Back to Journals » Infection and Drug Resistance » Volume 13
Characterization of Hypervirulent Extended-Spectrum β-Lactamase-Producing Klebsiella pneumoniae Among Urinary Tract Infections: The First Report from Iran
Authors Taraghian A, Nasr Esfahani B, Moghim S ![]() , Fazeli H
, Fazeli H
Received 9 June 2020
Accepted for publication 12 August 2020
Published 9 September 2020 Volume 2020:13 Pages 3103—3111
DOI https://doi.org/10.2147/IDR.S264440
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Azadeh Taraghian, Bahram Nasr Esfahani, Sharareh Moghim, Hossein Fazeli
Department of Microbiology, School of Medicine, Isfahan University of Medical Sciences, Isfahan, Iran
Correspondence: Hossein Fazeli Email [email protected]
Introduction: This study was conducted to identify the hypermucoviscosity, iron acquisition, and capsule serotypes of K. pneumoniae strains isolated from urinary tract infections among community-acquired patients (CA) and assess the frequency of plasmid-mediated quinolone resistance (PMQR) and extended-spectrum beta-lactamases (ESBL) genes between classic and hypervirulent strains.
Materials and Methods: A total of 105 K. pneumoniae were isolated from CA-UTI. Demographic data related to the underlying diseases and clinical manifestations were further collected. Antibiotic resistance pattern and molecular characterization were compared among ESBL-positive, ESBL-negative, hypervirulent, and classic isolates.
Results: The results revealed that 52.4% of the isolates were confirmed as ESBL producers and 11 (10.5%) were considered as hypervirulent K. pneumoniae (hvKp). Ciprofloxacin and nalidixic acid were the most inactive antibiotics with resistance rates of 68.6% and 64.8%, respectively. Molecular characterization revealed that 7.6% of all the isolates carried k1 and 66.6% carried K2 genes. The most frequent ESBL gene was blaSHV 63.8%, followed by blaTEM 59.0%, and blaCTX-M 58.1%. ESBL genes were significantly more in hvKp than in cKp. Moreover, 61 (84.7%), 47 (65.2%), and 16 (22.2%) of isolates harbored qnrB, qnrS, and qnrA. ESBL genes were detected in all hvKps, and blaSHV was observed in 90.9% of hvKp (P value= 0.048, 95%).
Discussion: This study reported the high frequency of antimicrobial and multidrug resistance among hvKp isolates. Coexistence of PMQR and ESBL genes in hvkp indicates the necessity to enhance the clinical knowledge and management of hvKp infections.
Keywords: hypervirulent Klebsiella pneumoniae, urinary tract infections, capsular group, ESBL, qnr
Introduction
Klebsiella pneumoniae is a common Gram-negative organism with worldwide distribution. It causes nosocomial and community-acquired infections, liver abscess, and urinary tract infections, mostly in patients with underlying diseases.1,2 This organism is among the top three pathogens of international concern documented in the World Priority List of Antibiotic-Resistant Bacteria released by the World Health Organization (WHO) in 2018.3 Over the past two decades, K. pneumoniae has been the second most prevalent etiological agent of community-acquired (CA) urinary tract infections (UTI).4 There are mainly two pathotypes that pose a threat to public health: hypervirulent K. pneumoniae (hvKp) and classical K. pneumoniae (cKp). The most common subtype of the K. pneumoniae strains is classic K. pneumoniae (cKp); unlike this subtype, the new “hypervirulent” K. pneumoniae (hvKp) with hypermucoviscosity has emerged as a clinically important infectious agent causing extremely invasive infections such as liver abscesses in both healthy and immunocompromised individuals.5,6 These strains can cause serious CA infections in healthy individuals, along with the reported global dissemination of the growing antibiotic resistance.7,8 Therefore, hvKp increases the public health costs, restricts the selection of antibiotics, and causes failure in medical therapy, leading to mortality and increased morbidity.9
The hvKp strains are of hypermucoviscous type due to the overproduction of the capsular polysaccharide. Hypermucoviscosity is a crucial in vitro parameter for identifying hvKp.10 Moreover, several virulence genes such as the regulator of mucoid phenotype A gene (rmpA) and aerobactin (iucA) have been identified to be associated with the hypervirulent variant of K. pneumoniae.11 Furthermore, the capsule, particularly serotypes K1 and K2, has long been known as a major virulence factor in K. pneumoniae.12
On one hand, fluoroquinolones such as ciprofloxacin and levofloxacin as well as β-lactam antibiotics are commonly recommended for complicated UTI treatment.13 On the other hand, there has been a global increase in extended-spectrum β-lactamases (ESBL)-producing Enterobacteriaceae and fluoroquinolone resistance.14 Plasmid-mediated quinolone resistance (PMQR) determinants have been further reported. PMQR includes Qnr (quinolone resistance) proteins (qnrA, qnrB, qnrC, qnrD, and qnrS) which protect the topoisomerase IV and DNA gyrase from quinolone inhibition, which acetylates aminoglycoside and ciprofloxacin and reduces their activity.15 PMQR genes are mostly on the same plasmid as ESBL genes.16 Resistance plasmids harboring genes encoding ESBLs can be transferred by conjugation, which helps disseminate PMQR determinants in different Enterobacteriaceae species.17
Multidrug-resistant (MDR) K. pneumoniae isolates are defined by non-susceptibility to at least one agent in three or more antibiotic categories, which severely limits antimicrobial treatment options.18 Hence, the coexistence of MDR-ESBLs and PMQR genes is a main concern, particularly in the community status.19,20 ESBL producers have been associated with resistance to non-β-lactam antimicrobials, including fluoroquinolones, aminoglycosides, and sulfonamides, which are often used to treat and prevent diseases. Additionally, plasmid-mediated quinolone resistance (PMQR) genes are associated with ESBL production, and the spread of co-expressing PMQRs and ESBL isolates contributes to the growing concerns regarding MDR isolates.21 Also, hvKp strains are of major importance as they are hypervirulent and highly transmissible.18
To our knowledge, there is a paucity of information concerning the frequency of hvKp in CA-UTI and their antimicrobial susceptibility. Therefore, the objective of this study was to identify the hypermucoviscosity, iron acquisition, and capsule serotypes of K. pneumoniae strains isolated from CA-UTI; the frequency of PMQR (qnrA, qnrB and qnrS) and ESBL genes (blaSHV, blaTEM, blaCTX-M) in ESBL and non-ESBL producer isolates was further assessed. In addition, cKp and hvKp strains were compared in terms of antimicrobial susceptibility and molecular characterizations.
