Back to Journals » Infection and Drug Resistance » Volume 19
Chikungunya Virus in 2025: Epidemiology, Immunopathogenesis, and Vaccine Development — A Narrative Review
Authors Zhao C, Ge Z, Zhang T, Jiang Z, Tian D, Chen Z ![]()
Received 12 September 2025
Accepted for publication 6 January 2026
Published 13 January 2026 Volume 2026:19 567134
DOI https://doi.org/10.2147/IDR.S567134
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Chenxi Zhao,* Ziruo Ge,* Tingyu Zhang, Zhouling Jiang, Di Tian, Zhihai Chen
National Key Laboratory of Intelligent Tracking and Forecasting for Infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhihai Chen, National Key Laboratory of Intelligent Tracking and Forecasting for infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, People’s Republic of China, Email [email protected] Di Tian, National Key Laboratory of Intelligent Tracking and Forecasting for infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, People’s Republic of China, Email [email protected]
Abstract: Chikungunya is an arboviral disease caused by infection with chikungunya virus (CHIKV), an alphavirus transmitted primarily by Aedes aegypti and Aedes albopictus. Over the past two decades, chikungunya has re-emerged across Africa, Asia, the Indian Ocean, Europe, and the Americas, with 119 countries and territories reporting local transmission up to 2024. This expansion reflects a widening geographic range with greater risk of introduction and spreads through mosquito vectors, leaving larger populations susceptible to infection. Following inoculation by an infected mosquito, viral replication leads to viremia and the abrupt onset of fever, severe joint pain with swelling, myalgia, and rash. These clinical manifestations are driven by the host immune response to infection, which also influence disease severity and clinical outcomes. Although most patients recover within weeks, a proportion develop persistent arthralgia or chronic arthritis lasting months to years, contributing to the overall disease burden. Although both the live-attenuated vaccine IXCHIQ and the virus-like particle vaccine VIMKUNYA have been approved, the license for IXCHIQ was recently suspended due to safety concerns. Recent outbreaks of chikungunya fever have driven sustained research, leading to deeper insights into disease pathogenesis, host immune responses and chronic inflammation. In this narrative review, we summarize recent advances up to 2025 in the epidemiology, host receptors, pathogenesis, clinical manifestations, immune responses, mother-to-child transmission, and vaccines development of CHIKV.
Keywords: chikungunya, arthritis, cytokine, mother-to-child transmission, vaccine
Introduction
Chikungunya was first described and epidemiologically documented during 1952–1953 in the Tanganyika Territory (present-day United Republic of Tanzania),1 after which sporadic cases and localized outbreaks were recorded across multiple African regions, reflecting early endemic circulation. In 1958, Thailand reported the first recorded urban outbreak in Asia.2 In 2004, an outbreak in Kenya initiated the spread to the Indian Ocean islands, and in 2005–2006 Réunion Island experienced a major epidemic with approximately 244,000 infections, bringing chikungunya virus (CHIKV) to prominence as a global public health concern.3 In 2007, the first reported autochthonous transmission event in Europe occurred in northern Italy, linked to an imported case from India.4 In 2013, CHIKV was detected in the Caribbean and then spread widely in the Americas.5 According to the Pan American Health Organization (PAHO)/World Health Organization (WHO), 411,560 confirmed cases with 515 deaths were reported in 2023,6 and 431,223 cases with 243 deaths were reported in 2024.7 In the first seven months of 2025, about 240,000 chikungunya cases and 90 related deaths had been reported across 16 countries and territories.8 As of August 23, 2025, 10,258 laboratory-confirmed cases were reported in Foshan, the largest recorded outbreak of chikungunya in China.9 Recent modeling study suggests that under current climate conditions, about 15% of global land area is suitable for CHIKV transmission, placing an estimated 6.78 billion people at risk, mostly in tropical and subtropical regions.10 As of December 2024, local transmission had been documented in 119 countries and territories worldwide, predominantly in Africa, Asia, and the Americas.11
This narrative review synthesizes current evidence on CHIKV, covering its epidemiology and transmission, viral structure and receptors, insights from animal models and human pathogenesis, clinical manifestations and immune responses, mother-to-child transmission, and vaccine development. Relevant literature was identified through targeted searches of PubMed and Web of Science, supplemented by key references and reports from international public health agencies. Epidemiological and vaccine-related data were reviewed up to August 2025, which served as the cutoff date for this review.
Epidemiology and Transmission
CHIKV is an enveloped positive-sense single-stranded RNA virus belonging to the genus Alphavirus within the family Togaviridae.12 Most alphavirus are broadly classified into two groups according to the clinical syndromes they cause: arthritogenic and encephalitic.13 The arthritogenic alphaviruses, CHIKV, Ross River virus (RRV), Barmah Forest virus (BFV), Sindbis virus (SINV), Mayaro virus (MAYV) and O’nyong-nyong virus (ONNV), predominantly induce acute and chronic arthralgia.14 The encephalitic alphaviruses, Eastern equine encephalitis virus (EEEV), Venezuelan equine encephalitis virus (VEEV) and Western equine encephalitis virus (WEEV), cause central nervous system disease such as meningitis and encephalitis.15 Arthritogenic alphaviruses are transmitted chiefly by Aedes mosquitoes and circulate in two epidemiologically distinct cycles.16
CHIKV is maintained in a rural enzootic (forest/savannah) transmission cycle between Aedes (Aedes africanus and Aedes furcifer) and vertebrate amplifying hosts, including nonhuman primates considered the principal reservoirs.17 A meta-analysis estimated CHIKV seroprevalence in nonhuman primates was as high as 35% (95% CI: 9–66%) in Africa, 7% (95% CI: 0–28%) in the Americas, and 6% (95% CI: 5–34%) in Asia.18 Moreover, CHIKV has been isolated from bats, rodents, palm squirrels, and birds, and CHIKV-specific antibodies have been detected in rats, birds, elephants, and reptiles, suggesting that these animals may participate in the natural transmission cycle of the virus and serve as potential reservoir hosts to some extent.19 However, evidence that other wild vertebrates, such as rodents, bats and birds, serve as maintenance hosts remains limited and inconsistent.20 Enzootic transmission mainly affects rural areas, with humans as incidental hosts infected via spillover mediated by sylvatic or bridge vectors. The urban cycle consists of sustained human–mosquito–human transmission driven by Aedes aegypti and Aedes albopictus,19 these species also transmit dengue virus (DENV), Zika virus (ZIKV), and yellow fever virus (YFV).21 Climate change and anthropogenic forces, such as land-use change, deforestation, urbanization, and human mobility, are expanding vector ranges and seasonal suitability, increasing opportunities for spillover from enzootic cycles and subsequent urban amplification.22 A recent systematic review of Aedes-borne arboviral infections in Europe (2000–2023) reported that climate change and international travel are key drivers of the increasing risk of autochthonous transmission by promoting vector expansion, enhancing viral replication within mosquitoes, and facilitating repeated viral introductions into previously unaffected regions.23 These trends are expected to increase the risk of arbovirus transmission and have prompted the exploration of emerging vector control strategies, including Wolbachia-based endosymbiont control, genetically modified mosquitoes, and integrated vector management.24,25
Following ingestion of an infectious blood meal, CHIKV replicate in the midgut epithelial cells of Aedes mosquitoes.26 Viral particles become detectable in saliva by 2 days post infection (dpi), are transmissible from this time point, and reach peak transmissibility around 6 dpi.27 CHIKV can disseminate to the ovaries, with virus detected in eggs by 6 dpi.28 Vertical transmission in Aedes mosquitoes has been demonstrated, with CHIKV persisting to F5 and F6 progeny, indicating maintenance of the virus across successive mosquito generations.29 This mode is considered an important mechanism for viral persistence under adverse environmental conditions. The desiccation-resistant eggs remain viable for long periods, allowing CHIKV retention and maintaining infectivity during dry seasons and winter when mosquito densities are low.30 Mavale et al observed that CHIKV-infected male Aedes mosquitoes transmitted the virus to females during mating, and the infected females subsequently infected suckling mice, highlighting the potential epidemiological significance of venereal transmission.31
