothelial Piezo1 to induce Ca2⁺ influx and NF-κB activation, upregulate VCAM-1/ICAM-1 expression, and thereby promote early atheroprone inflammation Piezo1 knockdown or inhibition significantly reduced plaque formation and suppressed endothelial inflammation.69 Friedrich et al further showed that mechanical stimulation can activate Piezo1-mediated Ca2⁺ influx and upregulate MT1-MMP, thereby promoting endothelial cell migration and tube formation and enhancing angiogenesis, whereas inhibition of Piezo1 or MT1-MMP markedly attenuated this effect70 These studies suggest that Piezo1 may exert context-dependent regulatory effects in the microvasculature: aberrant activation may contribute to endothelial inflammation and vascular injury, whereas appropriate signaling may support flow sensing and vascular remodeling. Similar mechanisms may exist in CRD, although their specific role remains to be clarified in future studies.
Taken together, although direct interventional studies of Piezo1 in breast cancer–related CRD after surgery are currently lacking, modulation of Piezo1 activity has been associated with protective effects in multiple preclinical models involving fibrosis, inflammatory regulation, and vascular remodeling. Therefore, the existing evidence more strongly supports Piezo1 as a potential mechanistic interventional target rather than an established clinically translatable therapeutic target. Future basic and clinical studies focused specifically on CRD are still needed to further evaluate its precise role and potential translational value.
Limitations and Controversies
Although recent studies increasingly suggest that Piezo1 may play an important role in mechanotransduction and tissue remodeling, its specific function in breast cancer postoperative radiotherapy–related CRD remains subject to several limitations and uncertainties. Therefore, the discussion of Piezo1 in this review should be understood as a mechanistic hypothesis based on CRD-specific pathological changes and evidence from related fields, rather than as a summary of an established causal relationship.
First, current evidence regarding Piezo1 in CRD remains limited and is mainly derived from related fields such as wound repair, organ fibrosis, and vascular biology.50,66,67 Direct studies examining changes in Piezo1 expression and its functional role during chronic remodeling of the skin and subcutaneous tissues after breast radiotherapy are still relatively scarce. Therefore, current understanding relies largely on extrapolation from cross-disease models, and its applicability to the specific pathological context of CRD should be interpreted with caution. In addition, differences among studies in experimental models, modes of mechanical stimulation, and outcome measures further limit the comparability and integration of the available evidence.
Second, mechanotransduction itself is a highly complex and dynamically regulated network process. In addition to Piezo1, multiple mechanosensitive molecules and systems, including integrins, transient receptor potential (TRP) channels, the cytoskeleton, and YAP/TAZ in the Hippo pathway, participate in the sensing and amplification of mechanical stimuli.14,59 During radiotherapy-induced chronic tissue remodeling, crosstalk and compensatory mechanisms may exist among these pathways, making it difficult to define the relative contribution of a single molecule to the overall mechanical response. Moreover, it remains unclear whether Piezo1 acts as an upstream driver in this process or primarily functions as a sensor of changes in ECM stiffness that contributes to downstream signal amplification. This causal relationship still requires further validation.15,50
Third, a clear gap remains between different levels of existing evidence. Most studies on Piezo1 are based on in vitro experiments or animal models, and their extrapolation to the complex setting of chronic radiation injury in humans remains limited. This limitation is particularly relevant for CRD, a pathological process shaped by the combined effects of surgical trauma, radiation injury, and long-term tissue remodeling, in which the role of a single signaling pathway may be substantially influenced by temporal dynamics and tissue-specific factors.19,27,28
In addition, Piezo1 is widely expressed across multiple tissues and cell types, including vascular endothelial cells, immune cells, and fibroblasts, which poses challenges for its clinical translation as a potential therapeutic target. Systemic modulation of Piezo1 may interfere with normal mechanosensing and vascular homeostasis; therefore, achieving local, specific, and controllable regulation remains an important unresolved issue. Meanwhile, most current Piezo1-targeted interventions remain at the experimental stage, and their specificity and feasibility for in vivo application require further evaluation.