Materials and Methods
A cross-sectional study was conducted at Milad Advanced Medical laboratory, Isfahan, Iran, between April and December 2019. All patient data accessed complied with the relevant data protection and privacy regulations. This study protocol was approved by the Ethics Committee of Isfahan University of Medical Sciences (IR.MUI.MED.REC.1398.388), but no human participants were included. Urine samples were collected from outpatients who were suspected of UTI and had not consumed any antibiotics for the past two weeks. Samples were collected from the clean-catch midstream urine of adults and children aged less than two years using a sterile urine bag or a suprapubic catheter for sample collection. Demographic data and information related to underlying diseases and clinical manifestations were further collected.4 Urine samples were transferred to the microbiology laboratory at Isfahan University of medical science, then cultivated on blood agar and EMB media (Merck, Germany) for 24 hours. As described previously, a minimum colony count of 105 CFU/mL was considered as positive urine culture for midstream urine.22 Cases of mixed infection were excluded where they might reflect contamination with normal flora of the periurethral, vaginal, and perianal regions.23 All isolates were primarily screened by their pigment production, colony morphology, and Gram-staining techniques. Isolates were then identified by conventional biochemical tests and standard culture methods. In this regard, citrate and urea positive, oxidase, indole, methyl red, and motility negative strains were identified as K. pneumoniae.
String Test
Isolates were considered positive for the hypermucoviscosity phenotype after subculture on blood agar at 37 C overnight if an inoculation loop touching the surface of the colony generated a viscous string of 5 mm or more when pulled away from the plate.24
Antimicrobial Susceptibility Testing
The antibiotic susceptibility profile was determined on Muller–Hinton agar (Merck, Germany) plates using the disk diffusion method (the modified Kirby–Bauer assay) as described by the Clinical and Laboratory Standards Institute (CLSI 2019).25 Quality control strains were included to monitor the test performance. K. pneumoniae clinical isolates were tested for their susceptibility to the following antimicrobial agents: amoxicillin-clavulanic acid (20/10 µg), cefotaxime (30 µg), ceftazidime (30 µg), cefixime (5 μg), cefepime (30 µg), imipenem (10 µg), meropenem (10 µg), gentamicin (10 µg), tobramycin (10 µg), amikacin (30 µg), trimethoprim–sulfamethoxazole (30 µg), nitrofurantoin (300 µg), ciprofloxacin (5 µg), nalidixic acid (30 µg), levofloxacin (5 µg), ofloxacin (5 µg), and piperacillin-tazobactam (100/10 µg).25,26 All disks were obtained from ROSCO Company, Denmark.
Screening of ESBL Producers
The preliminary screening test to detect ESBL activity was performed by the disk diffusion method according to the CLSI guidelines (2019). The inhibition zone size of ≤22 mm for ceftazidime (30 µg) and ≤27 mm for cefotaxime (30 µg) indicated ESBL production.27 The phenotypic confirmatory test for ESBL was carried on by combination disk method using cefotaxime (30 µg) and ceftazidime (30 µg) alone and in combination with clavulanic acid (10 µg).28,29 ESBL activities were identified by ≥5 mm increase in zone diameter around the disk with the antibiotic combined with clavulanic acid.30,31 In brief, Muller Hinton agar plates were swabbed with 0.5 mL of overnight bacterial culture; after 10 minutes the antibiotic disk was placed on the plates and incubated for 24. Afterwards, the inhibition zone was measured and reported in diameter.32 E. coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as the quality control strains.25,33
Molecular Detection of Capsular Types and Virulence Genes
Genomic DNA was extracted from all K. pneumoniae isolates as described previously.34 Polymerase chain reaction (PCR) for virulence-associated genes (such as rmpA and iucA) was carried out among the string test positive isolates. Moreover, using K1 and K2 specifying primers, all isolates were tested for the presence of K. pneumoniae capsular types.5
Molecular Detection of ESBL and PMQR Genes
ESBL genes (blaSHV, blaTEM, blaCTX-M) were detected in all isolated uropathogens.35 In addition, isolates that were phenotypically resistant to at least one fluoroquinolone were screened for qnrA, qnrB, and qnrS genes. PCR primers were previously described.10 Amplification was conducted as follows: initial denaturation at 95°C for five minutes, followed by 35 cycles at 94°C for 30 seconds, 55°C for 30 seconds, and 72°C for 30 seconds, and a final extension at 72°C for five minutes. The PCR products were analyzed by electrophoresis with 1% agarose gel in 1X Tris–Acetate-EDTA buffer. The gels were stained with stain load dye (CinnaGen Co, Iran) and the PCR products were visualized under UV light.36,37
Statistical Analysis
The relationship among antibiotic resistance, ESBL production, and PMQR determinants was evaluated by the Chi-square test or Fisher’s exact test. P-values≤0.05 were considered as statistically significant. The data were analyzed using the Statistical Package for Windows v.19.0 (SPSS Inc., Chicago, IL, USA).
