Back to Journals » Nature and Science of Sleep » Volume 18

Changed Serum Levels of Sirtuin1 and Sirtuin3 and Their Correlations with Sleep Quality and Cognitive Function in Patients with Chronic Insomnia Disorder
Authors Liu GX ![]() , Zhang Y, Song Y, Yang S
, Zhang Y, Song Y, Yang S ![]() , Su AX
, Su AX ![]() , Feng YZ
, Feng YZ ![]() , Zhou YS
, Zhou YS ![]() , Ma ZJ, Xia L, Chen GH
, Ma ZJ, Xia L, Chen GH ![]()
Received 26 January 2025
Accepted for publication 28 January 2026
Published 3 March 2026 Volume 2026:18 518388
DOI https://doi.org/10.2147/NSS.S518388
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Valentina Alfonsi
Gao-Xia Liu, Yu Zhang, Yang Song, Shuai Yang, Ai-Xi Su, Yi-Zhou Feng, Yu-Shun Zhou, Zi-Jie Ma, Lan Xia, Gui-Hai Chen
Department of Neurology (Sleep Disorders), The Affiliated Chaohu Hospital of Anhui Medical University, Hefei, People’s Republic of China
Correspondence: Gui-Hai Chen, Email [email protected] Lan Xia, Email [email protected]
Purpose: The purpose of this study is to investigate the relationship between the serum concentrations of the mitochondrial markers sirtuins-1 and sirtuins-3, sleep quality and cognitive function in individuals with chronic insomnia disorder (CID).
Patients and Methods: A cohort of 80 patients with CID and 42 healthy controls (HCs) were recruited for the study. Serum concentrations of SIRT1 and SIRT3 were quantified using enzyme-linked immunosorbent assays. Sleep quality was evaluated utilizing the Pittsburgh Sleep Quality Index (PSQI) and polysomnography. The cognitive assessment tools employed included the Chinese version of the Montreal Cognitive Assessment (MoCA-C) for evaluating overall cognitive function, the Nine Box Maze Test (NBMT) for assessing multidimensional memory, and the Blue Velvet Arena Test (BVAT) for examining spatial memory.
Results: In comparison to HCs, patients with CID exhibited a significantly elevated PSQI score (Z=− 9.094, P< 0.001), reduced MoCA score (Z=− 4.075, P< 0.001), and increased error distance in the BVAT task (Z=− 2.096, P< 0.05), with more errors in multi-memory components consisting of spatial reference (Z=− 2.963, P< 0.01), spatial working (Z=− 2.455, P< 0.05), object working (Z=− 2.356, P< 0.05), and object recognition (Z=− 2.025, P< 0.05) in the NBMT task. The patients also had increased serum levels of sirtuins-1 (Z=6.718, P< 0.001) and sirtuins-3 (Z=− 6.223, P< 0.001). Analysis of partial correlation revealed that the patients’ serum concentrations of sirtuins-1 and sirtuins-3 were positively associated with the PSQI score and spatial reference memory errors in the NBMT and negatively correlated with the delayed recall score in the MoCA-C. The sirtuins-1 level were positively associated with object recognition memory errors in the NBMT. Furthermore, sirtuins-3 level was negatively correlated with patients’ illness duration.
Conclusion: Patients with CID may have mitochondrial dysfunction in the brain which might be associated with worse sleep and cognition.
Keywords: cognition dysfunction, insomnia disorder, mitochondria, sirtuins
Introduction
Insomnia poses a significant challenge to public health, affecting nearly one-third of the population with its symptoms.1 Difficulties in initiating sleep, maintaining sleep continuity, or arising earlier than expected are the hallmarks of insomnia disorder, even when there is enough time for sleep.2 The ICSD-3 outlines that the diagnostic standards for Chronic Insomnia Disorder (CID) include a minimum duration of three months and a frequency of occurrence at least thrice a week.3 Every year, the prevalence of insomnia is rising. Long-term insomnia has a substantial adverse effect on physical and emotional well-being, life, work, study, and interpersonal communication, and can even lead to unexpected malignant events. Sleep can also lead to an increase in the incidence of physical diseases, such as hypertension, multiple sclerosis (MS), Parkinson’s disease (PD), Alzheimer’s disease (AD), stroke, traumatic brain injury (TBI), and epilepsy. Insomnia shows a particularly strong correlation with depression and can double the risk of suicidal ideation and behavior.4 Therefore, chronic insomnia imposes a significant burden on both individuals and communities. In recent years, progress has been made in treating chronic insomnia,5 but the mechanism of chronic insomnia is not yet clear.
The cognitive function impairment of insomnia patients is one of the main forms of daytime impairment, which is also some patients’ primary concern for medical treatment. A meta-analysis of clinical studies has shown that cognitive dysfunction in patients with insomnia mainly focuses on attention, memory, and executive function.6,7 However, the precise connection between insomnia and cognitive decline remains uncertain, and few studies have investigated this relationship, with inconsistent conclusions. In recent years, many scholars have studied the relationship between biomarkers and cognitive function in patients with CID.8,9 This study mainly reveals the relationship between mitochondria and cognitive impairment in patients with chronic insomnia, which has never been explored before.
While it is well known that sleep is an extremely complex physiological process and its biological basis has not been fully elucidated, there is strong evidence to support a connection between circadian rhythms, sleep, and metabolism, suggesting that mitochondria play an essential role in sleep,10 However, it is less clear how mitochondria and sleep interact. Animal studies have clearly indicated that mitochondria are involved in sleep, with oxidative stress and mitochondrial malfunction identified as potential underlying causes. Although limited, human studies correspond with animal research and further substantiate the role of sleep deprivation in inducing oxidative stress and mitochondrial dysfunction.11 This indicates that mitochondria play a role in regulating the sleep-wake cycle.
Mitochondria are important and independent organelles in eukaryotic cells with a variety of functions, from biological energy to cell signaling.12 Mitochondria are located in the axon terminals and dendrites of neurons, where they significantly contribute to synaptic function and plasticity. The association between mitochondrial dysfunction and reduced synaptic plasticity in the brain indicates that mitochondria are crucial for age-related memory decline. Nonetheless, the precise role of mitochondria in regulating the cellular mechanisms of learning and memory remains largely unclear.13 Genetic mutations and cumulative damage to mitochondria during aging are key factors in cognitive deficits in neurodevelopmental disorders and neurodegenerative diseases.14 Mitochondrial dysfunction is at the center of many neurodegenerative diseases and can lead to harmful effects on neural stem cell function, both during development and in the adult brain.15 Many mitochondrial diseases manifest as cognitive dysfunction in the central nervous system, leading to behavioral abnormalities.16 When sleep is insufficient, the brain is in a state of high energy demand for a long time,17 which leads to a series of negative events, starting from the increase in oxidative stress, leading to mitochondrial dysfunction. When mitochondrial function is damaged, ATP production decreases, which has an immeasurable impact on the plasticity of neurons and synapses, resulting in a decline in cognitive levels.
