Back to Journals » International Journal of Nanomedicine » Volume 21
cGAMP-Loaded M2e Nanovaccine Elicits Cross-Reactive Immunity and Mitigates H6N1 Avian Influenza Infection in Chickens
Authors Chen LY ![]() , Tsai HH
, Tsai HH ![]() , Peng L
, Peng L ![]() , Yao BY, Chiu J, Hu CMJ
, Yao BY, Chiu J, Hu CMJ ![]() , Chen HW
, Chen HW ![]()
Received 10 March 2026
Accepted for publication 29 May 2026
Published 15 June 2026 Volume 2026:21 607946
DOI https://doi.org/10.2147/IJN.S607946
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Kamakhya Prakash Misra
Li-Yuan Chen,1 Hsiao-Han Tsai,2 Lu Peng,1,3 Bing-Yu Yao,2 Jimmy Chiu,1 Che-Ming Jack Hu,2 Hui-Wen Chen1,3
1Department of Veterinary Medicine, National Taiwan University, Taipei, Taiwan; 2Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan; 3Animal Resource Center, National Taiwan University, Taipei, Taiwan
Correspondence: Hui-Wen Chen, Department of Veterinary Medicine, National Taiwan University, 1 sec 4 Roosevelt Road, Taipei, 10617, Taiwan, Email [email protected]
Introduction: Avian influenza virus (AIV) is a rapidly evolving zoonotic pathogen that causes significant economic loss in the poultry industry and threatens public health. In the development of a broadly protective influenza vaccine, the extracellular domain of matrix protein 2 (M2e) is a promising antigen candidate as it is highly conserved across most influenza A subtypes. M2e-induced immune protection is primarily mediated through antibody-dependent cellular cytotoxicity (ADCC), which enables the clearance of infected cells before viral release. However, the immunogenicity of M2e is limited due to its small size and low abundance on the viral surface, necessitating effective antigen delivery systems to enhance its presentation.
Methods: We developed a PLGA-based nanoshell vaccine co-encapsulating M2e peptide and a STING agonist and evaluated its immunogenicity and protective efficacy in specific-pathogen-free (SPF) chickens.
Results: Immunization of SPF chickens with the M2e nanoshell vaccine significantly enhanced immune-related gene expression and B-cell expansion, leading to a robust and durable M2e-specific antibody response. In addition, the induced antibodies demonstrated cross-reactivity by effectively recognizing the M2e protein expressed on MDCK cells infected with seven different strains of influenza viruses. Following H6N1 challenge, vaccinated chickens showed attenuated lesions in the kidneys and decreased viral load and viral antigen in the respiratory and intestinal lymphoid tissues, suggesting reduced viral replication. Furthermore, to explore potential Fc-mediated effector functions, an in vitro killing assay demonstrated that chicken splenic natural killer cells were capable of lysing influenza-infected target cells in the presence of serum from M2e nanoshell-immunized chickens, providing initial functional evidence of ADCC activity in the avian system.
Conclusion: Collectively, these results demonstrate that the M2e nanoshell vaccine elicits durable and cross-reactive immune responses and reduces tissue damage, potentially involving ADCC-mediated effector mechanisms. The M2e nanoshell vaccine shows potential as a broadly protective avian influenza vaccine and holds value for controlling AIV outbreaks in poultry. The image illustrates the mechanism of the M2e nanoshell vaccine in chickens. The vaccine consists of a nanoshell containing a STING agonist and M2e peptide, administered via injection. The vaccine activates dendritic cells, depicted as ’Activated DC’, through the STING pathway. These cells then stimulate the production of anti-M2e antibodies and T helper cells, labeled as ’Th1’. The antibodies bind to M2 proteins on infected cells, leading to antibody-dependent cellular cytotoxicity (ADCC) by natural killer cells. This process results in the lysis of infected cells, indicated by ’Infected cell lysis’. Various virus particles are shown, representing different stages of infection. The diagram highlights the immune response and cellular interactions involved in combating infection.Diagram of M2e nanoshell vaccine mechanism in chickens, showing activation of immune cells and infected cell lysis.
Keywords: avian influenza virus, M2e, STING agonist, nanoshell vaccine, antibody-dependent cellular cytotoxicity
Introduction
Avian influenza virus (AIV), classified as an influenza A virus within the family Orthomyxoviridae, is an enveloped, negative-sense RNA virus composed of eight gene segments encoding ten to eleven proteins.1 AIVs are classified into subtypes according to the antigenic properties of their two major surface glycoproteins, hemagglutinin (HA) and neuraminidase (NA), generating a large diversity of subtype combinations in nature. This extensive antigenic diversity poses a major challenge for conventional subtype-specific vaccine strategies and highlights the need for broadly protective vaccine approaches. Based on their pathogenicity in poultry, AIVs are further categorized into highly pathogenic avian influenza (HPAI) and low pathogenic avian influenza (LPAI). While LPAI (eg, H6 subtype) infections often cause mild clinical disease, HPAI strains, particularly H5 and H7 subtypes, can induce severe systemic illness and high mortality, resulting in substantial economic losses to the poultry industry.2 AIV is a zoonotic pathogen capable of crossing species barriers and infecting mammals. It mutates readily through antigenic drift, in which point mutations gradually accumulate in antigenic sites, and antigenic shift, which occurs when the viral RNA of different strains combines when co-infecting the same host. These mechanisms play a key role in the emergence of pandemic strains. Because of these characteristics, AIV remains a persistent threat to animal and global public health, highlighting the need for effective surveillance and control measures.3,4 Vaccination remains a cost-effective and practical approach to establishing immunity and minimizing mortality caused by AIV in poultry populations. Over the years, several vaccine platforms have been developed and applied to prevent and control AIV.5–7 Among them, inactivated vaccines are the most extensively used in the field and are typically produced by propagating the virus in chicken embryos and inactivated by chemical methods. These vaccines are safe and can induce strong humoral immunity, but they are subtype-specific and often require regular updates due to antigenic drift.5,8
M2e, a 23–amino acid peptide located on the N-terminus of the M2 protein, is highly conserved across influenza A subtypes and thus represents a promising target for universal influenza vaccine design.9 M2e can mediate protection primarily through Fc-dependent mechanisms such as antibody-dependent cellular cytotoxicity (ADCC), rather than through direct virus neutralization. ADCC is an essential immune mechanism in which effector cells recognize and kill target cells that are opsonized by specific antibodies. This mechanism demonstrates the ability to mediate cross-strain immunity and facilitate enhanced viral clearance, highlighting its potential in designing broadly protective vaccines.10 While ADCC has been extensively characterized in mammalian models of M2e-based vaccination, its presence and functional relevance in avian immune systems remain limited. However, despite its high conservation and cross-reactive potential, native M2e is intrinsically weakly immunogenic due to its short peptide length and limited abundance. Therefore, rational antigen delivery and adjuvant engineering are required to overcome these limitations and unlock its potential as a broadly protective vaccine target.11,12
Nanoparticle delivery systems, such as polymeric nanoparticles, virus-like particles, gold nanoparticles, and liposomes, have been widely explored in vaccine development due to their numerous advantages. These systems protect vaccine antigens from degradation, improve cellular uptake by antigen-presenting cells (APCs), and promote stronger humoral, cellular, and mucosal immune responses compared to soluble antigens alone.13,14 In particular, nanoshells typically comprise carboxyl-terminated poly (lactic-co-glycolic acid) (PLGA), a biodegradable and FDA-approved polymer recognized for its safety and excellent biocompatibility.15,16 PLGA forms negatively charged nanoparticles capable of encapsulating high-density antigens while protecting them from enzymatic degradation. This delivery strategy is especially advantageous for weakly immunogenic antigens, such as the conserved M2e of influenza A virus, which benefit from nanoshell-mediated protection and enhanced immunogenicity.17 This platform therefore provides a versatile nanodelivery strategy for co-presenting conserved antigens and immunostimulatory molecules to enhance vaccine efficacy.
Advancements in innate immunology have underscored the stimulator of interferon genes (STING) pathway as a critical mediator in the host’s early recognition of pathogens and an attractive target for vaccine adjuvant design.18,19 STING is a cytosolic adaptor protein that functions downstream of pattern recognition receptors (PRRs). It plays a pivotal role in sensing pathogen-associated molecular patterns (PAMPs) and triggering immune signaling cascades that link innate and adaptive immunity.20 STING activation triggers potent type I interferon responses and enhances dendritic cell activation, antigen cross-presentation, and subsequent adaptive immunity. STING has therefore emerged as a promising target for vaccine adjuvant development. Incorporating STING agonists into vaccine formulations has been shown to enhance dendritic cell maturation, promote antigen cross-presentation, and elicit robust CD8⁺ T cell responses.21–23 These properties make STING-based adjuvants especially valuable for vaccines against intracellular pathogens and subunit vaccines with poor inherent immunogenicity.