Results
Clinical Characteristics
Over the eight-month study period, 105 K. pneumoniae were isolated from the urine samples of CA patients in Isfahan. The mean age of patients was 46.50 years (1–81 years), and they comprised 32 (30.5%) males and 73 (69.5%) females. According to the demographic data of the patients, most had underlying diseases, including diabetes mellitus (33.5%), kidney transplant (13.3%), chronic renal insufficiency (20.9%), and immunosuppression (11.4%).
Out of the 105 positive urine cultures, 55 (52.4%) were confirmed as ESBL producer based on the combination disk method. MDR phenotype was detected in 59 out of 105 (56.1%) isolates. Of these 105 isolates, 94 (89.5%) were considered as cKp. Therefore, 11 isolates (10.5%) were string test positive and had two virulence genes (iucA and rmpA), hence considered as hvKp. As shown in Table 1, among these hvKp isolates, nine were females (81.8%), five (45.4%) were identified as MDR, and seven (63.6%) produced ESBL phenotype. Patients suffering from hvKp isolates had risk factors such as diabetes (7/11 (63.6%)), kidney transplant (3/11 (27.2%)), and immunosuppression (2/11 (18.1%)). Clinical and microbiological characteristics of the hvKp isolates are shown in Table 1.
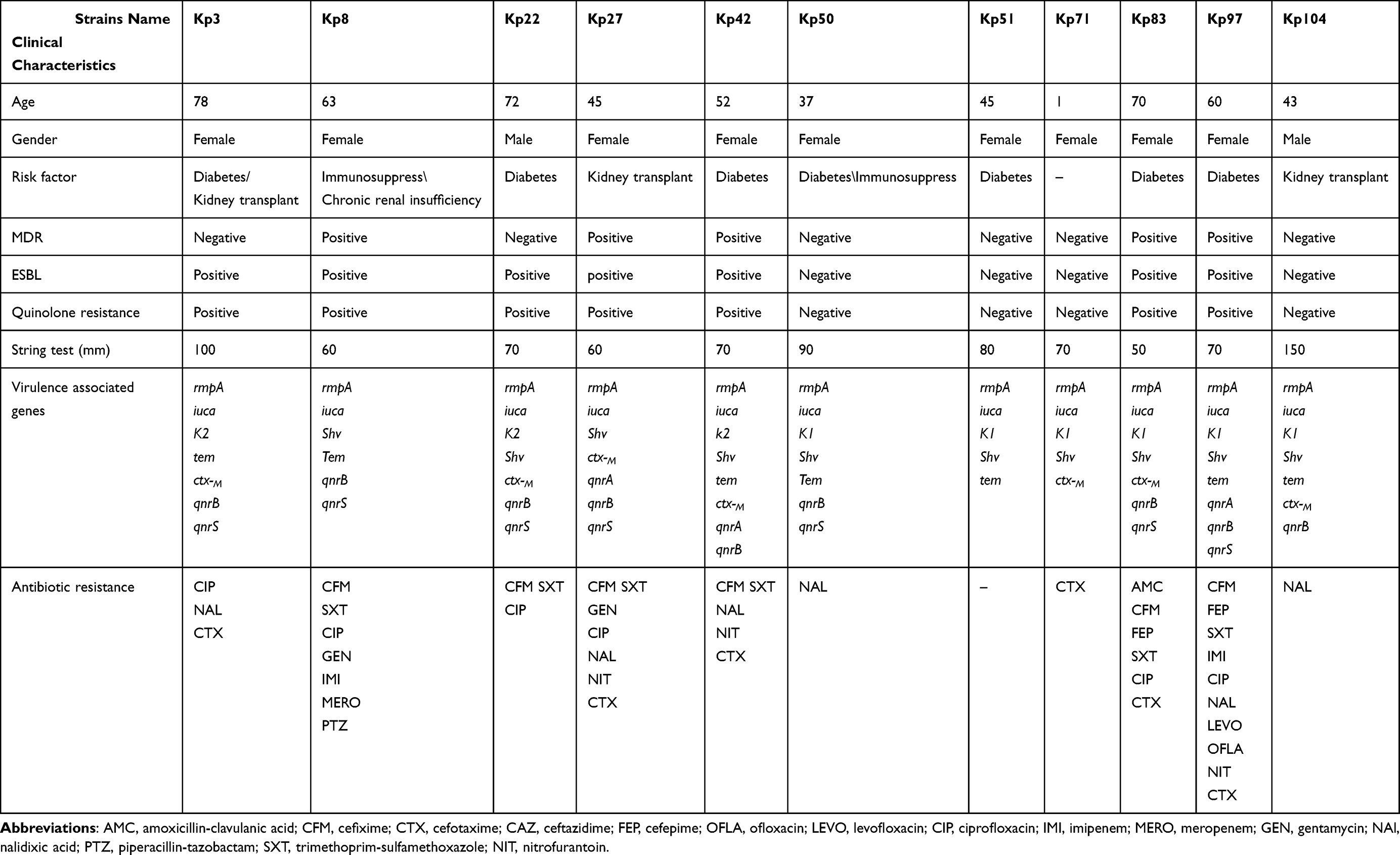 |
Table 1 Clinical and Microbiological Characteristics of hvKp Isolates |
Antimicrobial Resistance
The results revealed that ciprofloxacin and nalidixic acid were the most inactive antibiotics with resistance rates of 68.6% and 64.8%, respectively. However, the most effective antibiotics were meropenem, imipenem, amikacin, tobramycin, and piperacillin-tazobactam with resistance rates of 6.7%, 7.6% and 9.5%, respectively. There was no significant association among antimicrobial resistance, age, and gender (P>0.05). According to statistical analysis, the resistance rates of cefixime, cefotaxime, ciprofloxacin, ofloxacin, levofloxacin, and nalidixic acid were significantly higher in ESBL-positive than in ESBL-negative strains (P ≤0.05). The resistance rates to most tested antimicrobial agents were lower in hvKp than in cKp isolates. Nonetheless, cefixime, trimethoprim–sulfamethoxazole, nitrofurantoin, and carbapenems had higher resistance rates. Table 2 depicts the complete results of antibiotic resistance patterns and compares the ESBL-negative and ESBL-positive isolates.
