Back to Journals » Biologics: Targets and Therapy » Volume 18
Cholinesterase Inhibitor Reveals Synergistic Potential for Neural Stem Cell-Based Therapy in the 5xFAD Mouse Model of Alzheimer’s Disease
Authors Wu CC ![]() , Lee YK
, Lee YK ![]() , Tsai JK, Su YT, Ho YC, Chu TH, Chen KT
, Tsai JK, Su YT, Ho YC, Chu TH, Chen KT ![]() , Chang CL, Chen JS
, Chang CL, Chen JS
Received 3 August 2024
Accepted for publication 26 November 2024
Published 3 December 2024 Volume 2024:18 Pages 363—375
DOI https://doi.org/10.2147/BTT.S489683
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Shein-Chung Chow
Cheng-Chun Wu,1,2,* Yung-Kuo Lee,3,* Jui-Kang Tsai,4 Yu-Ting Su,5 Yu-Cheng Ho,2 Tian-Huei Chu,3 Kuang-Ti Chen,6,7 Chen-Lin Chang,3,4 Jui-Shen Chen1,8
1Graduate Institute of Medicine, College of Medicine, I-Shou University, Kaohsiung City, Taiwan; 2School of Medicine, College of Medicine, I-Shou University, Kaohsiung City, Taiwan; 3Medical Laboratory, Medical Education and Research Center, Kaohsiung Armed Forces General Hospital, Kaohsiung City, Taiwan; 4Department of Psychiatry, Kaohsiung Armed Forces General Hospital, Kaohsiung City, Taiwan; 5Department of Obstetrics and Gynecology, Kaohsiung Chang Gung Memorial Hospital and Chang Gung University College of Medicine, Kaohsiung City, Taiwan; 6School of Medicine, College of Medicine, China Medical University, Taichung City, Taiwan; 7Department of Veterinary Medicine, Nation Chung-Hsing University, Taichung City, Taiwan; 8Department of Neurosurgery, E-DA Hospital, I-Shou University, Kaohsiung City, Taiwan
*These authors contributed equally to this work
Correspondence: Chen-Lin Chang, Medical Laboratory, Medical Education and Research Center, Kaohsiung Armed Forces General Hospital, No. 2, Zhongzheng 1st Road, Lingya Dist, Kaohsiung, 802301, Taiwan, Tel +886-7-7496751-726528, Fax +886-7-7496751, Email [email protected] Jui-Shen Chen, Department of Neurosurgery, E-DA Hospital, I-Shou University, No. 1, E-Da Road, Yan-Cao Dist, Kaohsiung, 824005, Taiwan, Tel +886-7-6151100-7963, Fax +886-7-6151100, Email [email protected]
Background and Objectives: Stem cell therapy shows great promise for treating Alzheimer’s disease (AD). Cholinesterase inhibitors (ChEIs) like donepezil are well-established for alleviating AD symptoms. This study aimed to determine if combining ChEI treatment with stem cell therapy could improve therapeutic outcomes.
Methods: Neural stem cells (NSCs) were injected into the hippocampus of the 5xFAD AD mice using a stereotactic technique. Following this, donepezil or a placebo was administered for one month. We assessed behavioral improvements, survival and health of the grafts, and changes in synaptic density.
Results: The AD mice demonstrated cognitive impairment in both the Morris water maze and novel object recognition tests. In groups receiving stem cell therapy, donepezil enhanced the survival and neuronal differentiation of grafted NSCs, promoting the establishment of synaptic connections with the host brain. The combined treatment with donepezil and NSC transplantation more effectively increased synaptic density and improved behavioral performance in AD mice compared to NSC transplantation alone.
Conclusion: Combining ChEIs with NSC transplantation produces synergistic effects in AD treatment. This approach highlights the potential of integrating these therapies to develop more effective strategies for managing Alzheimer’s disease.
Keywords: Alzheimer’s disease, cholinesterase inhibitors, donepezil, neural stem cells, transplantation
Introduction
Alzheimer’s disease (AD) is a common neurodegenerative disorder closely linked to aging, characterized by the presence of amyloid-β (Aβ) plaques and neurofibrillary tangles.1 Currently, over 25 million people globally are affected by dementia, and by 2050, it’s anticipated that nearly 1 million new cases of Alzheimer’s dementia will emerge each year.2 Despite extensive research, effective treatments for AD remain elusive.
The cholinergic hypothesis suggests that the loss of cholinergic neurons in the basal forebrain and the subsequent decrease in central cholinergic transmission are central to the disease, it proposes that cognitive decline in AD is primarily due to impaired cholinergic signaling within the brain, resulting from a reduction in acetylcholine (ACh) levels and disrupted cholinergic pathways.3 Among the treatments approved by the US Food and Drug Administration, cholinesterase inhibitors (ChEIs) such as donepezil, galantamine, and rivastigmine are known to slow disease progression and offer neuroprotective benefits. In rodent models, donepezil has been shown to improve cognitive function,4,5 elevate brain-derived neurotrophic factor (BDNF) levels,6,7 reduce Aβ buildup,8 support neurogenesis, and restore synaptic density.8 However, the long-term effectiveness of donepezil in severe AD cases remains debated. This highlights the need for innovative combined therapies and identification of synergistic factors to improve treatment outcomes.
Stem cell transplantation is a promising strategy for treating various degenerative diseases,9 yet its success has not fully met expectations. Our previous studies revealed that neural stem cells (NSCs) engineered to overexpress brain-derived neurotrophic factor (BDNF-NSCs) could offer a new therapeutic approach for AD.10 Additionally, we found that granulocyte colony-stimulating factor (G-CSF) can mobilize bone marrow mesenchymal stem cells (BM-MSCs) to boost neurogenesis in an AD mouse model.11 These results underscore the critical role of neurogenesis in stem cell-based therapies for AD and the need for further strategies to maintain this neurogenic effect for effective treatment development.
Notably, cholinergic inputs play a crucial role in regulating neural stem cell (NSC) neurogenesis, particularly within brain regions such as the hippocampus and subventricular zone (SVZ).12 In the aged dentate gyrus, cholinergic activation has been shown to stimulate NSCs, promoting neurogenesis in a region that typically experiences age-related decline. Distinct populations of choline acetyltransferase-positive (ChAT+) neurons have been identified, which modulate NSC activity in an activity-dependent manner, particularly within the SVZ.13 Activation of cholinergic receptor pathways impacts apoptosis, cell proliferation, and neuronal differentiation, suggesting that cholinergic signaling exerts broad influence over the survival and maturation of NSCs.14 By enhancing NSC function and differentiation, cholinergic pathways contribute to the maintenance of neurogenesis and cognitive resilience, highlighting potential therapeutic implications for aging and neurodegenerative conditions. Consequently, we propose that combining cholinesterase inhibitor (ChEI) therapy with stem cell transplantation might create a synergistic effect in AD treatment, potentially improving behavioral outcomes and reducing synaptic loss in an AD mouse model.