Taken together, although Piezo1 provides an important clue for understanding the role of the mechanical microenvironment in chronic radiation-induced skin injury, its specific function and clinical translational value in breast cancer–related CRD after surgery remain to be validated through more systematic and disease-specific studies.
Future Perspectives
Future studies may further clarify the role of Piezo1-mediated mechanotransduction in chronic tissue remodeling within the specific pathological context of breast cancer postoperative radiotherapy–related CRD. Unlike radiation-induced skin injury at other anatomical sites, the postoperative chest wall region in breast cancer patients is characterized by surgical scar formation, abundant adipose tissue, and altered local tissue tension. These features may collectively shape a unique mechanical microenvironment and promote ECM deposition, increased tissue stiffness, and persistent fibrosis during the chronic stage.19,27,28 On this basis, future research should further define the specific mode of Piezo1 involvement in this process, including its dynamic changes across different pathological stages and its regulatory position within the broader mechanotransduction network.
At the in vitro level, three-dimensional skin-equivalent models incorporating multiple cellular components, such as keratinocytes, fibroblasts, vascular endothelial cells, and adipose-related cells, could be established. Alternatively, patient-derived organoids and skin organoid systems may be used to simulate the chronic tissue microenvironment after breast radiotherapy under controlled conditions.71–73 At the same time, hydrogels with tunable stiffness or other biomaterial-based platforms could be used to construct culture systems with different mechanical properties, allowing more precise evaluation of how changes in matrix stiffness affect Piezo1 activation and its downstream signaling pathways.8,59
For mechanistic validation, future studies should place greater emphasis on establishing causal relationships. For example, strategies such as genetic knockout or knockdown, overexpression, and rescue experiments could be used to evaluate the effects of Piezo1 modulation on fibrotic phenotypes, inflammatory responses, and related signaling pathway activity under different mechanical conditions. These approaches would also help clarify the interactions between Piezo1 and signaling axes such as YAP/TAZ, TGF-β, and cytoskeletal remodeling.14,15
At the in vivo level, it will be necessary to establish animal models that more closely reflect the clinical setting, such as combining local radiotherapy with mastectomy or chest wall injury to simulate chronic tissue remodeling driven by the combined effects of surgery and radiotherapy. In parallel, genetically modified animals or local gene-intervention strategies could be used to dynamically evaluate the role of Piezo1 in chronic radiation-induced skin injury and to explore its stage-specific functions during disease progression.65,68
At the translational level, given the broad expression of Piezo1 across multiple tissues, systemic intervention may carry potential safety risks; therefore, targeted strategies based on local microenvironmental modulation may be more feasible. Future studies could explore local injection, hydrogel dressings, or biomaterial-based delivery systems to deliver Piezo1-modulating agents, small-molecule inhibitors, or nucleic acid therapeutics specifically to chronically injured post-radiotherapy tissues, thereby enabling precise regulation of abnormal mechanotransduction pathways.8,63 By integrating clinical sample analyses, three-dimensional models, animal experiments, and other multilevel research approaches, future work may further clarify the role of Piezo1 in breast cancer radiotherapy–related CRD and provide a theoretical basis for the development of novel local therapeutic strategies.
Conclusion
Breast cancer postoperative radiotherapy–related CRD is a complex chronic tissue injury process driven by persistent inflammation, microvascular injury, and progressive fibrosis, and its long-term evolution is often accompanied by substantial structural remodeling and functional alterations. Although several clinical interventions targeting inflammation, microcirculation, and fibrosis are currently available, their overall efficacy remains limited, suggesting that key pathogenic mechanisms have not yet been fully elucidated.