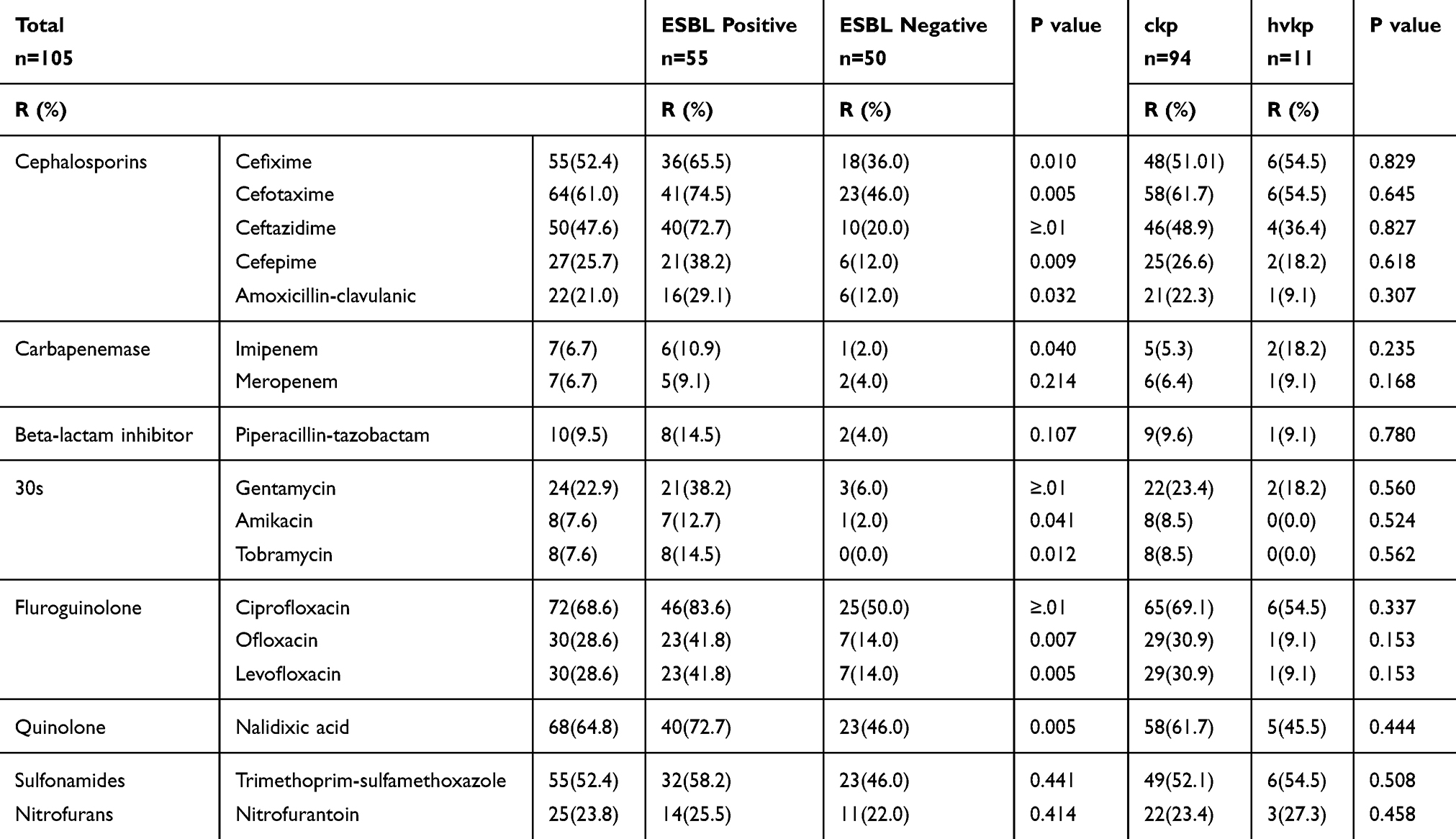 |
Table 2 Percentage of Antimicrobial Resistance of K. pneumoniae |
Molecular Analysis
Capsular Types
In this study, 8/105 (7.6%) isolates carried k1 capsule gene, all belonging to hvKp (P <0.0001). Furthermore, 70/105 (66.6%) carried K2 capsule gene, with only three (27.3%) associated with hvKp strain (P=0.005, Table 3). Twenty-seven isolates did not have k1 and K2 genes.