Materials and Methods
Experimental Animals and Procedures
The Institute of Animal Care and Use Committee at I-Shou University (ISU), Taiwan, approved all experimental and animal care procedures (IACUC-ISU 109029, valid from 2021/11/01 to 2023/08/30). The 5-familial Alzheimer`s disease (5XFAD) transgenic mice (B6SJL-Tg(APPSwFlLon, PSEN1*M146L*L286V)6799Vas/Mmjax) were obtained from the Jackson Laboratory (Bar Harbor, ME). These mice were kept on a 12-hour light/dark cycle and were fed ad libitum throughout the experiment to ensure unrestricted access to food and water, following the guidelines set by the ISU Animal Center. Male 6-month-old heterozygous (Tg+/−) mice were used in the all studies, age-matched Tg−/− (non-Tg wild type) mice were served as the control. Neural stem cell (NSC) transplantation was conducted according to our established protocol.10 Isoflurane anesthesia was administered to mice for induction and maintenance. For induction, a concentration of 3–5% isoflurane in oxygen was used, and for maintenance, 1–2% isoflurane was delivered via a nose mask. Anesthesia was monitored by assessing vital signs, and recovery was rapid upon stopping of the anesthetic. For transplantation, 100,000 cells suspended in 2 μL of Hank’s Balanced Salt Solution (HBSS) were injected into the hippocampal dentate gyrus using a stereotaxic apparatus. After transplantation, AD mice were treated with daily intraperitoneal injections of donepezil (0.3 mg/kg, Sigma D6821, St. Louis, MO) for 4 weeks, while the control group received saline solution. Behavioral assessments were conducted using the Morris water maze, probe test, and novel object recognition.10 Body weight measurements were taken weekly until sacrifice, with data showing no significant differences between the experimental groups.
NSCs and Neuron Primary Culture
Green fluorescent protein (GFP)-expressing neural stem cells (NSCs) were obtained from a transgenic mouse line engineered to express “enhanced” GFP cDNA (Jackson Laboratories). These GFP-expressing NSCs were isolated from the hippocampi of postnatal day 1 pups and expanded in adherent culture according to our previously established protocol.10 Additionally, primary neurons were cultured from postnatal day 1 wild-type C57BL/6 mouse pups, using the technique detailed in.15
In vitro Condition Medium Assays
Conditioned media were prepared from neurons that were cultured for 11 days and treated with either 50 μM donepezil or PBS for 24 hours. After treatment, this conditioned media was transferred to cultures of neural stem cells (NSCs) exposed to Aβ toxicity and maintained for 5 days, with media changes every 2 days. Aβ was prepared using the method described in.16 Neurite outgrowth was assessed by counting cells that were positive for both MAP2 and DAPI. Images were captured with the ImageXpress System (Molecular Devices, Sunnyvale, CA), and the Neurite Outgrowth Module of the MetaXpress software (Molecular Devices) was used to analyze the number of neurons, total neurite length per cell, and the number of neurite processes and branches per cell.
Immunofluorescence (IF) Staining
For immunofluorescence (IF) staining, mice were first anesthetized and then perfused through the heart with PBS containing 4% paraformaldehyde (PFA). The brains were removed and fixed in 4% PFA for 2 hours, followed by dehydration. The tissue was then sectioned using a cryostat to prepare slices. The slides with tissue sections were incubated with various primary antibodies: GFP (Genetex GTX26662; Hsinchu, Taiwan), MAP2 (Abcam ab5392; Cambridge, UK), calbindin (Millipore PC253L; Burlington, MA), ChAT (Millipore AB143), synaptophysin (Millipore MAB5258A4), and PSD95 (Millipore MABN68). For detection, AlexaFluor-conjugated secondary antibodies (Thermo Fisher Scientific, Waltham, MA) were used. The sections were then counterstained with DAPI and mounted with a coverslip using mounting medium (Dako, Glostrup, Denmark). Quantification of immunoreactive cells was performed in a blinded manner. Data were collected from 20 randomly chosen images per mouse, using a 20x objective lens, from coronal brain sections.
Western Blotting
Tissue extracts were obtained from the hippocampus following our established protocol.10 Protein concentrations were measured using a Bioray assay. Proteins were then separated by electrophoresis on a 10% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and transferred to a membrane for Western blot analysis. Detection was carried out using primary antibodies against PSD95, synaptophysin, brain-derived neurotrophic factor (BDNF) (GeneTex GTX17884), and β-actin (Sigma A5316). The bands were visualized and quantified with ImageJ software. To ensure accurate comparison of protein levels, the intensity of each band was normalized to β-actin.
Statistical Analysis
All data are expressed as the mean ± standard error of the mean (SEM). Data with a normal distribution were analyzed using one-way ANOVA, followed by Tukey’s post hoc test for multiple comparisons, or Student’s t-test for comparing two groups. Data that did not follow a normal distribution were analyzed using the Kruskal–Wallis test with Dunn’s post hoc test for multiple comparisons, or the Mann–Whitney U-test for comparing two groups. Statistical significance was set at P < 0.05.
Results
Assessment of the Combined Effect of ChEI Treatment and Stem Cell Transplantation in Alzheimer’s Disease Mice
In the experimental design, five groups were established: (1) Wildtype (WT) mice + vehicle (Veh), (2) 5xFAD transgenic (Tg) mice + Veh, (3) Tg + donepezil, (4) Tg + NSC transplantation, and (5) Tg + NSC + donepezil. GFP-expressing NSCs were transplanted into the hippocampal dentate gyrus of 6-month-old Tg mice. Donepezil was administered via daily subcutaneous injections at a dose of 0.3 mg/kg for 4 weeks, with saline used as the vehicle control. Together, 4 weeks post-NSC transplantation and donepezil/vehicle administration, the mice underwent behavioral testing and were then sacrificed for subsequent biochemical analyses.