Phylogenetic analyses classify CHIKV into three major lineages: West African, Asian, and East/Central/South African (ECSA), the latter of which includes the Indian Ocean lineage (IOL).32 Within the IOL, selection in Aedes albopictus has driven sequential adaptive changes in the envelope glycoproteins. The E1-A226V substitution increases Aedes albopictus infection by approximately 50~100-fold, followed by E2-L210Q, which provides an additional four to six-fold increase.33,34 These adaptive mutations markedly enhance transmission in Aedes albopictus while exerting little measurable effect on Aedes aegypti competence or on experimental models of human infection.35 Ongoing evolution of CHIKV envelope glycoproteins may therefore promote adaptation to more widely distributed vectors and facilitate further geographic spread.36 To date, no specific amino acid substitution has been linked to increased viral fitness in humans, such as higher viremia, greater clinical severity, or shifts in tissues and organs tropism.37
Viral Structure and Receptors
The CHIKV genome is approximately 11.8 kb and contains 5′ and 3′ untranslated regions and two open reading frames (ORFs) separated by a short junction region. The 5′ ORF is translated from genomic RNA to yield the non-structural polyprotein P1234, which is proteolytically processed into nsP1, nsP2, nsP3, nsP4.38 The nsP3 is a key component of the plasma-membrane–associated replication complex and forms cytoplasmic condensates/tubular networks that concentrate genomic RNA, capsid and requisite host factors, thereby promoting efficient replication and infection.39 The 3′ ORF is expressed from a subgenomic RNA and encodes the structural polyprotein that is processed into capsid (C), envelope (E)3, E2, 6K, and E1.40,41 The capsid protein is essential for selectively packaging the viral genome.42 E1 contains a hydrophobic fusion loop that drives membrane fusion process,43 while E2 is a major target of neutralizing antibodies that inhibit attachment, membrane fusion, and subsequent viral entry or egress.44 The E2 forms a complex with the E1, and three E2-E1 complexes form a trimeric envelope protein.45 E3 stabilizes the E2-E1 heterodimer and prevents premature exposure of the E1 fusion loop.43 CHIKV virions are spherical, enveloped particles approximately 65 nm in diameter.46 The T=4 icosahedral nucleocapsid encloses the positive-sense RNA genome and is wrapped by a host-derived lipid bilayer, within which 240 E1–E2 heterodimers assemble into 80 trimeric spikes that arrange an outer quasi-icosahedral glycoprotein lattice.47,48
Matrix remodeling–associated protein 8 (MXRA8), also known as DICAM, ASP3, or limitrin, is an adhesion molecule conserved across mammals, birds, and amphibians, with expression on epithelial, myeloid, and mesenchymal cells.49 Zhang et al identified MXRA8 as a host entry factor for several arthritogenic alphaviruses, including CHIKV, RRV, MAYV, and ONNV.50 Structural and functional studies show that MXRA8 binds CHIKV within the canyon between two adjacent protomers of the trimeric E spike, involving multiple interactions with E2–E1 heterodimers and the MXRA8 hinge region.45 Human MXRA8 facilitates CHIKV attachment to and infection of dermal fibroblasts, synovial fibroblasts, osteoblasts, chondrocytes, and skeletal muscle cells, and serves as a key receptor for viral entry into host cells.50 Consistently, an MXRA8-Fc fusion protein or anti-MXRA8 monoclonal antibodies effectively block CHIKV infection in vitro and reduce viral infection and joint swelling in mice.38 Live cell imaging indicates that MXRA8 binds CHIKV at the cell surface and co-internalizes with viral particles, many of which remain colocalized with MXRA8 during endosomal membrane fusion, further supporting its role as a bona fide entry receptor.51 Kim et al demonstrated that MXRA8 from mouse, rat, chimpanzee, dog, horse, goat, sheep, and human supports CHIKV infection in cell culture, whereas bovine MXRA8 does not because a 15-amino acid insertion in its ectodomain prevents viral binding. Deletion of this insertion in bovine MXRA8 enhances CHIKV binding and infection, while introducing the insertion into mouse MXRA8 blocks binding in cells and mitigates CHIKV-induced pathogenesis in mice. In contrast, avian MXRA8 (chicken, turkey, and duck) does not support CHIKV infection.52 However, MXRA8 is not expressed on all target cell types, and CHIKV infection of MXRA8 knockout mice leads to a reduced but not abolished level of replication, suggesting that the virus likely engages additional receptors in vivo.53,54 Other studies have identified the four-and-a-half-LIM domain protein 1 splice variant A (FHL1A) as a crucial host factor that interacts with the hypervariable domain of CHIKV nsP3 and is essential for viral RNA replication, with high expression in CHIKV target cells and marked abundance in muscle.55 In humans, FHL1 levels are elevated in both acute and chronic chikungunya disease, and FHL1 knockout mice infected with CHIKV exhibit reduced arthritis and myositis, fewer immune infiltrates, and decreased production of proinflammatory cytokines and chemokines compared with infected wild type controls.56 In addition, prohibitin, dendritic cell-specific intercellular adhesion molecule 3-grabbing non-integrin (DC-SIGN), basigin (CD147), and T cell immunoglobulin and mucin domain-containing protein 1 (TIM1) have also been proposed as entry or attachment factors for CHIKV.57–59
Animal Models and Human Pathogenesis
Mouse models of CHIKV infection provide useful tools for investigating viral and host factors that drive CHIKV pathogenesis.60,61 In mice with mild CHIKV infection, the virus primarily targets fibroblasts in muscle, joints, and skin, while in severe disease, it disseminates to the central nervous system and specifically targets the choroid plexus and leptomeninges.62 Several studies have reported that CHIKV-infected mice develop active synovitis with articular cartilage erosion, periostitis with cortical bone involvement, and myositis characterized by muscle fiber degeneration.63,64 Legros et al using an in vivo mouse model encompassing acute and chronic CHIKV infection, confirmed that the metatarsal joints supported viral replication after the acute phase and that chondrocytes were the main targets.65
In macaque models, CHIKV targets lymphoid tissues, liver, central nervous system, joints, and muscles, mainly infecting macrophages, dendritic cells, and endothelial cells during the acute phase. In the later stages, the virus persists in lymphoid organs, liver, joints, and muscles, and can be detected in macrophages for up to three months after inoculation.66 Macrophages have been identified as the primary cellular reservoir of persistent CHIKV infection during the chronic phase. Studies in rhesus macaques have revealed age-related differences, with older animals showing a general decline in both innate and adaptive immune responses and infectious CHIKV persisting in the spleen, liver, and muscle for up to 44 days postinfection.67 In cynomolgus macaques, higher inoculation doses correlate with earlier viremia peaks and higher peak viremia levels.68
CHIKV exhibits broad cellular tropism and infects various human cell lines and primary cells in vitro, including epithelial (HeLa, HEK293, Huh7), hepatocytic (HepG2), microglial (CHME5), and neuroblastoma lines, as well as primary fibroblasts (MRC5), skeletal muscle myoblasts (HSMMs), and monocyte-derived macrophages.69 These in vitro findings are supported by animal studies showing that CHIKV replicates in fibroblasts, muscle, joint, and immune cells, and disseminates to multiple organs under severe conditions. In natural infection, when an infected mosquito probes the host skin, saliva together with virus is injected, carrying a cocktail of molecules with anti-hemostatic and immunomodulatory properties.70 At the cutaneous inoculation site, CHIKV initially replicates in epidermal keratinocytes, melanocytes, and Langerhans cells, following which it spreads to dermal stromal cells such as fibroblasts and endothelial cells. Infected Langerhans cells and dermal dendritic cells subsequently undergo maturation upon antigenic stimulation and migrate to draining lymph nodes, where they initiate adaptive immune responses.71–73 By interacting with antigen-presenting cells in the skin, CHIKV enters the lymphatics, reaches the bloodstream and induces viremia.74 In this process, infiltrating monocytes support viral replication at the inoculation site and promote systemic dissemination.75 CHIKV subsequently disseminates hematogenously to organs including the liver, kidneys, spleen, lymph nodes, muscle, and joints.76 Type I interferons (IFN) induced during infection activate resident macrophages toward a classical phenotype, enhancing microbicidal activity and the production of proinflammatory mediators.77 Activated dendritic cells also secrete cytokines and chemokines.78 Human fibroblast-like synoviocytes have been shown to be permissive to CHIKV replication, creating a synovial environment that favors osteoclastogenesis, the resulting osteoclasts may contribute to CHIKV-associated arthralgia and arthritis.79 Quadriceps muscle biopsies from infected patients revealed scattered myofiber atrophy and necrosis with minimal inflammatory infiltration at the end of the acute phase. Around three months after infection, biopsies demonstrated extensive interstitial inflammation with polymorphonuclear cells, lymphocytes, and histiocytes, along with large areas of necrosis and collagen deposition.80 An autopsy study of fatal chikungunya cases demonstrated that CHIKV antigen exhibited marked tropism for mesenchymal cells (fibroblasts, adipocytes, and endothelial cells) and mononuclear cells, including tissue macrophages, blood monocytes, splenic follicular dendritic cells, and Kupffer cells, although no overt cytopathic effects were observed.81