Recent studies have increasingly recognized that ECM deposition and increased tissue stiffness after radiation injury can markedly alter the local mechanical microenvironment and may contribute to inflammatory regulation, vascular dysfunction, and fibrotic progression through mechanotransduction mechanisms. As a mechanosensitive ion channel, Piezo1 can convert mechanical stimuli into intracellular signals and exert regulatory effects in various forms of tissue injury and remodeling. However, current understanding of Piezo1 in breast cancer radiotherapy–related CRD is still largely derived from other diseases or experimental models, and direct evidence supporting its specific role in this condition remains lacking. In addition, mechanotransduction involves multiple interconnected regulatory pathways, and the precise position of Piezo1 within this network, as well as its relationship with other mechanosensing mechanisms, remains to be further clarified. Therefore, caution is warranted when interpreting Piezo1 as a single key regulatory factor. From a translational perspective, regulation of the abnormal mechanical microenvironment and its associated signaling pathways may provide a new research direction for CRD intervention. Future studies should systematically evaluate the effects of mechanical environmental changes on related signaling pathways in model systems that more closely reflect the clinical setting, and further establish their causal roles in chronic tissue remodeling. Meanwhile, precision intervention strategies based on local microenvironmental modulation, particularly targeted approaches combined with biomaterials or local delivery systems, may have potential application prospects, although their safety and feasibility require further validation.
Overall, re-examining the pathological process of CRD from the perspective of the mechanical microenvironment and mechanotransduction mechanisms may help expand current understanding of disease development and progression. At the same time, it also highlights important gaps in the existing evidence that should be systematically addressed in future studies.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this study.
Author Contributions
Zhiyao Wang: Writing – original draft; Writing – review & editing; Project administration; Conceptualization. Feiyu Chen: Writing – original draft; Visualization. Zhi Li: Writing – review & editing; Visualization; Methodology; Conceptualization; Project administration; Supervision. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Zhiyao Wang and Feiyu Chen share first authorship.
Funding
This work was supported by Shanxi Basic Research Program Free Exploration Category Project (Grant Number: 202303021221197) and Clinical Key Specialty Program of Shanxi Bethune Hospital.
Disclosure
The authors declare that they have no competing interests.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–15. PMID: 33538338. doi:10.3322/caac.21660
2. Kong W, Wang Z, Liu Y, Pei J, Guo F. The anterior intercostal artery perforator flap in immediate oncoplastic breast reconstruction: current applications and future perspectives. Front Oncol. 2026;16:1765679. PMID: 41727632; PMCID: PMC12920241. doi:10.3389/fonc.2026.1765679
3. Darby S, McGale P; Early Breast Cancer Trialists’ Collaborative Group (EBCTCG). Effect of radiotherapy after breast-conserving surgery on 10-year recurrence and 15-year breast cancer death: meta-analysis of individual patient data for 10,801 women in 17 randomised trials. Lancet. 2011;378(9804):1707–1716. PMID: 22019144; PMCID: PMC3254252. doi:10.1016/S0140-6736(11)61629-2
4. Whelan TJ, Pignol JP, Levine MN, et al. Long-term results of hypofractionated radiation therapy for breast cancer. N Engl J Med. 2010;362(6):513–520. PMID: 20147717. doi:10.1056/NEJMoa0906260
5. Behroozian T, Bonomo P, Patel P, et al; Multinational Association of Supportive Care in Cancer (MASCC) Oncodermatology Study Group Radiation Dermatitis Guidelines Working Group. Multinational Association of Supportive Care in Cancer (MASCC) clinical practice guidelines for the prevention and management of acute radiation dermatitis: international Delphi consensus-based recommendations. Lancet Oncol. 2023;24(4):e172–e185. PMID: 36990615. doi:10.1016/S1470-2045(23)00067-0
6. Burke G, Faithfull S, Probst H. Radiation induced skin reactions during and following radiotherapy: a systematic review of interventions. Radiography. 2022;28(1):232–239. doi:10.1016/j.radi.2021.09.006 PMID: 34649789.