Building on our previous findings that the M2e nanoshell vaccine incorporating STING agonists conferred strong cross-subtype protection against influenza A virus infection in a murine model,17 this study further extends the investigation to an avian model. Given the continuous evolution of the avian influenza virus and the limitations of conventional vaccines, this study aimed to evaluate an M2e-based nanoshell vaccine incorporating STING agonists as adjuvants in chickens. In particular, we sought to explore whether M2e-induced antibodies in chickens could engage Fc-mediated effector mechanisms, including ADCC, which remain largely unexplored in avian systems.
Materials and Methods
Cell and Virus Cultures
The Madin-Darby Canine Kidney (MDCK) cell line (ATCC CCL-34) was used in this study. The MDCK cell line was purchased from Taiwan’s Bioresource Collection and Research Center. The cells were maintained in Dulbecco’s Modified Eagle medium (DMEM) (Gibco, Grand Island, NY) supplemented with 10% of fetal bovine serum (FBS) (Gibco) and 1% of penicillin/streptomycin/amphotericin B (PSA) (Sigma-Aldrich, St. Louis, MO) and incubated at 37°C in a humidified atmosphere containing 5% CO2.
Seven different influenza viruses were used in this study. They were propagated in chicken embryos and titrated by TCID50, hemagglutination (HA) test, or EID50 assays. A/Taiwan/1/2013×PR8 (rgH7N9) was obtained from the Centers for Disease Control, Ministry of Health and Welfare, Taiwan. A/Vietnam/1203/2004×PR8 (rgH5N1) was kindly provided by Dr. Hui-Ling Yen at Hong Kong University. A/Chicken/Taiwan/3937/2012 (H6N1) was a field isolate from chickens in Taiwan. A/California/7/2009 (pdmH1N1) was kindly provided by Dr. Li-Min Huang, National Taiwan University Hospital. A/Puerto Rico/8/1934 (PR8 H1N1) was kindly provided by Dr. Shin-Ru Shih, Chang Gung University. A/Aichi/2/1968 (H3N2) was kindly provided by Dr. Hung-Chih Yang, Department of Microbiology, National Taiwan University College of Medicine. A/Swine/Tainan/46-4/2005 (H1N2) was kindly provided by Dr. Ivan-Chen Cheng, Department of Veterinary Medicine, National Taiwan University.
Preparation of M2e Nanoshell Vaccine
The nanoshells were prepared by a water-in-oil-in-water (W/O/W) double emulsion process. Briefly, carboxyl-terminated PLGA (50:50 lactide: glycolide; Mw 7000–17,000; Sigma-Aldrich) was dissolved in ethyl acetate (75 mg/mL) to prepare the polymer solution. An aqueous phase containing M2e peptide (80 μg; GenScript; purity >95%; SLLTEVETPIRNEWGCRCNDSSD) and STING agonists cGAMP (20 μg, InvivoGen, San Diego, CA) was emulsified into the polymer solution using probe sonication (40% amplitude, 1 s on/2 s off, 1 minute, on ice). The resulting primary emulsion was added to 5 mL of 10 mM NaHCO3 buffer and subjected to a second sonication (30% amplitude, 2 minutes). Solvent evaporation was performed at 40°C, gently stirring. The nanoshells were collected using a 100 kDa molecular weight cut-off (MWCO) Amicon filter (Sigma-Aldrich) to remove unencapsulated materials.
Transmission Electron Microscopy (TEM) and Dynamic Light Scattering (DLS)
The images and size distribution of nanoshells were determined by transmission electron microscopy (TEM) and dynamic light scattering (DLS). TEM analysis was performed using a Jeol JEM-1400 Transmission Electron Microscope at the Joint Center for Instrument and Research, College of Bioresources and Agriculture, National Taiwan University. M2e nanoshell vaccine samples were absorbed onto carbon-coated copper grids for 15 seconds. Excess liquid was gently blotted with filter paper, followed by a brief rinse with RNase-free water to remove unbound materials. The grids were then negatively stained with 2% (w/v) phosphotungstic acid (PTA, pH 7.0) for 30 seconds. After staining, the excess stain was removed by blotting, and the grids were air-dried at room temperature before imaging. Nanoparticle size was measured by DLS with Malvern Zetasizer Nano ZS (Malvern Instrument) according to the manufacturer’s instructions.
Quantitative Peptide Assays
Peptide concentrations in M2e nanoshell vaccine were quantified using a colorimetric peptide assay (Pierce™ Quantitative Colorimetric Peptide Assay Kit, ThermoFisher Scientific, Waltham, MA). The M2e nanoshell vaccine was first mixed with an equal volume of acetone and incubated at 56°C for 10 minutes to release the encapsulated peptides. Subsequently, 20 μL of the sample and the M2e peptide standard were added to a 96-well flat-bottom microplate in duplicate. Then, 180 μL of Working Reagent (WR; ThermoFisher Scientific) was added and gently tapped to mix. The reaction mixture was incubated at 37°C for 15 minutes. The OD was measured at 480 nm using an automated plate reader. Peptide concentrations were determined by interpolation to the M2e peptide standards.
STING Reporter Cell Assays
293-Dual™ hSTING-R232 cells (InvivoGen) were used to evaluate STING pathway activation. Free cGAMP (10 μg), nanoshell NS (M2e+cGAMP) (10 μg cGAMP, 1.25 mg PLGA) were diluted in 20 μL of PBS. Each formulation was 2-fold serially diluted and added to 180 μL of culture medium containing 1×105 293-Dual™ hSTING-R232 cells per well in a 96-well plate. Treatments were performed in duplicate and incubated at 37°C with 5% CO2 for 48 hours. After incubation, 20 μL of culture supernatant was transferred to a new 96-well flat-bottom plate with 180 μL QUANTI-Blue™ solution (InvivoGen). The mixture was incubated at 37°C for 3 hours, and the optical density (OD) was measured at 655 nm using an automated plate reader (Synergy H1 Hybrid Multi-Mode Microplate Reader, BioTek, Winooski, VT). The OD655 values correspond to SEAP enzymatic activity and serve as an indirect measure of STING pathway activation.
PCR Array
To evaluate the immune responses induced by the M2e nanoshell vaccine, gene expression profiling was performed using the RT2 Profiler Chicken Innate & Adaptive Immune Responses PCR Array (Qiagen, Hilden, Germany). Briefly, two-week-old SPF chickens were randomly divided into three groups (n = 3 per group): (1) NS (M2e+cGAMP); (2) M2e+ISA; (3) unvaccinated control. Each group received a single dose of the corresponding vaccine formulation. Spleen tissues were collected at 7 days post-immunization (dpi). Total RNA was extracted from spleen tissues using a preloaded DNA/RNA extraction kit (GeneReach Biotechnology Corp., Taichung, Taiwan) according to the manufacturer’s instructions. Complementary DNA (cDNA) was synthesized from 400 ng of total RNA using the RT2 First Strand Kit (Qiagen) following the manufacturer’s protocol. Quantitative PCR was performed using the QuantiNova SYBR Green PCR Kit (Qiagen). Reaction mixtures were prepared according to the manufacturer’s protocol and loaded onto 384-well PCR array plates. Amplification was conducted using the CFX384 Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA) under the following cycling conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. Relative gene expression levels were calculated using the ΔΔCt method and normalized to the housekeeping genes included in the array.
3D Immunofluorescence Staining
Confocal 3D immunofluorescence imaging of spleen tissues was performed following a previously reported protocol with minor modifications.24 Briefly, freshly collected spleen tissues were fixed in 10% neutral-buffered formalin, embedded in paraffin, and sectioned into 150-μm-thick sections using a microtome. After deparaffinization, the sections were rehydrated through a graded ethanol series in PBS and subsequently permeabilized with 2% Triton X-100 (V900502, Sigma-Aldrich). The sections were incubated with anti-chicken Bu-1a/b antibody (1:800; AV20, Santa Cruz Biotechnology, Dallas, TX), followed by poly-HRP-conjugated goat anti-mouse IgG secondary antibody for 2 hours each. Alexa Fluor 488 tyramide reagent (ThermoFisher Scientific) was then applied for an additional 2 hours. Sections were subsequently incubated with DAPI (5 µg/mL; D9542, Sigma-Aldrich) and optical clearing reagent (Invitrogen) overnight at room temperature. Cleared sections were mounted for 3D image acquisition. Three-dimensional fluorescence images were acquired using a benchtop confocal microscope (BC43, Andor, Belfast, United Kingdom). Images were stitched using ImarisStitcher 9.9.0 software (Bitplane) and exported using Imaris 10.2.0 software (Bitplane).