Clinical Manifestations and Immune Responses
Chikungunya symptoms usually develop within 4 to 8 days (range 2–12 days) after the bite of an infected mosquito and most cases are self-limiting.82 Approximately 15–35% of infected individuals remain asymptomatic.83 Chikungunya typically presents with fever, often accompanied by joint pain, while other common manifestations include myalgia, rash, headache, nausea, and fatigue (Figure 1). Joint pain usually resolves within days but may persist for weeks, months, or even years in some patients.84 Clinically, chikungunya progresses through acute, post-acute, and chronic phases, defined by the duration of joint symptoms ranging from transient arthralgia to chronic arthritis.85
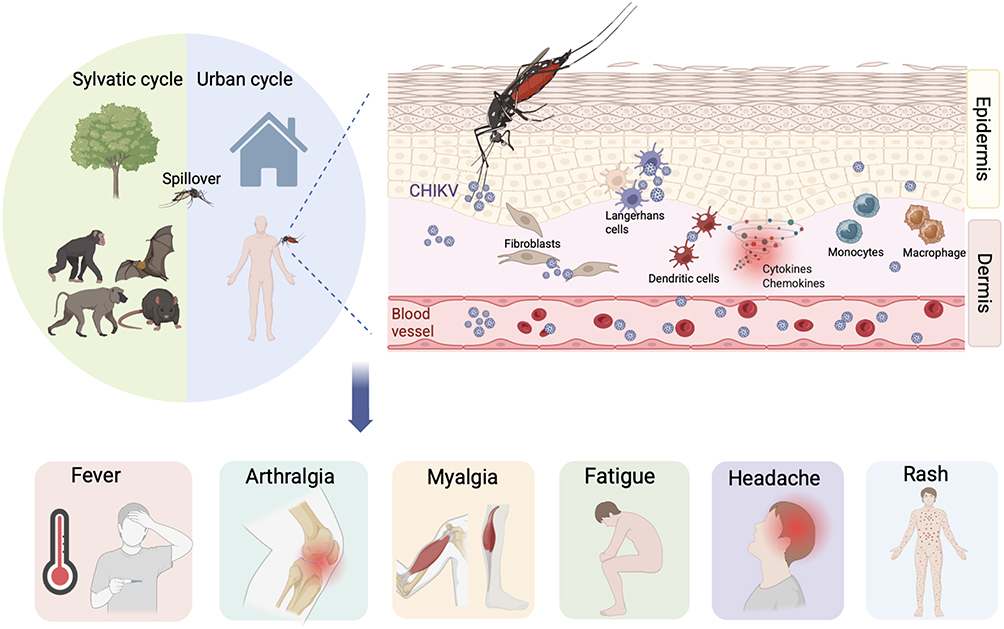 |
Figure 1 Chikungunya virus transmission, host infection, and clinical manifestations. |
The acute phase lasts up to two weeks and is characterized primarily by abrupt onset fever, severe arthralgia, joint swelling, and rash.46,69 Fever usually lasts for 4–5 days.86 Joint symptoms are usually symmetrical, predominantly affecting distal joints such as the wrists, ankles, and small joints of the hands, while the spine and sacroiliac joints are rarely affected.87 Older age has been associated with a higher risk of acute arthralgia.88 Skin manifestations often present as a transient macular or maculopapular rash involving the extremities, palms, feet, trunk, and face, usually appearing within 2–5 days of illness onset, pruritus occurs in approximately one quarter of cases.89 A recent meta-analysis of symptomatic adults found that arthralgia was the most common manifestation, affecting 89.7% of patients, followed by fever in 87.8%, myalgia in 62.9%, fatigue in 56.0%, joint swelling in 50.0%, headache in 49.5%, rash in 44.3%, and nausea in 34.7%, whereas arthritis was reported less frequently at 17.6%.90 Other atypical features include lymphadenopathy, conjunctivitis, optic neuritis, photosensitivity, exfoliative dermatitis, hyperpigmentation, vasculitic eruptions, erythema nodosum–like lesions, exacerbation of pre-existing dermatoses, encephalitis, meningitis, and mucosal ulceration.89,91–93 Chikungunya-associated neurological disease is regarded as the most common severe complication of infection.94 Autopsy findings from fatal cases confirmed the anatomical sites underlying these clinical manifestations, with CHIKV antigen detected in multiple tissues, including the skin, bone, muscle, spleen, liver, kidney, lung, and heart.81 CHIKV infection is highly symptomatic and marked by high-titer viremia that peaks within the first three days and can persist for up to eight days.95 Acute infection elicits robust activation of innate immune responses.96 The acute phase inflammatory response is characterized by elevated immune mediators and infiltration of immune cells into affected tissues, dominated by interleukin (IL)-6, IL-8, IFN-α, interferon gamma–induced protein (IP)-10, monocyte chemoattractant protein (MCP)-1, and monokine induced by gamma interferon (MIG).97–99 Higher circulating levels of IL-1β, IL-17A, IL-27, and granulocyte–macrophage colony-stimulating factor (GM-CSF) correlate with more severe joint pain.100 Consistent with other reports, IL-27, a member of the IL-6/IL-12 family, is associated with rheumatoid arthritis disease activity and correlates with tender joint counts in CHIKV infection.101 In mouse models, replication in skeletal muscle cells and IL-6 release are key mediators of disease, and IL-6 receptor blockade significantly reduces joint swelling.102 Correspondingly, high IL-6 levels have been most consistently associated with disease severity.103 Type-I IFN signaling is critical for controlling infection, and its deficiency is associated with severe disease.62 MCP-1 is produced primarily by monocytes, and its induction requires IFN-β–mediated communication with other leukocytes.104 IL-6 is one of the IFN-β–responsive cytokine produced by non-monocytic cells that further promotes this response.105 Despite early correlations between MCP-1 levels and viral titers, MCP-1 does not directly contribute to CHIKV replication.98,105 Overall, the balance among pro- and anti-inflammatory mediators is critical for an appropriate host response to CHIKV and is a key determinant of disease severity. Moreover, galectin-9 (GAL-9) and high mobility group box 1 protein (HMGB1) have been identified as potential biomarkers of acute infection.106,107 Several studies have indicated that CD8⁺ T cells are recruited early during CHIKV infection, where they play an important role in adaptive immunity by mediating cytolytic activity against target cells through granules exocytosis and granule-independent pathways.108,109 These cytotoxic functions contribute to viral clearance in the circulation.108
The post-acute phase spans from two weeks to three months after onset, during which joint manifestations may either persist continuously or recur intermittently following symptom-free periods.110 Most infected individuals recoverfully after the acute phase, but 30–40% progress to chronic arthritis that may last for months or even years.111 The chronic phase is defined by persistent of joint manifestations such as pain, edema, or stiffness for more than three months after the acute phase. During this stage, patients may also develop chronic arthritis attributable to chikungunya.85,112 Recent data suggest that arthritis can persist for up to seven years in some patients.113 Chronic CHIKV disease is characterized by ongoing inflammation and immune activation.114 Previous study reported that, in a patient 18 months postinfection, CHIKV RNA and antigens were detected in perivascular synovial macrophages surrounded by infiltrating NK cells, CD4+ T cells, and fewer cytotoxic CD8+ T cells.115 CHIKV antigen has also been detected in human muscle specimen more than three months after acute infection.80 However, synovial fluid analysis from 38 participants with CHIKV-associated arthritis at a median of 22 months postinfection was qRT-PCR negative, showed no viral proteins by mass spectrometry, and was culture negative.116 A meta-analysis shows that chronicity rates decline over time, from 43.9% at three months to 34.4% at six months and 31.9% at twelve months.[87] Similarly, a Latin American cohort study reported that approximately 25% of participants experienced persistent joint pain after 20 months of follow-up, highlighting the long-term burden of chikungunya-related arthritis.117 Higher viral loads and acute symptoms such as arthralgia, myalgia, and weakness are associated with an increased risk of developing chronic arthralgia.46 Elevated levels of IL-6 and GM-CSF have also been linked to persistent joint pain.99 In addition, elevated acute-phase levels of IL-6, IL-1β, TNF-α, IL-5, or IL-12 are associated with a greater risk of chronic joint pain, whereas low levels of IL-13, IL-2, and IL-4 are predictive of chronic outcomes, and a vigorous cytokine response appears necessary for effective viral clearance.103,118
Although severe disease and mortality are rare in chikungunya, neurological, cardiac, and renal involvement may result in high morbidity and mortality.119 Older adults (≥65 years), infants, young children, pregnant women, and individuals with comorbidities such as hypertension, diabetes, or cardiovascular disease are at increased risk of severe disease and prolonged recovery.82 After infection, IgM antibodies appear within days of symptom onset, with neutralizing anti-CHIKV IgG antibodies typically emerging by the second week, and the IgG3 isotype predominates in the humoral response.120 Current evidence suggests that recovery is followed by durable immunity against reinfection.121
Accurate laboratory diagnosis of CHIKV infection is essential for clinical management and epidemiological surveillance. Diagnostic testing is typically performed on plasma or serum samples from suspected cases. During the acute phase, viral RNA can be detected by RT-PCR, which offers high sensitivity and specificity and is considered the reference method for early diagnosis. Viral isolation using cell culture is largely restricted to research and reference laboratories.69 In later stages of infection, serological assays detecting CHIKV-specific IgM and IgG antibodies are widely used, including enzyme-linked immunosorbent assays (ELISA) and emerging point-of-care tests that facilitate rapid diagnosis in resource-limited settings.122 However, serological testing may be complicated by cross-reactivity among antigenically related arboviruses, potentially reducing diagnostic specificity in regions with overlapping transmission. Recent studies have demonstrated the feasibility and performance of affordable multiplex qRT-PCR assays for the simultaneous detection of CHIKV, DENV, ZIKV, YFV, and co-infections, as well as broader panels capable of identifying multiple medically important flaviviruses and alphaviruses in parallel.123–125 These multiplex platforms improve diagnostic efficiency and support outbreak response in endemic regions.