7. Ma L, Chen Y, Gong Q, et al. Cold atmospheric plasma alleviates radiation-induced skin injury by suppressing inflammation and promoting repair. Free Radic Biol Med. 2023;204:184–194. PMID: 37172912. doi:10.1016/j.freeradbiomed.2023.05.002
8. Cicchetti A, Mangili P, Fodor A, et al. Skin dose-volume predictors of moderate-severe late side effects after whole breast radiotherapy. Radiother Oncol. 2024;194:110183. PMID: 38423138. doi:10.1016/j.radonc.2024.110183
9. Spałek M. Chronic radiation-induced dermatitis: challenges and solutions. Clin Cosmet Invest Dermatol. 2016;9:473–482. PMID: 28003769; PMCID: PMC5161339. doi:10.2147/CCID.S94320
10. Allen BD, Limoli CL. Breaking barriers: neurodegenerative repercussions of radiotherapy induced damage on the blood-brain and blood-tumor barrier. Free Radic Biol Med. 2022;178:189–201. PMID: 34875340; PMCID: PMC8925982. doi:10.1016/j.freeradbiomed.2021.12.002
11. Kaidar-Person O, Marks LB, Jones EL. Pentoxifylline and vitamin E for treatment or prevention of radiation-induced fibrosis in patients with breast cancer. Breast J. 2018;24(5):816–819. doi:10.1111/tbj.13044 PMID: 29687536.
12. Finkelstein S, Kanee L, Behroozian T, et al. Comparison of clinical practice guidelines on radiation dermatitis: a narrative review. Support Care Cancer. 2022;30(6):4663–4674. PMID: 35067732. doi:10.1007/s00520-022-06829-6
13. Fan M, Feng M, Yuan S. Chinese clinical practice guidelines for the prevention and treatment of radiation-induced dermatitis. Precis Radiat Oncol. 2023;7(3):160–172. doi:10.1002/pro6.1208 PMID: 40337200; PMCID: PMC11935096.
14. Wynn TA, Ramalingam TR. Mechanisms of fibrosis: therapeutic translation for fibrotic disease. Nat Med. 2012;18(7):1028–1040. doi:10.1038/nm.2807 PMID: 22772564; PMCID: PMC3405917.
15. Lin Y, Wen D, Chen K, et al. Piezo1 and tissue fibrosis: insights into its role and potential for modulation. Burns Trauma. 2025;13:tkaf054. PMID: 41245993; PMCID: PMC12616850. doi:10.1093/burnst/tkaf054
16. Duffield JS, Lupher M, Thannickal VJ, Wynn TA. Host responses in tissue repair and fibrosis. Annu Rev Pathol. 2013;8:241–276. PMID: 23092186; PMCID: PMC3789589. doi:10.1146/annurev-pathol-020712-163930
17. Wilkinson L, Gathani T. Understanding breast cancer as a global health concern. Br J Radiol. 2022;95(1130):20211033. doi:10.1259/bjr.20211033 PMID: 34905391; PMCID: PMC8822551.
18. Dejonckheere CS, Dejonckheere E, Layer JP, et al. Barrier films for the prevention of acute radiation dermatitis in breast cancer: a systematic review and meta-analysis of randomised controlled trials. Breast. 2023;71:31–41. PMID: 37473629; PMCID: PMC10404536. doi:10.1016/j.breast.2023.07.001
19. Chakraborty MA, Lee SF, Wong HCY, et al. Chronic radiation dermatitis in breast cancer patients: pathophysiology, prevention and management strategies, and clinical impact. Ann Palliat Med. 2025;14(3):269–282. PMID: 40511570. doi:10.21037/apm-24-158
20. Campisi J. Cell biology: the beginning of the end. Nature. 2014;505(7481):35–36. Erratum in: Nature. 2014 Jan 9;505(7482):168. PMID: 24352243; PMCID: PMC4167797. doi:10.1038/nature12844
21. Vano-Galvan S, Fernandez-Lizarbe E, Truchuelo M, et al. Dynamic skin changes of acute radiation dermatitis revealed by in vivo reflectance confocal microscopy. J Eur Acad Dermatol Venereol. 2013;27(9):1143–1150. PMID: 22882643. doi:10.1111/j.1468-3083.2012.04680.x
22. Glover D, Harmer V. Radiotherapy-induced skin reactions: assessment and management. Br J Nurs. 2014;23(4):S28, S30–5. doi:10.12968/bjon.2014.23.Sup2.S28 PMID: 24619051.
