Back to Journals » Journal of Inflammation Research » Volume 15
Characterization of Early Inflammatory Events Leading to Provoked Vulvodynia Development in Rats
Authors Awad-Igbaria Y ![]() , Dadon S, Shamir A, Livoff A, Shlapobersky M, Bornstein J
, Dadon S, Shamir A, Livoff A, Shlapobersky M, Bornstein J ![]() , Palzur E
, Palzur E ![]()
Received 29 March 2022
Accepted for publication 16 June 2022
Published 11 July 2022 Volume 2022:15 Pages 3901—3923
DOI https://doi.org/10.2147/JIR.S367193
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Yaseen Awad-Igbaria,1,2 Shilo Dadon,1,2 Alon Shamir,3,4 Alejandro Livoff,5 Mark Shlapobersky,5 Jacob Bornstein,1,2,* Eilam Palzur2,*
1Faculty of Medicine in the Galilee, Bar-Ilan University, Safed, Israel; 2The Research Institute of Galilee Medical Center, Nahariya, Israel; 3Psychobiology Research Laboratory, Mazor Mental Health Center, Akko, Israel; 4The Ruth and Bruce Rappaport Faculty of Medicine, Technion – Israel Institute of Technology, Haifa, Israel; 5Pathology Department, Barzilai University Medical Center, Ashkelon, Israel
*These authors contributed equally to this work
Correspondence: Yaseen Awad-Igbaria, The Research Institute of Galilee Medical Center, PO Box 21, Nahariya, 22100, Israel, Tel +972 50-4500101, Email [email protected]
Background: Provoked vulvodynia (PV) is the main cause of vulvar pain and dyspareunia. The etiology of PV has not yet been elucidated. However, PV is associated with a history of recurrent inflammation, and its often accompanied by increases in the numbers of mast cells (MCs) and sensory hyperinnervation in the vulva. Therefore, this study aimed to examine the role of MCs and the early inflammatory events in the development of chronic vulvar pain in a rat model of PV.
Methods: Mechanical and thermal vulvar sensitivity was measured for 5 months following zymosan vulvar challenges. Vulvar changes in glutamate and nerve growth factor (NGF) were analyzed using ELISA. Immunofluorescence (IF) staining of the vulvar section after 20, 81, and 160 days of the zymosan challenge were performed to test MCs accumulation, hyperinnervation, and expression of pain channels (transient receptor potential vanilloid/ankyrin-1-TRPV1 & TRPA1) in vulvar neurons. Changes in the development of vulvar pain were evaluated following the administration of the MCs stabilizer ketotifen fumarate (KF) during zymosan vulvar challenges.
Results: Zymosan-challenged rats developed significant mechanical and thermal vulvar sensitivity that persisted for over 160 days after the zymosan challenge. During inflammation, increased local concentrations of NGF and glutamate and a robust increase in MCs degranulation were observed in zymosan-challenged rats. In addition, zymosan-challenged rats displayed sensory hyperinnervation and an increase in the expression of TRPV1 and TRPA1. Treatment with KF attenuated the upregulated level of NGF during inflammation, modulated the neuronal modifications, reduced MCs accumulation, and enhanced mechanical hypersensitivity after repeated inflammation challenges.
Conclusion: The present findings suggest that vulvar hypersensitivity is mediated by MCs accumulation, nerve growth, and neuromodulation of TRPV1 and TRPA1. Hence, KF treatment during the critical period of inflammation contributes to preventing chronic vulvar pain development.
Keywords: provoked vulvodynia, inflammation, mast cell, nerve growth factor, glutamate, hyperinnervation
Introduction
The International Society for the Study of Vulvar Diseases (ISSVD) defined vulvodynia as “vulvar pain of at least three months duration, without a clear identifiable cause, which may have potential associated factors“.1 The most common type of vulvodynia is “provoked vulvodynia” (PV), which affects 8% to 28% of women of all ages reproductive stages, and it is associated with severe sequelae on the quality of life.2–4 PV is a subset of vulvodynia that is provoked and localized to the vulvar vestibule. Women with PV report localized hypersensitivity and severe pain of the vulvar vestibule upon an attempt at vaginal penetration (vulvar mechanical allodynia; eg, intercourse, tampon use, light touching).5
The pathogenesis of vulvodynia is poorly understood, and the etiology is multifactorial.6 Previous studies have shown that hormonal imbalance promotes vulvar hyperinnervation and allodynia in an animal model;7,8 moreover, some evidence indicates that genetic factors contribute to neuronal sensitization among PV patients;9,10 central sensitization was also suggested to be involved in PV development;11 additionally, psychological distress and adverse childhood experiences including abuse, and social trauma (exclusion or lack of support) are also risk factors for PV.12
A substantial body of evidence, based on both animal and human studies, suggests that repeated vulvar inflammation is correlated with PV development. Interestingly, women with PV often report recurrent outbreaks of vulvovaginal candidiasis (RVVC).13 Conceivably, RVVC may be a primary trigger that causes inflammation in the vulva, resulting in stimulation of pain receptors, hyperinnervation, and impairment of nociceptors that lead to allodynia, persisting even after the inflammation itself subsides.6,14 Additionally, repeated vaginal yeast infection in laboratory animals was shown to cause hypersensitivity, allodynia, and increased pro-inflammatory mediators production in the vulvar vestibule.15
Findings concerning hypersensitivity and hyperinnervation in women with PV prompted a hypothesis that in response to signals from chemical mediators released during inflammation, the activity of the neuronal transduction proteins controlling the excitability of nociceptor terminals would lead to long-lasting neuronal modification.16,17 For example, the released glutamate from inflamed or damaged peripheral tissue activates mGluR5 (metabotropic glutamate receptor 5) in peripheral nerve terminals, leading to sensitization of nociceptors, including transient receptor potential cation channel vanilloid 1 (TRPV1), responsible for heat and burning sensations, and transient receptor potential ankyrin 1 (TRPA1), responsible for cold and mechanical sensation.17 Moreover, the released nerve growth factor (NGF) during inflammation from the immune cells or the peripheral tissue activates TRKA (Tropomyosin receptor kinase A) in peripheral nerve terminals, leading to hyperinnervation and nociceptors sensitization, including TRPA1 and TRPA1.18–21
Several findings have shown that hyperinnervation and mast cells (MCs) accumulation are the two most consistent features of vestibular biopsies from patients diagnosed with PV.14 This finding might explain the hypersensitivity and lower pain threshold among women with PV.22,23 MCs can discharge various mediators from their granules during inflammation, such as histamine, tryptase, chymase, and NGF, all involved in hyperinnervation and sensitization.24,25 In addition, the activation of MCs could establish a positive feedback loop that also contributes to maintaining pain in women with PV.26,27 Hyperinnervation and mast cells (MCs) accumulation in the vulva among PV has led to the hypothesis that early inflammatory events set the stage for long-term MC increase in the vulva, which in turn supports the overgrowth of the sensory neurons at the early stage of the inflammation, as well as the stability of the hyperinnervation of neurons after the inflammation subsides.14,26,28 However, whether mast cell-mediated inflammation can drive hyperinnervation and neuromodulation manifests in vulvar allodynia, and hyperalgesia remains unknown. Here, we investigated the long-term effects of inflammation on vulvar sensitivity, local inflammatory changes, and the role of mast cells in the development of chronic vulvar sensitivity.
Materials and Methods
Subjects
Female Sprague-Dawley rats (250–300g; 10 weeks age) were used in this study. All rat’s procedures were approved by the University Animal Care Committee, and were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. During the study, rats were housed in groups of 3–4 rats in a sterilized solid bottom cage with contact bedding under controlled temperature and a 12:12 h light/dark cycle. Rats were maintained on a standard pellet diet, and water was supplied ad libitum. All efforts were made to maintain rats suffering to a minimum.
A Rat Model of Vulvodynia
A rat model of vulvodynia was produced using a modified technique that had previously been described in mice by Farmer et al. Zymosan -a glycan derived from a yeast cell wall- was injected to achieve an inflammatory response. The injection was carried out under Isoflurane anesthesia. Each administered dose (10 mg/mL in 10μL of saline; 0.1 mg Zymosan, Sigma Z4250-1G) contained 300µL (1mL Syringe with 27Gxon needle) divided into 150µL into each side of the vulva to disperse the inflammatory factor throughout the vulva. Each subject rat was given three zymosan (or saline as control) injections in total, with one week period between each injection. After three rounds of zymosan, subject rats developed chronic vulvar pain that persisted long after the 3rd injection of zymosan.
Study Design
Rats were examined for baseline mechanical sensitivity of the vulva by an electronic Von Frey (VF) measuring device. Two and five days after each zymosan/saline administration (Round #1: day 2 and day 5; Round #2: day 9 and day 12; Round #3: day 16 and day 19), mechanical sensitivity of the vulva was measured. Following the 3rd round of zymosan/saline administration, mechanical sensitivity was measured on days 24 and 30 (Figure 1A). In addition, thermal sensitivity was tested by a dynamic hot plate (DHP) on day 32 (Figure 1A). In order to examine the long-term effects of zymosan administration, rats were tested every 20 days after the 3rd injection of zymosan\saline for a total period of 156 days (Figure 1B). Also, on day 172 (158 days after the last zymosan/saline administration), thermal sensitivity was tested by a dynamic hot plate (DHP) (Figure 1B).
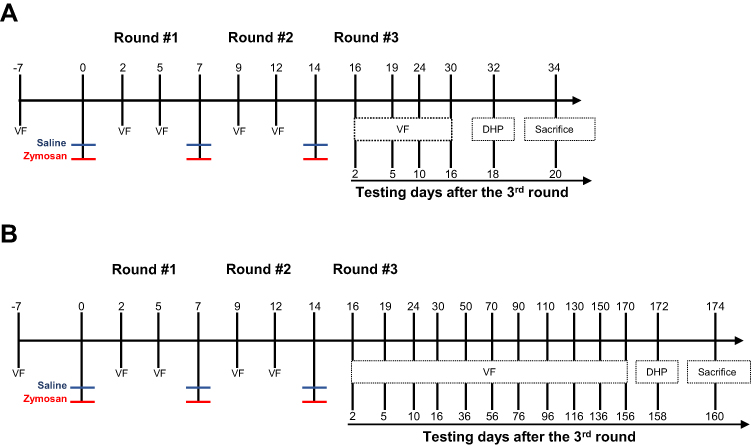 |
Figure 1 The experimental timeline illustrates the procedures across three rounds of vulvovaginal zymosan\saline administration. (A and B) Rats received zymosan\saline injections on days 0, 7, and 14; overall, three injections, 7 days between each one. Withdrawal responses of vulvar mechanical sensitivity were measured by electronic Von Frey (eVF) at baseline (−7 days) and +2 days and +5 days after zymosan\saline administration. eVF’s follow-up tests were taken at two points for the “short term“ effects of zymosan administration: day 24 (10 days after the 3rd round) and day 30 (16 days after the 3rd round). Thermal sensitivity was assessed using a Dynamic Hot plate test on day 32 (A). (B) for the ”long-term” effects of zymosan administration: eVF measurements were documented at nine points: day 24, day 30, day 50, day 70, day 90, day 110, day 130, day 150, day 170. Thermal allodynia was assessed using a Dynamic Hot plate test on day 172. |
Mast Cell Stabilizing
Mast cell stabilizer Ketotifen fumarate (KF; cat.0784, Tocris) was injected intraperitoneally (i.p.; 3mg/Kg). First, Ketotifen fumarate was dissolved in DMSO (1% of the final volume), and the stock solution was dissolved with physiological saline. Rats were injected with KF 45 minutes before each zymosan injection (300µL; 10 mg/mL in 10μL of saline; 0.1 mg) and every day to day 7 after the 3rd round, overall 21 days of treatment (Figure 8A).