To determine the impact of combining cholinesterase inhibitor (ChEI) treatment with stem cell transplantation in Alzheimer’s disease (AD) mice, we first evaluated spatial memory in AD mice post-treatment using the Morris water maze task. Results demonstrated that AD mice showed significant cognitive impairment in escape latency. Both NSC transplantation and donepezil treatment individually improved escape latency, but the combination therapy provided an even greater recovery in spatial memory (Figure 1A). Next, we performed the probe test to measure memory retention and consolidation after escape training. Results indicated that the combination of NSCs and donepezil significantly enhanced memory retention and consolidation (Figure 1B and C). Additionally, the novel object recognition test was used to assess recognition abilities in AD mice following treatment. Data indicated that the combined therapy was the most effective approach for improving recognition abilities in AD mice (Figure 1D). Overall, these findings suggest that the combined therapy of NSCs and donepezil produces synergistic effects, offering superior efficacy in improving behavioral outcomes in AD mice compared to individual treatments.
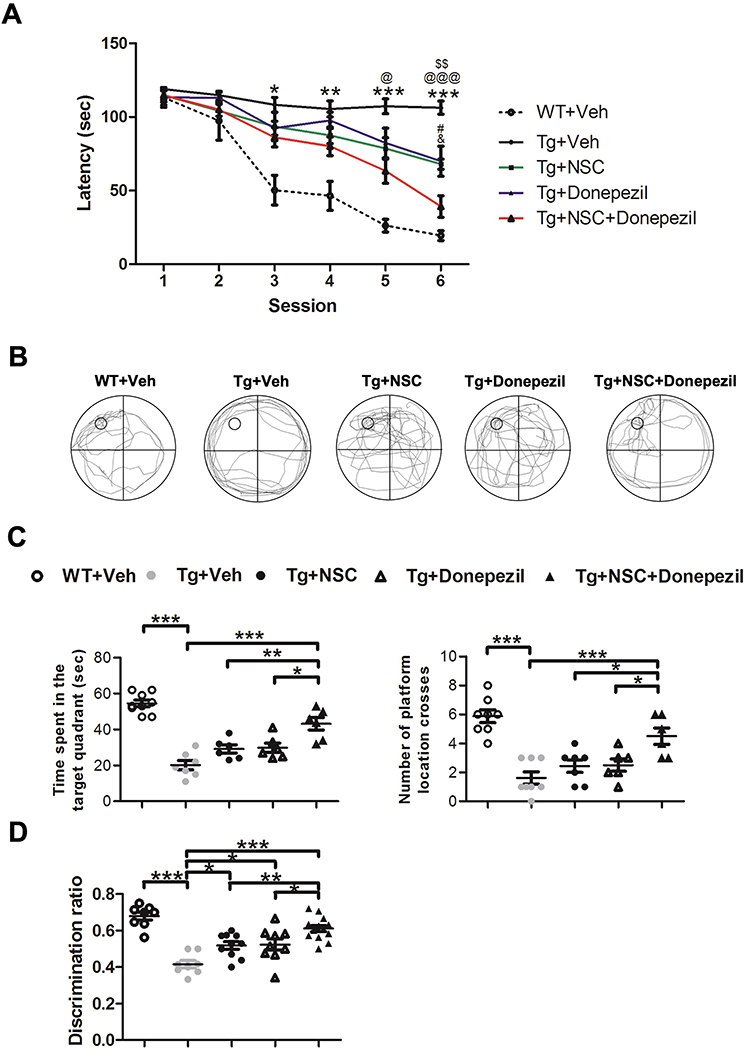 |
Figure 1 Donepezil treatment enhanced the effectiveness of NSC transplantation in improving behavioral outcomes in AD mice. (A) The Morris water maze escape latency was measured 4 weeks after NSC transplantation and donepezil treatment. *Transgenic (Tg) + Veh mice vs Wiltype (WT) + Vehicle (Veh) mice, @Tg + Veh mice vs Tg + NSC + Donepezil mice, $Tg + Veh mice vs Tg + NSC mice, #Tg + NSC mice vs Tg + NSC + Donepezil mice, &Tg + NSC mice vs Tg + NSC + Donepezil mice. Statistical significance is indicated by *, @, $, #, & for p < 0.05. **, $$ for p < 0.01. ***, @@@ for p < 0.001, as determined by the Kruskal–Wallis test with the Dunn post hoc test. (B) Representative paths from the probe trial test, captured from video recordings. (C) Quantitative results showing time spent in the target quadrant (left panel) and the number of times the hidden platform was crossed (right panel) in the probe test. (D) The novel object recognition test was conducted 4 weeks after NSC transplantation and donepezil treatment. Data are presented as mean ± SEM with N=6 per group. Statistical significance is indicated by * for p < 0.05, ** for p < 0.01, and *** for p < 0.001, as determined by the Kruskal–Wallis test with the Dunn post hoc test. |
Enhancement of Neural Stem Cell Survival and Neuronal Differentiation in Alzheimer’s Disease Mice by Donepezil Treatment
To understand how donepezil improves the effectiveness of combined therapy, we first investigated its impact on the survival and development of transplanted NSCs. We measured the number of surviving engrafted cells using immunofluorescence (IF) staining to detect GFP-expressing cells at 4 weeks after transplantation. Our results indicated that donepezil significantly enhanced the survival of these cells in the hippocampal dentate gyrus of Alzheimer’s disease (AD) mice (Figure 2A). We then analyzed the neuronal differentiation of the engrafted cells by staining for both GFP and the neuronal dendritic marker MAP2. Donepezil treatment led to a greater number of NSC-derived neurons and improved neuronal differentiation (Figure 2B). Furthermore, we assessed the potential for the engrafted cells to become granular neurons by staining for the granular cell marker calbindin (Figure 2C). The combination of donepezil with the NSCs resulted in increased cell survival and a higher proportion of granular neurons. These findings suggest that donepezil plays a key role in boosting the survival, neuronal differentiation, and development of granular neurons from transplanted NSCs in the AD brain.