Immunization Studies in Chickens
The animal experiments complied with institutional guidelines and were approved by the Institutional Animal Care and Use Committee (IACUC) of National Taiwan University, under approval number NTU-112-EL-00120. Chickens were housed at the Animal Resource Center, National Taiwan University. SPF chickens were randomly divided into three groups (n = 6 per group): (1) NS (M2e+cGAMP); (2) M2e+ISA; (3) unvaccinated control. For NS (M2e+cGAMP) vaccination, one-day-old SPF chickens (JD-SPF Biotech Co., Ltd., Miaoli, Taiwan) were immunized with 100 μL of nanoshell vaccine, which contains 80 μg of M2e peptides and 20 μg of cGAMP per dose. For ISA 71-adjuvanted vaccination (M2e+ISA), 80 μg of M2e peptides dissolved in NaHCO3 buffer was mixed with MONTANIDETM ISA 71 VG (Seppic, Paris, France) at a volume ratio of 3:7 (aqueous phase: oil phase) and emulsified by shaking at room temperature for 30 minutes before administration. SPF chickens were then immunized subcutaneously with the prepared emulsion. A three-dose vaccination regimen was administered with 3-week intervals between each immunization. Blood samples were collected at days 21, 42, 56, 77 and 98 post-immunization. Sera were centrifuged for 10 minutes at 3000 ×g and stored at −20°C.
Enzyme-Linked Immunosorbent Assay (ELISA)
96-well flat-bottom plates (Corning, Corning, NY) were coated with M2e peptide antigens (100 ng/well) in coating buffer (15 mM Na2CO3 (Sigma-Aldrich) and 35 mM NaHCO3 (YAKURI PURE CHEMICALS, Osaka, Japan) in ddH2O) at 4°C overnight. After washing with PBST (with 0.05% Tween80) three times, each well was blocked with a blocking buffer (5% skim milk in PBST) for one hour. Following washes, the test chicken sera were serially diluted and incubated for one hour at room temperature. After repeated washes, 100 μL of goat anti-chicken IgG HRP conjugate (Bethyl, Boston, MA) diluted 1:2000 in blocking buffer was added to each well and incubated for one hour at room temperature. After three further washes, 100 μL of KPL SureBlue ReserveTM TMB Microwell Peroxidase Substrate (SeraCare, Milford, MA) was added to each well for 10 minutes in the dark, followed by stopping the reaction with 100 μL of H2SO4. The result was read at OD 450 nm by an automated plate reader. M2e-specific titers were calculated on the endpoint titers of OD 0.2.
Immunocytochemical (ICC) Staining
MDCK cells were incubated in 8-well chamber slides (ThermoFisher Scientific) at a density of 1.5×104 cells per well for 24 hours. After washing with PBS, MDCK cells were infected with multiplicity of infection (MOI)=1 of H6N1, rgH7N9, H3N2, pdmH1N1, H1N1 PR8, or H1N2 virus or MOI=0.05 of rgH5N1 in infectious media. Following incubation for 24 hours, the infected MDCK cells were rinsed with PBS and then fixed in 10% formaldehyde at room temperature for 20 minutes. Fixed MDCK cells were blocked at room temperature with 100 μL 1% BSA-PBST for one hour, then the test chicken sera (1:50 dilution in 1% BSA-PBST) was added and incubated for one hour at room temperature. After incubation, the wells were washed with wash buffer (PBST, PBS with 0.1% Tween 20) three times (5 minutes for each) and incubated in 3% H2O2 in methanol for 15 minutes at room temperature to block endogenous peroxidase activity. After washing with wash buffer three times, each well received 200 μL of goat anti-chicken IgG conjugated to HRP (Jackson ImmunoResearch, West Grove, PA), diluted 1:400 in 1% BSA-PBST, and the slide was incubated at room temperature for 30 minutes. Following washing with wash buffer three times, a DAB Substrate kit (Dako, Carpinteria, CA) was used for visualization. The reaction time for visualization was determined under microscopic observation for each section. The slides were counterstained using Mayer’s hematoxylin (Sigma-Aldrich) and colored with carbonate-bicarbonate buffer (pH 9.4). Finally, the slides were dehydrated and covered with a coverslip.
Viral Challenge Studies in Chickens
SPF chickens were randomly divided into three groups (n = 6 per group): (1) NS (M2e+cGAMP), s.c.; (2) M2e + ISA, s.c.; (3) unvaccinated control. Each group received three doses of the corresponding vaccine formulation at 3-week intervals. Two weeks after the final immunization, all chickens were challenged intranasally with H6N1 influenza virus at 108.2 EID50 in 0.1 mL. On 7 days post-challenge (dpc), all chickens were humanely euthanized using CO2 inhalation at the end of the experiment following institutional animal care guidelines. Kidneys were then collected for histopathological examination using hematoxylin and eosin (H&E) staining. Tracheas and lungs were then collected for quantifying viral loads in tissues by RT-qPCR. Cecal tonsils were then collected for virus antigen IHC staining.
Histopathological Examination
Histopathological evaluation of kidney tissue sections stained with H&E staining was quantified and scored on a scale from 1 to 4 according to specific criteria. Scoring criteria were adapted and modified from previously published methods25,26 and tailored to assess pathological changes in the kidney. For kidney sections, scoring was determined by the degree of tubular degeneration, necrosis, and interstitial inflammation. Score 1: Minimal inflammation. Score 2: Multifocal tubular degeneration and mild inflammatory infiltration. Score 3: Widespread tubular necrosis with moderate interstitial inflammation. Score 4: Severe, diffuse tubular damage and necrosis with marked interstitial inflammation.
Virus RNA Extraction and RT-PCR
Total viral RNA from samples was extracted using a preloaded DNA/RNA extraction kit (GeneReach Biotechnology Corp.) following the manufacturer’s protocol. For reverse transcription, 5 µL of extracted RNA was mixed with 1 µL of 50 µM Uni12 primer, 1 µL of 10 µM dNTPs, and 5 µL of nuclease-free water (Ambion, Austin, TX) in a total volume of 12 µL. The mixture was heated at 65°C for 5 minutes and immediately placed on ice. Subsequently, 4 µL of 5X first-strand buffer, 2 µL of 0.1 M DTT, and 40 units of RNase inhibitor (Invitrogen) were added, followed by incubation at 37°C for 2 minutes. Reverse transcription was initiated by adding 200 units of M-MLV reverse transcriptase (Invitrogen), and the reaction was carried out at 25°C for 10 minutes, 37°C for 50 minutes, and finally at 72°C for 15 minutes to inactivate the enzyme.
Quantitative Real-Time PCR (qPCR)
qPCR was performed using the PrimeTime® Gene Expression Master Mix (Integrated DNA Technologies) according to the manufacturer’s instructions. Each 20 µL reaction contained 10 µL of 2X Master Mix, 1 µL of 4 µM FAM-labeled probe, 1 µL each of M gene-specific forward and reverse primers, 5 µL of RNase-free water, and 2 µL of cDNA template.27 A plasmid DNA standard curve (102 to 109 copies) of the H1N1 M gene was included for quantification. Reactions were run on a CFX Connect Real-Time PCR Detection System (Bio-Rad) with the following cycling conditions: 95°C for 3 minutes, followed by 45 cycles of 95°C for 15 seconds and 60°C for 1 minute.
Immunohistochemical (IHC) Staining
Cecal tonsils from chickens were collected and fixed with 10% formalin. The tissues were embedded in paraffin, sectioned, and subsequently deparaffinized using xylene, followed by rehydration through a graded ethanol series. Heat-induced antigen retrieval was then performed using antigen unmasking solution (Vector Laboratories, Burlingame, CA) under high pressure for 20 minutes. After washing with wash buffer three times for 1 minute each, endogenous peroxidase activity was blocked with 3% hydrogen peroxide in methanol for 10 minutes at room temperature and washed three times. Subsequently, non-specific binding was blocked by incubation with 1% BSA in PBST for 1 hour at room temperature. The sections were then incubated with the primary antibody (NS1 monoclonal antibody, 1:1000 dilution) for 1 hour at room temperature. After washing, the sections were incubated with the secondary antibody (Dako REAL EnVision HRP, rabbit-mouse) for 1 hour at room temperature. After three additional washes, immunoreactivity was visualized using a DAB Substrate Kit (Dako). The reaction time was determined by microscopic observation for each section. The slides were subsequently counterstained with Gill II hematoxylin (Epredia, Kalamazoo, MI), blued with Dako bluing buffer, and covered with a coverslip.