23. Hu SC, Hou MF, Luo KH, et al. Changes in biophysical properties of the skin following radiotherapy for breast cancer. J Dermatol. 2014;41(12):1087–1094. PMID: 25354814. doi:10.1111/1346-8138.12669
24. Tewary G, Freyter B, Al-Razaq MA, et al. Immunomodulatory effects of histone variant H2A.J in Ionizing radiation dermatitis. Int J Radiat Oncol Biol Phys. 2024;118(3):801–816. PMID: 37758068. doi:10.1016/j.ijrobp.2023.09.022
25. Ryan JL. Ionizing radiation: the good, the bad, and the ugly. J Invest Dermatol. 2012;132(3 Pt 2):985–993. doi:10.1038/jid.2011.411 PMID: 22217743; PMCID: PMC3779131.
26. Amber KT, Shiman MI, Badiavas EV. The use of antioxidants in radiotherapy-induced skin toxicity. Integr Cancer Ther. 2014;13(1):38–45. doi:10.1177/1534735413490235 PMID: 23729466.
27. Fijardo M, Kwan JYY, Bissey PA, Citrin DE, Yip KW, Liu FF. The clinical manifestations and molecular pathogenesis of radiation fibrosis. EBioMedicine. 2024;103:105089. PMID: 38579363; PMCID: PMC11002813. doi:10.1016/j.ebiom.2024.105089
28. Wang Y, Chen S, Bao S, et al. Deciphering the fibrotic process: mechanism of chronic radiation skin injury fibrosis. Front Immunol. 2024;15:1338922. PMID: 38426100; PMCID: PMC10902513. doi:10.3389/fimmu.2024.1338922
29. Poulsen MG, Riddle B, Keller J, Porceddu SV, Tripcony L. Predictors of acute grade 4 swallowing toxicity in patients with stages III and IV squamous carcinoma of the head and neck treated with radiotherapy alone. Radiother Oncol. 2008;87(2):253–259. doi:10.1016/j.radonc.2008.03.010 PMID: 18410976.
30. McLaughlin SA, Wright MJ, Morris KT, et al. Prevalence of lymphedema in women with breast cancer 5 years after sentinel lymph node biopsy or axillary dissection: objective measurements. J Clin Oncol. 2008;26(32):5213–5219. Erratum in: J Clin Oncol. 2010 Apr 1;28(10):1808. PMID: 18838709; PMCID: PMC2652091. doi:10.1200/JCO.2008.16.3725
31. Bray FN, Simmons BJ, Wolfson AH, Nouri K. Acute and chronic cutaneous reactions to ionizing radiation therapy. Dermatol Ther. 2016;6(2):185–206. doi:10.1007/s13555-016-0120-y PMID: 27250839; PMCID: PMC4906114.
32. Hindley A, Zain Z, Wood L, et al. Mometasone furoate cream reduces acute radiation dermatitis in patients receiving breast radiation therapy: results of a randomized trial. Int J Radiat Oncol Biol Phys. 2014;90(4):748–755. Erratum in: Int J Radiat Oncol Biol Phys. 2015 Mar 15;91(4):882. PMID: 25585779. doi:10.1016/j.ijrobp.2014.06.033
33. Ulff E, Maroti M, Serup J, Falkmer U. A potent steroid cream is superior to emollients in reducing acute radiation dermatitis in breast cancer patients treated with adjuvant radiotherapy. A randomised study of betamethasone versus two moisturizing creams. Radiother Oncol. 2013;108(2):287–292. doi:10.1016/j.radonc.2013.05.033 PMID: 23827771.