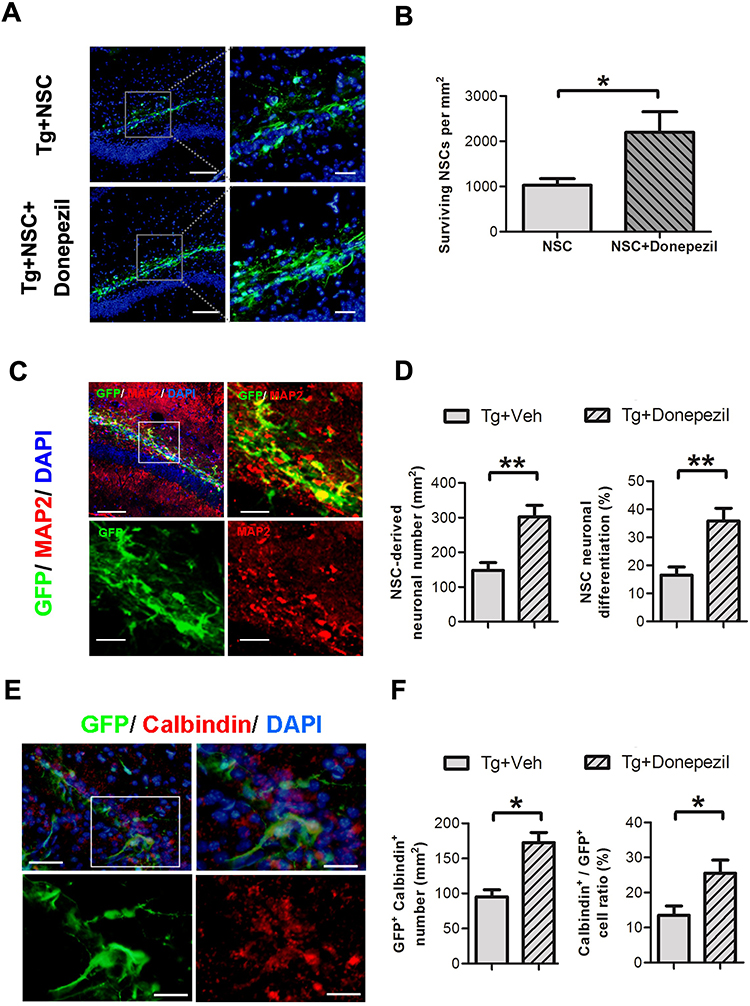 |
Figure 2 Donepezil treatment enhanced the survival, neuronal differentiation, and granular neuron formation of transplanted NSCs 4 weeks post-transplantation. (A) Representative images showing immunofluorescence (IF) staining of engrafted NSCs in the hippocampus of AD mice, with or without donepezil treatment. DAPI staining (blue) highlights the nuclei (left top and left bottom). Scale bar: 50 μm. The area outlined by the white dashed line is further magnified for presentation (right top and right bottom). Scale bar: 25 μm. (B) Quantification of green fluorescent protein (GFP)-expressing cells that survived, based on IF staining data (right panel). (C) IF staining for GFP and microtubule-associated protein 2 (MAP2) to assess neuronal differentiation 4 weeks after transplantation with or without donepezil treatment (left top). Scale bar: 50 μm. The area outlined by the white dashed line is further magnified for presentation (right top, left bottom, and right bottom). Scale bar: 25 μm. (D) Quantification of cells that are positive for both MAP2 and GFP (right panel). (E) IF staining for GFP and calbindin to evaluate granular neuron differentiation 4 weeks post-transplantation with or without donepezil treatment (left top). Scale bar: 50 μm. The area outlined by the white dashed line is further magnified for presentation (right top, left bottom, and right bottom). Scale bar: 25 μm. (F) Quantification of cells positive for both calbindin and GFP (right panel). Data are presented as mean ± SEM with N=6 per group. Statistical significance is indicated by * for p < 0.05 and ** for p < 0.01, as analyzed by the Mann–Whitney test. |
Donepezil Enhances Neural Stem Cell Survival and Differentiation Under Aβ Toxicity by Increasing ACh and BDNF Levels in Cultured Neurons
To explore how donepezil enhances the effects of transplanted neural stem cells (NSCs), we conducted a conditioned medium assay to analyze donepezil’s impact while NSCs present in Aβ toxicity. We first cultured primary neurons and evaluated their cell viability after donepezil treatment (Figure 3A). Furthermore, donepezil treatment elevated both ACh and brain-derived neurotrophic factor (BDNF) levels in these neurons (Figure 3B and C). We then collected conditioned medium from neurons treated with or without donepezil and applied it to NSCs exposed to Aβ toxicity, and conducted IF staining for MAP2 to evaluate levels of neuronal differentiation and neurite outgrowth (Figure 3D). The quantified results indicated that Aβ exposure severely impaired NSC survival, neuronal differentiation, and neurite outgrowth. However, the conditioned medium from donepezil-treated neurons preserved NSC survival and differentiation, and improved neurite outgrowth, including total neurite length and the number of branches. Importantly, the positive effects of the conditioned medium were reversed when ACh receptors of NSCs were blocked by atropine (Figure 3E). These results suggest that donepezil supports NSC survival, neuronal differentiation, and neurite development under Aβ toxicity by boosting ACh and BDNF levels from adjacent neurons.
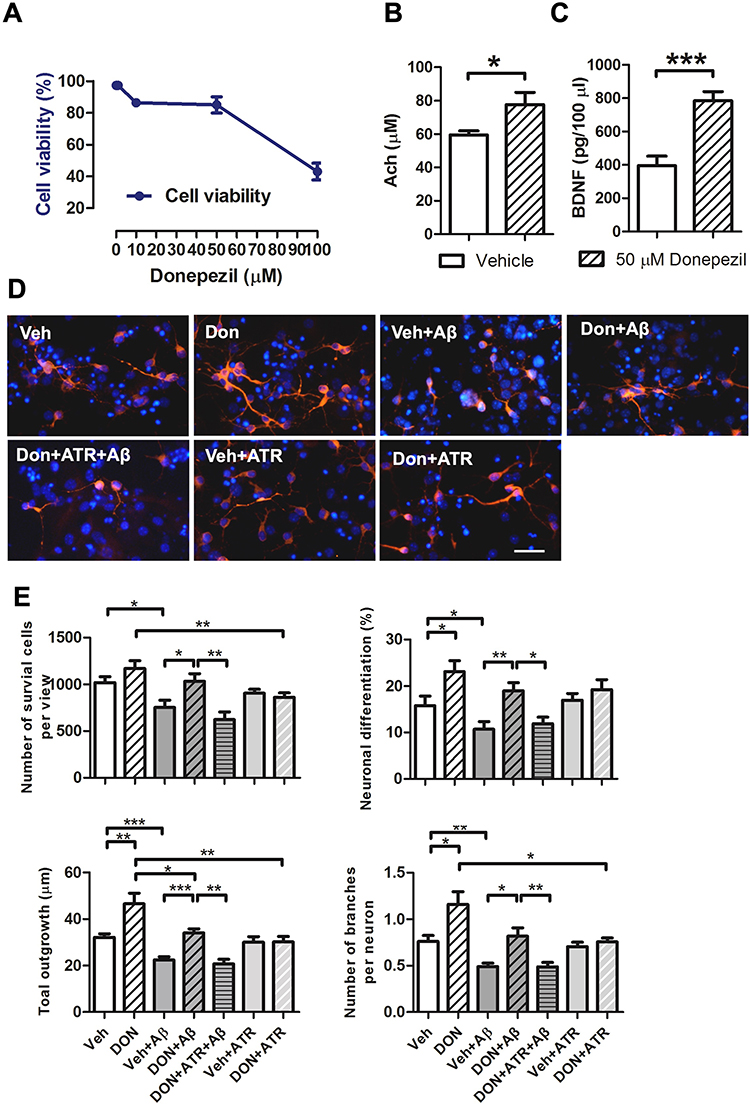 |
Figure 3 Donepezil-conditioned medium enhanced the survival, neuronal differentiation, and neurite outgrowth of NSCs exposed to Aβ toxicity. (A) Determination of the dose and cell viability for primary neuronal response to donepezil in vitro. (B) Enzyme-Linked Immunosorbent Assay (ELISA) results showing acetylcholine (ACh) levels in the culture medium from primary neurons with or without donepezil treatment. Data are presented as mean ± SEM with N=5 independent experiments per group. Statistical significance is indicated by * for p < 0.05, analyzed by the Mann–Whitney test. (C) ELISA results showing brain-derived neurotrophic factor (BDNF) levels in the culture medium from primary neurons with or without donepezil treatment. Data are presented as mean ± SEM with N=5 per group. Statistical significance is indicated by *** for p < 0.001, analyzed by the Mann–Whitney test. (D) Representative images of immunofluorescence (IF) staining for MAP2 in NSCs treated with Aβ for 5 days to assess neuronal differentiation. The effects of donepezil-conditioned medium on neuronal viability, differentiation, and neurite outgrowth were evaluated. Scale bar: 50 μm. (E) Quantification of cell viability, neuronal differentiation, total dendrite length, and number of branches per cell. Data are presented as mean ± SEM with N=5 independent experiments per group. Statistical significance is indicated by * for p < 0.05, ** for p < 0.01, and *** for p < 0.001, analyzed by the Kruskal–Wallis test with the Dunn post hoc test. Abbreviations: Veh, vehicle; Don, donepezil; ATR, atropine. |
Integration and Functionality of Engrafted Neural Stem Cells in the Hippocampal Circuits of Alzheimer’s Disease Mice
To explore the ability of engrafted cells to integrate into endogenous hippocampal circuits, we performed IF staining for GFP, PSD95, and ChAT in NSCs transplanted into AD mice, assessed 4 weeks post-transplantation (Figure 4A). The transplanted cells successfully integrated into the granular cell layer of the hippocampal dentate gyrus, extending dendrites into the molecular layer and expressing the post-synaptic marker PSD95. Notably, the PSD95 staining from engrafted cells colocalized with ChAT immunoreactivity, indicating that these cells are receiving cholinergic inputs from the host brain. This was observed in AD mice with or without donepezil treatment, suggesting that the engrafted cells can integrate into the hippocampal dentate gyrus and interact with the host’s cholinergic system.
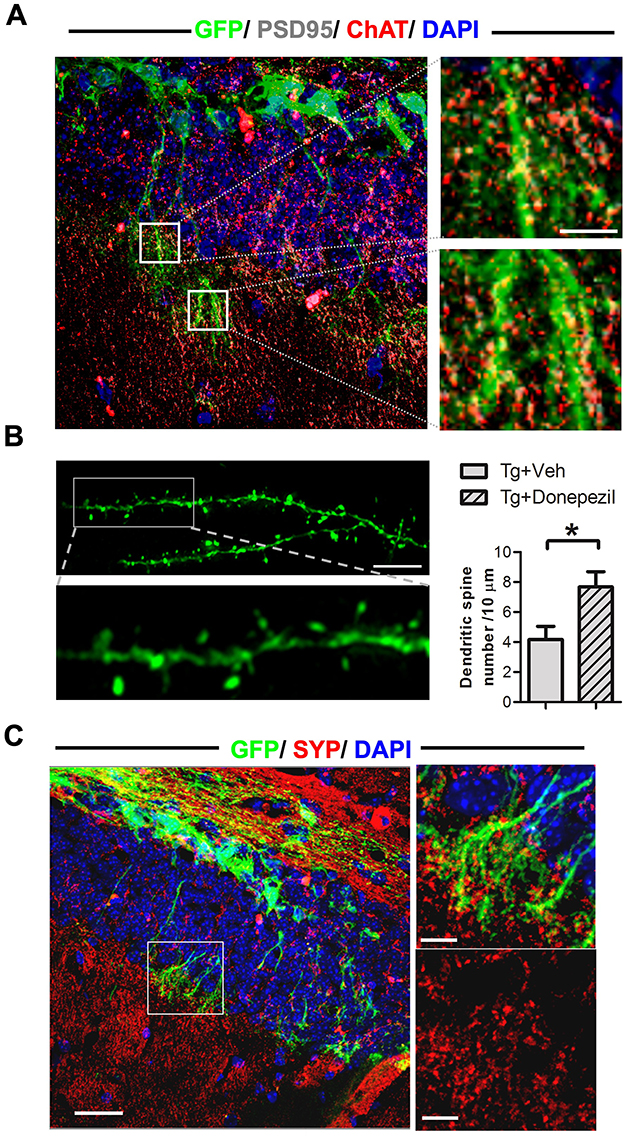 |
Figure 4 Engrafted cells received synaptic input from endogenous cholinergic neurons, developed dendritic spines, and established synaptic connections with native neurons in the brains of AD mice. (A) Left: IF staining for choline acetyl transferase (ChAT), postsynaptic density protein 95 (PSD95), and GFP in engrafted cells 4 weeks post-transplantation. Right: Magnified images from the boxed area in the left panel show PSD95 distribution on the dendrites of engrafted cells and its colocalization with ChAT immunoreactivity from endogenous neurons. Scale bar: 25 μm. (B) Left upper: Representative confocal microscopy image showing the structure of dendritic spines in engrafted cells. Scale bar: 10 μm. Left lower: Magnified images from the boxed area in the left upper panel. Right: Quantification of the number of dendritic spines in NSCs transplanted into AD mice, with or without donepezil treatment. Data are presented as mean ± SEM with N=6 per group. Statistical significance is indicated by * for p < 0.05, analyzed by the Mann–Whitney test. (C) Left: IF staining for GFP and synaptophysin (SYP) in NSCs transplanted into AD mice. Scale bar: 100 μm. Right Upper: Magnified image from the boxed area in the left panel. Scale bar: 25 μm. Right Lower: Magnified image focusing solely on SYP immunoreactivity from the same boxed area. Scale bar: 25 μm. |
To further investigate the potential of engrafted cells and the impact of donepezil treatment, we assessed dendritic spine formation in these cells 4 weeks post-transplantation. The engrafted cells displayed dendritic spine structures, and donepezil treatment enhanced dendritic spine density (Figure 4B), indicating an increased capacity for the development of excitatory synaptic structures.17 Additionally, we performed IF staining for GFP and synaptophysin to examine the capability of the engrafted cells to establish synaptic connections with host neurons. The co-localization of GFP immunoreactivity from engrafted cells and synaptophysin immunoreactivity from endogenous neurons was observed in NSC-transplanted mice, both with and without donepezil treatment (Figure 4C). These findings suggest that donepezil treatment enhances dendritic spine formation in engrafted cells and that these cells are capable of forming synaptic connections with the host brain circuits.