Mother-to-Child CHIKV Transmission
CHIKV infection does not increase the risk of miscarriage, intrauterine fetal death, or congenital malformations. However, maternal–neonatal transmission can occur in viremic women during childbirth.126 A meta-analysis estimated the overall risk of mother-to-child transmission at 15.5% (206/1331). The pooled risks of neonatal death were 0.6% (5/832) among maternal infections and 2.8% (5/182) among neonatal infections, while the risk of CHIKV-confirmed antepartum fetal death was 0.3% (3/1203). Symptomatic infection occurred in 50.0% (23/46) of intrapartum cases but in none of the 712 antepartum or peripartum maternal infections. Infected neonates were either asymptomatic or developed symptoms during the first week of life, rather than at birth, including fever, irritability, rash, hyperalgesia, diffuse limb edema, bullous dermatitis, and occasionally meningoencephalitis. Long-term neurodevelopmental delay was observed in 50% of symptomatic neonatal infections.127 In a two-year follow-up study from the Réunion Island outbreak, children exposed to perinatal mother-to-child CHIKV infection had significantly lower neurocognitive scores than unexposed peers (development quotient: 86.3 vs 100.2, p < 0.001). Among exposed children, those with severe encephalopathy had significantly worse outcomes than those with non-severe disease (mean development quotient: 77.6 vs 91.2, p < 0.001). Of 12 infants with neonatal encephalopathy, five developed microcephaly and four matched the criteria for cerebral palsy.128
Gérardin et al conducted a prospective study showing that mother-to-child CHIKV transmission occurs almost exclusively during intrapartum maternal viremia and is frequently associated with severe neonatal disease. Among 39 viremic mothers at delivery, 19 neonates became infected (vertical transmission rate, 48.7%), with severe illness observed in 10 cases, predominantly neonatal encephalopathy. Notably, placentas from infected neonates had significantly higher mean viral loads than those from uninfected neonates (p = 0.021).129 Another study reported similar findings, with a transmission rate of approximately 62% (18/29) among viremic pregnant women during delivery.130 Immunofluorescence labeling with anti-CHIKV antibodies failed to detect viral antigen in placentas or in an animal model of maternal–fetal CHIKV infection.62 In addition, RT-PCR on placental cells isolated by mechanical dissociation was consistently negative.129 These findings support passive contamination of the placenta by maternal blood-borne free viral particles rather than true placental infection. Mother-to-child transmission of CHIKV may result from placental barrier disruption caused by uterine contractions, allowing maternal blood with high viral loads to cross into the fetal circulation. Notably, emergency cesarean section delivery does not prevent transmission.129 Consistently, Leglet et al reported that among 151 women infected with CHIKV during pregnancy, 118 delivered outside the viremic period and no neonate showed evidence of infection. In contrast, among 33 women who delivered while viremic, 16 newborns (48.5%) developed neonatal chikungunya.131 Regarding management, Escobar et al suggested that deferring delivery in women with acute CHIKV infection may be an appropriate management strategy in the absence of obstetric contraindications. The mean interval from maternal symptom onset to delivery was 6.3 ± 1.4 days, which may provide sufficient time for passive transfer of maternal antibodies and thereby reduce the risk of symptomatic infection in the newborn.132
Vaccine Development
In November 2023, the US Food and Drug Administration (FDA) approved IXCHIQ (VLA1553), a live-attenuated vaccine developed by Valneva, as the first licensed vaccine against chikungunya (Table 1). IXCHIQ was approved for individuals ≥ 18 years at increased risk of CHIKV exposure and was administered as a single intramuscular dose. Its prescribing information included a warning that the vaccine contains live attenuated virus and may cause symptoms in recipients that are similar to those of natural chikungunya infection.133 In June 2024, the vaccine was unanimously endorsed by the Member States following a stringent assessment by the European Medicines Agency (EMA).134 Owing to the unpredictable nature of CHIKV epidemiology, vaccine licensure was based on a correlate of protection rather than traditional Phase 3 trials.135 On May 9, 2025, the FDA and the Centers for Disease Control and Prevention (CDC) issued a safety communication recommending suspension of IXCHIQ use in adults ≥60 years while post-marketing reports of serious neurologic and cardiac adverse events were under investigation. The suspension was lifted on August 6, 2025, following a risk-benefit assessment. Subsequently, on August 22, 2025, the FDA’s Center for Biologics Evaluation and Research (CBER) suspended the biologics license for IXCHIQ. CBER’s benefit–risk analysis concluded that the clinical benefit had not been adequately confirmed in clinical studies and the potential benefits of IXCHIQ did not outweigh the associated risks. Accordingly, CBER determined that the vaccine was not safe and that continued administration would pose a public health risk.136
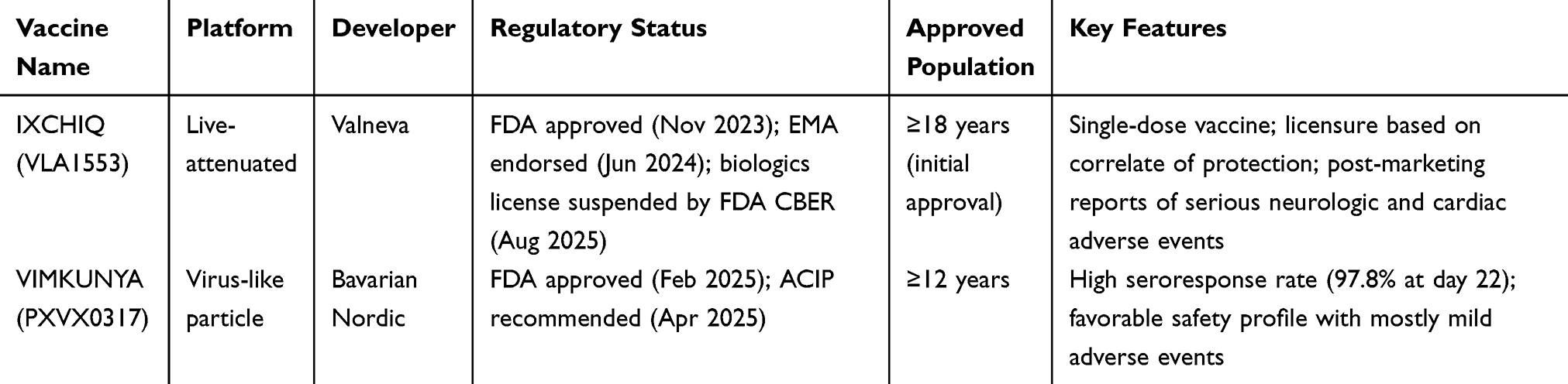 |
Table 1 Current Landscape of Licensed Chikungunya Vaccines (Updated to August 2025) |
In February 2025, the FDA approved VIMKUNYA (PXVX0317) for individuals ≥ 12 years, a virus-like particle vaccine developed by Bavarian Nordic. Phase 3 clinical trial results demonstrated a seroresponse rate of 97.8% with significantly elevated serum neutralizing antibody titers at day 22. In addition, VIMKUNYA also showed a favorable safety profile, with most adverse events reported as mild and self-limiting.137 In April 2025, the Advisory Committee on Immunization Practices (ACIP) issued recommendations for the use of VIMKUNYA in travelers to chikungunya endemic countries or territories and in laboratory workers at risk of exposure.138
Future Recommendations
The geographic suitability for CHIKV transmission continues to expand. Long-term studies of international travelers show that chikungunya cases in returning travelers reflect transmission activity in endemic regions and often provide early evidence of virus circulation beyond affected areas.139,140 Retrospective epidemiological analyses in mainland Europe from 2007 to 2023 indicate that the majority of human CHIKV infections were travel-related and that imported cases were frequently observed either before or closely associated with autochthonous transmission in areas where competent Aedes vectors are established.141 Travelers returning to non-endemic regions with CHIKV infection may initiate local clusters or outbreaks where competent mosquito vectors are present. Accordingly, individuals travelling to areas with active CHIKV circulation should be informed of infection risk and advised on appropriate preventive measures.142 An exploratory surveillance study conducted in Portugal demonstrated that continuous and active surveillance is essential for the timely detection and management of potential CHIKV outbreaks.143 These observations support the incorporation of travel-associated case detection into integrated surveillance frameworks, together with entomological monitoring and laboratory confirmation, to facilitate early risk assessment and timely public health response.
Conclusions
In conclusion, chikungunya remains a significant global health threat, with Aedes mosquitoes well adapted to urban environments and expanding their range due to climate change and human activity. Future studies should clarify how early innate and adaptive immune responses lead to chronic joint disease and develop validated biomarkers that predict persistence and severity. Differences in these responses between asymptomatic and symptomatic CHIKV infection, and between self-limited and chronic courses, need clearer definition. Mother-to-child care should be optimized to reduce transmission at the time of delivery. Developing effective therapeutic approaches is essential to shorten the disease course and reduce the incidence of chronic arthritis. Vaccine strategies should provide safe, durable, and broadly accessible protection for high-risk populations, with continuous monitoring of effectiveness and safety in real-world settings. Progress against chikungunya will require multidisciplinary efforts that integrate fundamental and translational research, clinical investigation, epidemiology, and public health practice.