Existing studies have shown that histone deacetylases (HDAC) play a role in protecting neurons and helping them survive.18 HDACs are categorized into four classes (I–IV), according to their functions and locations. Notably, class III HDACs, known as SIRTs, exhibit both neuroprotective and neurotoxic effects.19 Past reports have highlighted that some sirtuins, particularly silence-regulatory protein 1 (SIRT1) and 3 (SIRT3), are active in several brain regions.20 They support the maintenance of higher levels of brain function by representing the most significant sirtuins activity in neurons, which control the majority of important brain activities and are believed to be implicated in the pathophysiology of nearly all age-related and neurodegenerative brain diseases and protect neuronal cells from non-physiological factors and neuronal degeneration.21 SIRT1 and SIRT3 play a role in neuron survival, proliferation, and response to stress control, and their overexpression has protective effect on neurons.
SIRT1 is the most important and widely studied member of sirtuins and has been shown to be expressed primarily in neurons.22 It is present in the nucleus of most cells, has both nuclear localization and nuclear output signals, and can shuttle between the cytoplasm and the nucleus.23 Its activity is closely related to the biogenesis and turnover of mitochondria. Depletion of SIRT1 may negatively impact cognitive processes, such as short-term memory, classical conditioning, and spatial learning.24 This indicates that SIRT1 is essential for sustaining advanced brain function and synaptic remodeling.
SIRT3 is mainly located in the mitochondrial matrix, where it remains active. It is vital for reducing oxidative stress and is involved in the control of mitochondrial activities.25 Additionally, it regulates mitosis to eliminate modified mitochondria.26 SIRT3 has also been directly linked to increased longevity of nerve cells and human lifespan. Its overexpression and stimulation may shield neurons against pathological modifications.27 SIRT3 is essential for preserving mitochondrial integrity, making it a significant focus in investigating the pathogenesis of neurodegenerative encephalopathy, and therefore evaluating SIRT3 can be a possible target for therapy in managing neurological diseases produced by mitochondrial dysfunction.28
The objective of this study is to find out whether the concentration of mitochondrial biomarkers SIRT1 and SIRT3 changes in patients with CID and, if so, whether the change is related to the severity of insomnia and cognitive function decline in CID patients, and whether the serum marker is valuable for distinguishing patients with CID from healthy sleepers.
Materials and Methods
Subjects
Qualified and voluntary subjects were recruited from the Sleep Disorder Department of Chaohu Hospital affiliated with Anhui Medical University from October 2021 to July 2023. The study included 122 participants in total, including 42 HCs and 80 individuals with CID. The flow-chart illustrating the study participants is shown in Figure 1. The Inclusion criteria for patients were as follows: (1) CID group: meet the diagnostic guidelines of the third edition of the International Classification of Sleep Disorders,3 At the same time, meet the following requirements: ① Duration suffered from insomnia > 6 months; ② 18–60 years old; ③ Receiving education ≥ 9 years of, without any understanding and hearing impairment; ④ The scores for the Pittsburgh Sleep Quality Index (PSQI) is > 7, while those for the Hamilton Anxiety Scale-14 (HAMA-14) is < 14, and the Patient Health Questionnaire-9 (PHQ-9) scores are < 10; ⑤ There was no history of any sedative and hypnotic drugs in the two weeks prior to the visit. (2) Healthy control group: Gender, age, and education level were matched with the CID group, no insomnia complaints and family history, no mood disorders and cognitive dysfunction; PSQI score < 7, HAMA-14 score < 7, PHQ-9 score < 5, and score ≥ 26 on the Montreal Cognitive Assessment, Chinese Version (MoCA-C). The scales were used as part of the clinical assessment and were properly cited for transparency and accountability. Moreover, patients with any medical conditions (such as immunological, endocrine, cardiovascular, neurological, mitochondrial, or degenerative brain diseases) or psychological disorders were not included in the study. In addition, patients during pregnancy or breastfeeding or patients who have taken sedative hypnotic drugs or any other drugs that may affect sleep, mood, and cognitive function within two weeks were not eligible for the study.
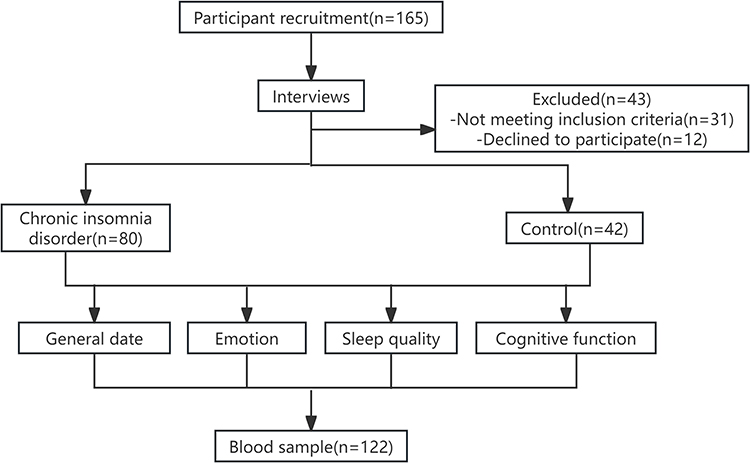 |
Figure 1 The flowchart for study participant selection. |
Baseline Data Collection
General information, such as name, age, gender, educational background, current and previous histories of all subjects, were collected.
Assessment of Subjects’ Emotions
We used the PHQ-929 and HAMA-1430 to assess the emotional state of the patients. The PHQ-9 has good reliability and validity for the diagnosis and evaluation of the severity of depression which contains 9 items that relate to the patient’s emotions. The range of total scores is 0–27, where 0 to 4 demonstrates no depression, 5 to 9 indicates minor depression, and 10 to 14 suggests moderate depression, and more than 15 signifies severe depression, with higher scores indicating more severe depression. The HAMA was used to assess anxiety levels, where a score of ≥ 7 suggests the presence of anxiety, while a score of < 7 shows an absence of anxiety.
Assessment of Sleep Status
Subjective sleep quality: The PSQI is a useful tool for measuring sleep quality and patterns,31 widely used in clinical and scientific research, it can evaluate the sleep quality of the subjects for nearly a month.32 The scale comprises seven variables, including subjective sleep quality, sleep onset latency, sleep duration, habitual sleep efficiency, sleep disorders, sleep medication use, and daytime sleep disturbance in the past month.33 The scale’s total score ranges from 0 to 21, with higher scores reflecting a deterioration in sleep quality. In China, the score line that distinguishes sleep disorders from healthy sleepers is PSQI score ≥ 7 points.34
Objective sleep quality: Polysomnography (PSG) can monitor a number of parameters related to sleep, such as breathing, blood oxygen saturation, snoring, heart rate, electroencephalogram, electrocardiogram, etc. Following the collection of data overnight, skilled technicians interpreted stages of sleep and sleep-related events in compliance with the most recent edition of the American Academy of Sleep Medicine Manual for the Scoring of Sleep and Associated Events,35 and the data indicators were finally obtained: total sleep time (TST), wake time after sleep onset (WASO), sleep efficiency (SE), sleep onset latency (SOL), wake (W), rapid eye movement (REM) sleep, REM latency (REMSL), non-rapid eye movement (NREM) sleep (including N1, N2, N3), and time percentage of each phase in TST (N1%, N2%, N3%, REM%).