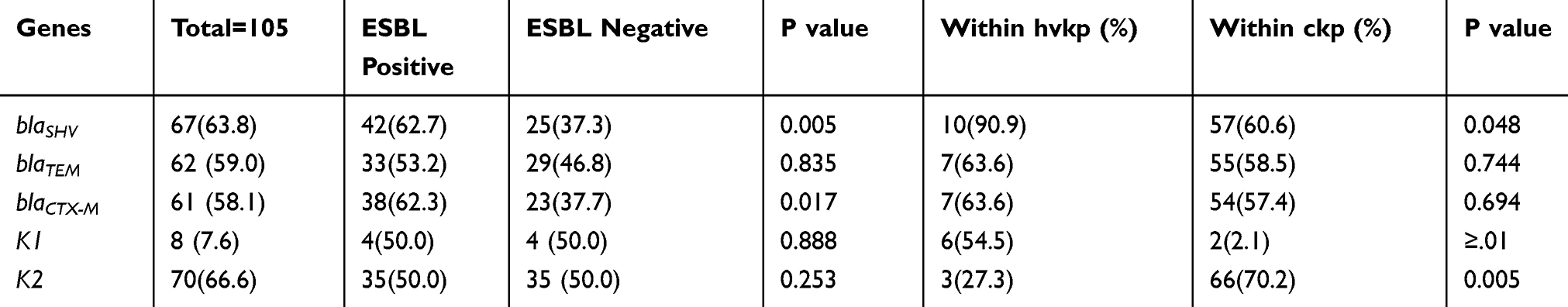 |
Table 3 Frequency of Different ESBL Genes and Capsular Serotype, Among ESBL Positive, ESBL Negative, hvkp and ckp |
Detection of ESBL Genes
Although ESBLs were phenotypically detected in 55 (52.4%) of the isolates, electrophoresis analysis revealed that all isolates had at least one ESBL gene. Table 3 represents the frequency of these genes among ESBL-positive and negative isolates. The most frequent ESBL gene in the tested isolates was blaSHV 67 (63.8%), followed by blaTEM 62 (59.0%) and blaCTX-M 61 (58.1%). ESBL genes were more observed in hvKp than in cKp. According to the table, blaSHV was found in 90.9% of hvKp group and 60.6% of cKp group (P= 0.048).
Detection of PMQR Genes
In the current study, ESBL-producing isolates had a high level of resistance to all tested quinolones compared with non-ESBL-producing ones. In addition, 72 isolates that were resistant to at least one quinolone antibiotics were tested for qnr genes. The most common qnr gene was qnrB 61 (84.7%), followed by qnrS 47 (65.2%) and qnrA 16 (22.2%). Table 4 describes the distribution of gnr genes among ESBL-positive, ESBL-negative, hvkp, and ckp strains.
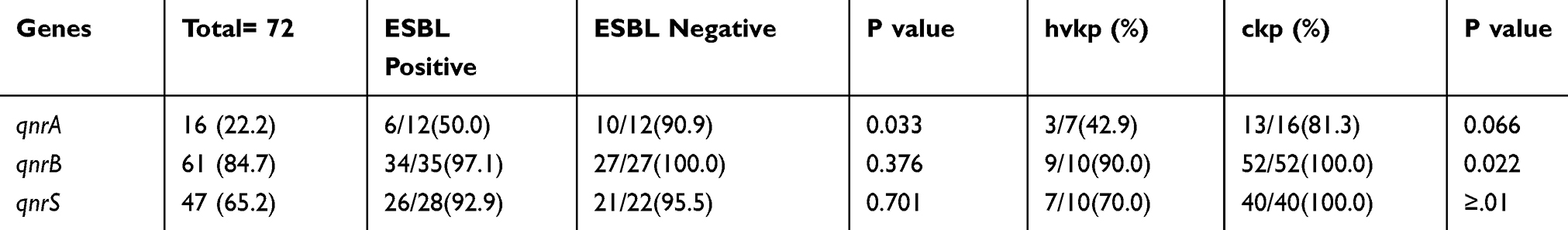 |
Table 4 Frequency of Different qnr Genes, Among ESBL Positive, ESBL Negative, hvkp and ckp Strains |
Results revealed that qnr subtypes could be present alone or in association with blaCTX−M, blaTEM, and blaSHV genes. The simultaneous presence of two or three qnr genes was also detected. qnrBS combinations were most frequently associated with blaCTX-M (54.11%), and 59.7% (43/72) of the isolates had both qnrB and blaSHV genes. Statistical analysis showed that unlike ESBL genes, qnr genes were mostly present in cKp isolates (P-value<0.05 (Table 4)).
Discussion
To our knowledge, this is the first report on the prevalence of hvKp isolates among UTI samples in Iran. Over the recent years, hvKp has emerged as a global health issue and a potential “superbug” for clinical settings, causing a wide variety of infections among community patients.38 In the current study, using a positive string test, 10.5% of K. pneumoniae were identified as hypervirulent, which was confirmed by the presence of the rmpA and iucA genes. Lin et al reported lower amount of hvkp in UTI samples (27.8%).4 In the present study, hvKp strain caused UTI mostly among old female patients. Diabetes mellitus was the most prevalent underlying disease 7/11 (63.6%). A previous study implied that patients with diabetes ran a higher risk for symptomatic UTI.39 Other contributing underlying diseases were chronic renal insufficiency, kidney transplant, and immunosuppression. These results are similar to the previous findings on UTI in China.4
Consistent with previous reports, the rate of resistance to common antibiotics in hvKp strains was lower than that in the cKp group with the exception of cefixime, trimethoprim-sulfamethoxazole, nitrofurantoin, and carbapenems.40 It is widely accepted that carbapenemase-producing hvKp (CR-hvKp) strains cause various life-threatening infections, especially in critical patients.41,42
Empirical use of beta-lactam drugs for CA-UTI ensues increased rates of resistance and outbreaks. In the present study, 52.4% of isolates were ESBL producers, which is higher than that obtained in Taiwan (20.7%) and South Korea (30.0%).43,44 In Turkey (2019), on the other hand, 47% of Klebsiella spp. were ESBL producers.45 These differences might be attributed to the excessive use of antibiotics and differences in risk factors. Unfortunately, phenotypic ESBLs were more in hvkp strains (7 (63.6%)) than in cKp (48 (51.1%)). In a study carried out in China in 2010, only 17% of hvkp were ESBL producers.40 In the current study, all hvKps carried ESBL genes, implying the compatibility of plasmids containing these genes with hvKp strains.46
Additionally, MDR-hvKp was identified in 5 out of 11 (45.4%) hvKp isolates. This shows the resistance to at least one agent in three or more antimicrobial categories, which is higher than a study conducted in China (2019), where 20% MDR-hvKp was detected.6 Taken together, these data reveal that MDR-hvKp isolates are emerging and antimicrobial resistance is growing among these strains, which is similar to previous studies in China.6 Therefore, new clinical and medical interventions concerning these strains are needed.40
Serotypes K1 and K2 have been generally considered as predominant virulent strains of K. pneumoniae, causing high resistance to phagocytosis.47 In this research, 54.5% of hvKp possessed k1 gene (p value<0.001) whereas 27.3% had K2 gene (p value= 0.005). This is in accordance with a study carried out in the US in 2018 (55% k1 vs 20% k2) and another performed in China in 2016 (33.3% k1 vs 17.2 k2).46,48
The resistance rates of the antibiotics to ESBL-positive strains between CA-UTIs were significantly higher than those of ESBL-negative strains (P<0.05) (Table 2). Based on these results, ESBL-producing Klebsiella spp. are not merely a nosocomial problem, rather they are a growing issue in CA-UTIs.