Synergistic Effects of Combined Therapy on Synaptic Density and BDNF Levels in Alzheimer’s Disease Mice
To further assess the contribution of engrafted cells to synaptic formation, we measured synaptic density in AD mice to evaluate the therapeutic potential of our strategies. Immunofluorescence (IF) staining for synaptophysin was conducted to quantify synaptic density. The data revealed that AD mice exhibited a significant reduction in synaptic density within the molecular layer of the hippocampus. Both donepezil injection and NSC transplantation alone had modest effects in promoting the recovery of synaptic density. However, the combination therapy demonstrated a significant improvement in synaptic density (Figure 5).
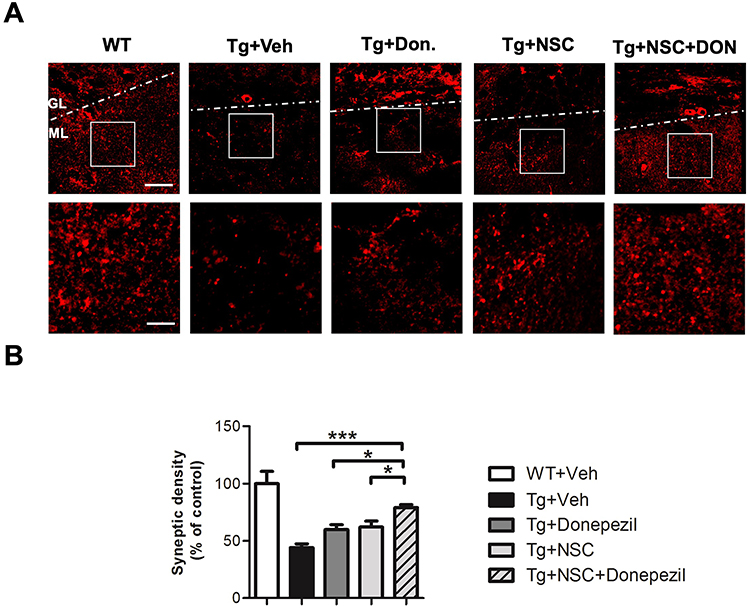 |
Figure 5 Combined donepezil and NSC transplantation treatment demonstrated the most effective recovery of synaptic density in AD mice. (A) Upper Panel: Representative confocal microscopy image showing SYP immunoreactivity in the hippocampus of AD mice 4 weeks after NSC transplantation. Scale bar: 50 μm. Lower Panel: Magnified images from the boxed region in the upper panel, highlighting details of SYP immunoreactivity. Scale bar: 25 μm. (B) Quantification of synaptic density from the immunofluorescence-stained images (lower panel). Data are shown as mean ± SEM (%), with N=6 per group. Statistical significance is indicated by * for p < 0.05 and *** for p < 0.001, analyzed by the Kruskal–Wallis test with the Dunn post hoc test. Abbreviations: GL, granule cell layer; ML, molecular layer. |
Additionally, we performed Western blot analysis on hippocampal lysates to measure the levels of synaptophysin, PSD95, and BDNF in vivo. The Western blot results were consistent with the immunostaining findings; the combined therapy showed a marked positive effect in mitigating synaptic loss in AD mice (Figure 6). Furthermore, BDNF levels were elevated in AD mice following the combined therapy. These results confirm that the combined therapy exerts a synergistic effect in ameliorating synaptic loss and increasing BDNF levels in AD mice.
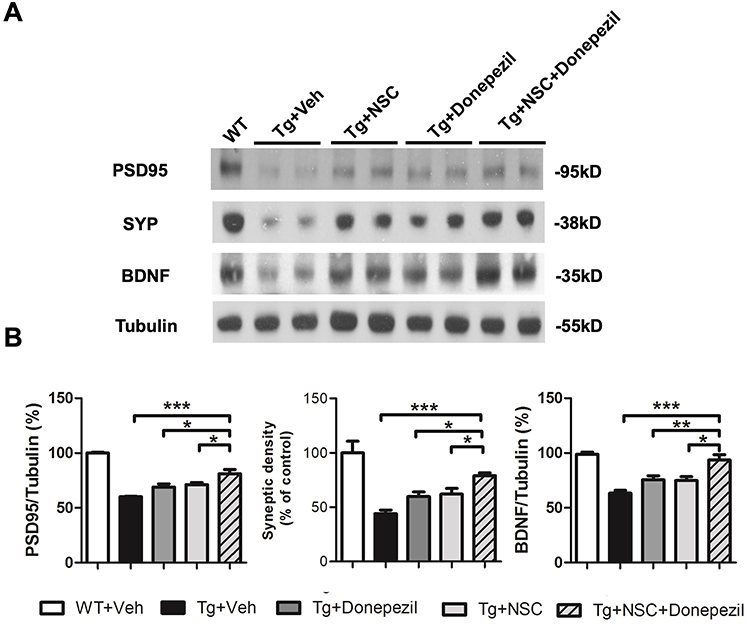 |
Figure 6 Combined donepezil treatment and NSC transplantation exhibited synergistic effects in restoring synaptic density and brain-derived trophic factor (BDNF) levels. (A) Representative Western blot images showing the expression of PSD95, SYP, and BDNF in the hippocampus 4 weeks after NSC transplantation. (B) Quantification of the protein levels for PSD95, SYP, and BDNF. Data are presented as mean ± SEM (%), with N=6 per group. Statistical significance is indicated by * for p < 0.05, ** for p < 0.01, and *** for p < 0.001, analyzed by the Kruskal–Wallis test with the Dunn post hoc test. |
Discussion
This study highlights the synergistic effects of combining donepezil treatment with NSC transplantation, demonstrating that this approach enhances cognitive function and restores synaptic density in AD mice. Notably, donepezil treatment improved both the survival and neuronal differentiation of the transplanted cells. To our knowledge, this is the first study to report such a synergistic effect between donepezil and NSC transplantation. Given the current lack of effective therapies for AD and the pressing healthcare needs, our findings offer valuable insights for developing new treatment strategies.