Evaluation of Cognitive Function
General cognitive function: The Chinese version of the Montreal cognitive assessment (MoCA-C) was used for the subjects’ general cognitive function assessment.36,37 MoCA-C covers eight cognitive domains, encompassing short-term memory, delayed recall, naming, attention, language, abstraction, executive function, orientation, and visuospatial and executive function. The scale has an overall score of 30 points. One extra point can be given if the person has not completed twelve years of education, but the final score should not exceed more than 30 points. The higher the MoCA-C score, the less severe the cognitive impairment.
Multidimensional memory function: In this study, the multi-dimensional memory function of all participants was evaluated using a modified Nine Box Maze Test (NBMT), including spatial working memories (SWM), object working memories (OWM), spatial reference memories (SRM), object Reference Memories (ORM), and object recognition memory (ORcM). The final result of the test is expressed as the number of errors. An increase in errors is associated with a decline in memory function. This experiment has been widely used in our research group, and the results are supported by relevant literature.38,39
Spatial memory function: The subjects’ spatial memory function was assessed using the Blue Velvet Arena Test (BVAT), which was introduced and modified by our research group.9 BVAT is a human simulation of the Morris Water Maze used in animal studies.40 The experiment was carried out in eight rounds, and the average of the results was taken, which was called the average error distance. An increase in the average error distance is indicative of a decline in spatial navigation ability, which consequently reflects deterioration in spatial memory ability.
Blood Sample Collection and Storage
Following the PSG procedure, a 3 mL venous blood sample was collected at approximately 8:00 am. The collected blood samples were left for half an hour before being centrifuged at 3000r/min for a duration of 5 min. The upper serum was extracted and placed in a refrigerator maintained at ultra-low temperatures of −80°C until the sample was tested. The enzyme-linked immunosorbent assay (ELISA) was applied for measuring the concentrations of SIRT1 and SIRT3 in all samples, in accordance with the manufacturer’s guidelines (Wuhan USCN, Wuhan, China).
Statistical Analysis
All experimental data were analyzed using SPSS26.0 statistical software. A normality test was conducted on the measurement data, and variables that followed a normal distribution were expressed as the mean ± standard deviation (SD). Comparison between the two groups was performed via T-test of the mean of two independent samples. Data conforming to non-normal distributions were represented by interquartile spacing [P50 (P25, P75)], and group differences were examined using the Mann–Whitney U-test. Classified data was compared using the Chi-square test. Partial correlation analysis was applied to the continuous variables, and the correlation between serological indicators in the CID group and clinical scales, PSG parameters, illness duration, number of errors in the NBMT, and mean error gap in the BVAT was evaluated by adjusting for confounding variables such as gender, age, education level, and PHQ-9. In order to evaluate the diagnostic accuracy of the biomarkers, receiver operating characteristic (ROC) curves were constructed, and the areas under the curve (AUCs) were computed. Statistical significance in all analyses was assessed using a threshold of P≤ 0.05.
Results
Baseline Information and Subjective Sleep Quality
Analysis revealed no significant changes between the two groups regarding the gender, age, body mass index (BMI), and education years, as indicated by P > 0.05 (Table 1). In comparison to the HC group, patients with CID exhibited markedly elevated scores on the PSQI, HAMA, and PHQ-9 assessments (P < 0.001, Table 1), suggesting that the subjects in the CID group had mild depression and moderate to severe insomnia. These results suggested that the subjects met the inclusion criteria.
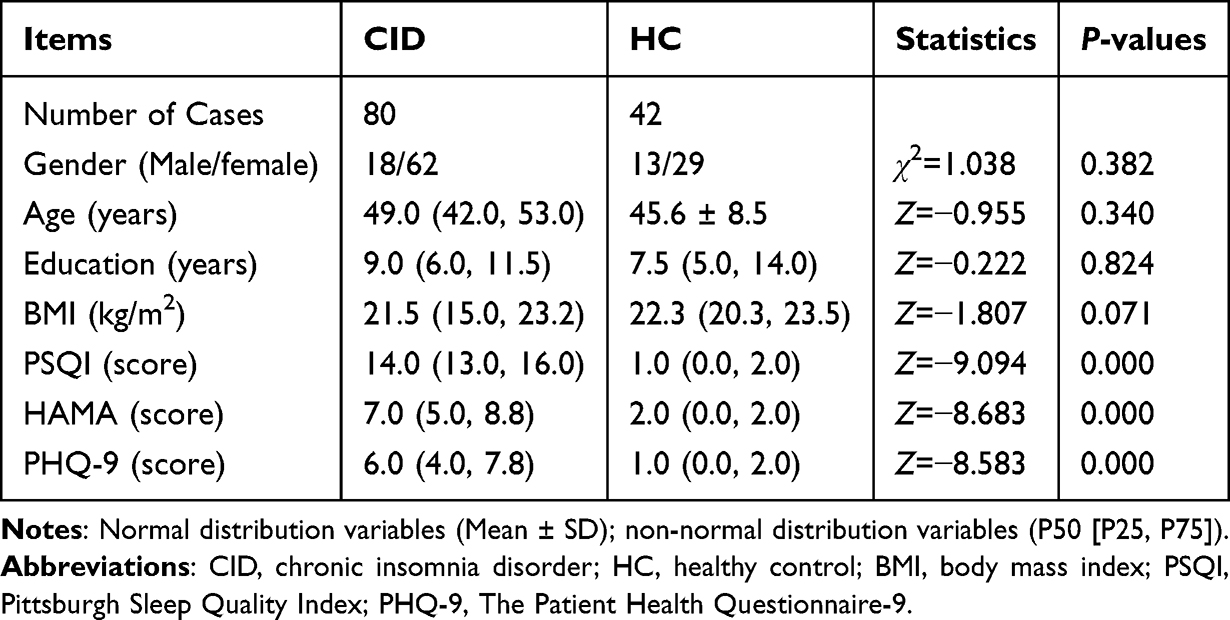 |
Table 1 General Demographic Data and PSQI and PHQ-9 Scores |
Cognitive Function
General Cognitive Function
In comparison to the HC group, patients with CID exhibited significantly lower MoCA-C scores (P<0.001, Table 2). The MoCA-C evaluation indicated that individuals with CID experienced significant reductions in scores across the visuospatial and executive function, language, delayed recall, and orientation domains (P< 0.05, Table 2).
 |
Table 2 Results From the MoCA-C Assessment |
Memory Function
In the CID group, there was a marked increase in the mean error distance of BVAT (P<0.05), and more errors in the SRM (P<0.01), SWM (P<0.05), OWM (P<0.05), and ORcM tasks of the NBMT (P<0.05, Table 3) in contrast to the HC.
 |
Table 3 Results of Blue Velvet Arena Test and Nine Box Maze Test Assessment |
Levels of Serum Biomarkers
Within the CID group, the concentrations of SIRT1 and SIRT3 were considerably higher than those observed in the HC group (P < 0.001, Table 4).