Preparation of Chicken NK Cells
Spleens were collected from three 9-week-old SPF chickens and were pooled for natural killer (NK) cell isolation. Briefly, spleens were passed through a 70 µm cell strainer (Corning) and were washed by adding cold PBS up to 50 mL, followed by centrifugation at 500 ×g for 10 minutes at 4°C. Pelleted cells were resuspended in 3 mL of cold PBS and gently layered onto an equal volume of Histopaque-1119 (Sigma-Aldrich). The gradient was centrifuged at 1000 ×g for 30 minutes at 4°C. Splenocytes at the interface were carefully collected and washed once with five volumes of PBS. Fluorescence-activated cell sorting (FACS) was performed to isolate CD3−CD8α⁺ NK cell populations for NK cell isolation.28 Prior to staining, cells were blocked with 2% heat-inactivated chicken serum in PBS for 30 minutes at 4°C. After centrifugation at 1500 rpm for 5 minutes, cells were washed with FACS buffer (1X PBS supplemented with 2% FBS and 1% PSA) and centrifuged again. Cells were stained with 0.5 µg of anti-chicken CD3-PACBLU (SouthernBiotech, Birmingham, AL) and 0.05 µg of anti-chicken CD8α-APC (SouthernBiotech) antibodies per 106 cells at 4°C for 1 hour. After staining, cells were washed with FACS buffer, centrifuged at 1500 rpm for 5 minutes, and passed through a 40 µm cell strainer (Corning) before sorting. CD3−CD8α⁺ NK cells were gated and collected using a BD FACSAria™ III Cell Sorter (BD Biosciences, San Jose, CA). Flow cytometric data were analyzed using FlowJo software (version 10.8.1; Tree Star Inc., Ashland, OR).
In vitro Killing Assay
An in vitro killing assay was performed to evaluate ADCC activity. MDCK cells (7 × 103 cells/well) were seeded into 96-well flat-bottom plates and incubated overnight. The culture medium was removed the following day, and the cells were gently washed once with 100 μL of sterile 1X PBS. Cells were then infected with recombinant rgH5N1 influenza virus at an MOI of 0.05 in infection medium and incubated at 37°C for 24 hours. After infection, the isolated chicken NK cells were co-cultured with the infected MDCK cells at an effector-to-target (E: T) ratio of 10:1 in the presence of heat-inactivated serum from vaccinated chickens. Each experimental condition was tested in triplicate. Following a 24-hour incubation at 37°C with 5% CO2, cell viability was measured using a CCK-8 assay (Dojindo, Japan) according to the manufacturer’s instructions. The OD at 450 nm and 650 nm was read using an automated plate reader. A reduction in cell viability indicated NK cell-mediated lysis of infected MDCK cells, facilitated by vaccine-induced antibodies.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism version 10.1.2 (GraphPad Software, San Diego, CA). For normally distributed datasets, differences between groups were analyzed using one-way analysis of variance (ANOVA) followed by either Dunnett’s or Tukey’s multiple comparisons test, depending on the experimental design. Unpaired t-tests were used for datasets involving two independent groups. For non-normally distributed data or ordinal scores (eg, histopathological lesion scores), the Kruskal–Wallis test was applied, followed by Dunn’s multiple comparisons test. Data are presented as mean ± standard error of the mean (SEM) for parametric data. A p-value of less than 0.05 was considered statistically significant. The specific statistical tests used for each experiment are indicated in the corresponding figure legends.
Results
Preparation and Characterization of the M2e Nanoshell Vaccine
M2e nanoshell vaccines were prepared using a double emulsion solvent evaporation method as previously described.17 Transmission electron microscopy revealed uniform spherical particles (Figure 1A). DLS analysis showed an average diameter of 143.6 nm with a low polydispersity index (PdI = 0.087), confirming narrow size distribution and good formulation quality (Figure 1B). The encapsulation efficiencies of M2e peptide were quantified in the nanoshell formulations. NS (M2e+cGAMP) exhibited an encapsulation efficiency of approximately 70% for M2e peptide (Figure 1C), demonstrating efficient antigen loading within the nanoshells.
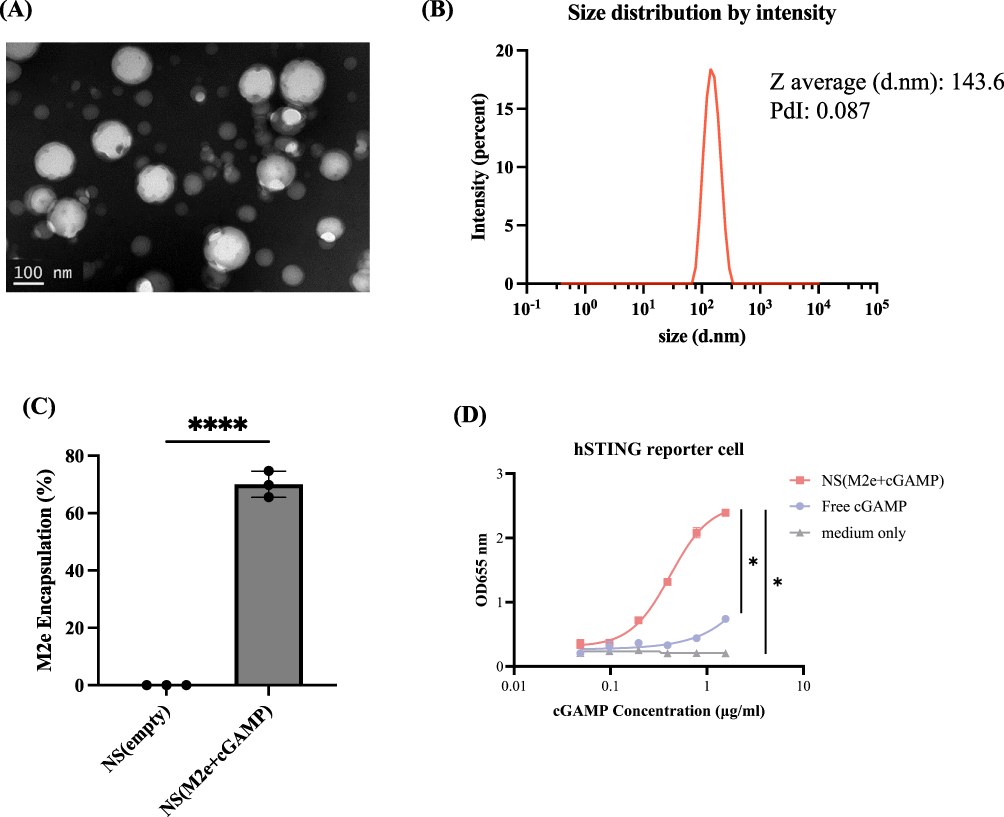 |
Figure 1 Characterization of M2e nanoshell vaccine. (A) The M2e nanoshell vaccine was imaged using TEM under 2% PTA negative staining. (Scale bar = 100 nm) (B) The size of the M2e nanoshell vaccine was measured using DLS and is presented in diameter in nanometers (d.nm). (C) M2e peptide encapsulation efficiency in nanoshells was determined by interpolation to the M2e peptide standards. All measurements were performed in triplicate (n = 3), and data are presented as mean ± SEM. Statistical analysis was performed using unpaired t-tests. (D) Assessment of STING activation curve of NS (M2e+cGAMP), free cGAMP, and medium only. 293-Dual™ hSTING-R232 cells were incubated with the indicated formulations for 48 hours. Dose–response curves were generated based on 2-fold serial dilutions, and SEAP activity in the culture supernatant was measured at 655 nm as an indicator of STING pathway activation. Data are presented as mean ± SEM, and statistical analysis was performed using ordinary one-way ANOVA followed by Tukey’s multiple comparisons test. (*p < 0.05; ****p < 0.0001). |
To assess the bioactivity of the STING agonist, both free cGAMP and NS (M2e+cGAMP) formulations were evaluated. The nanoshell formulation markedly enhanced STING pathway activation compared to free cGAMP. At the same concentration of 1.56 μg/mL, NS (M2e+cGAMP) induced a substantially higher response (OD655 = 2.394) than free cGAMP (OD655 = 0.740), indicating that nanoshell encapsulation promotes more efficient intracellular delivery and utilization of the STING agonist (Figure 1D). These findings suggest that the nanoshell serves as an effective carrier system to potentiate STING-mediated immune activation.
M2e Nanoshell Vaccination Activates Antiviral and Th1-Associated Gene Expression
The overall experimental design for evaluating vaccine-induced immune responses is illustrated in Figure 2A. A PCR array targeting 84 innate and adaptive immune–related genes was performed on spleen samples collected from chickens immunized with M2e+ISA, NS (M2e+cGAMP), or left unvaccinated. As shown in Figure 2B, global gene expression profiling revealed that the NS (M2e+cGAMP) group exhibited a broader and markedly higher magnitude of gene upregulation compared with M2e+ISA, when normalized to the unvaccinated controls. To further dissect the immune pathways involved, genes were categorized according to their functional relevance. Notably, in the NS (M2e+cGAMP) group, antiviral defense genes such as TLR7, STAT3, IRF7, MX1, and IFIH1 were markedly upregulated (Figure 2C), indicating robust activation of interferon-mediated antiviral signaling, which is essential for protection against viral challenge. At the same time, Th1-associated genes including STAT4, NFKBIA, IRF1, and CD40LG were significantly enhanced (Figure 2D). The Th1-polarized immune signature is particularly relevant given that ADCC, one of the major effector functions mediated by M2e-specific antibodies, relies on Th1-driven cytokine and effector cell activation. Together, these data demonstrate that co-delivery of M2e peptide and cGAMP within PLGA nanoshells amplifies both innate antiviral and Th1-type immune pathways, establishing a mechanistic foundation for the observed cross-reactive and cytotoxic immune responses elicited by the nanoshell vaccine.