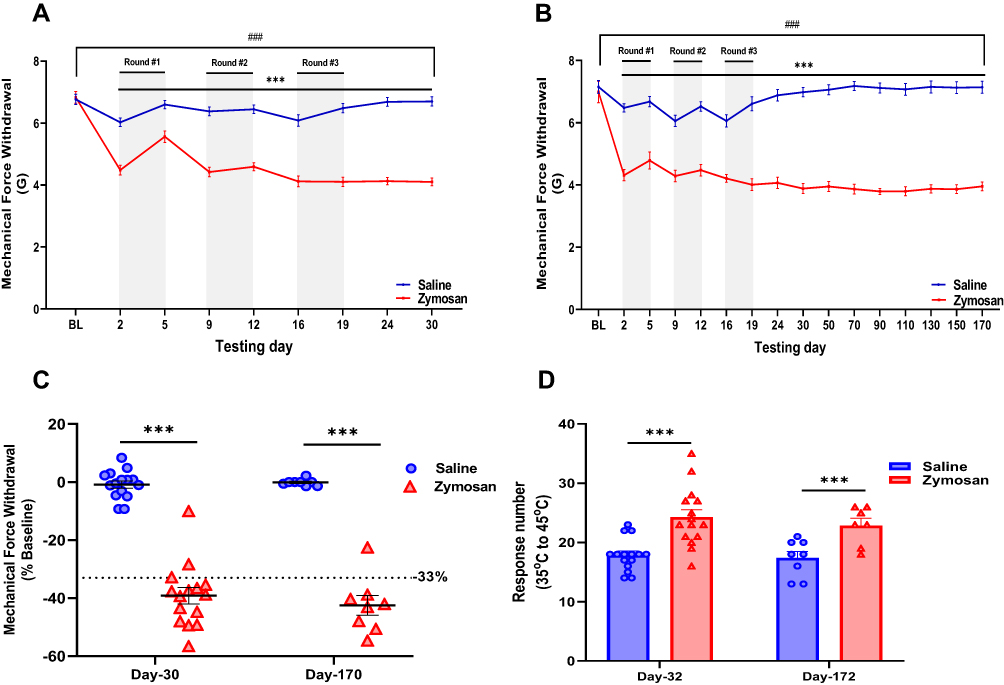 |
Figure 2 Development of vulvar mechanical and thermal sensitivity in rats after three rounds of zymosan administration. (A and B) Mechanical force withdrawal (G). There was a significant decrease in mechanical threshold among the saline group during each round. Afterward, the mechanical threshold returns to baseline ((A), short term n=15; (B), long term n=8). Among the zymosan group, the decrease in mechanical threshold was maintained throughout the zymosan challenges and 156 days after the 3rd round (day 170), compared to the baseline and saline group ((A), short term n=15; (B), long term n=8). (C) Mechanical force withdrawal (G) of day 30 and day 170 normalized to the baseline (D) Sum of nociceptive response (Vulva/hind paw licking, Rearing, and jumps) in the Dynamic Hot plate test (35°C to 45°C) after 18 days (day 32) and 158 days (day 172) of the 3rd round of zymosan/saline administration (n=8-15 per group). Mean ± SEM. ***P<0.001, zymosan vs saline, ###P<0.001, compared to Baseline. |
Mechanical (Von Frey) Sensitivity Testing
Mechanical sensitivity was assessed using an electronic Von Frey(eVF) device. For that purpose, rats were transferred to the testing room while in their cages for one hour for habituation. Thirty minutes before the measurement began, rats were placed in the measuring chambers (20 × 20 cm Plexiglas boxes equipped with a metallic screen-mesh floor) located 20 cm above the bench for acclimatization. An Electronic von Frey device (cat.38450 Ugo Basile, Varese, Italy) was used: the withdrawal threshold was evaluated by applying a force ranging from 0–30 g with a 0.2 g accuracy. The punctuate stimulus was delivered to the vulvar aspect of the rat vulva, and the withdrawal threshold was automatically displayed on the screen. Five values were collected by an observer blind to the experimental condition for each rat.
Thermal Sensitivity
Thermal sensitivity was assessed using a computer-controlled hot plate analgesia meter (cat.3515-022 Ugo Basile, Varese, Italy). The rat was placed in a Plexiglas cylinder (rats: 15.5-cm diameter, 25-cm height) with a drilled cover, where the plate can be heated up to 65°C. For the dynamic hot plate test, first, the rats were transferred to the measurement room while still in the home cage and stayed there for one hour as a habituation to the testing room. The rats were then placed on the plate for 15 minutes for habituation with a temperature of 35°C. For the dynamic hot plate (DHP), the plate temperature was increased up to 45°C with 1°C per min speed (r2=1). An observer blinded to the experimental condition documented hind paw\vulva lickings, escape behavior (jump), and rearing for each degree interval.
Immunofluorescent Staining and Microscopy
At the end of the experiment, rats were deeply anesthetized, sacrificed, and transcardially perfused with heparinized saline, 10% sucrose in buffered saline, and 4% buffered formaldehyde. The vaginal canal (from the external vulva to the cervix) was excised and fixated in 5% paraformaldehyde in phosphate buffer for one hour and then embedded in paraffin. Sections of 5μm thickness were cut from the vestibular area with a microtome, and every 10th section was collected for analysis. The sections were then mounted on Matsunami adhesive glass slide (TM-1190 TOMO, MATSUNAMI). In order to prepare the slides for histology and immunohistochemistry staining, Vestibular sections were deparaffinized and subjected to heat-induced epitope retrieval using OmniPrep (pH 9.0 (10X), Cat. No.: ZUC067-100, ZYTOMED SYSTEMS). According to the manufacturer’s instructions, briefly, the Coplin jar containing the slides in pre-warmed solution was placed in a water bath set to 85°C for 20 minutes. Followed by 3x wash with DDW at 85°C and rinsed with wash buffer (Zytomed systems) using a squirt bottle. Afterward, to remove background staining, we used Background Buster (#NB306-50, INNOVEX), followed by three cycles of wash buffer for two minutes each, then incubated for one hour at room temperature in a humidity chamber with one of the first antibodies: anti-rat TRPV1 channel ATTO-488 (1:240, cat.ACC-029-AG, Alomone labs, Jerusalem), anti-rat TRPA1 channel (1:240, cat.ACC-037, Alomone labs, Jerusalem), anti-rat protein gene product 9.5 (PGP 9.5, 1:500, NB300-676, Novus Biological), rabbit anti-Chymase (1:250, NBP2-257441, Novus Biological) and mouse anti-Tryptase (1:250, NBP2-26444, Novus Biological). After the first immunoreaction, the slides were washed in 1xPBS and incubated with the secondary antibody for one hour at room temperature in a humidity chamber. However, When the antibodies were primary antibodies, after the second immunoreaction, the slides were washed in 1xPBS and incubated for 1 hour with a secondary antibody: donkey anti-mouse IgG dylight® 594 (1:500, #A90-337D4, Bethyl Laboratories, Inc.), Goat anti-rabbit IgG dylight® 488 (1:500, #A120-201D2, Bethyl Laboratories, Inc.), F(ab`)2 Donkey x-Rabbit IgG dylight® 594 IgG® (1:500, #A120-216D4, Bethyl Laboratories, Inc.), Goat anti-rabbit IgG dylight® 488 (1:500, #A120-201D2, Bethyl Laboratories, Inc.), followed by three cycles of wash buffer for two minutes each. The sections were mounted with “Flouromount G with DAPI” (eBiosciences) and incubated for 30 seconds. Microscopic observation was done using the Eclipse Ci microscope (Nikon Corp., Japan). The images of 10 fields were captured by the Nikon DS-Ri1 camera (Nikon Corp., Japan) with the same microscope settings and exposure time. The images were analyzed using NIS Elements analysis software. Mast cell density was determined by the number of positively stained cells with chymase and tryptase, TRPV1 and TRPA1 channels density were determined by fluorescent pixel intensity measurements, and Fiber length was calculated with functions in the NIS Elements program configured to measure total fiber length per unit area. The images were analyzed by an observer blinded to the condition.
Assessment of Morphology After Zymosan/Saline Administration
Sections of 5μm thickness were cut from the vestibular area with a microtome onto IHC adhesive glass slide (TM-1190 TOMO, MATSUNAMI) and stained with H&E to identify gross vulvar morphology changes among saline groups and zymosan groups. Sections were also examined for inflammatory infiltrate and edema in the epithelium. Microscopic observation using the Eclipse Ci microscope (Nikon Corp., Japan), the images of 10 fields were captured by the Nikon DS-Ri1 camera (Nikon Corp., Japan), and the images were analyzed by a pathologist blinded to the condition. Morphological scores of inflammation in H&E-stained: Whole skin-absence of inflammation (score=0), Discrete–presence of few inflammatory cells (score=1), Moderate–many inflammatory cells (score=2), Severe–exaggerated inflammatory cellularity (score=3).
Biochemistry Measurements
Glutamate levels were determined in rat vulva extracts using the Glutamate Assay Kit (MAK004, Sigma-Aldrich). Briefly, the rats were sacrificed after 6, 24, 48 hours, and 30 days of zymosan (300µL; 10 mg/mL) or saline injection (300ul) in each round. Samples of vestibular extracts were homogenized and diluted with a Standard diluent buffer of Glutamate kit (1:100). NGF levels were determined according to the manufacturer’s instructions in rat vestibule extracts using the rat NGF Sandwich ELISA kit (BE69198, IBL America). Briefly, the rats were sacrificed after 24 hours and 30 days of zymosan (300µL; 10 mg/mL) or saline (300ul) injection in each round. The vestibular was excised. Samples of vestibular extracts were homogenized and diluted with Standard diluent buffer of NGF ELISA kit (1:100) and tested according to the manufacturer’s instructions.