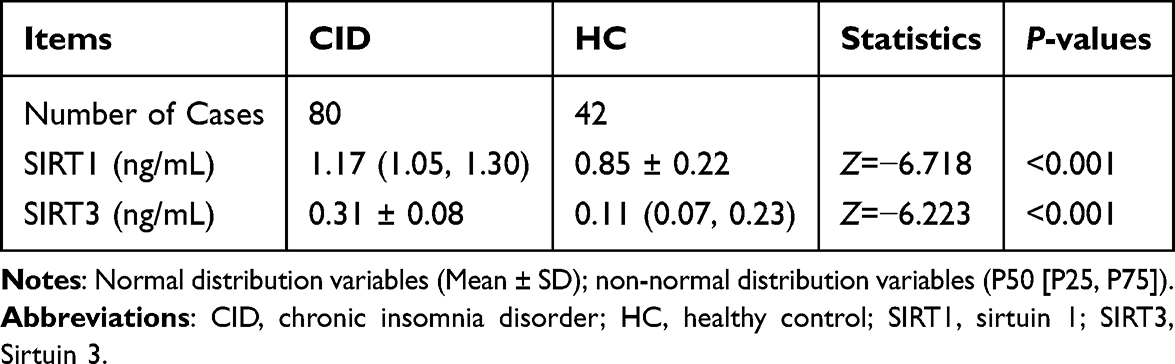 |
Table 4 Comparison of Serum Biomarkers Levels Between Two Groups |
Correlations Between Biomarkers and Sleep Quality and Illness Duration
Partial correlation analysis was conducted to evaluate the outcomes, with adjustments made for variables such as gender, age, education level, and PHQ-9 scores. The serum concentrations of SIRT1 and SIRT3 showed a positive correlation with PSQI scores (r=0.376, P < 0.001; r=0.441, P < 0.001), but not with the objective sleep parameters (P>0.05, Table 5). Serum SIRT1 level was marginally (r=−0.206, P=0.074), while SIRT3 level was significantly (r=−0.381, P=0.001) correlated with illness duration negatively.
 |
Table 5 Correlations of Biomarkers with Sleep Quality in the CID Group |
Correlations Between Biomarkers and Cognitive Function in CID
According to the identical partial correlation analysis, the serum concentrations of SIRT1 and SIRT3 were not significantly correlated with the total score but were negatively correlated with the delayed recall in the MoCA-C scale (r=−0.230, P=0.046; r=−0.297, P=0.009), with no significant correlation with other factors (P>0.05, Table 6). There was a positive correlation between SIRT1 and SIRT3 levels and NBMT’s SRM errors (r=0.299, P=0.009; r=0.250, P=0.029), and SIRT1 was positively correlated with ORcM errors (r=0.262, P=0.022; Table 6), whereas no significant correlation existed between SIRTs and the mean error distance in the BVAT.
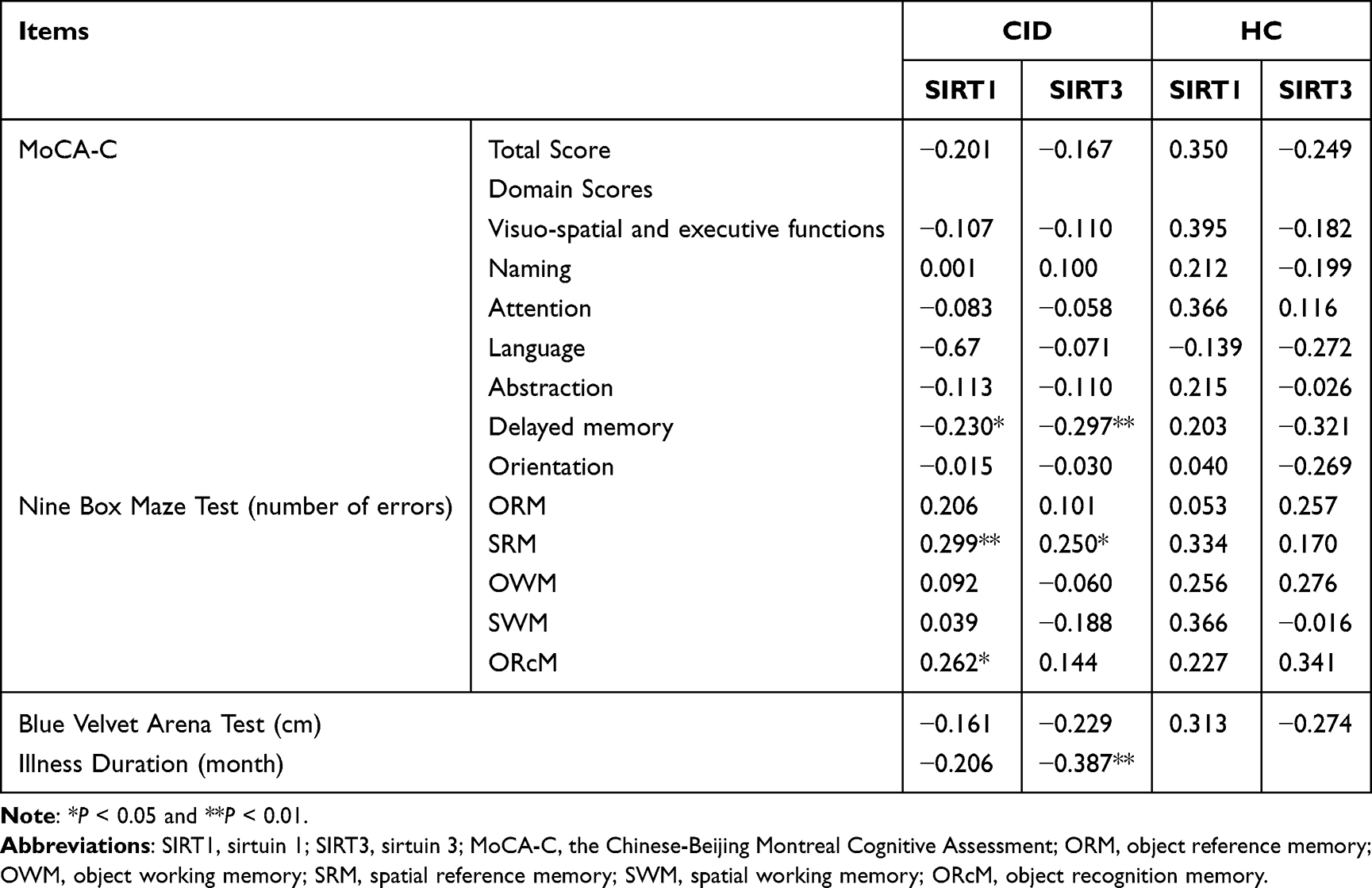 |
Table 6 Correlations of Biomarkers with Cognitive Function and Illness Duration in CID Group |
Diagnostic Value of Serum Biomarkers
Receiver operating characteristic (ROC) curves of SIRT1 and SIRT3 diagnosis of CID were plotted (Table 7). The AUC areas of SIRT1 and SIRT3 were 0.871 and 0.843, respectively, both of which had high sensitivity and specificity, suggesting that they were good predictors for distinguishing CID patients from healthy sleeper. The optimal cut-off values were 0.626 ng/mL and 0.698 ng/mL, respectively.