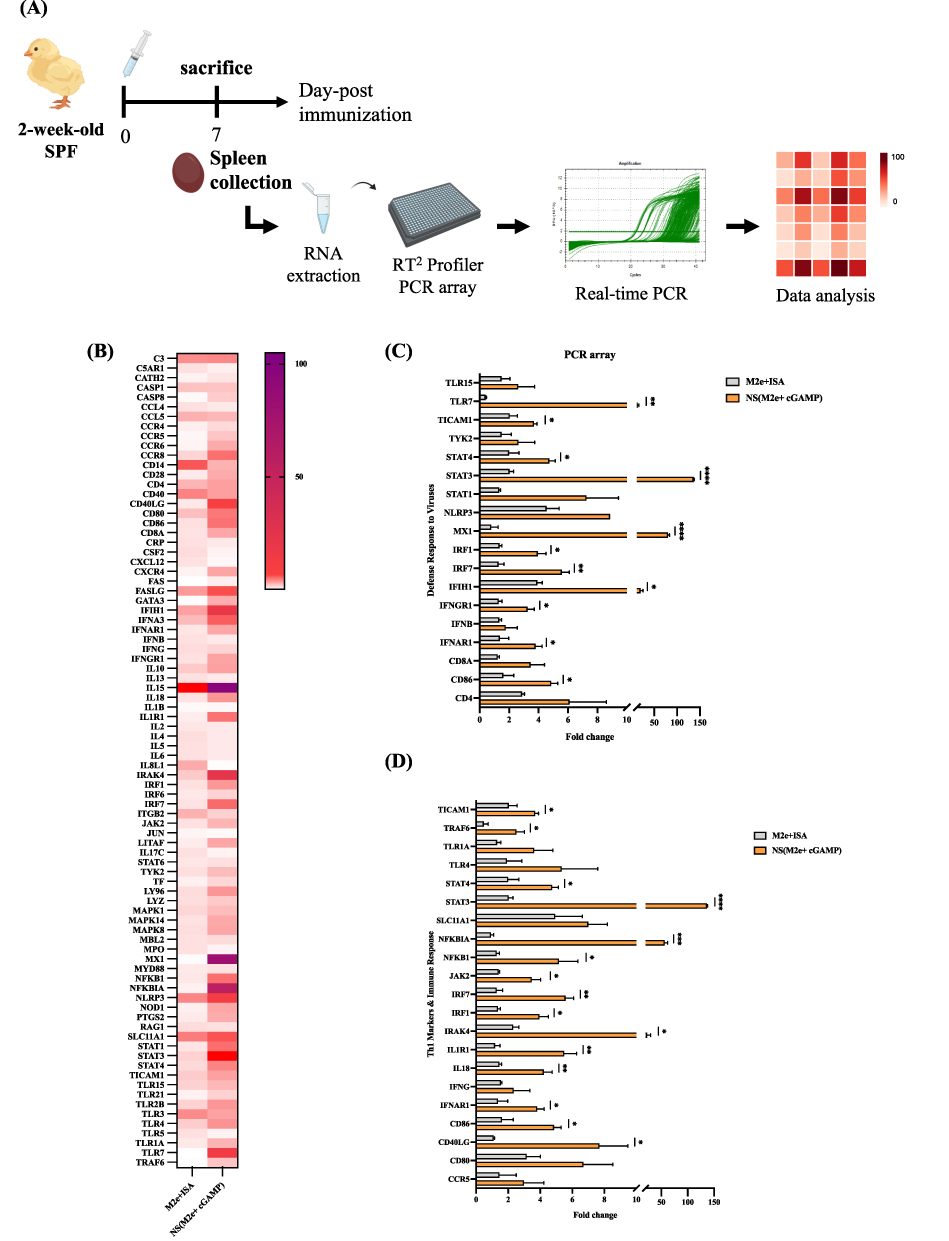 |
Figure 2 Transcriptomic profiling of immune-related genes in chicken spleens following M2e nanoshell vaccination. (A) Schematic illustration of experimental workflow for the PCR array analysis. Two-week-old chickens (n = 3 per group) were immunized with a single dose of NS (M2e+cGAMP), M2e+ISA, or remained unvaccinated, and spleens were collected for gene profiling. (B) Heatmap showing the relative fold change of 84 immune-related genes compared with the untreated control. (C) The relative fold change of the defense response to the virus genes. (D) The relative fold change of Th1-markers and immune response-related genes. Data are presented as mean ± SEM, and statistical analysis was performed using unpaired t-tests. (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001). |
M2e Nanoshell Vaccination Enhances Splenic B-Cell Responses
The overall experimental workflow for 3D immunofluorescence imaging of splenic architecture is illustrated in Figure 3A. In spleen tissue sections, chickens vaccinated with NS (M2e+cGAMP) exhibited markedly stronger and more widespread Bu-1a/b–positive B-cell signals compared with those immunized with M2e + ISA or left unvaccinated (Figure 3B). Quantitative analysis revealed that B-cell–positive regions accounted for 24.7% in the nanoshell-vaccinated group, 19.1% in the M2e+ISA group, and 7.1% in unvaccinated controls (Figure 3C). These results indicate that M2e nanoshell vaccination effectively stimulates B-cell expansion and lymphoid follicle development, further supporting that the M2e nanoshell vaccine provides a structural basis for the heightened antibody titers and functional immune protection observed in subsequent assays.
 |
Figure 3 B-cell response detected in different vaccinated chicken tissues by 3D immunofluorescence staining. (A) Schematic workflow of the 3D immunofluorescence staining. (B) Representative spleen sections from chickens immunized with NS (M2e+cGAMP), M2e+ISA, or unvaccinated groups, showing Bu-1a/b–positive B-cell signals (green). Nuclei were counterstained with DAPI (blue). (C) Quantification of B-cell–positive areas in spleen sections, expressed as a percentage of the total tissue area (n = 1 per group). |
M2e Nanoshell Vaccination Elicits Robust and Cross-Reactive M2e-Specific Antibody Responses
The immunogenicity of the M2e nanoshell vaccine was evaluated by quantifying M2e-specific IgG titers in chicken sera across a three-dose immunization schedule (Figure 4A). As shown in Figure 4B, NS (M2e+cGAMP) vaccination elicited strong IgG titers after the second immunization, with mean IgG titers reaching 1.5 log10, and approaching 2.0 log10 after the third immunization. These titers were substantially higher than those induced by the M2e+ISA group, which remained at 0.4 log10, demonstrating stronger immunogenicity of the NS (M2e+cGAMP) formulation compared with the M2e+ISA group. To further evaluate antibody cross-reactivity, serum from NS (M2e+cGAMP)-immunized chickens were applied to MDCK cells infected with seven different influenza A virus strains (Figure 4C). The antibodies showed no reactivity toward uninfected cells but exhibited distinct binding signals in infected cells across all tested subtypes, including H1N1 PR8, pdmH1N1, H1N2, H3N2, rgH5N1, H6N1 and rgH7N9, indicating that the induced antibodies have cross-reactive potential (Figure 4D). These findings suggest that vaccine-induced antibodies are capable of recognizing M2e expressed by diverse influenza virus subtypes in its native context. These results demonstrate that M2e nanoshell vaccine markedly enhances both the magnitude and persistence of the chicken immune response, while simultaneously broadening antigen recognition. This highlights the potential of the M2e nanoshell vaccine as a universal influenza vaccine platform capable of eliciting durable and cross-subtype protective immunity.