34. Meier EL, Mink van der Molen DR, Lansdorp CA, et al. Hyperbaric oxygen therapy for local late radiation toxicity in breast cancer patients: a systematic review. Breast. 2023;67:46–54. PMID: 36587606; PMCID: PMC9982272. doi:10.1016/j.breast.2022.12.009
35. Lanigan SW, Joannides T. Pulsed dye laser treatment of telangiectasia after radiotherapy for carcinoma of the breast. Br J Dermatol. 2003;148(1):77–79. doi:10.1046/j.1365-2133.2003.04861.x PMID: 12534598.
36. Rigotti G, Marchi A, Galiè M, et al. Clinical treatment of radiotherapy tissue damage by lipoaspirate transplant: a healing process mediated by adipose-derived adult stem cells. Plast Reconstr Surg. 2007;119(5):1409–1422. PMID: 17415234. doi:10.1097/01.prs.0000256047.47909.71
37. Delanian S, Porcher R, Balla-Mekias S, Lefaix JL. Randomized, placebo-controlled trial of combined pentoxifylline and tocopherol for regression of superficial radiation-induced fibrosis. J Clin Oncol. 2003;21(13):2545–2550. doi:10.1200/JCO.2003.06.064 PMID: 12829674.
38. Jacobson G, Bhatia S, Smith BJ, Button AM, Bodeker K, Buatti J. Randomized trial of pentoxifylline and vitamin E vs standard follow-up after breast irradiation to prevent breast fibrosis, evaluated by tissue compliance meter. Int J Radiat Oncol Biol Phys. 2013;85(3):604–608. doi:10.1016/j.ijrobp.2012.06.042 PMID: 22846413.
39. Hoang TT, Vu VQ, Trinh DT. Management of radiation-induced ulcers by singlestage reconstructive surgery: a prospective study. Ann Burns Fire Disasters. 2019;32(4):294–300. PMID: 32431580; PMCID: PMC7197913.
40. Ge J, Li W, Zhao Q, et al. Architecture of the mammalian mechanosensitive Piezo1 channel. Nature. 2015;527(7576):64–69. PMID: 26390154. doi:10.1038/nature15247
41. Zhao Q, Zhou H, Chi S, et al. Structure and mechanogating mechanism of the Piezo1 channel. Nature. 2018;554(7693):487–492. Erratum in: Nature. 2018 Nov;563(7730)E19 PMID: 29469092. doi:10.1038/nature25743
42. Guo YR, MacKinnon R. Structure-based membrane dome mechanism for Piezo mechanosensitivity. eLife. 2017;6:e33660. PMID: 29231809; PMCID: PMC5788504. doi:10.7554/eLife.33660
43. Saotome K, Murthy SE, Kefauver JM, Whitwam T, Patapoutian A, Ward AB. Structure of the mechanically activated ion channel Piezo1. Nature. 2018;554(7693):481–486. Erratum in: Nature. 2022 Jul;607(7919) PMID: 29261642; PMCID: PMC6010196. doi:10.1038/nature25453
44. Coste B, Mathur J, Schmidt M, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science. 2010;330(6000):55–60. PMID: 20813920; PMCID: PMC3062430. doi:10.1126/science.1193270
45. Retailleau K, Duprat F, Arhatte M, et al. Piezo1 in smooth muscle cells is involved in hypertension-dependent arterial remodeling. Cell Rep. 2015;13(6):1161–1171. PMID: 26526998. doi:10.1016/j.celrep.2015.09.072
46. Cahalan SM, Lukacs V, Ranade SS, Chien S, Bandell M, Patapoutian A. Piezo1 links mechanical forces to red blood cell volume. eLife. 2015;4:e07370. PMID: 26001274; PMCID: PMC4456639. doi:10.7554/eLife.07370
47. Eisenhoffer GT, Loftus PD, Yoshigi M, et al. Crowding induces live cell extrusion to maintain homeostatic cell numbers in epithelia. Nature. 2012;484(7395):546–549. PMID: 22504183; PMCID: PMC4593481. doi:10.1038/nature10999
48. Pathak MM, Nourse JL, Tran T, et al. Stretch-activated ion channel Piezo1 directs lineage choice in human neural stem cells. Proc Natl Acad Sci U S A. 2014;111(45):16148–16153. PMID: 25349416; PMCID: PMC4234578. doi:10.1073/pnas.1409802111
49. Murthy SE, Loud MC, Daou I, et al. The mechanosensitive ion channel Piezo2 mediates sensitivity to mechanical pain in mice. Sci Transl Med. 2018;10(462):eaat9897. PMID: 30305457; PMCID: PMC6709986. doi:10.1126/scitranslmed.aat9897
50. He J, Cheng X, Fang B, Shan S, Li Q. Mechanical stiffness promotes skin fibrosis via Piezo1-Wnt2/Wnt11-CCL24 positive feedback loop. Cell Death Dis. 2024;15(1):84. doi:10.1038/s41419-024-06466-3 PMID: 38267432; PMCID: PMC10808102.