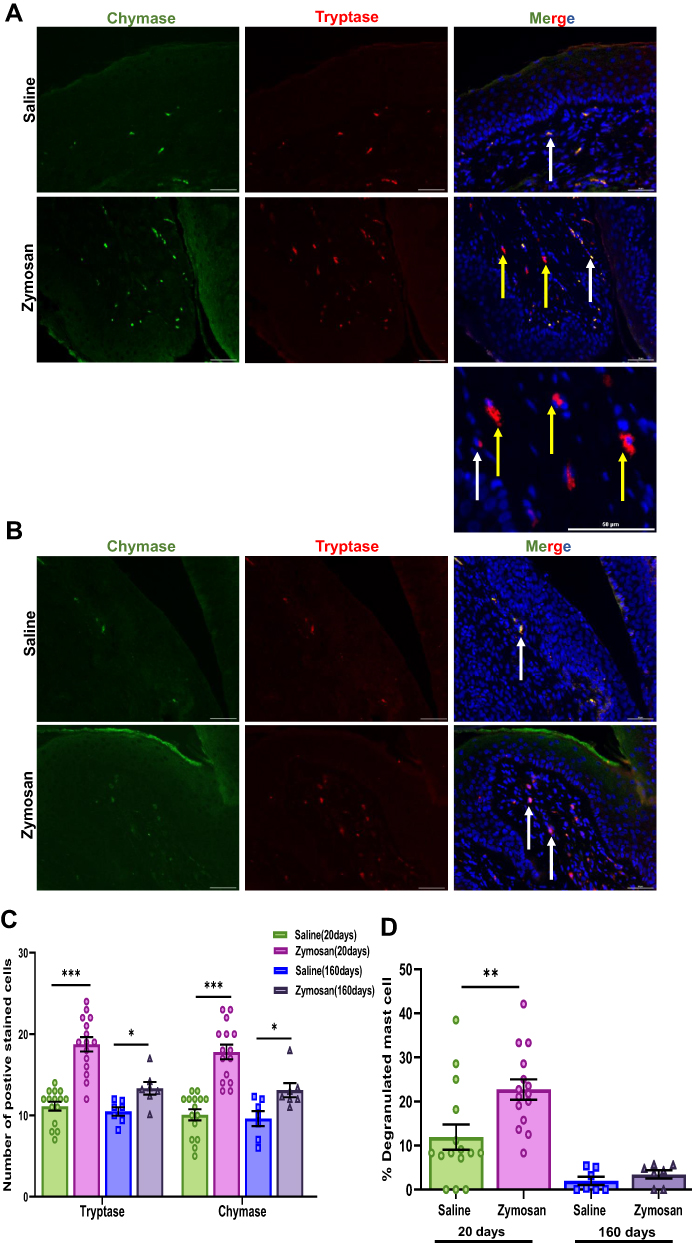 |
Figure 3 Short and long-term effects of zymosan challenges in the vulva on mast cell. (A) Positively stained cells with mast cell chymase and tryptase in the zymosan and the saline group after 20 days of the 3rd round. Mast cell chymase (green; White arrows) and tryptase (red; white arrows), diffuse tryptase (Yellow arrows) merged with dapi stain (blue) scale bar: 50 µm. (B) Positively stained cells with mast cell chymase and tryptase in the zymosan and the saline group after 160 days of the 3rd round. Mast cell chymase (green; White arrows) and tryptase (red; white arrows) merged with dapi stain (blue) scale bar: 50 µm. (C) The number of positively chymase or tryptase stained cells in zymosan and saline groups after 20 and 160 days of the 3rd round (n=7-15 per group). (D) The percentage of degranulated mast cells among zymosan and saline groups after 20 and 160 days of the 3rd round (n=7-15 per group). Mean ± SEM. *P<0.05, **P<0.005, ***P<0.001. |
Data Analysis
Statistical analyses were performed using IBM SPSS statistics version 25 and GraphPad Prism 9. All data were expressed as Mean ± SEM. Differences between groups were assessed by Student’s t-test, one-way ANOVA, two-way ANOVA, and mixed-model repeated-measures analysis of variance (GLM). Differences were determined Post hoc by Tukey’s test and Student’s t-test when significant main effects or interactions were detected. Non-parametric X2 analysis was used to test Mechanical vulva thresholds (decrease ≥33%, compared with baseline). The accepted significance value for all tests was set at P<0.05.
Results
Repeated Zymosan Administration Causes Mechanical and Thermal Allodynia
In each round, two days after administration, both the saline and the zymosan groups displayed a significant decrease in mechanical sensitivity threshold. The decrease in the mechanical threshold of the saline group disappeared after ten days of the 3rd round (Figure 2A and B). However, the mechanical threshold decrease in the zymosan group was maintained throughout 156 days after the 3rd round compared to baseline (Figure 2A and B). After 16 days of 3rd round of zymosan challenge (Figure 1A), persistent allodynia was present in the short-term group (X2=8.067, P=0.005; Figure 2C). More specifically, 87% of the rats had a 33% reduction in the mechanical sensitivity threshold. In the long-term groups, 156 days after the zymosan challenge, 86% of the rats had a 33% reduction in the mechanical sensitivity threshold (X2=4.50, P=0.034; Figure 2C). Furthermore, the zymosan groups showed a higher thermal sensitivity response than the saline groups after 18 and 158 days of the zymosan challenge. The Zymosan group also displayed more pain-related behaviors like escape behavior, rearing, and vulva licking were more observed among the zymosan groups (Figure 2D).
 |
Figure 4 Morphological and inflammatory effects of zymosan challenges in the vulva. (A) Visual inspection of hematoxylin and eosin (H&E)–stained sections, 20 and 160 days of zymosan/saline administration in the vulva. Saline administration in the vulva does not have morphological or inflammatory effects. However, evidence of an active inflammatory process, marked dermal lymphocytic infiltrate with exocytosis, increase in the number and the activation of mast cells (See Figure 3) after 20 days of the 3rd round of zymosan administration in the vulva. However, after 160 days of the 3rd round of zymosan administration, there was no evidence of inflammation. (B) Inflammation score of the saline and the zymosan groups after 20 and 160 of 3rd round of zymosan/saline administration (n=5 per group). Mean ± SEM. ***P<0.001. |
Three Zymosan Challenges to the Vulva Induce Local Accumulation of MC
We found a significant increase in MC number by double staining of tryptase and chymase proteases in the zymosan group compared to the saline group, 20 days after the 3rd zymosan/saline administration round (t(28)=−7.25, P<0.001; t(28)=−6.82, P<0.001, Tryptase, Chymase, respectively; n=15 per group; Figure 3A and C). In addition, we also observed an increase in the number of diffused tryptase staining around the MCs in the zymosan group. The diffuse staining around the MCs suggests that they were degranulated (t(28)=−2.94, P=0.006; Figure 3A). Thus, after repeated zymosan administrations, we found an almost 200% increase in MC number in the zymosan group compared to the saline group, and almost 25% of those MCs were degranulated (Figure 3C and D).
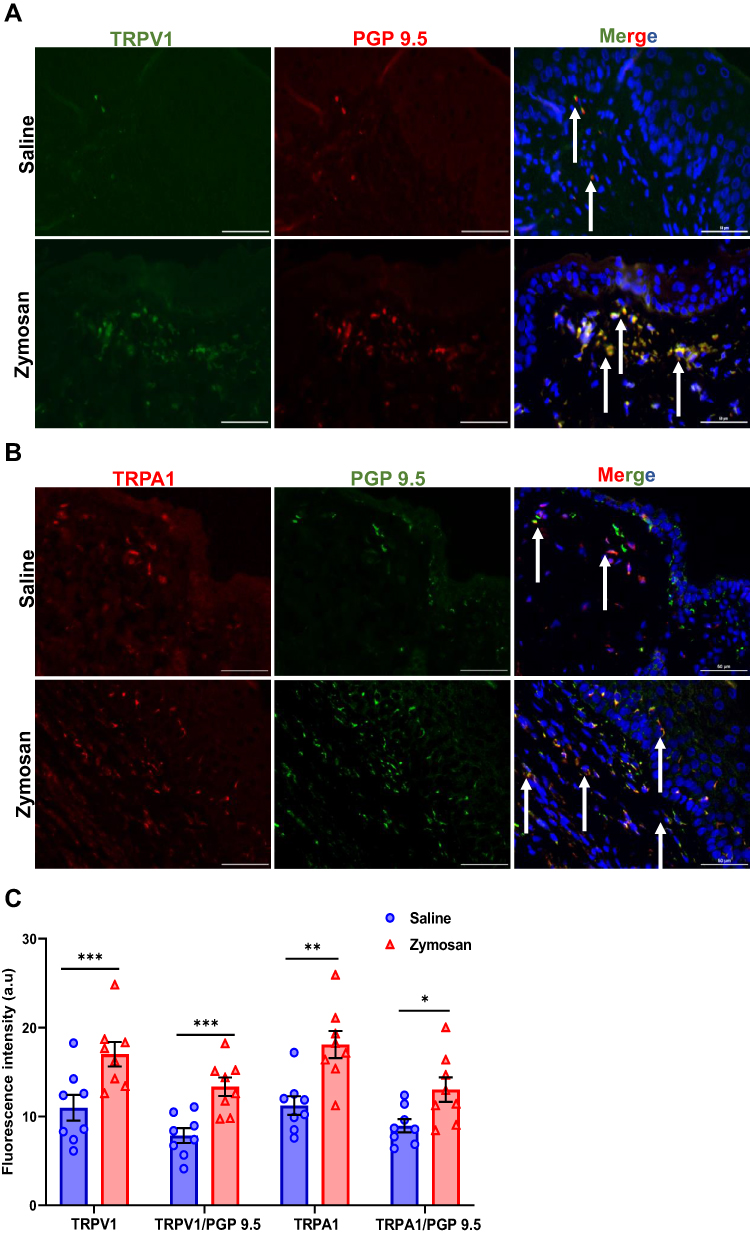 |
Figure 5 TRPV1 and TRPA1 expression in vulvar nerves after multiple rounds of zymosan challenge. (A) The expression of TRPV1 channel in vulva nerves, 20 days after the 3rd zymosan/saline administration. Coexpression of TRPV1 channel (green) and neuronal PGP 9.5 (red; White arrows) merged with dapi stain (blue). (B) The expression of TRPA1 channel in vulva nerves, 20 days after the 3rd round zymosan\saline administration. Coexpression of TRPA1 channel (red) and neuronal PGP 9.5 (green; White arrows) merged with dapi stain (blue). Scale bar: 50 µm. (C) Fluorescence intensity (arbitrary units) of TRPV1, TRPA1, TRPV1/PGP-9.5, and TRPA1/PGP-9.5 in the zymosan and the saline groups 20 days of the 3rd round (n=8 per group). Mean ± SEM. *P<0.05, **P<0.005, ***P<0.001. |
In order to examine the long-term effect of inflammation on the presence of MC, Immunofluorescence analyses were conducted on post-mortem rats’ vulvar tissue 160 days after the 3rd zymosan/saline administration round. Among the zymosan group, there was an increase in the presence of MC compared to the saline group (t(12)=−3.03, P=0.01; t(12)=−2.76, P=0.01, Tryptase, Chymase, respectively, n=7 per group, Figure 3B and C). However, MCs number in the zymosan group after 160 days was less than zymosan group that was tested at 20 days after the 3rd round of zymosan administration (t(20)=3.80, P<0.001; t(20)=3.23, P=0.004, Tryptase, Chymase, respectively). Moreover, there was no significant difference in the number of degranulated MCs between the saline and the zymosan groups after 160 days of the 3rd round (t(12)=−1.09, P=0.29, n=7 per group, Figure 3D).
The Effect of Zymosan Challenges on Histological Changes
Visual inspection of hematoxylin and eosin (H&E) stained vulvar sections obtained after 20 days and 160 days of the 3rd round of zymosan/saline administration revealed marked dermal lymphocytic infiltrate with exocytosis in the zymosan group compared to the saline group after 20 days of the 3rd round (P<0.05; n=5 per group; Figure 4A and B). However, these inflammatory effects of zymosan administration were not detectable after 160 days of the 3rd round. Thus, there were no detectable intergroup differences in the inflammatory infiltrate between the saline and the zymosan group (P=0.56; n=5 per group; Figure 4A and B).