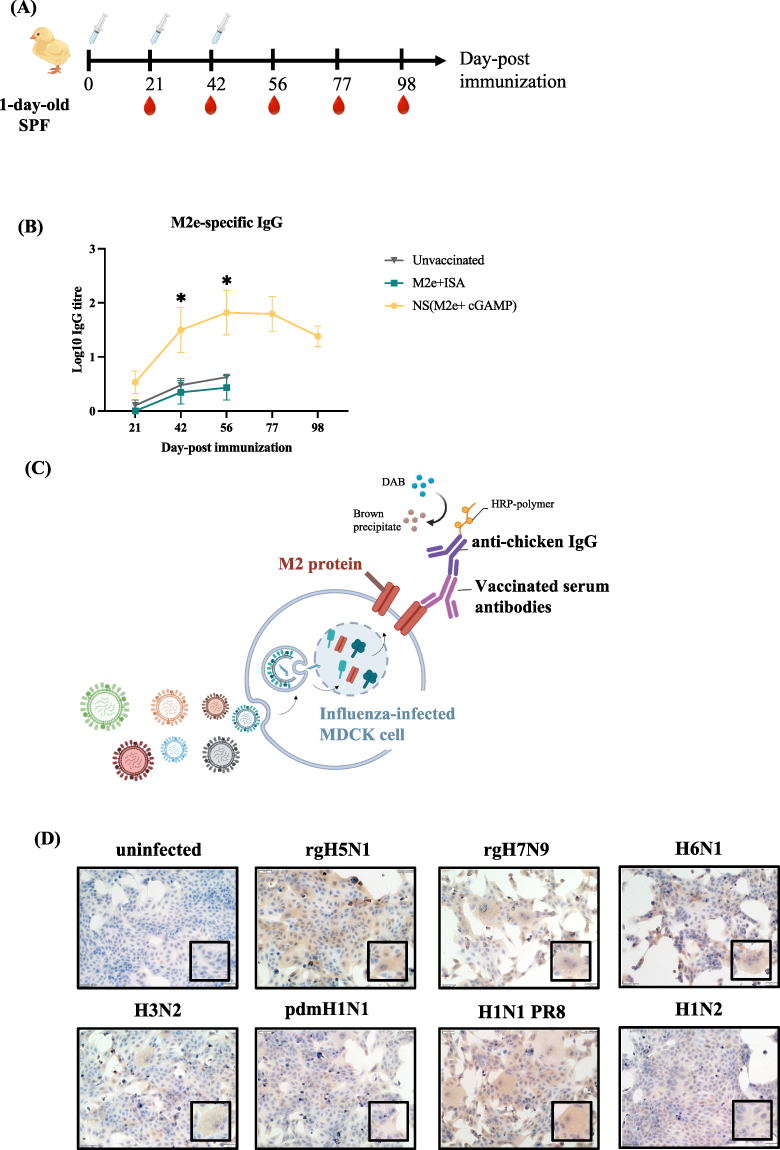 |
Figure 4 M2e-specific antibody responses and cross-type reactivity induced by the M2e nanoshell vaccine. (A) Schematic diagram of the immunization schedule. SPF chickens were divided into three groups (n = 6 per group) and subcutaneously immunized on days 0, 21, and 42 with NS (M2e+cGAMP), M2e formulated with ISA 71 VG (oil-based adjuvant), or left unvaccinated. Blood samples were collected on days 21, 42, 56, 77, and 98 for serological analysis. (B) Detection of M2e-specific IgG titers in sera collected at multiple time points after immunization. M2e-specific IgG levels were determined by ELISA, and endpoint titers were defined as the highest serum dilution yielding an OD450 value of 0.2. Statistical analysis was performed using one-way ANOVA followed by Dunnett’s multiple comparisons test at each time point to compare antibody titers among the three groups. Data are expressed as mean ± SEM. (*p < 0.05). (C) Schematic illustration of cross-reactivity ICC testing. Sera from vaccinated chickens were incubated with MDCK cells infected with seven influenza A strains (rgH5N1, H6N1, rgH7N9, H1N1 PR8, H1N2, pdmH1N1, and H3N2) to evaluate antibody recognition of influenza-infected cells expressing M2e. (D) Cells infected with different viruses were fixed and incubated with sera collected from chickens immunized subcutaneously with a single dose of NS (M2e+cGAMP), followed by staining with anti-chicken IgG and DAB chromogenic detection. Uninfected MDCK cells served as negative controls. Positive brown staining indicates specific binding of M2e-induced antibodies to influenza-infected cells. (Scale bar = 50 µm). |
M2e Nanoshell Vaccination Protects Chickens Against H6N1 Challenge
The protective potential of the M2e nanoshell vaccine was evaluated in SPF chickens challenged with H6N1 influenza virus two weeks after the final immunization (Figure 5A). Previous studies have reported renal lesions and urate deposition in chickens infected with H6N1 AIV,29 thus, the kidney was selected as a representative target organ, as it is a major site of AIV–induced pathology in chickens. At 7 dpc, kidneys were collected for gross and histopathological evaluation. Gross examination revealed that unvaccinated control and M2e+ISA chickens developed marked renal lesions, including swelling and extensive urate deposition on the renal surface, indicative of severe virus-induced nephritis. In contrast, chickens immunized with the NS (M2e+cGAMP) vaccine displayed kidneys of normal size, color, and texture, without visible urate accumulation or tissue damage (Figure 5B). Histopathological analysis of H&E–stained sections revealed inflammatory infiltration and tubular epithelial swelling in the kidneys of unvaccinated chickens. In vaccinated groups, renal architecture was largely preserved, with minimal inflammatory infiltration and intact tubular morphology (Figure 5C). Consistently, the proportion of chickens showing kidney lesions was lower in the NS (M2e+cGAMP) group, where only 2 out of 6 chickens exhibited gross renal pathology compared to 4/6 in the ISA-adjuvanted group, and 5/6 in the unvaccinated group (Figure 5D). To quantitatively assess renal microscopic damage, kidney sections were scored on a scale of 1 to 4 based on the severity of tubular degeneration, necrosis, and interstitial inflammation, where 1 = normal to minimal changes and 4 = severe and widespread pathology. The NS (M2e+cGAMP) group exhibited lower renal lesion scores compared to other two groups (Figure 5E), suggesting that the nanoshell vaccine mitigated H6N1-induced renal pathology.
 |
Figure 5 Protective efficacy of M2e nanoshell vaccine in H6N1-infected chickens. (A) The schedule of immunization and challenge in chickens. Three doses of vaccine were administered to SPF chickens at three-week intervals. Chickens (n = 6 per group) were intranasally challenged two weeks after the final vaccination with 108.2 EID50 of Chicken/Taiwan/3937/2012 (H6N1) virus. At 7 dpc, chickens were euthanized by CO2 inhalation, and tissues were collected for virus load detection and histological evaluation. (B) Representative gross pathology of kidneys collected from each group. Images were taken immediately after necropsy. (C) Representative H&E staining of kidney tissue sections collected from each group (Scale bar = 100 µm). (D) Summary of gross kidney lesions across different treatment groups. Data are shown as the number of chickens with or without visible gross kidney lesions. (E) Histopathological lesion scores of kidney tissues in each group, evaluated based on tubular degeneration, necrosis, and interstitial inflammation. Scores range from 1 (minimal) to 4 (severe). Statistical analysis was performed using the Kruskal–Wallis and Dunn’s multiple comparisons tests. (*p < 0.05). (F) Viral load in the trachea and lungs. Quantitative RT-PCR was performed on homogenized samples, and viral load was normalized and expressed as viral RNA copies per gram of tissue. Data are shown as mean ± SEM. (G) IHC staining of NS1 protein in cecal tonsils (Scale bar = 100 µm). |
To further assess viral burden in respiratory tissues, lung and trachea samples were collected at 7 dpc for viral RNA quantification by RT-qPCR. Compared with the unvaccinated and ISA-adjuvanted groups, a trend toward reduced viral RNA levels was detected in chickens immunized with the M2e nanoshell vaccine, although the difference did not reach statistical significance (Figure 5F). To further evaluate viral replication at the tissue level, IHC staining was performed on cecal tonsil sections collected at 7 dpc using an NS1-specific monoclonal antibody. Strong NS1 signals were observed in the cecal tonsils of unvaccinated and ISA-adjuvanted chickens, indicating active viral infection. In contrast, NS1 staining was reduced in chickens immunized with the NS (M2e+cGAMP) vaccine (Figure 5G), suggesting decreased viral presence in the vaccinated group. Collectively, these findings suggest that the M2e nanoshell vaccine is associated with reduced renal pathology and decreased viral burden, indicating protective potential against H6N1 infection in this experimental model.
M2e Nanoshell Vaccination Elicits NK Cell-Mediated ADCC
Avian NK cells are recognized as a unique population of innate lymphoid cells that express CD8α homodimers on the surface but lack T- or B-cell-specific markers, such as CD3. Based on this immunophenotypic profile, chicken NK cells are commonly identified as CD3−CD8α⁺ cells. In this study, splenocytes were harvested from SPF chickens and isolated via density gradient centrifugation. The cells were stained with fluorescently labeled antibodies against CD3 and CD8α and analyzed using Fluorescence-activated cell sorting. As shown in Figure 6A, lymphocytes were first gated based on forward and side scatter properties (FSC/SSC). Within the lymphocyte gate, NK cells were defined as CD3− and CD8α⁺. The proportion of CD3−CD8α⁺ cells accounted for approximately 1.06% of total splenocytes. These sorted NK cells were subsequently used in downstream in vitro ADCC assays, where their cytotoxic potential was assessed in the presence of vaccine-induced antibodies and influenza-infected target cells.