 |
Table 7 Characteristics of Potential Serum Biomarkers for CID in ROC Analysis |
Discussion
CID Patients Might Have Mitochondrial Dysfunction
Mitochondrial dysfunction is a fundamental factor in the development of many neurodegenerative conditions, such as Alzheimer’s, Parkinson’s, and Huntington’s diseases. Insomnia is a neurodegenerative disease closely related to circadian rhythm disturbances.41,42 Substantial evidence demonstrates that mitochondria play a crucial role in regulating sleep and wake cycles.10 According to related research, sleep disorders are often connected with primary mitochondrial diseases, and the clinical phenotype affects the type of sleep disorders associated with mitochondrial dysfunction.43 Currently, the majority of research investigating the relationship between sleep and mitochondrial dysfunction has been conducted in animal models, with limited studies available in human subjects. In rodent models, sleep deprivation which can result in oxidative stress, is associated with mitochondrial dysfunction.44 A European study involving human participants revealed that monozygotic twins with shorter sleep durations and diminished sleep efficiency exhibited a lower count of mitochondrial DNA copies.45 Therefore, mitochondrial markers SIRT1 and 3, which are closely related to neurons, were selected for this study.
SIRT1 and 3 are regulators of cell and mitochondrial activity with beneficial effects and are protective biomarkers. Mitochondria are key targets when it comes to oxidative stress. SIRT1 plays a role in boosting mitochondrial efficiency and alleviates oxidative stress. The process of autophagy triggered by oxidative stress via SIRT1 leads to less mitochondrial damage, which in turn results in reduced apoptosis and enhanced cell protection, indicating that autophagy initiated by SIRT1 plays a role in preserving mitochondrial function and ameliorating mitochondrial biogenesis, thus protecting the cells.46 In mitochondria, SIRT3, the primary deacetylase, performs the deacetylation activity and is dependent on NAD+. It plays a role in all stages of mitochondrial metabolism, including synthesis and mobility, which is essential for maintaining mitochondrial function and mitochondrial homeostasis.24 It can strengthen the mitochondrial defense mechanisms and decrease the damage caused by oxidative stress.47
Sleep is regulated by homeostasis and circadian processes that control all aspects of sleep behavior and related variables. In mammals, systemic circadian regulation is accomplished through central oscillators in the anterior hypothalamic suprachiasmatic nucleus (SCN).48 SIRT1 acts as a negative regulator by deacetylating BMAL1 and histones at circadian gene promoters to promote the repression of the inhibitory component of the oscillator.49 It plays a regulatory role in the expression of circadian clock genes. Change of SIRT1 may lead to circadian clock dysfunction, leading to the occurrence and development of age-related diseases.50 It has been found that SIRT1 appears to be central to the age-related decline in central circadian function.51 SIRT3 is the major mitochondrial deacetylase, and its activity is controlled by the circadian clock. The circadian clock regulates SIRT3 activity by controlling nicotinamide adenine dinucleotide concentration, which in turn regulates mitochondrial function.52 SIRT3 also affects locus coeruleus noradrenergic neurons (LCns), a type of arousal neurons that are highly sensitive to metabolic stress, by maintaining mitochondrial redox homeostasis. Therefore, LCns may be susceptible to the metabolic demands of prolonged wakefulness.53 Short-term sleep deprivation is a metabolic stressor for LCns, and mitochondria are involved in the homeostatic reaction to sleep insufficiency through SIRT3.
This study is the first to research the combination of serum markers SIRT1 and SIRT3 in CID patients. According to this study, individuals with CID showed significantly elevated serum SIRT1 and SIRT3 levels compared to HCs. This suggests that CID patients might experience a certain level of mitochondrial dysfunction. However, as protective biomarkers, elevated blood levels of SIRT 1 and SIRT 3 might also be a compensatory mechanism. We cannot rule out this possibility, and further exploration is required.
Mitochondrial Biomarkers is Associated with Insomnia Severity and Duration
Sleep serves a vital function in safeguarding and restoring the mitochondria. Studies on sleep deprivation and sleep apnea reveal that a significant benefit of sleep is its capacity to mitigate mitochondrial dysfunction and protect mtDNA from damage.54 Lack of sleep not only affects the shape of mitochondria but also reduces their quantity and leads to mitochondrial dysfunction. Additionally, this dysfunction is linked to the development and progression of various neurological diseases associated with sleep disorders.55
Currently, there is a lack of studies exploring the connection between serum SIRT1 and 3 levels and sleep quality in individuals with CID. Based on previous studies, we speculated that SIRT1 and SIRT3 reflect sleep status to a certain extent. In the present study, patients’ subjective and objective sleep quality was measured using the PSQI and PSG. Higher serum SIRT1 and SIRT3 levels were significantly related to increased overall PSQI scores, indicating poorer subjective sleep quality in patients. However, no statistically significant correlation was found with objective sleep quality. (as indicated by the PSG results). We also found that SIRT3 expression was significantly negatively correlated with the duration of disease. It can be inferred that the levels of serum SIRT1 and SIRT3 can reflect a certain extent of subjective severity of insomnia. However, their impact on objective sleep is unclear. This suggests that subjective and objective sleep arise from different neurophysiological processes. We conclude that a range of confounding factors could be involved in this process, which remains a promising avenue for future exploration.