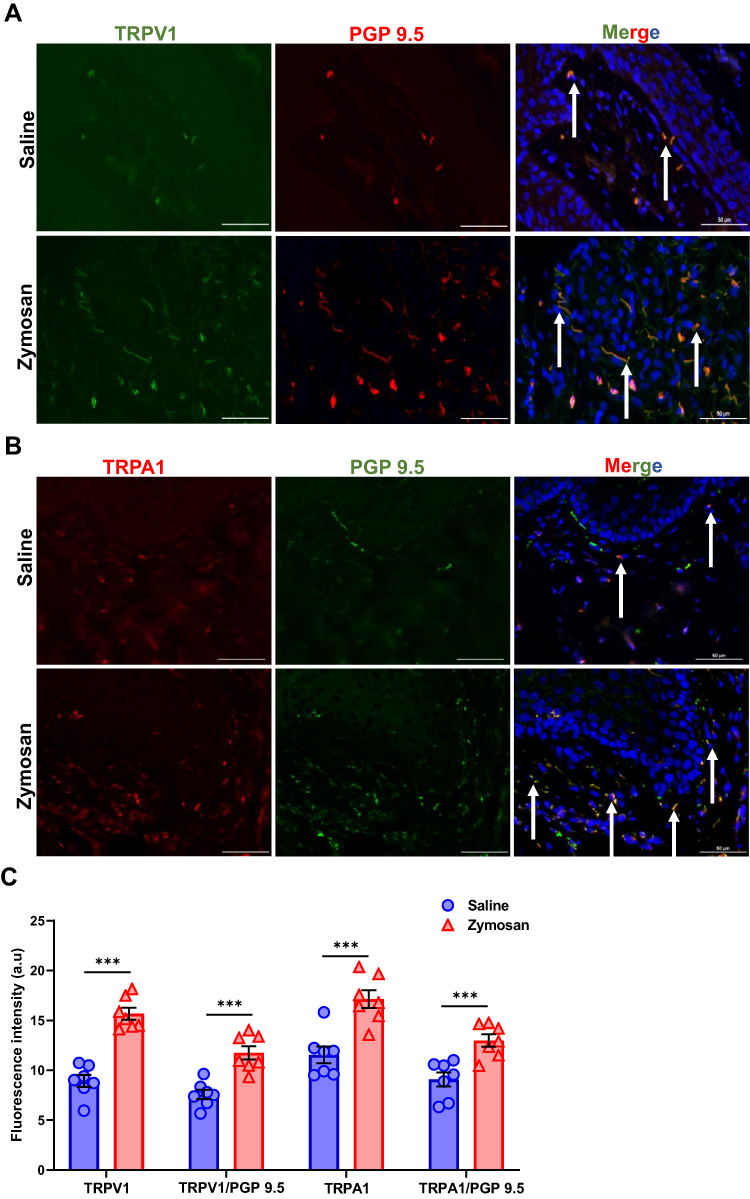 |
Figure 6 TRPV1 and TRPA1 expression in vulvar nerves after 160 days of the 3rd round of zymosan/saline administration. (A) The expression of TRPV1 channel in vulva nerves, 20 days after the 3rd zymosan/saline administration. Coexpression of TRPV1 channel (green) and neuronal PGP 9.5 (red; White arrows) merged with dapi stain (blue). (B) The expression of TRPA1 channel in vulva nerves, 20 days after the 3rd round zymosan\saline administration. Coexpression of TRPA1 channel (red) and neuronal PGP 9.5 (green; White arrows) merged with dapi stain (blue). Scale bar: 50 µm. (C) Fluorescence intensity (arbitrary units) of TRPV1, TRPA1, TRPV1/PGP-9.5, and TRPA1/PGP-9.5 in the zymosan and the saline groups 160 days of the 3rd round (n=8 per group). Mean ± SEM. ***P<0.001. |
Neuromodulation and Hyperinnervation of Vulva Nerves After Multiple Rounds of Zymosan Challenge
There was a significant increase of TRPV1 and TRPA1 expression in vulva neurons [as detected by immunoreactivity (IR) for the pan-axonal marker protein gene product 9.5 (PGP 9.5)] in the zymosan group compared to the saline group 20 days after the 3rd round (t(14)= −4.14, P<0.001; t(14)=−2.60, P=0.021, TRPV1, TRPA1, respectively; n=8 per group; Figure 5A–C). In addition, after 160 days from the 3rd round, we found a high expression of TRPV1 and TRPA1 in the zymosan group compared to the saline group (t(12)=−5.08, P<0.001; t(12)=−4.12, P<0.001, TRPV1, TRPA1, respectively; n=7 per group; Figure 6A–C).
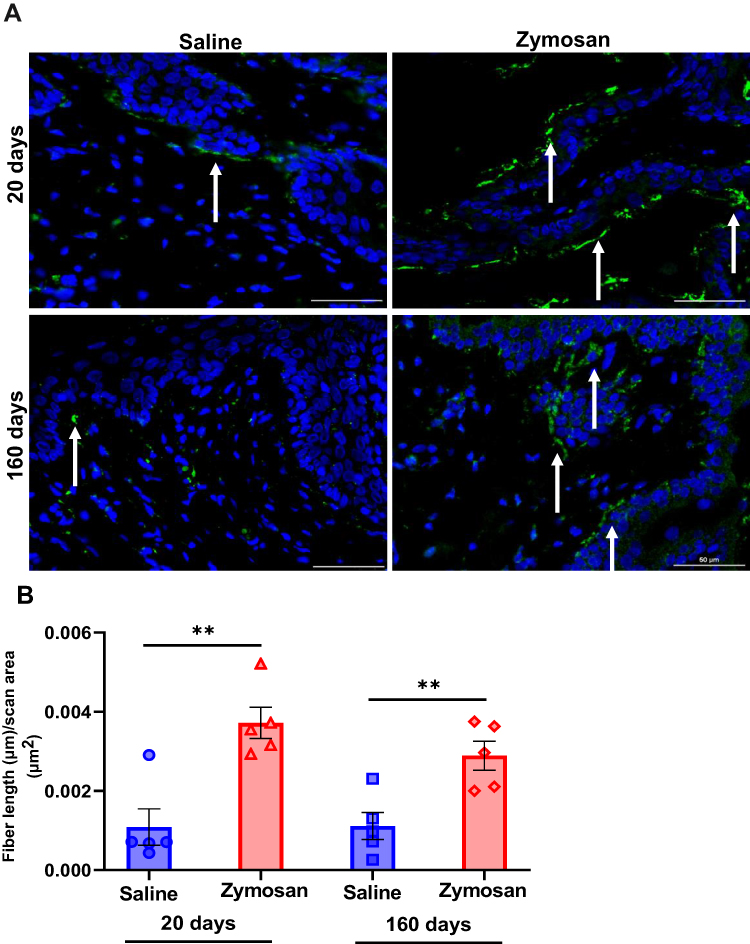 |
Figure 7 Hyperinnervation of vulvar nerves after multiple rounds of zymosan administration. (A) Nerve fibers [as detected by immunoreactivity (IR) for the pan-axonal marker protein gene product 9.5 (PGP 9.5; green)] merged with dapi stain (blue). Scale bar: 50 µm. (B) Fiber length (µm) per unit area (µm2) in the zymosan and saline groups after 20 and 160 days of the 3rd round of zymosan/saline administration (n=5 per group). Mean ± SEM. **P<0.005. |
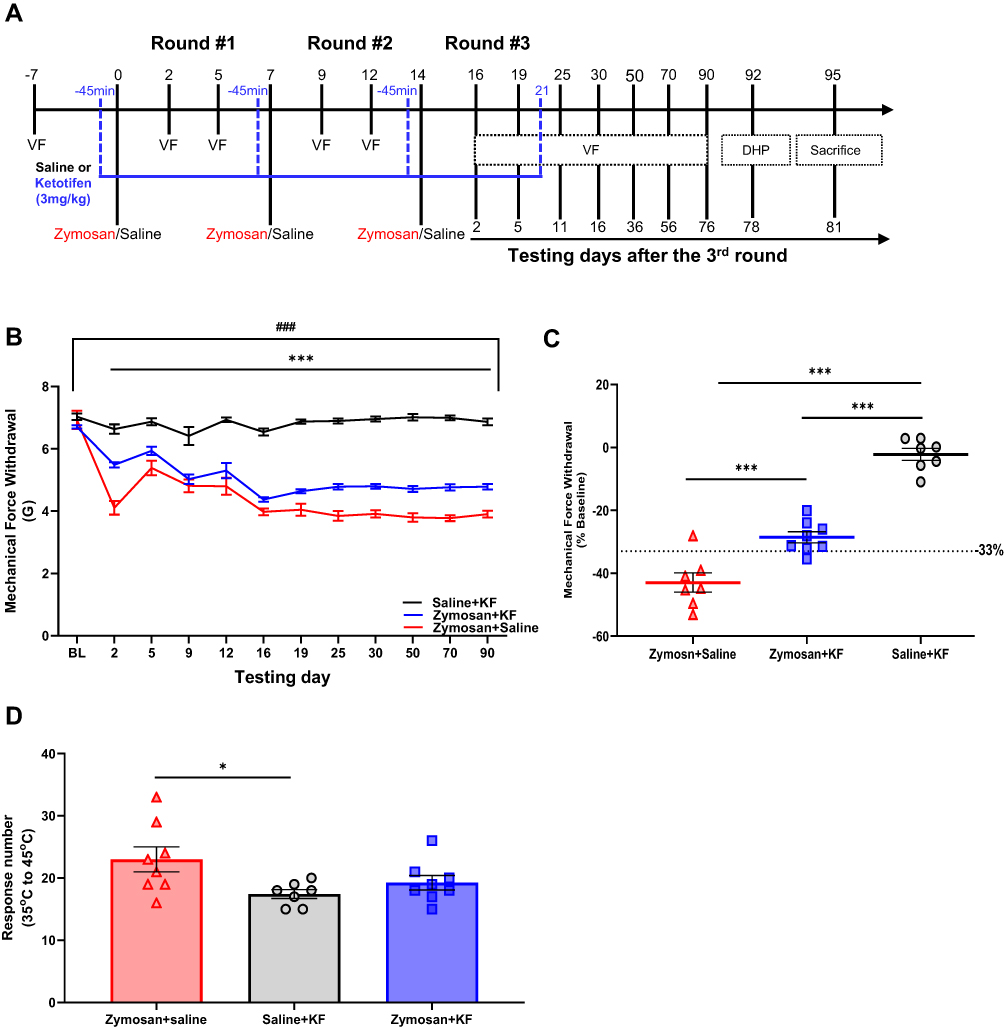 |
Figure 8 Treatment with Ketotifen fumarate during inflammation suppressed the development of vulvar hypersensitivity. (A) The experimental timeline illustrates the experimental procedures across three rounds of vulvar inflammation. Vulvar rat injected with zymosan or saline (n=7-8 per group) on days 0, 7, and 14; overall, three injections, 7 days between each one. The groups were injected with ketotifen (i.p; 3mg/Kg) or saline (i.p; 300ul) 45 minutes before the zymosan/saline administration into the vulva, and every day to day 7 after the 3rd round, overall 21 days of treatment. Vulvar mechanical sensitivity was measured by Von Frey (VF). Thermal sensitivity was assessed using a Dynamic Hot plate test on day 92 after the 3rd round. (B) Mechanical force withdrawal (G) of the zymosan-saline, the zymosan-ketotifen, and the saline-ketotifen group. There was no significant change in mechanical threshold among the saline- ketotifen group. There was a significant decrease in mechanical threshold in zymosan-saline and zymosan-ketotifen groups during the three rounds compared to the baseline. The decrease in mechanical threshold was maintained 76 days after the 3rd round in both groups. Also, there was a significant difference in mechanical force withdrawal between the two zymosan groups and the saline-ketotifen group at each time point (Excluding baseline). (C) Mechanical force withdrawal test of day 90 normalized to the baseline (n=7-8 per group). (D) Sum of nociceptive response in the Dynamic Hot plate test (35°C to 45°C) in zymosan-saline, zymosan-ketotifen, and saline-ketotifen group after 78 of the 3rd round of zymosan/saline administration (n=8 per group). Mean ± SEM. *P<0.05, ***P<0.001, difference between groups, ###P<0.001, Compared to the baseline. |
Moreover, we observed an increase in the density of nerve fibers [as detected by immunoreactivity (IR) for the pan-axonal marker protein gene product 9.5 (PGP 9.5)] after 20 days of the 3rd round of zymosan/saline administration in the zymosan group compared to the saline group (t(8)= −4.33, P=0.003; Figure 7A and B). The increase in the density of the nerve fibers is reflected in the number of fibers and their length. In addition, among the zymosan group, nerve fibers were located near the epithelial surface (Figure 7A). Unsurprisingly, 160 days after the 3rd round, we observed an increase in the density of nerve fibers in the zymosan group compared to the saline group (t(8)=−3.53, P=0.008; Figure 7A and B). The increased density was reflected in the number of fibers and their length. Hence, the nerve fibers almost reached the epithelial surfaces in the zymosan group (Figure 7A).