Emerging evidence indicates that impaired neurogenesis contributes to AD progression.18 Significant neuronal loss is observed even in early stages of AD,19 and various molecules involved in AD, such as ApoE, PS1, and APP, have been linked to the modulation of neurogenesis.20 Several existing drugs, including erythropoietin,21 granulocyte colony-stimulating factor,16 and certain antidepressants,22,23 have shown potential to promote neurogenesis in animal models. Notably, donepezil also appears to support neurogenesis,24 making it a promising candidate for AD intervention or prevention. Our previous research emphasized the importance of neurogenesis in NSC-based therapies for AD, particularly for the survival and integration of transplanted NSCs into the hippocampal circuitry.10,11 This suggests that the effectiveness of such therapies is closely tied to the neuronal fate of the transplanted cells. Our data demonstrate that donepezil treatment enhances cell survival and neuronal differentiation in AD mice, leading to significant improvements in behavioral outcomes. Additionally, environmental factors influence the survival, differentiation, and functional maturation of NSCs,25 and the adverse conditions in the AD brain, such as harmful proteins and inflammatory factors, can reduce the effectiveness of NSC transplantation.10 Our study shows that donepezil supplementation improves the survival, neuronal specification, and growth of engrafted cells, supporting the viability and effectiveness of NSC-based therapies for AD.
Cholinergic projections play a role in regulating the activity of resident NSCs, affecting their proliferation, survival, and neurogenesis.13 Acetylcholine (ACh) is known to influence neuronal differentiation during development. Both muscarinic and nicotinic ACh receptors affect processes like apoptosis, cellular proliferation, and neuronal differentiation by activating downstream signaling pathways such as Ras-mitogen activated protein kinase, phosphatidylinositol 3-kinase-Akt, protein kinase C, and c-Src.14 Our study shows that transplanted cells form synaptic connections with endogenous cholinergic terminals, and we propose that donepezil enhances these cells’ survival and differentiation through these signaling pathways.
Previous studies have shown that donepezil can increase levels of brain-derived neurotrophic factor (BDNF) and restore synaptic density,6–8,26 which are important for mitigating AD-related pathogenesis. We confirmed that donepezil treatment raises BDNF levels both in vivo and in vitro. We propose that BDNF, released by engrafted cells and enhanced by donepezil, plays a crucial role in the synergistic effects of combined donepezil and NSC transplantation. BDNF is vital for synaptic plasticity, neuronal survival, differentiation, and learning/memory processes.27 It also has neuroprotective properties, helping neurons resist Aβ toxicity and reducing Aβ deposition via the APOE receptor SORL1.28 Additionally, BDNF supports neurite outgrowth and synaptic plasticity.29 Given its role in synaptic plasticity, which is closely linked to AD progression and is reversible and dynamic,30–32 BDNF’s involvement in restoring synaptic density and cognitive function suggests that it is a key player in the combined therapeutic effects of donepezil and NSC transplantation.
Conclusions
Our findings indicate that donepezil enhances the effectiveness of NSC-based therapies. Donepezil treatment improves the survival and neuronal differentiation of transplanted NSCs by interacting with endogenous cholinergic neurons through synaptic connections. This results in a synergistic effect that leads to better behavioral outcomes in AD mice. Additionally, the combined therapy helps restore key pathological features, such as synaptic density and BDNF levels. Overall, these results offer valuable insights for developing new treatment strategies for AD and emphasize the potential of using ChEIs to boost the effectiveness of NSC-based therapies.
Abbreviations
AD, Alzheimer’s disease; Ach, Acetylcholine; AchR, Acetylcholine receptor; Aβ, β- amyloid; BDNF, Brain-derived neurotrophic factor; ChEI, Cholinesterase inhibitor; ChAT, Choline Acetyltransferase; GFP, green fluorescent protein; MAP2, microtubule associate protein 2; IF, immunofluorescent staining.
Data Sharing Statement
The authors confirm that all the data, materials, and methods will be available upon request.
Ethical Approval and Consent to Participate
The Institute of Animal Care and Use Committee of ISU, Taiwan approved all experimental procedures and animal care procedures. Mice were housed in a 12/12-hour light/dark cycle with food and water provided ad libitum in accordance with the guideline of Animal Center of ISU and maintained in the Animal Center at ISU.
Acknowledgments
The authors acknowledge the technical support by the Basic Medical Core Laboratory, I-Shou University College of Medicine.
Funding
This study was supported by grants from E-Da Hospital (EDDHP113002, EDAHP113003, EDAHP112003, EDPJ111061) and Kaohsiung Armed Forces General Hospital (KAFGH-D-113018, KAFGH-D-113032), Taiwan.
Disclosure
The authors all have no conflict of interest in this study.
References
1. Selkoe DJ, Hardy J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol Med. 2016;8:595–608. doi:10.15252/emmm.201606210