Abbreviations
CHIKV, chikungunya virus; PAHO, Pan American Health Organization; WHO, World Health Organization; RRV, Ross River virus; BFV, Barmah Forest virus; SINV, Sindbis virus; MAYV, Mayaro virus; ONNV, O’nyong-nyong virus; EEEV, Eastern equine encephalitis virus; VEEV, Venezuelan equine encephalitis virus; WEEV, Western equine encephalitis virus; DENV, dengue virus; ZIKV, Zika virus; YFV, yellow fever viruses; dpi, days post infection; ORF, open reading frame; C, capsid; E, envelope; ECSA, East/Central/South African; IOL, Indian Ocean lineage; MXRA8, Matrix remodeling–associated protein 8; FHL1A, four-and-a-half-LIM domain protein 1 splice variant A; DC-SIGN, dendritic cell-specific intercellular adhesion molecule 3-grabbing non-integrin; TIM1, T cell immunoglobulin and mucin domain-containing protein 1; IL, interleukin; IFN, interferon; IP, interferon gamma–induced protein; MCP, monocyte chemoattractant protein; MIG, monokine induced by gamma interferon; GM-CSF, granulocyte–macrophage colony-stimulating factor; GAL-9, galectin-9; HMGB1, high mobility group box 1 protein; ELISA, enzyme-linked immunosorbent assays; FDA, Food and Drug Administration; EMA, European Medicines Agency; CDC, Centers for Disease Control and Prevention; CBER, Center for Biologics Evaluation and Research; ACIP, Advisory Committee on Immunization Practices.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Key Research and Development Program of China (2022YFF1203201).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Robinson MC. An epidemic of virus disease in southern province, tanganyika territory, in 1952–53. I. Clinical features. Trans R Soc Trop Med Hyg. 1955;49(1):28–14. doi:10.1016/0035-9203(55)90080-8
2. Hammon WM, Rudnick A, Sather GE. Viruses associated with epidemic hemorrhagic fevers of the Philippines and Thailand. Science. 1960;131(3407):1102–1103. doi:10.1126/science.131.3407.1102
3. Renault P, Solet JL, Sissoko D, et al. A major epidemic of chikungunya virus infection on Reunion Island, France, 2005–2006. Am J Trop Med Hyg. 2007;77(4):727–731. [PMID: 17978079]. doi:10.4269/ajtmh.2007.77.727
4. Rezza G, Nicoletti L, Angelini R, et al. Infection with chikungunya virus in Italy: an outbreak in a temperate region. Lancet. 2007;370(9602):1840–1846. doi:10.1016/S0140-6736(07)61779-6
5. Olowokure B, Francis L, Polson-Edwards K, et al. The Caribbean response to chikungunya. Lancet Infect Dis. 2014;14(11):1039–1040. doi:10.1016/S1473-3099(14)70948-X
6. Epidemiological update for dengue and other arboviruses. Available from: https://ais.paho.org/ha_viz/Arbo/Arbo_Bulletin_2023.asp?env=pri.
7. Epidemiological update for dengue and other arboviruses. Available from: https://ais.paho.org/ha_viz/Arbo/Arbo_Bulletin_2024.asp?env=pri.
8. Chikungunya virus disease worldwide overview. 2025. Available from: https://www.ecdc.europa.eu/en/chikungunya-monthly.
9. Guangdong Provincial center for disease control and Prevention. Available from: https://cdcp.gd.gov.cn/ywdt/zdzt/yfjkkyr/yqxx/index.html.
10. Dai Z, Chen Y, Jiang Y, et al. Global Assessment of current and future chikungunya virus transmission risk using optimized maxent modeling. Acta Trop. 2025;269:107756. doi:10.1016/j.actatropica.2025.107756
11. Chikungunya epidemiology update - June 2025. Available from: https://www.who.int/publications/m/item/chikungunya-epidemiology-update-june-2025.
12. Current ICTV taxonomy release | ICTV. Available from: https://ictv.global/taxonomy.
13. Chen W, Foo SS, Sims NA, Herrero LJ, Walsh NC, Mahalingam S. Arthritogenic alphaviruses: new insights into arthritis and bone pathology. Trend Microbiol. 2015;23(1):35–43. doi:10.1016/j.tim.2014.09.005
14. Suhrbier A, Jaffar-Bandjee MC, Gasque P. Arthritogenic alphaviruses—an overview. Nat Rev Rheumatol. 2012;8(7):420–429. doi:10.1038/nrrheum.2012.64
15. Ronca SE, Dineley KT, Paessler S. Neurological sequelae resulting from encephalitic alphavirus infection. Front Microbiol. 2016;7:959. doi:10.3389/fmicb.2016.00959
16. Zaid A, Burt FJ, Liu X, et al. Arthritogenic alphaviruses: epidemiological and clinical perspective on emerging arboviruses. Lancet Infect Dis. 2021;21(5):e123–e133. doi:10.1016/S1473-3099(20)30491-6
17. Diallo D, Sall AA, Buenemann M, et al. Landscape ecology of sylvatic chikungunya virus and mosquito vectors in southeastern senegal. PLoS Negl Trop Dis. 2012;6(6):e1649. doi:10.1371/journal.pntd.0001649
18. Mongkol N, Wang FS, Suthisawat S, Likhit O, Charoen P, Boonnak K. Seroprevalence of Chikungunya and Zika virus in nonhuman primates: a systematic review and meta-analysis. One Health. 2022;15:100455. doi:10.1016/j.onehlt.2022.100455
19. Weaver SC, Chen R, Diallo M. Chikungunya virus: role of vectors in emergence from enzootic cycles. Annu Rev Entomol. 2020;65:313–332. doi:10.1146/annurev-ento-011019-025207
20. de Souza WM, Gaye A, Ndiaye EH, et al. Serosurvey of chikungunya virus in old world fruit bats, Senegal, 2020–2022. Emerg Infect Dis. 2024;30(7):1490–1492. doi:10.3201/eid3007.240055
21. Weaver SC, Lecuit M. Chikungunya virus and the global spread of a mosquito-borne disease. New Engl J Med. 2015;372:1231–9. doi:10.1056/NEJMra1406035
22. de Souza WM, Weaver SC. Effects of climate change and human activities on vector-borne diseases. Nat Rev Microbiol. 2024;22(8):476–491. doi:10.1038/s41579-024-01026-0
23. Hedrich N, Bekker-Nielsen Dunbar M, Grobusch MP, Schlagenhauf P. Aedes-borne arboviral human infections in Europe from 2000 to 2023: a systematic review and meta-analysis. Travel Med Infectious Dis. 2025;64:102799. doi:10.1016/j.tmaid.2025.102799
24. Lühken R, Brattig N, Becker N. Introduction of invasive mosquito species into Europe and prospects for arbovirus transmission and vector control in an era of globalization. Infect Dis Poverty. 2023;12(1):109. doi:10.1186/s40249-023-01167-z
25. Tuñon A, García J, Carrera LC, Chaves LF, Lenhart AE, Loaiza JR. Chemical control of medically important arthropods in Panama: a systematic literature review of historical efforts. Acta Trop. 2024;255:107217. doi:10.1016/j.actatropica.2024.107217
26. Dong S, Balaraman V, Kantor AM, et al. Chikungunya virus dissemination from the midgut of Aedes aegypti is associated with temporal basal lamina degradation during bloodmeal digestion. PLoS Negl Trop Dis. 2017;11(9):e0005976. doi:10.1371/journal.pntd.0005976
27. Dubrulle M, Mousson L, Moutailler S, Vazeille M, Failloux AB. Chikungunya virus and Aedes mosquitoes: saliva is infectious as soon as two days after oral infection. PLoS One. 2009;4(6):e5895. doi:10.1371/journal.pone.0005895
28. Vazeille M, Moutailler S, Coudrier D, et al. Two chikungunya isolates from the outbreak of La Reunion (Indian Ocean) exhibit different patterns of infection in the mosquito, Aedes albopictus. PLoS One. 2007;2(11):e1168. doi:10.1371/journal.pone.0001168
29. Chompoosri J, Thavara U, Tawatsin A, et al. Vertical transmission of Indian Ocean lineage of chikungunya virus in aedes aegypti and aedes albopictus mosquitoes. Parasit Vectors. 2016;9:227. doi:10.1186/s13071-016-1505-6
30. Heath CJ, Grossi-Soyster EN, Ndenga BA, et al. Evidence of transovarial transmission of chikungunya and dengue viruses in field-caught mosquitoes in Kenya. PLoS Negl Trop Dis. 2020;14(6):e0008362. doi:10.1371/journal.pntd.0008362
31. Mavale M, Parashar D, Sudeep A, et al. Venereal transmission of chikungunya virus by aedes aegypti mosquitoes (Diptera: culicidae). Am J Trop Med Hyg. 2010;83(6):1242–1244. doi:10.4269/ajtmh.2010.09-0577
32. de Souza WM, Ribeiro GS, de Lima STS, et al. Chikungunya: a decade of burden in the Americas. Lancet Reg Health Am. 2024;30:100673. doi:10.1016/j.lana.2023.100673
33. Tsetsarkin KA, Chen R, Weaver SC. Interspecies transmission and chikungunya virus emergence. Curr Opin Virol. 2016;16:143–150. doi:10.1016/j.coviro.2016.02.007
34. Tsetsarkin KA, Vanlandingham DL, McGee CE, Higgs S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007;3(12):e201. doi:10.1371/journal.ppat.0030201
35. Tsetsarkin KA, Chen R, Yun R, et al. Multi-peaked adaptive landscape for chikungunya virus evolution predicts continued fitness optimization in aedes albopictus mosquitoes. Nat Commun. 2014;5:4084. doi:10.1038/ncomms5084
36. Kraemer MUG, Reiner RC, Brady OJ, et al. Past and future spread of the arbovirus vectors aedes aegypti and aedes albopictus. Nat Microbiol. 2019;4(5):854–863. doi:10.1038/s41564-019-0376-y
37. de Souza WM, Lecuit M, Weaver SC. Chikungunya virus and other emerging arthritogenic alphaviruses. Nat Rev Microbiol. 2025:1–17. doi:10.1038/s41579-025-01177-8