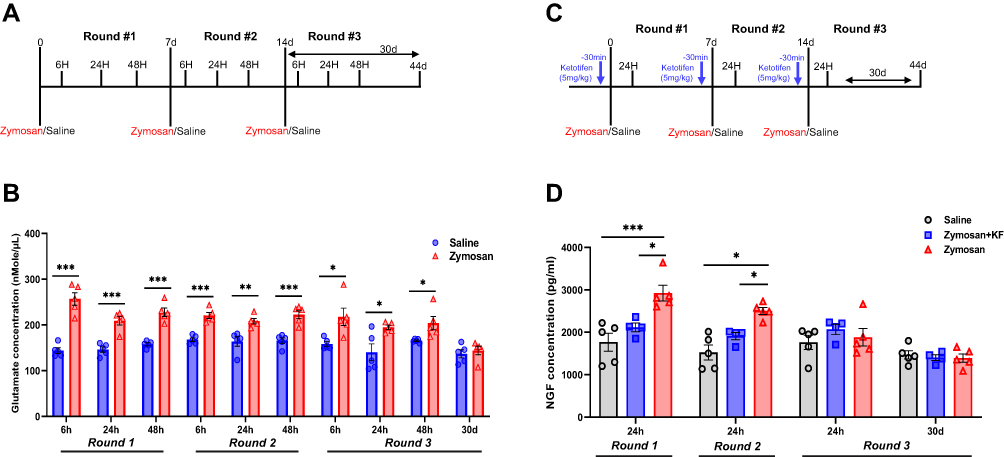 |
Figure 9 Glutamate and NGF concentration in vulva tissue after zymosan or saline administration. (A) Rats received zymosan\saline injection (in the vulva) on days 0, 7, and 14; overall, three injections, 7 days between each one. Rats were sacrificed after 6, 24, and 48 hours of each injection and after 30 days of the 3rd injection of zymosan/saline. (B) Glutamate concentration in the vulva tissue of the saline and the zymosan group (n=5 per time point). (C) Rats were injected with ketotifen (i.p; 3mg/kg) 45 minutes before each zymosan/saline administration (in the vulva); overall, three injections of zymosan/saline, 7 days between each one. Rats were sacrificed after 24 hours of each injection and after 30 days of the 3rd injection of zymosan/saline. (D) NGF concentration in the vulva tissue of the saline, zymosan-saline, and zymosan-ketotifen group (n=4-5 per group at each time point). Mean ± SEM. *P <0.05, **P<0.005, ***P<0.001. |
The Treatment with MC Stabilizer During Inflammation Suppressed the Development of Vulvar Hypersensitivity
To examine the role of MC in the development of vulvar mechanical and thermal sensitivity, we used MCs stabilizer Ketotifen fumarate (KF). Rats were injected 45 minutes with ketotifen (i.p; 3mg/k) before zymosan/saline administration and every day to day 7 after the 3rd round (overall 21 days of treatment; Figure 8A). There was no significant decrease in mechanical sensitivity threshold in the saline group injected with Ketotifen fumarate (Saline-KF; P=0.075). However, among both zymosan groups: the zymosan group that was treated with Ketotifen fumarate (Zymosan-KF) and the zymosan group that was treated with saline (Zymosan-saline), there was a significant decrease in mechanical sensitivity threshold during each round, which persisted even 76 days after the 3rd round compared to baseline (Figure 8B). Furthermore, there was a significant difference in mechanical threshold sensitivity between the zymosan-KF group and the zymosan-saline group at each time point (Excluding Baseline, 5 days after the 1st round, 2 and 5 days after the 2nd round). More specifically, the zymosan-saline group’s mechanical threshold decrease was higher than the zymosan-KF group. Also, there was a significant difference in mechanical threshold sensitivity at each time point between the saline group and both of the zymosan groups: zymosan-KF and the zymosan-saline group (Figure 8B). In addition, persistent allodynia was present in the zymosan-saline group (X2=3.57, P=0.033), given that 85% of the rats had a 33% reduction in the mechanical sensitivity threshold, while only 25% percent of the zymosan-KF treated rats had a 33% reduction in the mechanical sensitivity threshold (X2=2.0, P=0.157; Figure 8C). Furthermore, the zymosan-saline group showed a higher thermal sensitivity response than the saline-KF group. However, there was no significant difference in thermal sensitivity between the zymosan-saline KF and the two other groups: the saline-KF and the zymosan-saline group (Figure 8D).
Glutamate Concentration After Zymosan Administration in Vulva Tissue
Glutamate concentration in the vulva was significantly higher in the zymosan group compared to the saline group after 6 hours, 24 hours, and 48 hours in the three rounds (P<0.05; n=5 per group; Figure 9A and B). The increase in glutamate concentration was maintained 24–48 hours after the zymosan challenge. However, there was no significant difference in glutamate concentration in the vulva 30 days after the 3rd round of zymosan\saline administration between the saline and the zymosan group (P=0.56; Figure 9B).
Stabilization of MCs During Inflammation Reduces Nerve Growth Factor Concentration and MCs Accumulation in the Vulva
Rats were injected with ketotifen fumarate (i.p; 3mg/k) 45 minutes before zymosan or saline administration in the vulva. There were three zymosan/saline administration rounds with 7 days between each injection (Figure 9B). After 24 hours of the 1st and the 2nd round of zymosan or saline injection, the nerve growth factor (NGF) concentration was significantly higher in the zymosan group compared to the saline and the zymosan group that was treated with ketotifen fumarate (P<0.05; n=4-5 per group; Figure 9B), while no such difference was observed after 24 hours and 30 days of the 3rd round between the three groups (Figure 9B).
In order to examine the effect of MCs stabilizer ketotifen fumarate on the presence of MCs, Immunofluorescence analyses were conducted on post-mortem rats’ vulvar tissue 81 days after the 3rd zymosan/saline administration round (Figure 8A). There was a significant difference in MCs number between the three groups; saline-KF, zymosan-KF, and zymosan-saline (F(2,18)=10.22, P<0.001; F(2,18)=5.18, P=0.017, Tryptase, Chymase, respectively, n=7 per group, Figure 10A and B). Post-hoc analyses show that the number of MCs in the vulva tissue was higher in the zymosan group that was treated with saline (zymosan-saline) during the three rounds of zymosan/saline administration in the vulva, compared to both ketotifen fumarate treatment groups: the saline-KF and the zymosan-KF group (P<0.05; Tryptase, Chymase). There was no significant difference in the number of MCs between the zymosan-KF and the saline-KF groups (P=0.48; P=0.83, Tryptase, Chymase, respectively; Figure 10A and B). Also, there was no significant difference in the number of MCs degranulated between groups (F(2,18)=1.33, P=0.282; n=7 per group; Figure 10C).
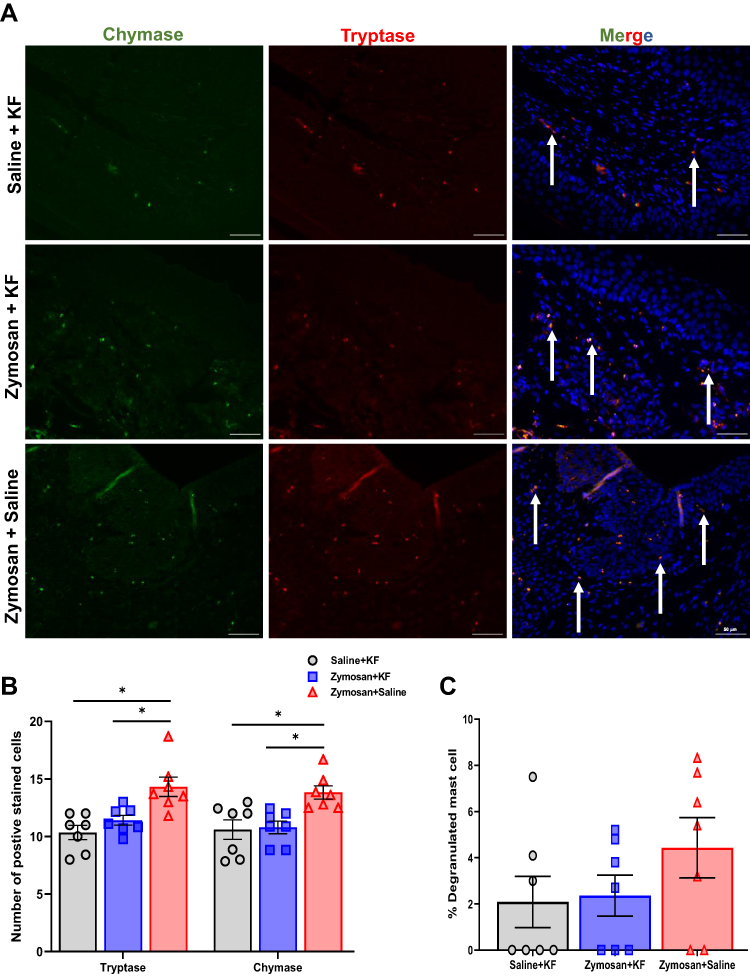 |
Figure 10 Long-term effects of mast cells stabilizer KF on the presence of mast cell, 81 days after the 3rd round. (A) Positively stained cells with mast cell chymase (green; white arrows) and tryptase (red; white arrows) merged with dapi stain (blue) in the Saline- ketotifen, the zymosan- ketotifen, and the zymosan-saline group. Scale bar: 50 µm. (B) The number of positively stained cells with chymase or tryptase in the Saline- ketotifen, the zymosan- ketotifen, and the zymosan-saline group after 81 days of the 3rd round (n=7 per group). (C) The percentage of degranulated mast cells among the Saline- ketotifen, the zymosan- ketotifen, and the zymosan-saline group after 81 days of the 3rd round (n=7 per group). Mean ± SEM. *P<0.05. |
Stabilization of MCs During Inflammation Modulates the Neuromodulation and Hyperinnervation of Vulva Nerves
There was a significant difference in TRPV1 and TRPA1 expression in vulva neurons [as detected by immunoreactivity (IR) for the pan-axonal marker protein gene product 9.5 (PGP 9.5)] between the three groups: the zymosan-saline, the saline-KF, and the zymosan-KF group. 81 day after the 3rd round (F(2,18)=6.18, P=0.009; F(2,18)=9.82, P<0.001, TRPV1, TRPA1, respectively; n=7 per group; Figure 11A–C). Post-hoc analyses showed that the expression of TRPV1 in vulva neurons was higher in the zymosan group that was treated with saline (zymosan-saline) during the three rounds of zymosan/saline administration in the vulva, compared to both ketotifen fumarate treatment groups: the saline-KF and the zymosan-KF group (P<0.05). There was no significant difference in the expression of the TRPV1 channel between the saline-KF group and the zymosan-KF group (P=0.80). Post-hoc analyses showed that the expression of TRPA1 in vulva neurons was higher in both zymosan groups (the zymosan-saline and zymosan-KF groups) compared to the saline-KF group (P<0.05; Figure 11C), and there was no significant difference in the expression of TRPA1 in vulva neurons between the zymosan-saline and the zymosan-KF group (P=0.18; Figure 11C). However, the zymosan-KF group tended, on average, to show a slight decrease in the expression of TRPA1 in vulva neurons compared to the zymosan-saline group.