2. Association As. 2017 Alzheimer`s disease facts and figures. Alzheimers Dement. 2017;13:325–373.
3. Terry AV Jr, Buccafusco JJ. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: recent challenges and their implications for novel drug development. J Pharmacol Exp Ther. 2003;306:821–827. doi:10.1124/jpet.102.041616
4. Romberg C, Mattson MP, Mughal MR, Bussey TJ, Saksida LM. Impaired attention in the 3xTgAD mouse model of Alzheimer’s disease: rescue by donepezil (Aricept). J Neurosci. 2011;31:3500–3507. doi:10.1523/JNEUROSCI.5242-10.2011
5. Yuede CM, Dong H, Csernansky JG. Anti-dementia drugs and hippocampal-dependent memory in rodents. Behav Pharmacol. 2007;18:347–363. doi:10.1097/FBP.0b013e3282da278d
6. Johansson J, Formaggio E, Fumagalli G, Chiamulera C. Choline up-regulates BDNF and down-regulates TrkB neurotrophin receptor in rat cortical cell culture. Neuroreport. 2009;20:828–832. doi:10.1097/WNR.0b013e32832b7324
7. Leyhe T, Stransky E, Eschweiler GW, Buchkremer G, Laske C. Increase of BDNF serum concentration during donepezil treatment of patients with early Alzheimer’s disease. Eur Arch Psychiatry Clin Neurosci. 2008;258:124–128. doi:10.1007/s00406-007-0764-9
8. Dong H, Yuede CM, Coughlan CA, Murphy KM, Csernansky JG. Effects of donepezil on amyloid-beta and synapse density in the Tg2576 mouse model of Alzheimer’s disease. Brain Res. 2009;1303:169–178. doi:10.1016/j.brainres.2009.09.097
9. Fang Y, Gao T, Zhang B, Pu J. Recent advances: decoding Alzheimer’s disease with stem cells. Front Aging Neurosci. 2018;10:77. doi:10.3389/fnagi.2018.00077
10. Wu CC, Lien CC, Hou WH, Chiang PM, Tsai KJ. Gain of BDNF function in engrafted neural stem cells promotes the therapeutic potential for Alzheimer’s disease. Sci Rep. 2016;6:27358. doi:10.1038/srep27358
11. Wu CC, Wang IF, Chiang PM, Wang LC, Shen CJ, Tsai KJ. G-CSF-mobilized bone marrow mesenchymal stem cells replenish neural lineages in Alzheimer’s disease mice via CXCR4/SDF-1 chemotaxis. Mol Neurobiol. 2016;54:6198–6212. doi:10.1007/s12035-016-0122-x
12. Itou Y, Nochi R, Kuribayashi H, Saito Y, Hisatsune T. Cholinergic activation of hippocampal neural stem cells in aged dentate gyrus. Hippocampus. 2011;21:446–459. doi:10.1002/hipo.20761
13. Paez-Gonzalez P, Asrican B, Rodriguez E, Kuo CT. Identification of distinct ChAT(+) neurons and activity-dependent control of postnatal SVZ neurogenesis. Nat Neurosci. 2014;17:934–942. doi:10.1038/nn.3734
14. Resende RR, Adhikari A. Cholinergic receptor pathways involved in apoptosis, cell proliferation and neuronal differentiation. Cell Commun Signal. 2009;7:20. doi:10.1186/1478-811X-7-20
15. Majumdar D, Gao Y, Li D, Webb DJ. Co-culture of neurons and glia in a novel microfluidic platform. J Neurosci Methods. 2011;196:38–44. doi:10.1016/j.jneumeth.2010.12.024
16. Tsai KJ, Tsai YC, Shen CK. G-CSF rescues the memory impairment of animal models of Alzheimer’s disease. J Exp Med. 2007;204:1273–1280. doi:10.1084/jem.20062481
17. Lai KO, Ip NY. Structural plasticity of dendritic spines: the underlying mechanisms and its dysregulation in brain disorders. Biochim Biophys Acta. 2013;1832:2257–2263. doi:10.1016/j.bbadis.2013.08.012
18. Hollands C, Bartolotti N, Lazarov O. Alzheimer’s disease and hippocampal adult neurogenesis; exploring shared mechanisms. Front Neurosci. 2016;10:178. doi:10.3389/fnins.2016.00178
19. Gomez-Isla T, Price JL, McKeel DW Jr, Morris JC, Growdon JH, Hyman BT. Profound loss of layer II entorhinal cortex neurons occurs in very mild Alzheimer’s disease. J Neurosci. 1996;16:4491–4500. doi:10.1523/JNEUROSCI.16-14-04491.1996
20. Ghosal K, Stathopoulos A, Pimplikar SW. APP intracellular domain impairs adult neurogenesis in transgenic mice by inducing neuroinflammation. PLoS One. 2010;5:e11866. doi:10.1371/journal.pone.0011866
21. Hassouna I, Ott C, Wustefeld L, et al. Revisiting adult neurogenesis and the role of erythropoietin for neuronal and oligodendroglial differentiation in the hippocampus. Mol Psychiatry. 2016;21:1752–1767. doi:10.1038/mp.2015.212
22. Taupin P. Neurogenesis and the effect of antidepressants. Drug Target Insights. 2006;1:13–17. doi:10.1177/117739280600100005
23. Hanson ND, Owens MJ, Nemeroff CB. Depression, antidepressants, and neurogenesis: a critical reappraisal. Neuropsychopharmacology. 2011;36:2589–2602. doi:10.1038/npp.2011.220
24. Kwon KJ, Kim MK, Lee EJ, et al. Effects of donepezil, an acetylcholinesterase inhibitor, on neurogenesis in a rat model of vascular dementia. J Neurol Sci. 2014;347:66–77. doi:10.1016/j.jns.2014.09.021
25. Marutle A, Ohmitsu M, Nilbratt M, Greig NH, Nordberg A, Sugaya K. Modulation of human neural stem cell differentiation in Alzheimer (APP23) transgenic mice by phenserine. Proc Natl Acad Sci. 2007;104:12506–12511. doi:10.1073/pnas.0705346104
26. Srivareerat M, Tran TT, Salim S, Aleisa AM, Alkadhi KA. Chronic nicotine restores normal abeta levels and prevents short-term memory and E-LTP impairment in abeta rat model of Alzheimer’s disease. Neurobiol Aging. 2011;32:834–844. doi:10.1016/j.neurobiolaging.2009.04.015
27. Binder DK, Scharfman HE. Brain-derived neurotrophic factor. Growth Factors. 2004;22:123–131. doi:10.1080/08977190410001723308
28. Young JE, Boulanger-Weill J, Williams DA, et al. Elucidating molecular phenotypes caused by the SORL1 Alzheimer’s disease genetic risk factor using human induced pluripotent stem cells. Cell Stem Cell. 2015;16:373–385. doi:10.1016/j.stem.2015.02.004
29. Figurov A, Pozzo-Miller LD, Olafsson P, Wang T, Lu B. Regulation of synaptic responses to high-frequency stimulation and LTP by neurotrophins in the hippocampus. Nature. 1996;381:706–709. doi:10.1038/381706a0
30. McDole B, Isgor C, Pare C, Guthrie K. BDNF over-expression increases olfactory bulb granule cell dendritic spine density in vivo. Neuroscience. 2015;304:146–160. doi:10.1016/j.neuroscience.2015.07.056
31. Matsuzaki M, Honkura N, Ellis-Davies GC, Kasai H. Structural basis of long-term potentiation in single dendritic spines. Nature. 2004;429:761–766. doi:10.1038/nature02617
32. Trachtenberg JT, Chen BE, Knott GW, et al. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature. 2002;420:788–794. doi:10.1038/nature01273
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.