Relationship Between Serum Mitochondrial Biomarkers Levels and Cognitive Impairment
Sirtuins are crucial in the processes of aging and neurodegenerative diseases, particularly SIRT1 and SIRT3, which are present in most areas of the brain. They contribute to the maintenance of higher-level brain function, and protect brain cells from adverse insults and neurodegeneration.27 Moreover, they are highly active in neurons and control the majority of vital brain processes. SIRT1 is mainly localized in the nucleus of hippocampal pyramidal and granule neurons, a key cognitive process structure that plays a key role in memory formation and maintenance of normal cognitive function.56 The absence of SIRT1 negatively affects cognitive functions, such as immediate, associative, and spatial memory, as well as classical conditioning. This indicates that SIRT1 plays a vital role in sustaining advanced brain function and synaptic plasticity. A research study involving animals discovered that the absence of SIRT1 in mice led to cognitive challenges, which were connected to problems with synaptic plasticity. Conversely, mice exhibiting elevated SIRT1 expression in their brains displayed normal synaptic plasticity and memory, indicating that SIRT1 is crucial for typical learning, memory, and synaptic plasticity in mice.22
SIRT3 is mainly active in neuronal mitochondria, and its expression in the brain is not limited to neurons, but also includes astrocytes and microglia. Studies have shown that SIRT3 expression is significantly decreased in the cortex and hippocampus of aging rats, suggesting that it plays an important role in aging-related brain diseases.57 It has been reported that overexpression of SIRT3 prevents neuronal degeneration, whereas loss of SIRT3 accelerates neurodegeneration in excitotoxic brain tissue.58 Neurodegeneration will lead to cognitive dysfunction, synaptic loss and impaired synaptic plasticity.59,60 A study has shown that SIRT3 regulation can restore the health of mitochondria and neurons in AD.61 Animal studies have shown that SIRT3 depletion leads to AD in mouse models.47 And SIRT3 deficient mice were shown to have reduced neuronal number, synaptic plasticity, and poor long-range memory, thus significantly increasing the neuronal vulnerability.26,62 The increased expression of SIRT3 contributes to the prolongation of neuronal lifespan.63 These suggest that SIRT3 is highly relevant to the regulation of cognitive function.
Our study revealed that SIRT1 and SIRT3 did not exhibit a significant connection with the overall MoCA-C score. Nonetheless, both SIRT1 and SIRT3 were found to have a negative association with the delayed recall component of MoCA-C, with SIRT3 demonstrating a notably strong negative correlation (P<0.001), indicating that the higher the concentration of SIRT1 and SIRT3, the worse the delayed recall ability of the patients. SIRT1 and SIRT3 may not be suitable for the evaluation of overall cognitive function, but can be used to reflect the impairment of memory function in patients, which may be related to the fact that CID patients often complain of memory decline. Besides these, SIRT1 was positively correlated with SRM (P<0.01) and ORcM, however, SIRT3 was only positively correlated with SRM. These findings indicate that shifts in the serum concentrations of SIRT1 and SIRT3 in patients with CID were linked to partial memory deficits, particularly affecting spatial memory to a certain extent. But there was no correlation between the average error distance of BVAT and SIRT1, SIRT3. We think this may be due to the fact that the detection property of BVAT is different from the MoCA-C. One of them represents a objective detection method, while another one is subjective. Or the number of samples collected by BVAT is too small to reflect the problem. It is worth further exploration in the future.
In view of the previous papers published by myself and our research group,64–66 this study has a certain novelty. This study sets the stage for further investigation to identify the fundamental mechanisms of chronic insomnia and opens an innovative pathway for future research.
Limitations
There are still some shortcomings in this study. First, as this study is cross-sectional, we are unable to establish a causal link between mitochondria-related markers and the pathogenesis of CID. Second, the sample size is quite small, and further expansion is needed to make the results more convincing. Third, due to the limitation of subjects’ compliance and economic conditions, the number of PSG and BVAT completed in this study was low, so there may be data bias in the statistical analysis of the results of these two experiments.
Conclusions
In CID patients, the serum levels of SIRT1 and SIRT3 markers were increased, suggesting that there was mitochondrial function damage, which was correlated with the severity of insomnia and cognitive dysfunction. The detection of mitochondrial dysfunction biomarkers is of great significance for the diagnosis of CID and evaluation of the degree of cognitive impairment.
Data Sharing Statement
The raw data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The study was approved by the Clinical Trial Ethics Committee in the Affiliated Chaohu Hospitals of Anhui Medical University (No: KYXM- 202108- 005). This study complied with the Declaration of Helsinki. All subjects and their families signed informed consent forms voluntarily.
Acknowledgments
Thanks to all the subjects who participated in this study, and to all research team members. Thanks to Dr. Zhang Ping for her great contribution to the ethical support of this study.
Author Contributions
All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Gao-Xia Liu: Writing – Original Draft, Writing – Review & Editing, Methodology, Investigation, Conceptualization. Yu Zhang: Writing – Review & Editing, Formal Analysis, Visualization. Yang Song: Writing – Review & Editing, Methodology. Shuai Yang: Writing – Review & Editing, Resources, Validation. Ai-Xi Su: Writing – Review & Editing, Data Curation. Yi-Zhou Feng: Writing – Original Draft, Software, Conceptualization. Yu-Shun Zhou: Writing – Original Draft, Formal Analysis, Visualization. Zi-Jie Ma: Writing – Original Draft, Methodology, Investigation. Lan Xia: Writing – Review & Editing, Investigation, Conceptualization. GuiHai Chen: Writing – Review & Editing, Data Curation, Software, Conceptualization.
Funding
This study was supported by the National Natural Science Foundation of China (grant number: 81671316).
Disclosure
The authors declare no conflicts of interest.