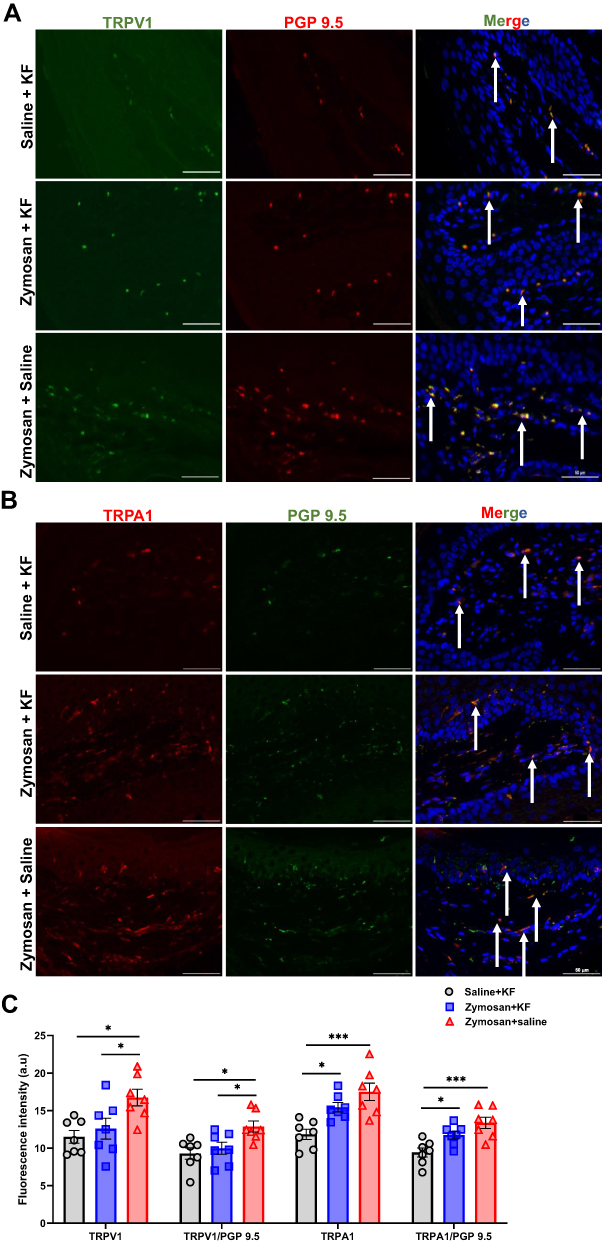 |
Figure 11 Long-term effects of mast cells stabilizer KF on TRPV1 and TRPA1 expression in vulvar nerves after 81 days of the 3rd round of zymosan/saline administration. (A) The expression of TRPV1 channel in vulva nerves, 81 days after the 3rd zymosan/saline administration. Coexpression of TRPV1 channel (green) and neuronal PGP 9.5 (red; white arrows) merged with dapi stain (blue). (B) The expression of TRPA1 channel in vulva nerves, 81 days after the 3rd round zymosan\saline administration. Coexpression of TRPA1 channel (red) and neuronal PGP 9.5 (green; white arrows) merged with dapi stain (blue). Scale bar: 50 µm. (C) Fluorescence intensity (arbitrary units) of TRPV1, TRPA1, TRPV1/PGP-9.5, and TRPA1/PGP-9.5 in the Saline- ketotifen, the zymosan- ketotifen, and the zymosan-saline group 81 of the 3rd round (n=7 per group). Mean ± SEM. *P<0.05, ***P<0.001. |
Furthermore, we found a significant increase in the density of nerve fibers in the vulva [as detected by immunoreactivity (IR) for the pan-axonal marker protein gene product 9.5 (PGP 9.5)] after 81 days of the 3rd round of zymosan/saline administration, in the zymosan-saline group compared to the zymosan-KF and the saline-KF group (F(2,12)=11.61, P=0.002; Figure 12A and B) with no significant difference between the saline-KF and the zymosan-KF group. In the zymosan-saline group, the increase in nerve fibers density is reflected in the number of fibers and their length (Figure 12A).
 |
Figure 12 Long-term effects of mast cells stabilizer KF on vulvar hyperinnervation. (A) Nerve fibers [as detected by immunoreactivity (IR) for the pan-axonal marker protein gene product 9.5 (PGP 9.5; green)] merged with dapi stain (blue). Scale bar: 50 µm. (B) Fiber length (µm) per unit area (µm2) in the saline- ketotifen, the zymosan-Ketotifen, and the zymosan-saline group (n=5 per group). Mean ± SEM. *P<0.05, ***P<0.001. |
Discussion
The intricacy of the link between the MCs and the nervous system during inflammation in the development of chronic pain conditions such as PV has been addressed before.29–32 However, the current study established several steps in the development of persistent PV in a rat model. First, it should be acknowledged that the vulvar pain model that was implemented here successfully induced long-lasting mechanical and thermal vulvar hypersensitivity after three rounds of zymosan challenge for an experimental period of approximately six months. Undoubtedly the long-lasting behavioral changes underscore the usefulness of the model.
During the three zymosan/saline injections, both the saline and zymosan groups displayed a significant decrease in the mechanical sensitivity threshold. There were no significant long-term effects of saline injection in the vulva, given that the decrease in the mechanical threshold of the saline groups disappeared after 10 days of the 3rd round. On the other hand, in the zymosan-injected groups, the mechanical sensitivity initially recovered to nonallodynic levels (defined as >66% of baseline threshold) after the 1st and 2nd zymosan challenges but never recovered to the baseline after the 3rd. In addition, after the 3rd challenge, the zymosan group showed a low mechanical threshold that was maintained throughout, even after 156 days, compared to the baseline. Moreover, we evaluated vulvar hypersensitivity using a dynamic hot plate test to evaluate hypersensitivity to heat, as observed in women with vulvodynia.33 Indeed, we observed a significant level of thermal hypersensitivity, including vulvar thermal hypersensitivity, after 18 and 158 days of the 3rd round of administration in the zymosan group compared with the saline group. The zymosan groups showed significant nociceptive responses such as vulva licking, rearing, and jumps compared to the saline groups.
As expected, we observed vulvar allodynia during and immediately after each round of the zymosan challenge. Zymosan is prepared from the yeast cell wall and consists of protein-carbohydrate complexes, and its administration induces inflammation.34 Thus, the onset of vulvar hypersensitivity during the zymosan challenge may have resulted from acute zymosan-induced inflammation. Interestingly, this result is compatible with a previous animal study15 and clinical reports of vulvovaginal pain during candida infection and inflammation.35 Notable, the inflammatory response to zymosan in the rat model was associated with marked dermal lymphocytic infiltrate with exocytosis, accompanied by a significant increase (almost 200%) in the number of MCs.
MCs are multifunctional innate immune cells implicated in inflammatory responses, such as hypersensitivity and allergic reactions.36 Zymosan can activate MCs through Toll-like receptor 2 (TLR-2), leading to the degranulation of pro-inflammatory molecules such as histamines and NGF and proteases such as chymase and tryptase.37 Almost 25% of the MCs in the zymosan group in our study were degranulated. Therefore, these results suggests that the vulvar allodynia and hypersensitivity that persisted 20 days after the 3rd zymosan challenge have been initiated by inflammation.
Once activated, MCs secrete pro-inflammatory mediators and signaling molecules involved in mediating vulvar nerve sensitization.38,39 Hence, the stabilization of MCs in repeated inflammation cases can potentially prevent chronic vulvar pain development. To examine this hypothesis, we used the MCs stabilizer-ketotifen fumarate, an antiallergic and antihistaminic agent that inhibits the calcium-dependent degranulation of MCs and noncompetitively blocks histamine at the H1-receptor.40 In the current study, rats were pre-treated with ketotifen fumarate (i.p; 3mg/k) before each zymosan challenge in the vulva and every day to day 7 after the 3rd zymosan challenge (overall 21 days of treatment). During the zymosan challenge, there was a significant decrease in mechanical threshold compared to baseline in both zymosan groups-The zymosan group treated with ketotifen fumarate (zymosan-KF) and the zymosan group treated with saline (zymosan-saline). No significant decrease in mechanical threshold was observed in the saline group treated with ketotifen fumarate (saline-KF). In each round, after the zymosan administration, the zymosan-KF group showed an advantage over the zymosan-saline group in terms of reduced pain response. Even though the difference in pain sensitivity reduction between the zymosan-KF and the zymosan-saline groups did not reach statistical significance (5 days in the 1st round, 2 and 5 days in the 2nd round), the zymosan-KF group tended on average to show a slight decrease in mechanical threshold compared to the zymosan-saline groups that displayed a robust decrease in mechanical threshold. This pattern of reduced pain response during inflammation as a result of KF treatment is consistent with previous studies reporting a reduction in acute inflammatory nociception in animals following KF treatment.41,42
Furthermore, the difference in mechanical sensitivity between the zymosan-saline and the zymosan-KF group was maintained even after 76 days following the 3rd zymosan challenge and long after the treatment period ended. It should be noted that the mechanical threshold of the zymosan-saline and the zymosan-KF group was not recovered to the baseline. However, only the zymosan-KF group exhibited a significant recovery to the nonallodynic levels (defined as >66% of baseline threshold) compared to the zymosan-saline group, in which persistent allodynia was present in 25% of rats in the zymosan-KF group in contrast 85% in the zymosan-saline group. However, there was no significant difference in the number of nociceptive responses in the thermal sensitivity test between the zymosan-saline and the zymosan-KF groups.
In the present study, vulvar allodynia persisted for at least 156 days after the edema and the inflammation subsided. However, even at 81 days and 160 days of the 3rd round of zymosan challenge, we found a significant difference in the number of MCs, but not of degranulated cells, between the zymosan and the saline group. In addition, there were fewer MCs in the zymosan group 160 days after the 3rd round of the zymosan challenge compared to after 20 days. MCs accumulation in the allodynic rats long after the 3rd zymosan challenge is compatible with histopathological studies of human biopsy tissues in case of chronic pain conditions (eg, vulvodynia and chronic knee pain) in which hyperinnervation is reported to occur, accompanied by an increase in the MCs presence with no evidence of inflammation.43,44 However, it should be noted that PV is a pain syndrome with a suspected inflammatory background, thus the lack of inflammation does not exclude a diagnosis of PV.45
Regarding KF treatment, the current findings depict a decrease in MCs accumulation long after the end of the treatment period in the zymosan group that was treated with KF during the inflammation, compared to the zymosan-saline group. Previous work report that KF treatment reduces MCs number and degranulation. For instance, pre-treatment with KF reduced the presence and the activation of MCs in the inflammation model of Complete Freund’s adjuvant;42 in addition, 14 days of KF treatment led to decreasing in MCs number in a rat model of endometriosis.46 However, the beneficial effect of KF in MCs modulation was mainly examined immediately after the end of the treatment. Therefore, the current results (to our knowledge) represent the first report demonstrating that KF treatment during inflammation in a rat model of PV reduced MCs accumulation long after the treatment period. This result raises the possibility that early inflammatory events, partially mediated by MCs, are involved in setting the stage for long-term MCs increases in the vulvar. Moreover, since the activation of MCs could establish a positive feedback loop that might also contributes to maintaining the pain,47 it is possible that limiting MCs activation may disrupt this loop and reduce pain. Indeed, future studies and clinical trials are needed to examine whether the modulation of MCs activity in the case of idiopathic pain (eg, Provoked vulvodynia, fibromyalgia) contributes to relieve pain.
A substantial body of evidence, based on both animal and human studies, demonstrates that MC-derived NGF plays a pivotal role in the development of hypersensitivity during inflammation.16,48,49 Therefore, it is no surprise that we found a significant increase in NGF concentration in the vulvar tissue after 24 hours of zymosan administration in the 1st and the 2nd round compared to the saline group. More critically, pretreatment with MCs stabilizer KF prevented the upregulation of NGF concentration in the vulva 24 hours after the zymosan administration in the 1st and the 2nd challenge. Indeed, the prevention of upregulation of NGF during inflammation has been suggested as one of the mechanisms whereby KF inhibits inflammatory pain.42,50 However, after 24 hours of the last zymosan injection, there was no significant difference in NGF concentration between the saline, the zymosan-KF, and the zymosan group. It might be suggested that the inflammation challenge occurring at a short time interval affects the inflammation response pathways in a way that causes the peak of NGF release to occur at different time points compared to the 1st and 2nd rounds.