The PCR revealed the prevalence of ESBL genes in the current study. Nevertheless, we found that blaSHV was the most prevalent ESBL gene, followed by blaTEM and blaCTX-M. In a similar investigation done in Iraq (2019), 64.7% blaTEM, 35.2% blaSHV, and 41.1% blaCTX-M genes existed in the isolates of K. pneumoniae.49
ESBL producer strains are mostly resistant to common antibiotics such as fluoroquinolones.50 Moreover, plasmid-mediated quinolone resistance might facilitate the spread and increased frequency of quinolone-resistant strains. A high proportion of isolates (68%) in this study were resistant to fluoroquinolones. According to the findings, resistance to the tested fluoroquinolones in ESBL-producing isolates was significantly higher than in non-ESBL-producing isolates, which is similar to another study performed in the Asia-Pacific region.51 In the current research, 46 (83.6%) of ESBL-positive K. pneumoniae isolates were resistant to ciprofloxacin, and their prevalence was higher than that of Meshkat (37.5%) in 2017.52
So far, qnr genes have been widely detected in different parts of the world.53 The frequencies found in the current research were higher than those reported in a Moroccan community in 2014, where the prevalence of qnr gene between 50% Klebsiella and 18.7% E. Coli was 23% qnrB, 10% qnr A, and 3% qnrS.54 PMQR are plasmid-mediated and can be easily transmitted across the Enterobacteriaceae family members, thereby facilitating the epidemic spread of quinolone resistance through horizontal gene transfer.55 Coexistence of PMQR and ESBL genes was identified in four out of 11 hvKp isolates. Although ESBL genes were mostly observed in hvKp than in cKp, qnr genes mostly existed in cKp strains (P< 0.05). These results imply that hvKp strains are capable of acquiring various types of antibiotic resistance mechanisms via the transmission of mobile genetic elements. In addition to the disconcerting antimicrobial resistance pattern of hvKp strains, these isolates underscore the need for specific surveillance programs.
Conclusions
In this study, the frequency of hvkp was lower than similar studies in other regions; however, the higher rates of resistance to antimicrobial agents, particularly carbapenems, and the high rates of MDR emergence indicate that these strains have emerged as major issues in health-care centers. Moreover, the coexistence of PMQR and ESBL genes severely affects the clinical outcomes due to the complications in therapy, hence the necessity to enhance the clinical knowledge and management of hvKp infections. It is also important to take appropriate supportive measures regarding empirical antibiotic therapy so as to control the spread of resistant strains.
Acknowledgments
The authors would like to acknowledge the Department of Microbiology of Isfahan University of Medical Sciences.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Melot B, Colot J, Guerrier G. Bacteremic community-acquired infections due to Klebsiella pneumoniae: clinical and microbiological presentation in New Caledonia, 2008–2013. Int J Infect Dis. 2015;41:29–31. doi:10.1016/j.ijid.2015.10.013
2. Ghanavati R, Darban-Sarokhalil D, Navab-Moghadam F, Kazemian H, Irajian G, Razavi S. First report of coexistence of AmpC beta-lactamase genes in Klebsiella pneumoniae strains isolated from burn patients. Acta Microbiol Immunol Hung. 2017;64(4):455–462. doi:10.1556/030.64.2017.028
3. Shrivastava SR, Shrivastava PS, Ramasamy J. World health organization releases global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. J Med Soc. 2018;32(1):76.
4. Lin W-H, Wang M-C, Tseng -C-C, et al. Clinical and microbiological characteristics of Klebsiella pneumoniae isolates causing community-acquired urinary tract infections. Infection. 2010;38(6):459–464.
5. Prokesch BC, TeKippe M, Kim J, Raj P, TeKippe EM, Greenberg DE. Primary osteomyelitis caused by hypervirulent Klebsiella pneumoniae. Lancet Infect Dis. 2016;16(9):e190–e5. doi:10.1016/S1473-3099(16)30021-4
6. Liu C, Guo J. Hypervirulent Klebsiella pneumoniae (hypermucoviscous and aerobactin positive) infection over 6 years in the elderly in China: antimicrobial resistance patterns, molecular epidemiology and risk factor. Ann Clin Microbiol Antimicrob. 2019;18(1):4. doi:10.1186/s12941-018-0302-9
7. Zhang Y, Zeng J, Liu W, et al. Emergence of a hypervirulent carbapenem-resistant Klebsiella pneumoniae isolate from clinical infections in China. J Infect. 2015;71(5):553–560.
8. Zhan L, Wang S, Guo Y, et al. Outbreak by hypermucoviscous Klebsiella pneumoniae ST11 isolates with carbapenem resistance in a tertiary hospital in China. Front Cell Infect Microbiol. 2017;7:182.