References
1. Dopheide JA. Insomnia overview: epidemiology, pathophysiology, diagnosis and monitoring, and nonpharmacologic therapy. Am J Manag Care. 2020;26(4 Suppl):S76–13. doi:10.37765/ajmc.2020.42769
2. Sutton EL. Insomnia. Ann Intern Med. 2021;174(3):ITC33–ITC48. doi:10.7326/AITC202103160
3. Sateia MJ. International classification of sleep disorders-third edition: highlights and modifications. Chest. 2014;146(5):1387–1394. doi:10.1378/chest.14-0970
4. Riemann D, Nissen C, Palagini L, et al. The neurobiology, investigation, and treatment of chronic insomnia. Lancet Neurol. 2015;14(5):547–558. doi:10.1016/S1474-4422(15)00021-6
5. Kay-Stacey M, Attarian H. Advances in the management of chronic insomnia. BMJ. 2016;354:i2123. doi:10.1136/bmj.i2123
6. Lejri I, Agapouda A, Grimm A, et al. Mitochondria- and oxidative stress-targeting substances in cognitive decline-related disorders: from molecular mechanisms to clinical evidence. Oxid Med Cell Longev. 2019;2019:9695412. doi:10.1155/2019/9695412
7. Brownlow JA, Miller KE, Gehrman PR. Insomnia and cognitive performance. Sleep Med Clin. 2020;15(1):71–76. doi:10.1016/j.jsmc.2019.10.002
8. Zhang P, Tan CW, Chen GH, et al. Patients with chronic insomnia disorder have increased serum levels of neurofilaments, neuron-specific enolase and S100B: does organic brain damage exist? Sleep Med. 2018;48:163–171. doi:10.1016/j.sleep.2017.12.012
9. Su AX, Ma ZJ, Li ZY, et al. Serum levels of neurotensin, pannexin-1, and sestrin-2 and the correlations with sleep quality or/and cognitive function in the patients with chronic insomnia disorder. Front Psychiatry. 2024;15:1360305. doi:10.3389/fpsyt.2024.1360305
10. Kempf A, Song SM, Talbot CB, Miesenbock G. A potassium channel beta-subunit couples mitochondrial electron transport to sleep. Nature. 2019;568:230–234. doi:10.1038/s41586-019-1034-5
11. Melhuish Beaupre LM, Brown GM, Braganza NA, et al. Mitochondria’s role in sleep: novel insights from sleep deprivation and restriction studies. World J Biol Psychiatry. 2022;23(1):1–13. doi:10.1080/15622975.2021.1907723
12. Picard M, Shirihai OS. Mitochondrial signal transduction. Cell Metab. 2022;34(11):1620–1653. doi:10.1016/j.cmet.2022.10.008
13. Todorova V, Blokland A. Mitochondria and synaptic plasticity in the mature and aging nervous system. Curr Neuropharmacol. 2017;15(1):166–173. doi:10.2174/1570159x14666160414111821
14. Khacho M, Harris R, Slack RS. Mitochondria as central regulators of neural stem cell fate and cognitive function. Nat Rev Neurosci. 2019;20(1):34–48. doi:10.1038/s41583-018-0091-3
15. Khacho M, Clark A, Svoboda DS, et al. Mitochondrial dynamics impacts stem cell identity and fate decisions by regulating a nuclear transcriptional program. Cell Stem Cell. 2016;19(2):232–247. doi:10.1016/j.stem.2016.04.015
16. Fattal O, Budur K, Vaughan AJ, Franco K. Review of the literature on major mental disorders in adult patients with mitochondrial diseases. Psychosomatics. 2006;47(1):1–7. doi:10.1176/appi.psy.47.1.1
17. Jung CM, Melanson EL, Frydendall EJ, Perreault L, Eckel RH, Wright KP. Energy expenditure during sleep, sleep deprivation and sleep following sleep deprivation in adult humans. J Physiol. 2011;589(Pt 1):235–244. doi:10.1113/jphysiol.2010.197517
18. Kukucka J, Wyllie T, Read J, Mahoney L, Suphioglu C. Human neuronal cells: epigenetic aspects. Biomol Concepts. 2013;4(4):319–333. doi:10.1515/bmc-2012-0053
19. Kumar V, Kundu S, Singh A, Singh S. Understanding the role of histone deacetylase and their inhibitors in neurodegenerative disorders: current targets and future perspective. Curr Neuropharmacol. 2022;20(1):158–178. doi:10.2174/1570159X19666210609160017
20. Sidorova-Darmos E, Wither RG, Shulyakova N, et al. Differential expression of sirtuin family members in the developing, adult, and aged rat brain. Front Aging Neurosci. 2014;6:333. doi:10.3389/fnagi.2014.00333
21. Donmez G, Outeiro TF. SIRT1 and SIRT2: emerging targets in neurodegeneration. EMBO Mol Med. 2013;5(3):344–352. doi:10.1002/emmm.201302451
22. Cai Y, Xu L, Xu H, Fan X. SIRT1 and neural cell fate determination. Mol Neurobiol. 2016;53(5):2815–2825. doi:10.1007/s12035-015-9158-6
23. Tang BL. Sirt1 and the mitochondria. Mol Cells. 2016;39(2):87–95. doi:10.14348/molcells.2016.2318
24. Ng F, Wijaya L, Tang BL. SIRT1 in the brain-connections with aging-associated disorders and lifespan. Front Cell Neurosci. 2015;9:64. doi:10.3389/fncel.2015.00064
25. Zhang J, Xiang H, Liu J, et al. Mitochondrial Sirtuin 3: new emerging biological function and therapeutic target. Theranostics. 2020;10(18):8315–8342. doi:10.7150/thno.45922
26. Vernucci E, Tomino C, Molinari F, et al. Mitophagy and oxidative stress in cancer and aging: focus on sirtuins and nanomaterials. Oxid Med Cell Longev. 2019;2019:6387357. doi:10.1155/2019/6387357
27. Kim SH, Lu HF, Alano CC. Neuronal Sirt3 protects against excitotoxic injury in mouse cortical neuron culture. PLoS One. 2011;6(3):e14731. doi:10.1371/journal.pone.0014731
28. Anamika KA, Acharjee P, Acharjee A, Trigun SK. Mitochondrial SIRT3 and neurodegenerative brain disorders. J Chem Neuroanat. 2019;95:43–53. doi:10.1016/j.jchemneu.2017.11.009
29. Manea L, Gilbody S, McMillan D. A diagnostic meta-analysis of the patient health questionnaire-9 (PHQ-9) algorithm scoring method as a screen for depression. Gen Hosp Psychiatry. 2015;37(1):67–75. doi:10.1016/j.genhosppsych.2014.09.009
30. Maier W, Buller R, Philipp M, Heuser I. The Hamilton anxiety scale: reliability, validity and sensitivity to change in anxiety and depressive disorders. J Affect Disord. 1988;14(1):61–68. doi:10.1016/0165-0327(88)90072-9
31. Backhaus J, Junghanns K, Broocks A, Riemann D, Hohagen F. Test-retest reliability and validity of the Pittsburgh Sleep Quality Index in primary insomnia. J Psychosom Res. 2002;53(3):737–740. doi:10.1016/s0022-3999(02)00330-6
32. Sancho-Domingo C, Carballo JL, Coloma-Carmona A, Buysse DJ. Brief version of the Pittsburgh Sleep Quality Index (B-PSQI) and measurement invariance across gender and age in a population-based sample. Psychol Assess. 2021;33(2):111–121. doi:10.1037/pas0000959
33. Buysse DJ, Reynolds CF, Monk TH, et al. The Pittsburgh sleep quality index: a new instrument for psychiatric practice and research. Psychiatry Res. 1989;28(2):193–213. doi:10.1016/0165-1781(89)90047-4
34. Tsai PS, Wang SY, Wang MY, et al. Psychometric evaluation of the Chinese version of the Pittsburgh Sleep Quality Index (CPSQI) in primary insomnia and control subjects. Qual Life Res. 2005;14(8):1943–1952. doi:10.1007/s11136-005-4346-x