 |
Figure 6 Isolation of chicken NK cells and ADCC activity induced by vaccinated serum. (A) Flow cytometric analysis for isolation of chicken NK cells. Splenocytes isolated from three SPF chickens were pooled and stained with anti-chicken CD3 and CD8α antibodies. NK cells were defined as CD3−CD8α⁺ and gated based on FSC/SSC profiles. (B) Schematic illustration of the in vitro killing assay setup. MDCK cells infected with rgH5N1 (MOI=0.05) were co-incubated with chicken NK cells and vaccinated serum. After 24 h, cell viability was measured using a CCK-8 assay to determine the killing rate. (C) Cell-killing rate was assessed using the CCK-8 assay. Data are shown as mean ± SEM. Statistical analysis was performed using ordinary one-way ANOVA followed by Tukey’s multiple comparisons test. (***p < 0.001; ****p < 0.0001). Representative cell images of infected MDCK cells (D) alone, (E) treated with vaccinated chicken serum, (F) treated with NK cells, and (G) treated with both NK cells and vaccinated chicken serum. Arrows indicate representative regions of reduced cell density and cell detachment, consistent with cytotoxic effects. |
In vitro killing assays were performed to investigate whether antibodies elicited by the M2e nanoshell vaccine could mediate ADCC. In this assay, MDCK cells were experimentally infected with the influenza virus under controlled conditions before co-culture, ensuring that target cells were uniformly infected for subsequent ADCC evaluation. A schematic illustration of the in vitro killing assays is shown in Figure 6B. As shown in Figure 6C, the group that received both vaccinated serum and NK cells exhibited an approximately 32% MDCK cell killing rate, which was significantly higher than that observed in the control group (0%, normalized), or in groups treated with NK cells alone (13%) or serum alone (4%). These findings indicate that antibodies generated by the M2e nanoshell vaccine can bind to virus-infected cells and effectively recruit NK cells for targeted lysis. Representative images of influenza-infected cells under different conditions are shown in Figure 6D–G. Cells in the control group (Figure 6D), which were infected but not treated with either chicken antibodies or NK cells, largely maintained normal morphology. Infected cells incubated with antibodies alone (Figure 6E) or NK cells alone (Figure 6F) showed moderate cytopathic changes and partial cell loss. In contrast, infected cells treated with both antibodies and NK cells (Figure 6G) exhibited extensive cytotoxicity, characterized by pronounced cell death and markedly reduced cell density. These findings suggest that vaccine-induced antibodies can bridge infected cells and NK cells, promoting NK cell activation and lysis of the infected targets. This ADCC effector activity demonstrates that the M2e nanoshell vaccine produces functionally competent antibodies capable of mediating viral clearance through non-neutralizing immune pathways.
Discussion
M2e has been considered a candidate for broadly reactive influenza vaccine development because of its high conservation across influenza A viruses. However, its poor immunogenicity has limited its application.30–33 Consistent with previous M2e-based vaccine studies, our findings support the need for an appropriate delivery and immune-stimulating platform to enhance M2e-directed immune responses. The PLGA nanoshell platform provides a biocompatible and thermally stable carrier that maintains antigen integrity, making it suitable for large-scale poultry vaccination.17 In this study, we extended this platform to an avian model to examine whether conserved-antigen vaccination could be effectively translated into chickens.
The present study contributes to underexplored areas in avian influenza vaccine research: the application of STING-targeted innate immune adjuvant strategies in poultry and the functional relevance of M2e-mediated Fc-dependent immune mechanisms in the avian host. Although the cGAS–STING pathway has emerged as a powerful target for vaccine adjuvant development in mammalian systems, particularly for viral vaccines, its translational application in poultry remains limited. STING agonists such as cGAMP have been shown in influenza vaccine studies to enhance type I interferon responses, dendritic cell activation, antigen cross-presentation, and heterologous protection, but most of these investigations have been conducted in mice or other mammalian models rather than food-producing avian species.34 The avian immune system differs substantially from mammalian systems in innate sensing pathways, antigen-presenting cell biology, and downstream effector responses, making direct extrapolation uncertain. In this context, our findings provide translational evidence that cGAMP-containing nanoshell vaccination in chickens is associated with enhanced antiviral gene expression, including upregulation of TLR7, IRF7, MX1, IFIH1, and Th1-associated immune pathways. These observations suggest that STING-targeted innate immune modulation may be a viable adjuvant strategy in poultry vaccine design, particularly for poorly immunogenic subunit antigens. Importantly, because extracellular cGAMP is rapidly degraded and requires efficient cytosolic delivery for optimal STING activation, the nanoshell platform may address a major translational limitation of cyclic dinucleotide adjuvants by facilitating intracellular delivery in avian immune cells.
Another important contribution of this study relates to the functional immunology of M2e-directed protection in poultry. M2e-based universal influenza vaccine strategies have been extensively investigated in mammalian systems, where protection is generally understood to rely less on classical neutralizing antibody activity and more on Fc-mediated effector mechanisms, particularly ADCC and phagocyte-mediated clearance of infected cells.11,35,36 However, whether a similar mechanism operates in chickens has remained largely unresolved because of limited characterization of avian Fc receptor biology, fewer validated immune effector assays, and incomplete understanding of NK cell-mediated antiviral responses in birds.28,35 Our study helps bridge this gap by demonstrating that serum antibodies elicited by M2e nanoshell vaccination can promote killing of influenza-infected target cells in the presence of chicken-derived CD3−CD8α⁺ NK-enriched splenocyte populations. While this does not constitute definitive mechanistic proof of classical Fc receptor-dependent ADCC, it provides initial functional evidence that M2e-directed antibody effector activity may be operational in poultry. This distinction is important because most avian influenza vaccines primarily rely on neutralizing antibodies directed against subtype-matched HA antigens, which are inherently vulnerable to antigenic drift and vaccine mismatch. By contrast, conserved-antigen strategies that engage infected-cell recognition and Fc-mediated immune clearance may represent a complementary path toward broader influenza control in poultry. Collectively, these findings extend the conceptual framework of M2e vaccine biology from mammalian proof-of-concept studies into a poultry-relevant immunological setting.
Despite these strengths, the current study has certain limitations. First, in the immunogenicity assay, the comparator formulation (M2e+ISA) does not independently isolate the relative contribution of nanoshell encapsulation versus STING agonism in the avian model. While our previous murine study established the optimized co-encapsulated M2e and STING agonist formulation as the lead candidate,17 the present work was designed as a translational validation study in chickens rather than mechanistic deconvolution of individual formulation components. Second, only one predefined formulation was evaluated for each vaccine group without dose titration, as the study was designed as a proof-of-concept evaluation rather than a formulation optimization study. Although a three-dose immunization regimen was used to evaluate maximal immune responses under experimental conditions, future studies will be needed to determine whether simplified vaccination schedules can achieve practical protection in field settings. Finally, in vivo protective efficacy was evaluated only in an H6N1 infection model. Although cross-reactive antibody responses against multiple influenza A subtypes were observed in vitro, these findings do not necessarily translate into in vivo protection against other strains, particularly highly pathogenic avian influenza viruses such as H5N1 or H7N9.37,38
Overall, this study suggests that the M2e nanoshell vaccine is a promising platform for inducing cross-reactive antibody responses and reducing disease severity following H6N1 infection in chickens. The simplicity, stability, and scalability of the nanoshell formulation further support its potential for practical field application in avian influenza control programs.
Conclusion
This study demonstrates that the M2e nanoshell vaccine significantly enhances the immunogenicity of the conserved M2e antigen and induces robust, durable, and cross-reactive immune responses in chickens. The vaccine promotes B cell activation, M2e-specific antibody production, and Fc-mediated cytotoxic responses, which may be supported by the upregulation of Th1-associated immune pathways. Following H6N1 challenge, a trend toward reduced viral RNA levels and tissue damage was observed. Importantly, in vitro killing assays revealed direct cytotoxic activity mediated by vaccine-induced antibodies and chicken NK cells, providing initial functional evidence of ADCC-related activity in the avian system. Overall, the M2e nanoshell vaccine represents a promising strategy for developing broadly protective avian influenza vaccines and provides potential value for controlling influenza outbreaks in poultry.
Ethics Statement
The study adhered to relevant institutional and national regulations. The animal experiments took place at the Animal Resource Center at National Taiwan University and followed established guidelines. All animal-related procedures were approved by the IACUC of National Taiwan University with the approval number NTU-112-EL-00120.