9. Patel PK, Russo TA, Karchmer AW, editors. Hypervirulent Klebsiella pneumoniae. In: Open Forum Infectious Diseases. Oxford University Press; 2014.
10. Liu YM, Zhang YY, Zhang W, Shen H, Li H, Cao B. Clinical and molecular characteristics of emerging hypervirulent Klebsiella pneumoniae bloodstream infections in mainland China. Antimicrob Agents Chemother. 2014;58:5379–5385. doi:10.1128/AAC.02523-14
11. Ye M, Tu J, Jiang J, et al. Clinical and genomic analysis of liver abscess-causing Klebsiella pneumoniae identifies new liver abscess-associated virulence genes. Front Cell Infect Microbiol. 2016;6(165). doi:10.3389/fcimb.2016.00165
12. Zhao Y, Zhang X, Torres VL, et al. An outbreak of carbapenem-resistant and hypervirulent Klebsiella pneumoniae in an intensive care unit of a major teaching hospital in Wenzhou, China. Front Public Health. 2019;7:229.
13. Bajaj P, Singh NS, Virdi JS. Escherichia coli β-lactamases: what really matters. Front Microbiol. 2016;7:417.
14. Ni Q, Tian Y, Zhang L, et al. Prevalence and quinolone resistance of fecal carriage of extended-spectrum β-lactamase-producing Escherichia coli in 6 communities and 2 physical examination center populations in Shanghai, China. Diagn Microbiol Infect Dis. 2016;86(4):428–433.
15. Strahilevitz J, Jacoby GA, Hooper DC, Robicsek A. Plasmid-mediated quinolone resistance: a multifaceted threat. Clin Microbiol Rev. 2009;22(4):664–689.
16. García-Fulgueiras V, Bado I, Mota MI, et al. Extended-spectrum β-lactamases and plasmid-mediated quinolone resistance in enterobacterial clinical isolates in the paediatric hospital of Uruguay. J Antimicrob Chemother. 2011;66(8):1725–1729. doi:10.1093/jac/dkr222
17. Rodríguez-Martínez JM, Velasco C, Pascual Á, Cano ME. Plasmid-mediated quinolone resistance: an update. J Infect Chemother. 2011;17(2):149–182. doi:10.1007/s10156-010-0120-2
18. Magiorakos A-P, Srinivasan A, Carey R, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281.
19. Mansouri S, Abbasi S. Prevalence of multiple drug resistant clinical isolates of extended-spectrum beta-lactamase producing Enterobacteriaceae in Southeast Iran. Iran J Med Sci. 2015;35(2):101–108.
20. Di Tella D, Tamburro M, Guerrizio G, et al. Molecular epidemiological insights into Colistin-resistant and Carbapenemases-producing clinical Klebsiella pneumoniae isolates. Infect Drug Resist. 2019;12:3783.
21. Liu X, Liu H, Wang L, et al. Molecular characterization of extended-spectrum β-lactamase-producing multidrug resistant Escherichia coli from swine in Northwest China. Front Microbiol. 2018;9:1756.
22. CDC/HICPAC. Urinary Tract Infection (UTI) Event for Long- Term Care Facilities. Atlanta, GA, USA: CDC; 2009.
23. Sathiananthamoorthy S, Malone-Lee J, Gill K, et al. Reassessment of routine midstream culture in diagnosis of urinary tract infection. J Clin Microbiol. 2019;57(3):e01452–18.
24. Cubero M, Marti S, MÁ D, González-Díaz A, Berbel D, Ardanuy C. Hypervirulent Klebsiella pneumoniae serotype K1 clinical isolates form robust biofilms at the air-liquid interface. PLoS One. 2019;14(9):e0222628.
25. Clinical and Laboratory Standards Institute. 2019. Performance standards for antimicrobial susceptibility testing; Twentynineth informational supplement CdM-SCaLSI, Wayne, PA.
26. Shoja S, Ansari M, Faridi F, et al. Identification of carbapenem-resistant Klebsiella pneumoniae with emphasis on New Delhi metallo-beta-lactamase-1 (bla NDM-1) in Bandar Abbas, South of Iran. Microb Drug Resist. 2018;24(4):447–454.
27. Rajivgandhi G, Maruthupandy M, Veeramani T, Quero F, Li WJ. Anti-ESBL investigation of chitosan/silver nanocomposites against carbapenem resistant Pseudomonas aeruginosa. Int J Biol Macromol. 2019;132:1221–1234.
28. Rajivgandhi G, Maruthupandy M, Ramachandran G, Priyanga M, Manoharan N. Detection of ESBL genes from ciprofloxacin resistant Gram negative bacteria isolated from urinary tract infections (UTIs). Front Lab Med. 2018;2(1):5–13.
29. Rajivgandhi G, Vijayan R, Maruthupandy M, Vaseeharan B, Manoharan N. Antibiofilm effect of Nocardiopsis sp. GRG 1 (KT235640) compound against biofilm forming Gram negative bacteria on UTIs. Microb Pathog. 2018;118:190–198.
30. Rajivgandhi G, Ramachandran G, Maruthupandy M, Senthil R, Vaseeharan B, Manoharan N. Molecular characterization and antibacterial investigation of marine endophytic actinomycetes Nocardiopsis sp. GRG 2 (KT 235641) compound against isolated ESBL producing bacteria. Microb Pathog. 2018.
31. Rajivgandhi G, Maruthupandy M, Muneeswaran T, et al. Biologically synthesized copper oxide nanoparticles enhanced intracellular damage in ciprofloxacin resistant ESBL producing bacteria. Microb Pathog. 2019;127:267–276.