38. Basore K, Kim AS, Nelson CA, et al. Cryo-EM structure of chikungunya virus in complex with the Mxra8 receptor. Cell. 2019;177(7):1725–1737.e16. doi:10.1016/j.cell.2019.04.006
39. Kril V, Hons M, Amadori C, et al. Alphavirus nsP3 organizes into tubular scaffolds essential for infection and the cytoplasmic granule architecture. Nat Commun. 2024;15(1):8106. doi:10.1038/s41467-024-51952-z
40. Voss JE, Vaney MC, Duquerroy S, et al. Glycoprotein organization of chikungunya virus particles revealed by X-ray crystallography. Nature. 2010;468(7324):709–712. doi:10.1038/nature09555
41. Wang M, Wang L, Leng P, Guo J, Zhou H. Drugs targeting structural and nonstructural proteins of the chikungunya virus: a review. Int J Biol Macromol. 2024;262(Pt 2):129949. doi:10.1016/j.ijbiomac.2024.129949
42. Martin CK, Yin P, Kielian M. The sticky business of alphavirus capsid-host interactions. Trends Microbiol. 2025;33(3):321–339. doi:10.1016/j.tim.2024.11.003
43. Li L, Jose J, Xiang Y, Kuhn RJ, Rossmann MG. Structural changes of envelope proteins during alphavirus fusion. Nature. 2010;468(7324):705–708. doi:10.1038/nature09546
44. Fox JM, Long F, Edeling MA, et al. Broadly neutralizing alphavirus antibodies bind an epitope on E2 and inhibit entry and egress. Cell. 2015;163(5):1095–1107. doi:10.1016/j.cell.2015.10.050
45. Song H, Zhao Z, Chai Y, et al. Molecular basis of arthritogenic alphavirus receptor MXRA8 binding to chikungunya virus envelope protein. Cell. 2019;177(7):1714–1724.e12. doi:10.1016/j.cell.2019.04.008
46. Bartholomeeusen K, Daniel M, LaBeaud DA, et al. Chikungunya fever. Nat Rev Dis Prim. 2023;9(1):17. doi:10.1038/s41572-023-00429-2
47. Chmielewski D, Schmid MF, Simmons G, Jin J, Chiu W. Chikungunya virus assembly and budding visualized in situ using cryogenic electron tomography. Nat Microbiol. 2022;7(8):1270–1279. doi:10.1038/s41564-022-01164-2
48. Button JM, Qazi SA, Wang JCY, Mukhopadhyay S. Revisiting an old friend: new findings in alphavirus structure and assembly. Curr Opin Virol. 2020;45:25–33. doi:10.1016/j.coviro.2020.06.005
49. Jung YK, Jin JS, Jeong JH, Kim HN, Park NR, Choi JY. DICAM, a novel dual immunoglobulin domain containing cell adhesion molecule interacts with alphavbeta3 integrin. J Cell Physiol. 2008;216(3):603–614. doi:10.1002/jcp.21438
50. Zhang R, Kim AS, Fox JM, et al. Mxra8 is a receptor for multiple arthritogenic alphaviruses. Nature. 2018;557(7706):570–574. doi:10.1038/s41586-018-0121-3
51. Feng F, Bouma EM, Hu G, et al. Colocalization of chikungunya virus with its receptor MXRA8 during cell attachment, internalization, and membrane fusion. J Virol. 2023;97(5):e0155722. doi:10.1128/jvi.01557-22
52. Kim AS, Zimmerman O, Fox JM, et al. An evolutionary insertion in the Mxra8 receptor-binding site confers resistance to alphavirus infection and pathogenesis. Cell Host Microbe. 2020;27(3):428–440.e9. doi:10.1016/j.chom.2020.01.008
53. Zhang R, Earnest JT, Kim AS, et al. Expression of the Mxra8 receptor promotes alphavirus infection and pathogenesis in mice and drosophila. Cell Rep. 2019;28(10):2647–2658.e5. doi:10.1016/j.celrep.2019.07.105
54. Single cell type - MXRA8 - the human protein atlas. Available from: https://www.proteinatlas.org/ENSG00000162576-MXRA8/single+cell#single_cell_type_summary.
55. Meertens L, Hafirassou ML, Couderc T, et al. FHL1 is a major host factor for chikungunya virus infection. Nature. 2019;574(7777):259–263. doi:10.1038/s41586-019-1578-4
56. Ng WH, Liu X, Ling ZL, et al. FHL1 promotes chikungunya and o’nyong-nyong virus infection and pathogenesis with implications for alphavirus vaccine design. Nat Commun. 2023;14(1):6605. doi:10.1038/s41467-023-42330-2
57. Wintachai P, Wikan N, Kuadkitkan A, et al. Identification of prohibitin as a chikungunya virus receptor protein. J Med Virol. 2012;84(11):1757–1770. doi:10.1002/jmv.23403
58. De Caluwé L, Ariën KK, Bartholomeeusen K. Host factors and pathways involved in the entry of mosquito-borne alphaviruses. Trends Microbiol. 2021;29(7):634–647. doi:10.1016/j.tim.2020.10.011
59. Caluwé L D, Coppens S, Vereecken K, et al. The CD147 protein complex is involved in entry of chikungunya virus and related alphaviruses in human cells. Front Microbiol. 2021;12:615165. doi:10.3389/fmicb.2021.615165
60. Morrison TE, Oko L, Montgomery SA, et al. A mouse model of chikungunya virus-induced musculoskeletal inflammatory disease: evidence of arthritis, tenosynovitis, myositis, and persistence. Am J Pathol. 2011;178(1):32–40. doi:10.1016/j.ajpath.2010.11.018
61. Chan YH, Lum FM, Ng LFP. Limitations of current in vivo mouse models for the study of chikungunya virus pathogenesis. Med Sci. 2015;3(3):64–77. doi:10.3390/medsci3030064
62. C T, C F, S C, et al. A mouse model for Chikungunya: young age and inefficient type-I interferon signaling are risk factors for severe disease. PLoS Pathogens. 2008;4(2). doi:10.1371/journal.ppat.0040029
63. Chang AY, Tritsch SR, Porzucek AJ, et al. A mouse model for studying post-acute arthritis of chikungunya. Microorganisms. 2021;9(9):1998. doi:10.3390/microorganisms9091998
64. Goupil BA, McNulty MA, Martin MJ, McCracken MK, Christofferson RC, Mores CN. Novel lesions of bones and joints associated with chikungunya virus infection in two mouse models of disease: new insights into disease pathogenesis. PLoS One. 2016;11(5):e0155243. doi:10.1371/journal.pone.0155243
65. Legros V, Belarbi E, Jeannin P, et al. Use of recombinant chikungunya virus expressing nanoluciferase to identify chondrocytes as target cells in an immunocompetent mouse model. J Infect Dis. 2025;232(2):e223–e233. doi:10.1093/infdis/jiaf232
66. L K, L T, J C, et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J Clin Invest. 2010;120(3). doi:10.1172/JCI40104
67. Messaoudi I, Vomaske J, Totonchy T, et al. Chikungunya virus infection results in higher and persistent viral replication in aged rhesus macaques due to defects in anti-viral immunity. PLoS Negl Trop Dis. 2013;7(7):e2343. doi:10.1371/journal.pntd.0002343
68. Chen CI, Clark DC, Pesavento P, et al. Comparative pathogenesis of epidemic and enzootic chikungunya viruses in a pregnant rhesus macaque model. Am J Trop Med Hyg. 2010;83(6):1249–1258. doi:10.4269/ajtmh.2010.10-0290
69. Khongwichit S, Chansaenroj J, Chirathaworn C, Poovorawan Y. Chikungunya virus infection: molecular biology, clinical characteristics, and epidemiology in Asian countries. J Biomed Sci. 2021;28(1):84. doi:10.1186/s12929-021-00778-8
70. Agarwal A, Joshi G, Nagar DP, et al. Mosquito saliva induced cutaneous events augment Chikungunya virus replication and disease progression. Infect Genet Evol. 2016;40:126–135. doi:10.1016/j.meegid.2016.02.033
71. Broeckel R, Haese N, Messaoudi I, Streblow DN. Nonhuman primate models of chikungunya virus infection and disease (CHIKV NHP Model). Pathogens. 2015;4(3):662–681. doi:10.3390/pathogens4030662
72. Matusali G, Colavita F, Bordi L, et al. Tropism of the chikungunya virus. Viruses. 2019;11(2):175. doi:10.3390/v11020175
73. Fong SW, Kini RM, Ng LFP. Mosquito saliva reshapes alphavirus infection and immunopathogenesis. J Virol. 2018;92(12):e01004–17. doi:10.1128/JVI.01004-17
74. Briant L, Desprès P, Choumet V, Missé D. Role of skin immune cells on the host susceptibility to mosquito-borne viruses. Virology. 2014;464–465:26–32. doi:10.1016/j.virol.2014.06.023
75. Holmes AC, Lucas CJ, Brisse ME, et al. Ly6C+ monocytes in the skin promote systemic alphavirus dissemination. Cell Rep. 2024;43(3):113876. doi:10.1016/j.celrep.2024.113876
76. Schwartz O, Albert ML. Biology and pathogenesis of chikungunya virus. Nat Rev Microbiol. 2010;8(7):491–500. doi:10.1038/nrmicro2368
77. Martinez FO, Gordon S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep. 2014;6:13. doi:10.12703/P6-13
78. Brulefert A, Kraemer M, Cumin M, et al. Chikungunya virus envelope protein E2 provides a vector for targeted antigen delivery to human dermal CD14+ dendritic cells. J Invest Dermatol. 2021;141(12):2985–2989.e5. doi:10.1016/j.jid.2021.04.027
79. Phuklia W, Kasisith J, Modhiran N, et al. Osteoclastogenesis induced by CHIKV-infected fibroblast-like synoviocytes: a possible interplay between synoviocytes and monocytes/macrophages in CHIKV-induced arthralgia/arthritis. Virus Res. 2013;177(2):179–188. doi:10.1016/j.virusres.2013.08.011
80. Ozden S, Huerre M, Riviere JP, et al. Human muscle satellite cells as targets of chikungunya virus infection. PLoS One. 2007;2(6):e527. doi:10.1371/journal.pone.0000527
81. Sharp TM, Keating MK, Shieh WJ, et al. Clinical characteristics, histopathology, and tissue immunolocalization of chikungunya virus antigen in fatal cases. Clin Infect Dis. 2021;73(2):e345–e354. doi:10.1093/cid/ciaa837