51. Chen J, Rodriguez M, Miao J, et al. Mechanosensitive channel Piezo1 is required for pulmonary artery smooth muscle cell proliferation. Am J Physiol Lung Cell Mol Physiol. 2022;322(5):L737–L760. PMID: 35318857; PMCID: PMC9076422. doi:10.1152/ajplung.00447.2021
52. Tu S, Li Y, Li J, et al. Mechanical stretch-mediated fibroblast activation: the pivotal role of Piezo1 channels. Biochim Biophys Acta Mol Cell Res. 2025;1872(7):120008. PMID: 40517844. doi:10.1016/j.bbamcr.2025.120008
53. Kumar R, Griffin M, Adigbli G, Kalavrezos N, Butler PE. Lipotransfer for radiation-induced skin fibrosis. Br J Surg. 2016;103(8):950–961. doi:10.1002/bjs.10180 PMID: 27169866.
54. Atcha H, Jairaman A, Holt JR, et al. Mechanically activated ion channel Piezo1 modulates macrophage polarization and stiffness sensing. Nat Commun. 2021;12(1):3256. PMID: 34059671; PMCID: PMC8167181. doi:10.1038/s41467-021-23482-5
55. Wang Y, Zhang Z, Yang Q, et al. Immunoregulatory role of the mechanosensitive ion channel Piezo1 in inflammation and cancer. Molecules. 2022;28(1):213. PMID: 36615408; PMCID: PMC9822220. doi:10.3390/molecules28010213
56. Zhu B, Li F, Yu J, et al. PIEZO1 mediates matrix stiffness-induced tumor progression in kidney renal clear cell carcinoma by activating the Ca2+/Calpain/YAP pathway. Biochim Biophys Acta Mol Cell Res. 2025;1872(1):119871. PMID: 39490703. doi:10.1016/j.bbamcr.2024.119871
57. Duan J, Li H, Zhang J, et al. PIEZO1 affects cell growth and migration via microfilament-mediated YAP trans-latitudinal regulation. Anal Chem. 2025;97(1):147–156. PMID: 39729436. doi:10.1021/acs.analchem.4c03420
58. Li W, Ze K, He X, et al. PIEZO1 promotes psoriasis-like skin inflammation in mice via NF-κB/IL-17 signaling pathway activation. Mol Med. 2025;31(1):225. PMID: 40495117; PMCID: PMC12150513. doi:10.1186/s10020-025-01279-2
59. Liu H, Hu J, Zheng Q, et al. Piezo1 channels as force sensors in mechanical force-related chronic inflammation. Front Immunol. 2022;13:816149. PMID: 35154133; PMCID: PMC8826255. doi:10.3389/fimmu.2022.816149
60. Rode B, Shi J, Endesh N, et al. Piezo1 channels sense whole body physical activity to reset cardiovascular homeostasis and enhance performance. Nat Commun. 2017;8(1):350. PMID: 28839146; PMCID: PMC5571199. doi:10.1038/s41467-017-00429-3
61. Douguet D, Patel A, Xu A, Vanhoutte PM, Honoré E. Piezo ion channels in cardiovascular mechanobiology. Trends Pharmacol Sci. 2019;40(12):956–970. doi:10.1016/j.tips.2019.10.002 PMID: 31704174.