32. Maruthupandy M, Muneeswaran T, Song J-M, Manoharan N. Biologically synthesized zinc oxide nanoparticles as nanoantibiotics against ESBLs producing gram negative bacteria. Microb Pathog. 2018;121:224–231. doi:10.1016/j.micpath.2018.05.041
33. Halaji M, Shahidi S, Atapour A, et al. Characterization of extended-spectrum β-lactamase-producing uropathogenic Escherichia coli among Iranian kidney transplant patients. Infect Drug Resist. 2020;13:1429.
34. Chen W-P, Kuo -T-T. A simple and rapid method for the preparation of gram-negative bacterial genomic DNA. Nucleic Acids Res. 1993;21(9):2260.
35. Rajivgandhi G, Maruthupandy M, Manoharan N. Detection of TEM and CTX-M genes from ciprofloxacin resistant Proteus mirabilis and Escherichia coli isolated on urinary tract infections (UTIs). Microb Pathog. 2018;121:123–130.
36. Dallenne C, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp498
37. Chen X, Zhang W, Pan W, et al. Prevalence of qnr, aac(6′)-Ib-cr, qepA, and oqxAB in Escherichia coli isolates from humans, animals, and the environment. Antimicrob Agents Chemother. 2012;56(6):3423–3427. doi:10.1128/AAC.06191-11
38. Lee C-H, Chuah S-K, Tai W-C, Chang -C-C, Chen F-J. Delay in human neutrophil constitutive apoptosis after infection with Klebsiella pneumoniae serotype K1. Front Cell Infect Microbiol. 2017;7:87.
39. Stamm WE, Hooton TM, Johnson JR, et al. Urinary tract infections: from pathogenesis to treatment. J Infect Dis. 1989;159(3):400–406.
40. Li W, Sun G, Yu Y, et al. Increasing occurrence of antimicrobial-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae isolates in China. Clin Infect Dis. 2014;58(2):225–232.
41. Zhang R, Lin D, Chan EW-C, et al. Emergence of carbapenem-resistant serotype K1 hypervirulent Klebsiella pneumoniae strains in China. Antimicrob Agents Chemother. 2016;60(1):709–711.
42. Ripabelli G, Sammarco ML, Scutellà M, Felice V, Tamburro M. Carbapenem-resistant KPC-and TEM-producing Escherichia coli ST131 isolated from a hospitalized patient with urinary tract infection: first isolation in Molise region, Central Italy, July 2018. Microb Drug Resist. 2020;26(1):38–45.
43. Kao C-Y, Udval U, Huang Y-T, et al. Molecular characterization of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella spp. isolates in Mongolia. J Microbiol Immunol Infect. 2016;49(5):692–700.
44. Yang Y-S, Ku C-H, Lin J-C, et al. Impact of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae on the outcome of community-onset bacteremic urinary tract infections. J Microbiol Immunol Infect. 2010;43(3):194–199.
45. Koksal E, Tulek N, Sonmezer MC, et al. Investigation of risk factors for community-acquired urinary tract infections caused by extended-spectrum beta-lactamase Escherichia coli and Klebsiella species. Investig Clin Urol. 2019;60(1):46–53.
46. Zhang Y, Zhao C, Wang Q, et al. High prevalence of hypervirulent klebsiella pneumoniae infection in china: geographic distribution, clinical characteristics, and antimicrobial resistance. Antimicrob Agents Chemother. 2016;60:6115–6120. doi:10.1128/AAC.01127-16
47. Lin J-C, Chang F-Y, Fung C-P, et al. High prevalence of phagocytic-resistant capsular serotypes of Klebsiella pneumoniae in liver abscess. Microbes Infect. 2004;6(13):1191–1198.
48. Russo TA, Olson R, Fang C-T, et al. Identification of biomarkers for differentiation of hypervirulent Klebsiella pneumoniae from classical K. pneumoniae. J Clin Microbiol. 2018;56(9):e00776–18.
49. Pishtiwan AH, Khadija KM. Prevalence of blaTEM, blaSHV, and blaCTX-M genes among ESBL-producing Klebsiella pneumoniae and Escherichia coli isolated from thalassemia patients in Erbil, Iraq. Mediterr J Hematol Infect Dis. 2019;11(1).
50. Yadav KK, Khadka R, Pant AD, Shah B. Multidrug resistant Enterobacteriaceae and extended spectrum beta-lactamase producing Escherichia coli: a cross-sectional study in National Kidney Center, Nepal. Antimicrob Resist Infect Control. 2015;4:42.
51. Lu P-L, Liu Y-C, Toh H-S, et al. Epidemiology and antimicrobial susceptibility profiles of Gram-negative bacteria causing urinary tract infections in the Asia-Pacific region: 2009–2010 results from the study for monitoring antimicrobial resistance trends. Int J Antimicrob Agents. 2012;40:S37–S43. doi:10.1016/S0924-8579(12)70008-0
52. Izadi N, Nasab MN, Mood EH, Meshkat Z. The frequency of qnr genes in extended-spectrum β-lactamases and non-ESBLs Klebsiella pneumoniae species isolated from patients in Mashhad, Iran. Iran J Pathol. 2017;12(4):377.
53. Salah FD, Soubeiga ST, Ouattara AK, et al. Distribution of quinolone resistance gene (qnr) in ESBL-producing Escherichia coli and Klebsiella spp. in Lomé, Togo. Antimicrob Resist Infect Control. 2019;8(1):104.
54. Jamali L, Haouzane F, Bouchajou M. Prevalence of plasmid mediated quinolone resistance genes among enterobacteria isolates in Moroccan community. Int J Innov Sci Res J. 2014;11:387–399.
55. Jacoby G, Strahilevitz J, Hooper D. Plasmid-mediated quinolone resistance. Microbiol Spectr. 2014;2. doi:10.1128/microbiolspec.PLAS-0006-2013
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.