82. Chikungunya. Available from: https://www.who.int/news-room/questions-and-answers/item/chikungunya.
83. CDC. Chikungunya. Yellow book. 2025. Available from: https://www.cdc.gov/yellow-book/hcp/travel-associated-infections-diseases/chikungunya.html.
84. Chikungunya. Available from: https://www.who.int/health-topics/chikungunya.
85. Case definitions, clinical classification, and disease phases. Available from: https://www.medbox.org/document/case-definitions-clinical-classification-and-disease-phases.
86. Paul BJ, Sadanand S. Chikungunya Infection: a Re-emerging Epidemic. Rheumatol Ther. 2018;5(2):317–326. doi:10.1007/s40744-018-0121-7
87. Pathak H, Mohan MC, Ravindran V. Chikungunya arthritis. Clin Med. 2019;19(5):381–385. doi:10.7861/clinmed.2019-0035
88. Warnes CM, Bustos Carrillo FA, Zambrana JV, et al. Longitudinal analysis of post-acute chikungunya-associated arthralgia in children and adults: a prospective cohort study in Managua, Nicaragua (2014–2018). PLoS Negl Trop Dis. 2024;18(2):e0011948. doi:10.1371/journal.pntd.0011948
89. Thiberville SD, Moyen N, Dupuis-Maguiraga L, et al. Chikungunya fever: epidemiology, clinical syndrome, pathogenesis and therapy. Antiviral Res. 2013;99(3):345–370. doi:10.1016/j.antiviral.2013.06.009
90. Rama K, de Roo AM, Louwsma T, et al. Clinical outcomes of chikungunya: a systematic literature review and meta-analysis. PLoS Negl Trop Dis. 2024;18(6):e0012254. doi:10.1371/journal.pntd.0012254
91. Sonego B, Schettini A, Talhari S, Maciel L, Massone C. Dermatological manifestations during dengue, chikungunya, and zika infections. Curr Opin Infect Dis. 2025;38(2):92. doi:10.1097/QCO.0000000000001077
92. Porto Silva CN, Crispim JG, Pereira MC, Galdino da Rocha Pitta M, Barreto de Melo Rêgo MJ, Melgarejo da Rosa M. The communication between chikungunya infection and the central nervous system. Microb Pathog. 2025;206:107747. doi:10.1016/j.micpath.2025.107747
93. da Silva LCM, da Silva Platner F, da Silva Fonseca L, et al. Ocular manifestations of chikungunya infection: a systematic review. Pathogens. 2022;11(4):412. doi:10.3390/pathogens11040412
94. Mehta R, Gerardin P, de Brito CAA, Soares CN, Ferreira MLB, Solomon T. The neurological complications of chikungunya virus: a systematic review. Rev Med Virol. 2018;28(3):e1978. doi:10.1002/rmv.1978
95. Appassakij H, Khuntikij P, Kemapunmanus M, Wutthanarungsan R, Silpapojakul K. Viremic profiles in asymptomatic and symptomatic chikungunya fever: a blood transfusion threat? Transfusion. 2013;53(10 Pt 2):2567–2574. doi:10.1111/j.1537-2995.2012.03960.x
96. Rueda JC, Arcos-Burgos M, Santos AM, et al. Human genetic host factors and its role in the pathogenesis of chikungunya virus infection. Front Med. 2022;9:654395. doi:10.3389/fmed.2022.654395
97. Burt FJ, Chen W, Miner JJ, et al. Chikungunya virus: an update on the biology and pathogenesis of this emerging pathogen. Lancet Infect Dis. 2017;17(4):e107–e117. doi:10.1016/S1473-3099(16)30385-1
98. Teng TS, Kam YW, Lee B, et al. A systematic meta-analysis of immune signatures in patients with acute chikungunya virus infection. J Infect Dis. 2015;211(12):1925–1935. doi:10.1093/infdis/jiv049
99. Chow A, Her Z, Ong EKS, et al. Persistent arthralgia induced by chikungunya virus infection is associated with interleukin-6 and granulocyte macrophage colony-stimulating factor. J Infect Dis. 2011;203(2):149–157. doi:10.1093/infdis/jiq042
100. Gualberto Cavalcanti N, MeloVilar K, Branco Pinto Duarte AL, et al. IL-27 in patients with chikungunya fever: a possible chronicity biomarker? Acta Trop. 2019;196:48–51. doi:10.1016/j.actatropica.2019.05.005
101. Lai X, Wang H, Cao J, et al. Circulating IL-27 is elevated in rheumatoid arthritis patients. Molecules. 2016;21(11):1565. doi:10.3390/molecules21111565
102. Lentscher AJ, McCarthy MK, May NA, et al. Chikungunya virus replication in skeletal muscle cells is required for disease development. J Clin Invest. 2020;130(3):1466–1478. doi:10.1172/JCI129893
103. Chirathaworn C, Chansaenroj J, Poovorawan Y. Cytokines and chemokines in chikungunya virus infection: protection or induction of pathology. Pathogens. 2020;9(6):415. doi:10.3390/pathogens9060415
104. Pattison MJ, MacKenzie KF, Elcombe SE, Arthur JSC. IFNβ autocrine feedback is required to sustain TLR induced production of MCP-1 in macrophages. FEBS Lett. 2013;587(10):1496–1503. doi:10.1016/j.febslet.2013.03.025
105. Ruiz Silva M, van der Ende-Metselaar H, Mulder HL, Smit JM, Rodenhuis-Zybert IA. Mechanism and role of MCP-1 upregulation upon chikungunya virus infection in human peripheral blood mononuclear cells. Sci Rep. 2016;6:32288. doi:10.1038/srep32288
106. Gualberto Cavalcanti N, Melo Vilar K, Branco Pinto Duarte AL, et al. Increased serum levels of galectin-9 in patients with chikungunya fever. Virus Res. 2020;286:198062. doi:10.1016/j.virusres.2020.198062
107. Rocha DCP, Souza TMA, Nunes PCG, et al. Increased circulating levels of High Mobility Group Box 1 (HMGB1) in acute-phase chikungunya virus infection: potential disease biomarker. J Clin Virol. 2022;146:105054. doi:10.1016/j.jcv.2021.105054
108. Dias de CNS, Gois BM, Lima VS, et al. Human CD8 T-cell activation in acute and chronic chikungunya infection. Immunology. 2018;155(4):499–504. doi:10.1111/imm.12992
109. Wauquier N, Becquart P, Nkoghe D, Padilla C, Ndjoyi-Mbiguino A, Leroy EM. The acute phase of Chikungunya virus infection in humans is associated with strong innate immunity and T CD8 cell activation. J Infect Dis. 2011;204(1):115–123. doi:10.1093/infdis/jiq006
110. Zaid A, Gérardin P, Taylor A, Mostafavi H, Malvy D, Mahalingam S. Review: chikungunya arthritis: implications of acute and chronic inflammation mechanisms on disease management. Arthritis Rheumatol. 2018;70(4):484–495. doi:10.1002/art.40403