Acknowledgments
Financial support from the National Science and Technology Council and National Taiwan University was greatly appreciated. The authors thank the Animal Resource Center of National Taiwan University for their dedication to animal care. The authors also thank the Technology Commons, College of Life Science, National Taiwan University for their technical support.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors thank the funding from National Science and Technology Council, Taiwan (NSTC113-2327-B-002-006, NSTC114-2327-B-002-008) and National Taiwan University (115L8401).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Spackman E. A brief introduction to avian influenza virus. Methods Mol Biol. 2020;2123:83–19. doi:10.1007/978-1-0716-0346-8_7
2. Nabeshima K, Asakura S, Iwata R, et al. Sequencing methods for HA and NA genes of avian influenza viruses from wild bird feces using Oxford Nanopore sequencing. Comp Immunol Microbiol Infect Dis. 2023;102:102076. doi:10.1016/j.cimid.2023.102076
3. Blagodatski A, Trutneva K, Glazova O, et al. Avian influenza in wild birds and poultry: dissemination pathways, monitoring methods, and virus ecology. Pathogens. 2021;10(5):630. doi:10.3390/pathogens10050630
4. He J, Kam YW. insights from avian influenza: a review of its multifaceted nature and future pandemic preparedness. Viruses. 2024;16(3):458. doi:10.3390/v16030458
5. Xu H, Zhu S, Govinden R, Chenia HY. Multiple vaccines and strategies for pandemic preparedness of avian influenza virus. Viruses. 2023;15(8):1694. doi:10.3390/v15081694
6. Mo J, Spackman E, Swayne DE. Prediction of highly pathogenic avian influenza vaccine efficacy in chickens by comparison of in vitro and in vivo data: a meta-analysis and systematic review. Vaccine. 2023;41(38):5507–5517. doi:10.1016/j.vaccine.2023.07.076
7. Lim CML, Komarasamy TV, Adnan N, Radhakrishnan AK, Balasubramaniam VRMT. Recent advances, approaches and challenges in the development of universal influenza vaccines. Influenza Other Respir Viruses. 2024;18(3):e13276. doi:10.1111/irv.13276
8. Cargnin Faccin F, Perez DR. Pandemic preparedness through vaccine development for avian influenza viruses. Hum Vaccin Immunother. 2024;20(1):2347019. doi:10.1080/21645515.2024.2347019
9. Bimler L, Ronzulli SL, Song AY, et al. Matrix protein 2 extracellular domain-specific monoclonal antibodies are an effective and potentially universal treatment for influenza A. J Virol. 2021;95(5). doi:10.1128/jvi.01027-20
10. Von Holle TA, Moody MA. Influenza and antibody-dependent cellular cytotoxicity. Review. Front Immunol. 2019;10. doi:10.3389/fimmu.2019.01457
11. Jegerlehner A, Schmitz N, Storni T, Bachmann MF. Influenza A vaccine based on the extracellular domain of M2: weak protection mediated via antibody-dependent NK cell activity. J Immunol. 2004;172(9):5598–5605. doi:10.4049/jimmunol.172.9.5598
12. Tao W, Gill HS. M2e-immobilized gold nanoparticles as influenza A vaccine: role of soluble M2e and longevity of protection. Vaccine. 2015;33(20):2307–2315. doi:10.1016/j.vaccine.2015.03.063
13. Pati R, Shevtsov M, Sonawane A. Nanoparticle vaccines against infectious diseases. Front Immunol. 2018;9:2224. doi:10.3389/fimmu.2018.02224
14. Chattopadhyay S, Chen JY, Chen HW, Hu CJ. Nanoparticle vaccines adopting virus-like features for enhanced immune potentiation. Nanotheranostics. 2017;1(3):244–260. doi:10.7150/ntno.19796
15. Wibowo D, Jorritsma SHT, Gonzaga ZJ, Evert B, Chen S, Rehm BHA. Polymeric nanoparticle vaccines to combat emerging and pandemic threats. Biomaterials. 2021;268:120597. doi:10.1016/j.biomaterials.2020.120597
16. Lin LCW, Huang CY, Yao BY, et al. Viromimetic STING agonist-loaded hollow polymeric nanoparticles for safe and effective vaccination against middle east respiratory syndrome coronavirus. Adv Funct Mater. 2019;29(28). doi:10.1002/adfm.201807616
17. Tsai HH, Huang PH, Lin LC, et al. Lymph node follicle-targeting STING agonist nanoshells enable single-shot M2e vaccination for broad and durable influenza protection. Adv Sci. 2023;10(17):e2206521. doi:10.1002/advs.202206521
18. Liu Z, Zhou J, Xu W, et al. A novel STING agonist-adjuvanted pan-sarbecovirus vaccine elicits potent and durable neutralizing antibody and T cell responses in mice, rabbits and NHPs. Cell Res. 2022;32(3):269–287. doi:10.1038/s41422-022-00612-2
19. Elmanfi S, Yilmaz M, Ong WWS, et al. Bacterial cyclic dinucleotides and the cGAS-cGAMP-STING pathway: a role in periodontitis? Pathogens. 2021;10(6):675. doi:10.3390/pathogens10060675
20. Takahama S, Yamamoto T. Pattern recognition receptor ligands as an emerging therapeutic agent for latent HIV-1 infection. Front Cell Infect Microbiol. 2020;10:216. doi:10.3389/fcimb.2020.00216
21. Shen Y, Huang W, Nie J, Zhang L. Progress update on STING agonists as vaccine adjuvants. Vaccines. 2025;13(4):371. doi:10.3390/vaccines13040371
22. Wang X, Huang Z, Xing L, et al. STING agonist-based ER-targeting molecules boost antigen cross-presentation. Nature. 2025;641(8061):202–210. doi:10.1038/s41586-025-08758-w
23. Lin PH, Liang CY, Yao BY, et al. Robust induction of T(RM)s by combinatorial nanoshells confers cross-strain sterilizing immunity against lethal influenza viruses. Mol Ther Methods Clin Dev. 2021;21:299–314. doi:10.1016/j.omtm.2021.03.010
24. Lee YH, Huang CY, Hsieh YH, et al. A novel computer-assisted tool for 3D imaging of programmed death-ligand 1 expression in immunofluorescence-stained and optically cleared breast cancer specimens. BMC Cancer. 2024;24(1):121. doi:10.1186/s12885-023-11748-8
25. Bóna M, Földi J, Dénes L, Harnos A, Paszerbovics B, Mándoki M. Evaluation of the virulence of low pathogenic H9N2 avian influenza virus strains in broiler chickens. Vet Sci. 2023;10(12). doi:10.3390/vetsci10120671
26. Landmann M, Scheibner D, Graaf A, et al. A semiquantitative scoring system for histopathological and immunohistochemical assessment of lesions and tissue tropism in avian influenza. Viruses. 2021;13(5):868. doi:10.3390/v13050868
27. Ward CL, Dempsey MH, Ring CJ, et al. Design and performance testing of quantitative real time PCR assays for influenza A and B viral load measurement. J Clin Virol. 2004;29(3):179–188. doi:10.1016/s1386-6532(03)00122-7
28. Jansen CA, van de Haar PM, van Haarlem D, et al. Identification of new populations of chicken natural killer (NK) cells. Develop Comparat Immunol. 2010;34(7):759–767. doi:10.1016/j.dci.2010.02.009
29. Zhu WZ, Wen YC, Lin SY, Chen TC, Chen HW. Anti-influenza protective efficacy of a H6 virus-like particle in chickens. Vaccines. 2020;8(3):465. doi:10.3390/vaccines8030465
30. Subbiah J, Oh J, Kim K-H, et al. A chimeric thermostable M2e and H3 stalk-based universal influenza A virus vaccine. Npj Vaccines. 2022;7(1):68. doi:10.1038/s41541-022-00498-6
31. Hajam IA, Senevirathne A, Hewawaduge C, Kim J, Lee JH. Intranasally administered protein coated chitosan nanoparticles encapsulating influenza H9N2 HA2 and M2e mRNA molecules elicit protective immunity against avian influenza viruses in chickens. Vet Res. 2020;51(1):37. doi:10.1186/s13567-020-00762-4
32. Lotfi Z, Golchin M, Khalili-Yazdi A, Khalili M. Immunological properties of the SLLTEVET epitope of Influenza A virus in multiple display on filamentous M13 phage. Comp Immunol Microbiol Infect Dis. 2019;65:76–80. doi:10.1016/j.cimid.2019.05.004
33. Calzas C, Mao M, Turpaud M, et al. Immunogenicity and protective potential of mucosal vaccine formulations based on conserved epitopes of influenza a viruses fused to an innovative ring nanoplatform in mice and chickens. Front Immunol. 2021;12:772550. doi:10.3389/fimmu.2021.772550
34. Wang J, Li PY, Wu MX. Natural STING agonist as an “ideal” adjuvant for cutaneous vaccination. J Invest Dermatol. 2016;136(11):2183–2191. doi:10.1016/j.jid.2016.05.105
35. El Bakkouri K, Descamps F, De Filette M, et al. Universal vaccine based on ectodomain of matrix protein 2 of influenza A: fc receptors and alveolar macrophages mediate protection. J Immunol. 2011;186(2):1022–1031. doi:10.4049/jimmunol.0902147
36. Vanderven HA, Jegaskanda S, Wheatley AK, Kent SJ. Antibody-dependent cellular cytotoxicity and influenza virus. Curr Opin Virol. 2017;22:89–96. doi:10.1016/j.coviro.2016.12.002
37. Shi J, Zeng X, Cui P, Yan C, Chen H. Alarming situation of emerging H5 and H7 avian influenza and effective control strategies. Emerg Microbes Infect. 2023;12(1):2155072. doi:10.1080/22221751.2022.2155072
38. Neumann G, Eisfeld AJ, Kawaoka Y. Viral factors underlying the pandemic potential of influenza viruses. Microbiol Mol Biol Rev. 2025;89(2):e00066–24. doi:10.1128/mmbr.00066-24
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.