35. Berry RB, Budhiraja R, Gottlieb DJ, et al. Rules for scoring respiratory events in sleep: update of the 2007 AASM manual for the scoring of sleep and associated events. J Clin Sleep Med. 2012;8:597–619. doi:10.5664/jcsm.2172
36. Lu J, Li D, Li F, et al. Montreal cognitive assessment in detecting cognitive impairment in Chinese elderly individuals: a population-based study. J Geriatr Psychiatry Neurol. 2011;24(4):184–190. doi:10.1177/0891988711422528
37. Nasreddine ZS, Phillips NA, Bédirian V, et al. The montreal cognitive assessment, MoCA: a brief screening tool for mild cognitive impairment. J Am Geriatr Soc. 2005;53(4):695–699. doi:10.1111/j.1532-5415.2005.53221.x
38. Chen GH, Xia L, Wang F, et al. Patients with chronic insomnia have selective impairments in memory that are modulated by cortisol. Psychophysiology. 2016;53(10):1567–1576. doi:10.1111/psyp.12700
39. He S, Chen XX, Ge W, et al. Are anti-inflammatory cytokines associated with cognitive impairment in patients with insomnia comorbid with depression? A pilot Study. Nat Sci Sleep. 2021;13:989–1000. doi:10.2147/NSS.S312272
40. Majerová V, Kalinčík T, Laczó J, et al. Disturbance of real space navigation in moderately advanced but not in early Huntington’s disease. J Neurol Sci. 2012;312(1–2):86–91. doi:10.1016/j.jns.2011.08.016
41. Xie Y, Tang Q, Chen G, et al. New insights into the circadian rhythm and its related diseases. Front Physiol. 2019;10:682. doi:10.3389/fphys.2019.00682
42. Lack LC, Micic G, Lovato N. Circadian aspects in the aetiology and pathophysiology of insomnia. J Sleep Res. 2023;32(6):e13976. doi:10.1111/jsr.13976
43. Brunetti V, Della Marca G, Servidei S, Primiano G. Sleep disorders in mitochondrial diseases. Curr Neurol Neurosci Rep. 2021;21(7):30. doi:10.1007/s11910-021-01121-2
44. Rodrigues NR, Macedo GE, Martins IK, et al. Short-term sleep deprivation with exposure to nocturnal light alters mitochondrial bioenergetics in Drosophila. Free Radic Biol Med. 2018;120:395–406. doi:10.1016/j.freeradbiomed.2018.04.549
45. Wrede JE, Mengel-From J, Buchwald D, et al. Mitochondrial DNA copy number in sleep duration discordant monozygotic twins. Sleep. 2015;38(10):1655–1658. doi:10.5665/sleep.5068
46. Ou X, Lee MR, Huang X, et al. SIRT1 positively regulates autophagy and mitochondria function in embryonic stem cells under oxidative stress. Stem Cells. 2014;32(5):1183–1194. doi:10.1002/stem.1641
47. Someya S, Yu W, Hallows WC, et al. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell. 2010;143:802–812.
48. Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010;330(6009):1349–1354. doi:10.1126/science.1195027
49. Asher G, Gatfield D, Stratmann M, et al. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell. 2008;134(2):317–328. doi:10.1016/j.cell.2008.06.050
50. Osum M, Serakinci N. Impact of circadian disruption on health; SIRT1 and Telomeres. DNA Repair. 2020;96:102993. doi:10.1016/j.dnarep.2020.102993
51. Chang HC, Guarente L. SIRT1 mediates central circadian control in the SCN by a mechanism that decays with aging. Cell. 2013;153(7):1448–1460. doi:10.1016/j.cell.2013.05.027
52. Peek CB, Affinati AH, Ramsey KM, et al. Circadian clock NAD+ cycle drives mitochondrial oxidative metabolism in mice. Science. 2013;342(6158):1243417. doi:10.1126/science.1243417.
53. Zhang J, Zhu Y, Zhan G, et al. Extended wakefulness: compromised metabolics in and degeneration of locus ceruleus neurons. J Neurosci. 2014;34(12):4418–4431. doi:10.1523/JNEUROSCI.5025-12.2014
54. Demine S, Renard P, Arnould T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 2019;8(8):795. doi:10.3390/cells8080795
55. Zhang W, Liu D, Yuan M, et al. The mechanisms of mitochondrial abnormalities that contribute to sleep disorders and related neurodegenerative diseases. Ageing Res Rev. 2024;97:102307. doi:10.1016/j.arr.2024.102307
56. Wei RM, Zhang YM, Zhang KX, et al. An enriched environment ameliorates maternal sleep deprivation-induced cognitive impairment in aged mice by improving mitochondrial function via the Sirt1/PGC-1α pathway. Aging. 2024;16(2):1128–1144. doi:10.18632/aging.205385
57. Braidy N, Poljak A, Grant R, et al. Differential expression of sirtuins in the aging rat brain. Front Cell Neurosci. 2015;9:167. doi:10.3389/fncel.2015.00167
58. Silberman DM. Metabolism, neurodegeneration and epigenetics: emerging role of Sirtuins. Neural Regen Res. 2018;13(3):417–418. doi:10.4103/1673-5374.228719
59. Chandramowlishwaran P, Vijay A, Abraham D, Li G, Mwangi SM, Srinivasan S. Role of sirtuins in modulating neurodegeneration of the enteric nervous system and central nervous system. Front Neurosci. 2020;14:614331. doi:10.3389/fnins.2020.614331
60. Wei RM, Zhang MY, Fang SK, et al. Melatonin attenuates intermittent hypoxia-induced cognitive impairment in aged mice: the role of inflammation and synaptic plasticity. Psychoneuroendocrinology. 2025;171:107210. doi:10.1016/j.psyneuen.2024.107210
61. Lloret A, Beal MF. PGC-1α, Sirtuins and PARPs in Huntington’s disease and other Neurodegenerative conditions: NAD+ to rule them all. Neurochem Res. 2019;44(10):2423–2434. doi:10.1007/s11064-019-02809-1
62. Dai SH, Chen T, Wang YH, et al. Sirt3 protects cortical neurons against oxidative stress via regulating mitochondrial Ca2+ and mitochondrial biogenesis. Int J Mol Sci. 2014;15(8):14591–14609. doi:10.3390/ijms150814591
63. Tao R, Coleman MC, Pennington JD, et al. Sirt3-mediated deacetylation of evolutionarily conserved lysine 122 regulates MnSOD activity in response to stress. Mol Cell. 2010;40(6):893–904. doi:10.1016/j.molcel.2010.12.013
64. Yang S, Kong X, Li X, et al. Serum C-reactive protein level and sleep characteristics in obstructive sleep apnea syndrome comorbid with panic disorder: a preliminary study. BMC Psychiatry. 2023;23(1):859. doi:10.1186/s12888-023-05376-6
65. Feng YZ, Chen JT, Hu ZY, et al. Effects of sleep reactivity on sleep macro-structure, orderliness, and cortisol after stress: a preliminary study in healthy young adults. Nat Sci Sleep. 2023;15:533–546. doi:10.2147/NSS.S415464
66. Yang S, Kong XY, Hu T, et al. Aquaporin-4, Connexin-30, and Connexin-43 as biomarkers for decreased objective sleep quality and/or cognition dysfunction in patients with chronic insomnia disorder. Front Psychiatry. 2022;13:856867. doi:10.3389/fpsyt.2022.856867
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.