111. Chikungunya virus disease. 2017. Available from: https://www.ecdc.europa.eu/en/chikungunya-virus-disease.
112. Amaral JK, Bilsborrow JB, Schoen RT. Chronic chikungunya arthritis and rheumatoid arthritis: what they have in common. Am J Med. 2020;133(3):e91–e97. doi:10.1016/j.amjmed.2019.10.005
113. Chang AYH, Hernández AS, Mejía JF, et al. The natural history of post-chikungunya viral arthritis disease activity and T-cell immunology: a cohort study. J Cell Immunol. 2024;6(2):64–75. doi:10.33696/immunology.6.191
114. Young AR, Locke MC, Cook LE, et al. Dermal and muscle fibroblasts and skeletal myofibers survive chikungunya virus infection and harbor persistent RNA. PLoS Pathog. 2019;15(8):e1007993. doi:10.1371/journal.ppat.1007993
115. Hoarau JJ, Jaffar Bandjee MC, Krejbich Trotot P, et al. Persistent chronic inflammation and infection by chikungunya arthritogenic alphavirus in spite of a robust host immune response. J Immunol. 2010;184(10):5914–5927. doi:10.4049/jimmunol.0900255
116. Chang AY, Martins KAO, Encinales L, et al. Chikungunya arthritis mechanisms in the americas: a cross-sectional analysis of chikungunya arthritis patients twenty-two months after infection demonstrating no detectable viral persistence in synovial fluid. Arthritis Rheumatol. 2018;70(4):585–593. doi:10.1002/art.40383
117. Chang AY, Encinales L, Porras A, et al. Frequency of chronic joint pain following chikungunya virus infection: a Colombian cohort study. Arthritis Rheumatol. 2018;70(4):578–584. doi:10.1002/art.40384
118. Chang AY, Tritsch S, Reid SP, et al. The cytokine profile in acute chikungunya infection is predictive of chronic arthritis 20 months post infection. Diseases. 2018;6(4):95. doi:10.3390/diseases6040095
119. Costa DMDN, Gouveia PADC, Silva GEDB. The relationship between chikungunya virus and the kidneys: a scoping review. Rev Med Virol. 2023;33(1):e2357. doi:10.1002/rmv.2357
120. Kam YW, Simarmata D, Chow A, et al. Early appearance of neutralizing immunoglobulin G3 antibodies is associated with chikungunya virus clearance and long-term clinical protection. J Infect Dis. 2012;205(7):1147–1154. doi:10.1093/infdis/jis033
121. Auerswald H, Boussioux C, In S, et al. Broad and long-lasting immune protection against various chikungunya genotypes demonstrated by participants in a cross-sectional study in a Cambodian rural community. Emerg Microbes Infect. 2018;7(1):13. doi:10.1038/s41426-017-0010-0
122. Andrew A, Navien TN, Yeoh TS, et al. Diagnostic accuracy of serological tests for the diagnosis of Chikungunya virus infection: a systematic review and meta-analysis. PLoS Negl Trop Dis. 2022;16(2):e0010152. doi:10.1371/journal.pntd.0010152
123. Malik MR, Walaskar S, Majji R, et al. Development of an affordable multiplex quantitative RT-PCR assay for early detection and surveillance of Dengue, chikungunya, and co-infections from clinical samples in resource-limited settings. PLoS Negl Trop Dis. 2025;19(8):e0013250. doi:10.1371/journal.pntd.0013250
124. Santiago GA, Vázquez J, Courtney S, et al. Performance of the trioplex real-time RT-PCR assay for detection of Zika, dengue, and chikungunya viruses. Nat Commun. 2018;9(1):1391. doi:10.1038/s41467-018-03772-1
125. Xu Z, Peng Y, Yang M, et al. Simultaneous detection of Zika, chikungunya, dengue, yellow fever, West Nile, and Japanese encephalitis viruses by a two-tube multiplex real-time RT-PCR assay. J Med Virol. 2022;94(6):2528–2536. doi:10.1002/jmv.27658
126. Simon F, Javelle E, Cabie A, et al. French guidelines for the management of chikungunya (acute and persistent presentations). November 2014. Médecine et Maladies Infectieuses. 2015;45(7):243–263. doi:10.1016/j.medmal.2015.05.007
127. Contopoulos-Ioannidis D, Newman-Lindsay S, Chow C, LaBeaud AD. Mother-to-child transmission of Chikungunya virus: a systematic review and meta-analysis. PLoS Negl Trop Dis. 2018;12(6):e0006510. doi:10.1371/journal.pntd.0006510
128. Gérardin P, Sampériz S, Ramful D, et al. Neurocognitive outcome of children exposed to perinatal mother-to-child chikungunya virus infection: the CHIMERE cohort study on reunion Island. PLoS Negl Trop Dis. 2014;8(7):e2996. doi:10.1371/journal.pntd.0002996
129. Gérardin P, Barau G, Michault A, et al. Multidisciplinary prospective study of mother-to-child chikungunya virus infections on the island of La Réunion. PLoS Med. 2008;5(3):e60. doi:10.1371/journal.pmed.0050060
130. Ferreira FC, Filippis AM, Moreira ME, et al. Perinatal and neonatal chikungunya virus transmission: a case series. J Pediatric Infect Dis Soc. 2024;13(11):576–584. doi:10.1093/jpids/piae102
131. Lenglet Y, Barau G, Robillard PY, et al. Chikungunya infection in pregnancy: evidence for intrauterine infection in pregnant women and vertical transmission in the parturient. Survey of the reunion Island outbreak. J Gynecol Obstet Biol Reprod. 2006;35(6):578–583. doi:10.1016/s0368-2315(06)76447-x
132. Escobar M, Nieto AJ, Loaiza-Osorio S, Barona JS, Rosso F. Pregnant women hospitalized with chikungunya virus infection, Colombia, 2015. Emerg Infect Dis. 2017;23(11):1777–1783. doi:10.3201/eid2311.170480
133. Commissioner O of the. FDA approves first vaccine to prevent disease caused by chikungunya virus. FDA. 2024. Available from: https://www.fda.gov/news-events/press-announcements/fda-approves-first-vaccine-prevent-disease-caused-chikungunya-virus.
134. Commission authorises chikungunya vaccine and funds new mosquito eradication programme*. European Commission - European Commission. Available from: https://ec.europa.eu/commission/presscorner/detail/en/https:\/\/ec.europa.eu\/commission\/presscorner\/detail\/en\/ip_24_3502.
135. Ribeiro Dos Santos G, Jawed F, Mukandavire C, et al. Global burden of chikungunya virus infections and the potential benefit of vaccination campaigns. Nat Med. 2025;31(7):2342–2349. doi:10.1038/s41591-025-03703-w
136. Research C for BE and. FDA update on the safety of ixchiq (Chikungunya Vaccine, Live). FDA. 2025. Available from: https://www.fda.gov/vaccines-blood-biologics/safety-availability-biologics/fda-update-safety-ixchiq-chikungunya-vaccine-live.
137. Richardson JS, Anderson DM, Mendy J, et al. Chikungunya virus virus-like particle vaccine safety and immunogenicity in adolescents and adults in the USA: a phase 3, randomised, double-blind, placebo-controlled trial. Lancet. 2025;405(10487):1343–1352. doi:10.1016/S0140-6736(25)00345-9
138. CDC. Chikungunya vaccine information for healthcare providers. Chikungunya Virus. 2025. Available from: https://www.cdc.gov/chikungunya/hcp/vaccines/index.html.
139. Trojánek M, Grebenyuk V, Manďáková Z, et al. Epidemiology of dengue, chikungunya and Zika virus infections in travellers: a 16-year retrospective descriptive study at a tertiary care centre in Prague, Czech Republic. PLoS One. 2023;18(2):e0281612. doi:10.1371/journal.pone.0281612
140. Gossner CM, Fournet N, Dias JG, et al. Risks related to chikungunya infections among European Union travelers, 2012–2018. Emerg Infect Dis. 2020;26(6):1067–1076. doi:10.3201/eid2606.190490
141. Liu Q, Shen H, Gu L, Yuan H, Zhu W. Chikungunya virus in Europe: a retrospective epidemiology study from 2007 to 2023. PLoS Negl Trop Dis. 2025;19(3):e0012904. doi:10.1371/journal.pntd.0012904
142. Simon F, Caumes E, Jelinek T, Lopez-Velez R, Steffen R, Chen LH. Chikungunya: risks for travellers. J Travel Med. 2023;30(2):taad008. doi:10.1093/jtm/taad008
143. Monteiro S, Pimenta R, Nunes F, Cunha MV, Santos R. Detection of dengue virus and chikungunya virus in wastewater in Portugal: an exploratory surveillance study. Lancet Microbe. 2024;5(11):100911. doi:10.1016/S2666-5247(24)00150-2
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.