62. Jiang M, Zhang YX, Bu WJ, et al. Piezo1 channel activation stimulates ATP production through enhancing mitochondrial respiration and glycolysis in vascular endothelial cells. Br J Pharmacol. 2023;180(14):1862–1877. PMID: 36740831. doi:10.1111/bph.16050
63. Lin D, Chai Y, Ma Y, Duan B, Yuan Y, Liu C. Rapid initiation of guided bone regeneration driven by spatiotemporal delivery of IL-8 and BMP-2 from hierarchical MBG-based scaffold. Biomaterials. 2019;196:122–137. PMID: 29449015. doi:10.1016/j.biomaterials.2017.11.011
64. Langlois S, Nyalendo C, Di Tomasso G, et al. Membrane-type 1 matrix metalloproteinase stimulates cell migration through epidermal growth factor receptor transactivation. Mol Cancer Res. 2007;5(6):569–583. doi:10.1158/1541-7786.MCR-06-0267 PMID: 17541067.
65. Xu L, Li T, Cao Y, et al. PIEZO1 mediates periostin+ myofibroblast activation and pulmonary fibrosis in mice. J Clin Invest. 2025;135(11):e184158. PMID: 40454481; PMCID: PMC12126248. doi:10.1172/JCI184158
66. Liu W, Zou H, Wei J, Dong L, Hou W. Piezo1: the potential novel target for radiation-induced liver fibrosis by regulating FAP + fibroblasts. Mol Cell Biochem. 2025;480(10):5297–5308. doi:10.1007/s11010-025-05327-7 PMID: 40526362; PMCID: PMC12515122.
67. He J, Fang B, Shan S, et al. Mechanical stretch promotes hypertrophic scar formation through mechanically activated cation channel Piezo1. Cell Death Dis. 2021;12(3):226. PMID: 33649312; PMCID: PMC7921104. doi:10.1038/s41419-021-03481-6
68. Luo S, Zhao X, Jiang J, et al. Piezo1 specific deletion in macrophage protects the progression of liver fibrosis in mice. Theranostics. 2023;13(15):5418–5434. Erratum in: Theranostics. 2025 Jan 1;15(3):1156–1157. PMID: 37908726; PMCID: PMC10614683. doi:10.7150/thno.86103
69. Lan Y, Lu J, Zhang S, et al. Piezo1-mediated mechanotransduction contributes to disturbed flow-induced atherosclerotic endothelial inflammation. J Am Heart Assoc. 2024;13(21):e035558. PMID: 39450718; PMCID: PMC11935715. doi:10.1161/JAHA.123.035558
70. Kang H, Hong Z, Zhong M, et al. Piezo1 mediates angiogenesis through activation of MT1-MMP signaling. Am J Physiol Cell Physiol. 2019;316(1):C92–C103. PMID: 30427721; PMCID: PMC6383143. doi:10.1152/ajpcell.00346.2018
71. Lakeh B, Shafiee A. Advancing dermatology with skin equivalents and organoids in pathophysiology and drug testing. Acta Biomater. 2025;207:120–130. PMID: 41072597. doi:10.1016/j.actbio.2025.10.008
72. Abaci HE, Guo Z, Coffman A, et al. Human skin constructs with spatially controlled vasculature using primary and iPSC-derived endothelial cells. Adv Healthc Mater. 2016;5(14):1800–1807. PMID: 27333469; PMCID: PMC5031081. doi:10.1002/adhm.201500936
73. Wang Z, Hou M, Pei J, Gao F, Li Z. Skin Organoids in Diabetic Chronic Wounds: current Status and Future Perspectives. Diabetes Metab Syndr Obes. 2026;19:598026. PMID: 42021845; PMCID: PMC13096021. doi:10.2147/DMSO.S598026
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.