Tissue inflammation induces glutamate release from the area of the inflammation, resulting in nerve sensitization that is reflected in hypersensitivity and a decrease of the pain threshold.17,51 In the current study, we examined the glutamate concentration at the early stage of the inflammation in the vulvar tissue; we found a significant increase in glutamate concentration in the vulvar tissue after 6, 24, and 48 hours of the zymosan challenge. However, the increase in glutamate and NGF concentrations was less detectable by day 30 after the 3rd zymosan injection. This result suggests that the transient changes that include a significant increase in NGF and glutamate levels during the inflammation might be involved in the development of long-lasting vulvar pain.
Numerous studies have shown that NGF and glutamate induce considerable alterations in nerve fiber distribution and influence innervating neurons’ phenotype, including growth-associated structural proteins, neuromodulators, and ion channels.17,52–54 Therefore, and since NGF and glutamate levels are substantially increased in inflamed tissues, it is not surprising that nerves’ hyperinnervation and neuromodulation occur during the inflammatory process.17,55,56 In line with these reports, substantial modifications in nerve fiber distribution were observed in the current study in the zymosan group, 20 days after the 3rd zymosan challenge, including hyperinnervation of the vulvar nerves and an increase in the expression of pain channels TRPV1 and TRPA1. This result is also with previous evidence showing the presence of sensory hyperinnervation and the overexpression of pain channels (TRPV1 and TRPA1) during an inflammatory response mediated by NGF and glutamate, which was reflected in mechanical and thermal hypersensitivity.51,52,57
Notably, in the current study, we demonstrate that even in the absence of overt inflammation, robust sensory hyperinnervation and overexpression of pain channels were found in the allodynic rats long after the 3rd zymosan challenge. This result suggests that vulvar hypersensitivity that persists long after the resolution of inflammation may be due to an abnormal persistence of the hyperinnervation and the overexpression of the pain channels. These findings might be consistent with some of the characteristics of provoked vulvodynia (PV). Indeed, hyperinnervation and overexpression of pain channels are often reported in women with PV.14,58 Similar results of vulvar hypersensitivity and hyperinnervation resulting from inflammation challenge were also reported in the mice model of vulvar pain.15,32,59
More importantly, we found that treatment with an MC stabilizer during inflammation induced by zymosan (21 days of treatment) attenuates neuromodulation and hyperinnervation in the vulva. After 81 days of the 3rd zymosan challenge, we found a significant difference in the expression of pain channels (especially the TRPV1 channel) and nerve density in the vulva between the zymosan-KF group and the zymosan-saline group. As for TRPA1, there was no significant difference in the expression level between the two groups, but when looking at the group level, the expression of the TRPA1 channel in the zymosan-KF group was diminished compared to the zymosan-saline group. These results are consistent with earlier studies.46 It was suggested that increased local nerve density and overexpression of pain channels might be MC-dependent and that the depletion of MCs reduces innervation and downregulates pain-channels expressions.26,32,59,60 Following current results, we speculate that modulation of MCs activity by KF during inflammation in the vulva may explain the difference in hyperinnervation and neuromodulation between the zymosan-KF group and the zymosan-saline group.
Several limitations of our study should be acknowledged. First, multiple studies suggest that inflammation may play a role in the development of PV.5,28,61 The current result support this notion, indeed, we found that repeated inflammation led to long-lasting mechanical and thermal vulvar hypersensitivity. However, in the current study we focused only on inflammation in the development of PV, and this a limitation of the current study. And, it should be noted that the development of chronic vulvar pain is not solely induced by inflammation.6,62 PV is a complex pain disorder, and the etiology is multifactorial.6 Thus, it is possible that in women’s the combined effects and the interaction of several factors (eg, psychological stress, genetic predisposition, hormonal imbalance, and recurrent inflammation) are essential in promoting PV development. Therefore, future studies could assess the combined effects and the interaction between these factors in the development of provoked chronic vulvar pain. Moreover, despite the clinical evidence that suggests a correlation between vulvar fungal infections (ie, topical infection) that lead to inflammation, and PV. Here, we used zymosan injection in the vulvar to induce inflammation, and this may a limitation of the current model.
The current study focused on PV, which is the most common type of vulvodynia,4,63 thus the results obtained in the current study may not provide information to understand the development of other types of vulvodynia (eg, generalized vulvodynia) and this a limitation of the current model. However, our findings may have an impact, in the future, on the vast majority of vulvodynia sufferers. Finally, our findings indicate that stabilization of MCs by KF reversed inflammatory-induced vulvar mechanical allodynia, suggesting that limiting MC activation is might crucial to resolve the ongoing inflammation and consequently, modulation of chronic vulvar pain development. Thus, the translational value of our findings is substantial as it suggests that stabilizing MCs in women with vulvar inflammation history may be useful to promote analgesia during inflammation and prevent chronic vulvar pain development, especially among women with a history of seasonal allergies, which are twice as likely to develop vulvodynia than allergy-free controls.64 Therefore, we encourage future clinical trials of KF to be used as a preventive treatment for PV in the case of repeated vulvar inflammation.
Conclusion
Taken together, we have developed a new, clinically relevant animal model of persistent vulvar pain. Furthermore, our results reveal robust evidence that mechanical and thermal sensitivity induced by inflammation is mediated partly by MC accumulation, nerve growth, and the increase of pain channels expression in the vulvar neurons, resulting in hypersensitivity reflected in decreased pain threshold, eventually manifested as allodynia and hyperalgesia, which are some of the reported symptoms of PV. Current results align with previous reports demonstrating that recurrent vulvar inflammation induces long-lasting neuronal modifications reflected in local hypersensitivity.15 Nevertheless, following current results, we can speculate, at least to some degree, that the early inflammatory events such as MCs degranulation and glutamate and NGF upregulation that we described mediate the neuronal changes that were observed in allodynic rats.
However, a more thorough examination of a specific inflammatory pathway that takes into account mast cell-centered tissue response at the early inflammatory events revealed that MCs stabilization by ketotifen fumarate attenuates the upregulated level of NGF during inflammation, modulate the neuronal modifications, reduces MCs accumulation, and enhanced mechanical hypersensitivity that occurs after repeated inflammation challenge (Figure 13). Considering that MCs are known to be involved in the development of chronic pain conditions like PV, insight derived from the present study may provide vital information towards an improved understanding of the role of MCs and the early inflammatory events in the development of provoked vulvodynia (Figure 13). In addition, the current findings encourage future clinical studies aimed at testing KF’s efficacy in reducing and preventing chronic vulvar pain development, especially in women with predisposition to allergy. Furthermore, the current findings might also have a possible implication on prevention and treatment of chronic idiopathic pain conditions such as fibromyalgia, that co-occur with different types of allergies.42
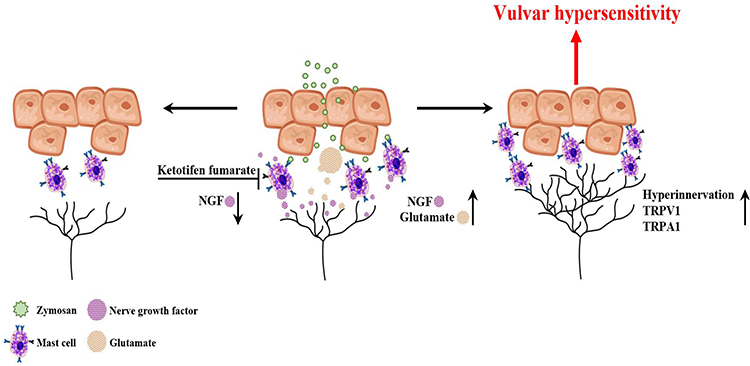 |
Figure 13 Diagram illustrating the involvement of mast cells in chronic vulvar pain development. Inflammation induced by zymosan leads to glutamate and NGF release from the inflammation area. The early inflammatory events (MC degranulation, NGF, and glutamate release) set the stage for long-term MC accumulation, nerves neuromodulation, and hyperinnervation manifested as vulvar hypersensitivity. In turn, mast cell stabilization by ketotifen fumarate attenuates the upregulated level of NGF during inflammation, modulates the neuronal modifications, reduces mast cells accumulation, and enhances mechanical hypersensitivity after repeated inflammation challenges. |
Abbreviations
PV, provoked vulvodynia; MC, mast cell; NGF, nerve growth factor; TRPA1, transient receptor potential vanilloid-1; TRPV1, transient receptor potential ankyrin-1- TRPA1; KF, ketotifen fumarate.
Data Sharing Statement
The datasets during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethical Review Committee Statement
All animal procedures were approved by the Bar-Ilan University Animal Care Committee (code# 84-11-2019, November 2019) and were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Acknowledgments
The authors would like to thank Jabarin R, and Sakas R, for their valuable comments.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. JB and EP contributed equally to this work.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Disclosure
The authors declare that they have no conflicts of interest in relation to this work.
References
1. Bornstein J, Goldstein AT, Stockdale CK, et al. 2015 ISSVD, ISSWSH and IPPS consensus terminology and classification of persistent vulvar pain and vulvodynia. Obstet Gynecol. 2016;127(4):745–751. doi:10.1097/AOG.0000000000001359
2. Woolf CJ, Ma Q. Nociceptors—noxious stimulus detectors. Neuron. 2007;55(3):353–364. doi:10.1016/j.neuron.2007.07.016
3. Bao C, Noga H, Allaire C, et al. Provoked vestibulodynia in women with pelvic pain. Sex Med. 2019;7(2):227–234. doi:10.1016/j.esxm.2019.03.002
4. Henzell H, Berzins K, Langford JP. Provoked vestibulodynia: current perspectives. Int J Women's Health. 2017;9:631–642. doi:10.2147/IJWH.S113416
5. Akopians AL, Rapkin AJ. Vulvodynia: the role of inflammation in the etiology of localized provoked pain of the vulvar vestibule (vestibulodynia). Semin Reprod Med. 2015;33(4):239–245. doi:10.1055/s-0035-1554919
6. Lev-Sagie A, Witkin SS. Recent advances in understanding provoked vestibulodynia. F1000Research. 2016;5:2581. doi:10.12688/f1000research.9603.1
7. Aalto A, Huotari-Orava R, Luhtala S, Mäenpää J, Staff S. Expression of estrogen-related receptors in localized provoked vulvodynia. Biores Open Access. 2020;9(1):13–21. doi:10.1089/biores.2019.0049
8. Liao Z, Smith PG. Persistent genital hyperinnervation following progesterone administration to adolescent female rats. Biol Reprod. 2014;91(6):1–9. doi:10.1095/biolreprod.114.121103
9. Kalfon L, Azran A, Farajun Y, et al. Localized provoked vulvodynia: association with nerve growth factor and transient receptor potential vanilloid type 1 genes polymorphisms. J Low Genit Tract Dis. 2019;23(1):58–64. doi:10.1097/LGT.0000000000000445
10. Heddini U, Bohm-Starke N, Grönbladh A, Nyberg F, Nilsson KW, Johannesson U. Serotonin receptor gene (5HT‐2A) polymorphism is associated with provoked vestibulodynia and comorbid symptoms of pain. J Sex Med. 2014;11(12):3064–3071. doi:10.1111/jsm.12685
11. Zhang Z, Zolnoun DA, Francisco EM, Holden JK, Dennis RG, Tommerdahl M. Altered central sensitization in subgroups of women with vulvodynia. Clin J Pain. 2011;27(9):755. doi:10.1097/AJP.0b013e31821c98ec
12. Khandker M, Brady SS, Stewart EG, Harlow BL. Is chronic stress during childhood associated with adult-onset vulvodynia? J Women's Health. 2014;23(8):649–656. doi:10.1089/jwh.2013.4484
13. Witkin SS, Gerber S, Ledger WJ. Differential characterization of women with vulvar vestibulitis syndrome. Am J Obstet Gynecol. 2002;187(3):589–594. doi:10.1067/mob.2002.125889
14. Bornstein J, Palzur E. Neuroproliferative processes in vulvodynia. In: Female Sexual Pain Disorders. Wiley; 2020:43–51.
15. Farmer MA, Taylor AM, Bailey AL, et al. Repeated vulvovaginal fungal infections cause persistent pain in a mouse model of vulvodynia. Sci Transl Med. 2011;3(101):101ra91. doi:10.1126/scitranslmed.3002613
16. Gangadharan V, Kuner R. Pain hypersensitivity mechanisms at a glance. Dis Model Mech. 2013;6(4):889–895. doi:10.1242/dmm.011502
17. Honda K, Shinoda M, Kondo M, et al. Sensitization of TRPV1 and TRPA1 via peripheral mGluR5 signaling contributes to thermal and mechanical hypersensitivity. Pain. 2017;158(9):1754–1764. doi:10.1097/j.pain.0000000000000973
18. Schnegelsberg B, Sun TT, Cain G, et al. Overexpression of NGF in mouse urothelium leads to neuronal hyperinnervation, pelvic sensitivity, and changes in urinary bladder function. Am J Physiol. 2010;298(3):R534–47. doi:10.1152/ajpregu.00367.2009
19. Ji RR, Samad TA, Jin SX, Schmoll R, Woolf CJ. p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. Neuron. 2002;36(1):57–68. doi:10.1016/S0896-6273(02)00908-X
20. Obata K, Katsura H, Mizushima T, et al. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J Clin Invest. 2005;115(9):2393. doi:10.1172/JCI25437
21. Zhang X, Huang J, McNaughton PA. NGF rapidly increases membrane expression of TRPV1 heat-gated ion channels. EMBO J. 2005;24(24):4211–4223. doi:10.1038/sj.emboj.7600893
22. Chadha S, Gianotten WL, Drogendijk AC, Weijmar Schultz WCM, Blindeman LAJ, van der Meijden WI. Histopathologic features of vulvar vestibulitis. Int J Gynecolog Pathol. 1998;17(1):7–11. doi:10.1097/00004347-199801000-00002
23. Regauer S, Eberz B, Beham-Schmid C. Mast cell infiltrates in vulvodynia represent secondary and idiopathic mast cell hyperplasias. APMIS. 2015;123(5):452–456. doi:10.1111/apm.12372
24. Schmelz M, Zeck S, Raithel M, Rukwied R. Mast cell tryptase in dermal neurogenic inflammation. Clin Exp Allerg. 1999;29(5):695–702. doi:10.1046/j.1365-2222.1999.00514.x
25. Leon A, Buriani A, Dal Toso R, et al. Mast cells synthesize, store, and release nerve growth factor. Proc Natl Acad Sci U S A. 1994;91(9):3739–3743. doi:10.1073/pnas.91.9.3739
26. Chatterjea D, Martinov T. Mast cells: versatile gatekeepers of pain. Mol Immunol. 2015;63(1):38. doi:10.1016/j.molimm.2014.03.001
27. Keppel Hesselink JM, Kopsky DJ, Sajben N. New topical treatment of vulvodynia based on the pathogenetic role of cross talk between nociceptors, immunocompetent cells, and epithelial cells. J Pain Res. 2016;9:757–762. doi:10.2147/JPR.S115407
28. Paavonen J, Eschenbach DA. Localized provoked vulvodynia-an ignored vulvar pain syndrome. Front Cell Infect Microbiol. 2021;11:550. doi:10.3389/fcimb.2021.678961
29. Wesselmann U, Bonham A, Foster D. Vulvodynia: current state of the biological science. Pain. 2014;155(9):1696–1701. doi:10.1016/j.pain.2014.05.010
30. Ren K, Dubner R. Interactions between the immune and nervous systems in pain. Nat Med. 2010;16(11):1267. doi:10.1038/nm.2234
31. Gupta K, Harvima IT. Mast cell-neural interactions contribute to pain and itch. Immunol Rev. 2018;282(1):168. doi:10.1111/imr.12622
32. Landry J, Martinov T, Mengistu H, et al. Repeated hapten exposure induces persistent tactile sensitivity in mice modeling localized provoked vulvodynia. Dileepan KN, ed. PLoS One. 2017;12(2):e0169672. doi:10.1371/journal.pone.0169672
33. Basha ME, Kellogg-Spadt S, Burrows LJ, et al. Thermal and mechanical pain thresholds of women with provoked localized vulvodynia: a pilot study. J Am Osteopathic Assoc. 2019;119(3):164–172. doi:10.7556/jaoa.2019.027
34. Young SH, Robinson VA, Barger M, Porter DW, Frazer DG, Castranova V. Acute inflammation and recovery in rats after intratracheal instillation of a 1→3-β-glucan (zymosan A). J Toxicol Environ Health. 2001;64(4):311–325. doi:10.1080/152873901316981303
35. Sobel JD. Vulvovaginal candidosis. Lancet. 2007;369(9577):1961–1971. doi:10.1016/S0140-6736(07)60917-9
36. da Silva EZM, Jamur MC, Oliver C. Mast cell function: a new vision of an old cell. J Histochem Cytochem. 2014;62(10):698–738. doi:10.1369/0022155414545334
37. Graham AC, Temple RM, Obar JJ. Mast cells and influenza A virus: association with allergic responses and beyond. Front Immunol. 2015;6. doi:10.3389/fimmu.2015.00238
38. Gupta S, Kakkar V, Bhushan I. Crosstalk between vaginal microbiome and female health: a review. Microb Pathog. 2019;136:103696. doi:10.1016/j.micpath.2019.103696
39. Theoharides TC, Alysandratos KD, Angelidou A, et al. Mast cells and inflammation. Biochim Biophys Acta. 2012;1822(1):21. doi:10.1016/j.bbadis.2010.12.014
40. Ben-Eli H, Solomon A. Topical antihistamines, mast cell stabilizers, and dual-action agents in ocular allergy: current trends. Current Opinion in Allergy & Clinical Immunology. 2018;18:411–416. doi:10.1097/ACI.0000000000000473
41. Anoush M, Mohammad Khani MR. Evaluating the anti-nociceptive and anti-inflammatory effects of ketotifen and fexofenadine in rats. Adv Pharm Bull. 2015;5(2):217. doi:10.15171/apb.2015.030
42. Meloto CB, Ingelmo P, Perez EV, et al. Mast cell stabilizer ketotifen fumarate reverses inflammatory but not neuropathic-induced mechanical pain in mice. Pain Rep. 2021;6(2):e902. doi:10.1097/PR9.0000000000000902
43. Sanchis-Alfonso V, Roselló -sastre E. Immunohistochemical analysis for neural markers of the lateral retinaculum in patients with isolated symptomatic patellofemoral malalignment a neuroanatomic basis for anterior knee pain in the active young patient. Am J Sports Med. 2000;28(5):725–731.
44. Bornstein J, Cohen Y, Zarfati D, Sela S, Ophir E. Involvement of heparanase in the pathogenesis of localized vulvodynia. Int J Gynecolog Pathol. 2008;27(1):136–141. doi:10.1097/pgp.0b013e318140021b
45. Tommola P, Bützow R, Unkila-Kallio L, Paavonen J, Meri S. Activation of vestibule-associated lymphoid tissue in localized provoked vulvodynia. Am J Obstet Gynecol. 2015;212(4):
46. Zhu T, Zou G, Ding S, et al. Mast cell stabilizer ketotifen reduces hyperalgesia in a rodent model of surgically induced endometriosis. J Pain Res. 2019;12:1359–1369. doi:10.2147/JPR.S195909
47. Vincent L, Vang D, Nguyen J, et al. Mast cell activation contributes to sickle cell pathobiology and pain in mice. Blood. 2013;122(11):1853–1862. doi:10.1182/blood-2013-04-498105
48. Barker PA, Mantyh P, Arendt-Nielsen L, Viktrup L, Tive L. Nerve growth factor signaling and its contribution to pain. J Pain Res. 2020;13:1223–1241. doi:10.2147/JPR.S247472
49. Mai L, Liu Q, Huang F, He H, Fan W. Involvement of mast cells in the pathophysiology of pain. Front Cell Neurosci. 2021;15:205. doi:10.3389/fncel.2021.665066
50. Massaad CA, Safieh-Garabedian B, Poole S, Atweh SF, Jabbur SJ, Saadé NE. Involvement of substance P, CGRP and histamine in the hyperalgesia and cytokine upregulation induced by intraplantar injection of capsaicin in rats. J Neuroimmunol. 2004;153(1–2):171–182. doi:10.1016/j.jneuroim.2004.05.007
51. Masuoka T, Yamashita Y, Yoshida J, et al. Sensitization of glutamate receptor-mediated pain behaviour via nerve growth factor-dependent phosphorylation of transient receptor potential V1 under inflammatory conditions. Br J Pharmacol. 2020;177(18):4223–4241. doi:10.1111/bph.15176
52. Almarestani L, Longo G, Ribeiro-da-Silva A. Autonomic fiber sprouting in the skin in chronic inflammation. Mol Pain. 2008;4:
53. Pitake S, Middleton LJ, Abdus-Saboor I, Mishra SK. Inflammation induced sensory nerve growth and pain hypersensitivity requires the N-type calcium channel Cav2.2. Front Neurosci. 2019;13:1009. doi:10.3389/fnins.2019.01009
54. Chung MK, Ro JY. Peripheral glutamate receptor and transient receptor potential channel mechanisms of craniofacial muscle pain. Molecular Pain. 2020;174480692091420. doi:10.1177/1744806920914204
55. Carr MJ, Undem BJ. Inflammation-induced plasticity of the afferent innervation of the airways. Environ Health Perspect. 2001;109(SUPPL. 4):567–571. doi:10.1289/ehp.01109s4567
56. Gegelashvili G, Bjerrum OJ. High-affinity glutamate transporters in chronic pain: an emerging therapeutic target. J Neurochem. 2014;131(6):712–730. doi:10.1111/jnc.12957
57. Diogenes A, Akopian AN, Hargreaves KM, El-Bialy T. NGF up-regulates TRPA1: implications for orofacial pain. The Angle Orthodontist. 2016;86(6):550–555. doi:10.1177/154405910708600612
58. Tympanidis P, Casula MA, Yiangou Y, Terenghi G, Dowd P, Anand P. Increased vanilloid receptor VR1 innervation in vulvodynia. Eur J Pain. 2004;8(2):129–133. doi:10.1016/S1090-3801(03)00085-5
59. Martinov T, Glenn-Finer R, Burley S, et al. Contact hypersensitivity to oxazolone provokes vulvar mechanical hyperalgesia in mice. PLoS One. 2013;8(10):e78673. doi:10.1371/journal.pone.0078673
60. van Diest SA, Stanisor OI, Boeckxstaens GE, de Jonge WJ, van den Wijngaard RM. Relevance of mast cell–nerve interactions in intestinal nociception. Biochimica et Biophysica Acta. 2012;1822(1):74–84. doi:10.1016/j.bbadis.2011.03.019
61. Tommola P, Unkila-Kallio L, Paetau A, Meri S, Kalso E, Paavonen J. Immune activation enhances epithelial nerve growth in provoked vestibulodynia. Am J Obstet Gynecol. 2016;215(6):
62. Torres-Cueco R, Nohales-Alfonso F, Luisa M, Ferrer S, Moran Sanchez I, Gonzalo JJA. Vulvodynia—it is time to accept a new understanding from a neurobiological perspective. Int J Environ Res Public Health. 2021;18(12):6639. doi:10.3390/ijerph18126639
63. Sadownik LA. Etiology, diagnosis, and clinical management of vulvodynia. Int J Women's Health. 2014;6(1):437–449. doi:10.2147/IJWH.S37660
64. Harlow BL, He W, Nguyen RHN. Allergic Reactions and Risk of Vulvodynia. Ann Epidemiol. 2009;19(11):771. doi:10.1016/j.annepidem.2009.06.006
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.