Back to Journals » ImmunoTargets and Therapy » Volume 14
Cell-Based Therapeutic Strategies for Autoimmune Diseases
Authors Quero FB, Troncoso-Bravo T, Farías MA, Kalergis AM
Received 23 December 2024
Accepted for publication 15 April 2025
Published 28 April 2025 Volume 2025:14 Pages 501—514
DOI https://doi.org/10.2147/ITT.S513629
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Flavio Salazar-Onfray
Francisco B Quero,1 Tays Troncoso-Bravo,1 Mónica A Farías,1 Alexis M Kalergis1,2
1Millennium Institute on Immunology and Immunotherapy, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Santiago, Chile; 2Departamento de Endocrinología, Facultad de Medicina, Pontificia Universidad Católica de Chile, Santiago, Chile
Correspondence: Alexis M Kalergis, Millennium Institute on Immunology and Immunotherapy, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Av. Portugal 49, Santiago, E-8330025, Chile, Tel +56-2-23542846, Email [email protected]
Abstract: Currently, the management of autoimmune disorders still being a challenge in terms of safety, efficiency, and specificity. Cell-based therapeutic strategies have emerged as a novel approach for autoimmune disease treatment, employing different cell therapy platforms, including tolerogenic dendritic cells, regulatory T cells, conventional and regulatory chimeric antigen receptor-T cells, mesenchymal and hematopoietic stem cells, each with their biological features. Here, we discuss the different cell therapy platforms, their immunological mechanisms of action, their therapeutic potential and benefits in autoimmune diseases, and challenges related to their production, scaling up, risks, and patient safety.
Keywords: autoimmunity, cell therapy, CAR T cell, tolerogenic dendritic cell, regulatory T cell, stem cells
Introduction
Autoimmune diseases encompass various disorders characterized by the loss of self-immune tolerance, resulting in autoreactive T and B cell activation. This dysregulation can lead to specific damage to individual tissues or organs, which can be seen in conditions such as type 1 diabetes (T1D) and multiple sclerosis (MS), or provoke systemic chronic inflammation, evident in disorders like systemic lupus erythematosus (SLE) and rheumatoid arthritis (RA). The etiology of autoimmune diseases is complex, influenced by multiple factors, including genetic predisposition,1 and particularly related to human leucocyte antigen (HLA) genes,1–3 epigenetic modifications,4–6 environmental factors,7,8 gestational conditions,9,10 hormones,11–13 gender,14 and ethnicity.15–17
Current treatment strategies for autoimmune diseases predominantly involve using glucocorticoids, hydroxychloroquine, and a range of immunosuppressants.18,19 While these medications can be effective in managing symptoms, they often lead to significant adverse events.20,21 Common side effects include increased susceptibility to infections, gastrointestinal intolerance, leukopenia, and complications such as heart failure, hypertension, osteoporosis, and anorexia.22,23 These adverse effects can limit treatments’ long-term usability and negatively impact patients’ life quality.24 Furthermore, although biological therapies like monoclonal antibodies against the cluster of differentiation 20 (CD20),25 tumor necrosis factor-alpha (TNF-α),26 interleukin-1β (IL-1β),27 or IL-6 receptor,28,29 have shown improved efficacy and a reduced incidence of side effects compared to traditional immune suppressor drugs, their high costs restrict accessibility, particularly in low- and middle-income countries, and it is prioritized for non-responder patients.30,31 Thus, substantial challenges involve the management of autoimmune diseases, including the need for more personalized treatments and safer alternatives to minimize side effects and improve the life quality of people with these affections.
Cell-based therapies have emerged as a promising therapeutic alternative in various fields,32–36 including stem cell applications for regenerative medicine,32 cancer treatment using chimeric antigen receptor (CAR) T cells33,34 or dendritic cell-based vaccines.35,36 These innovative therapeutic platforms offer new opportunities for treating autoimmune diseases by potentially restoring immune tolerance. Approaches like tolerogenic dendritic cells, regulatory T cells (Tregs), and CAR Treg therapies aim to modulate the immune response in an antigen-specific way.37–39 On the other hand, currently approved CAR T cell therapies can be applied for autoimmunity, deleting autoreactive B cells.40 Finally, stem cell-based therapies using mesenchymal stem cells focus on tissue repair and immune modulation.41 However, in the case of hematopoietic stem cells, they can achieve an “immune reset” after a conditioning regimen, leading to recovery of self-tolerance.42 Every therapy strategy mentioned above will be extensively addressed in this article, including findings in preclinical models and clinical trials in autoimmune diseases like T1D, MS, neuromyelitis optica (NMO), RA, idiopathic inflammatory myositis (IIM) and systemic sclerosis (SS).
This article discusses the novel cell therapy strategies developed for treating autoimmune diseases, focusing on their potential to restore immune balance and offer more targeted, long-lasting solutions. It addresses their mechanism of action and the challenges related to access, scalability, and safety for clinical applications. Highlighting the importance of this approach could represent a change in the management of autoimmune diseases that affect a large part of the population, offering safer and more effective treatments.
Emerging Cell Therapy Platforms
Tolerogenic Dendritic Cells
Dendritic cells (DCs) are the immune system’s most potent antigen-presenting cells (APCs).43 Through their ability to capture, process, and present antigens, they play a critical role in the onset of adaptive immune responses.44 DCs express many pathogen-associated molecular patterns (PAMPs) receptors and cytokine receptors, which trigger DC maturation by their activation.44–47 This maturation process is marked by a substantial increase in the expression of co-stimulatory molecules, such as CD40, CD80, CD83, and CD86, major histocompatibility complex (MHC) molecules and the chemokine receptor CCR7, which guide their migration to lymphoid tissues.47–49 In these lymph nodes, mature DCs present antigens to T cells, activating and differentiating naïve T cells into effector T cells, ultimately shaping immune responses against pathogens or abnormal cells such as tumor cells.45,49
In contrast to these highly immunogenic DCs, immature DCs exhibit a distinct phenotype characterized by their low expression of co-stimulatory molecules, MHC, and chemokine receptors.50,51 These cells, known as tolerogenic dendritic cells (tolDCs), are pivotal in maintaining immune homeostasis and promoting peripheral tolerance.52–57 TolDCs prevent autoimmune reactions by presenting self-antigens to T cells in a manner that leads to their inactivation or the generation of regulatory T cells (Tregs) rather than triggering an inflammatory immune response.50,51,58 The ability of tolDCs to suppress immune activation positions them as a key mechanism in avoiding autoimmune diseases and maintaining self-tolerance.52,59
The therapeutic potential of tolDCs has shown interest in recent years as a strategy for treating transplant rejection,60 allergies,61 and specialty for autoimmune diseases like SLE, MS, and RA.37,51,62–65 These cells can be generated ex vivo from precursor cells, such as hematopoietic stem cells or monocytes, and their tolerogenic properties can be enhanced using specific agents that inhibit their maturation.64,66,67 Agents such as dexamethasone, a potent glucocorticoid,66,68–70 active form of vitamin D3,69,71 interfering RNA (siRNA) to silencing B-cell activating factor (BAFF),67 and the peroxisome proliferator-activated receptor gamma (PPAR-γ) agonist rosiglitazone,66,68 have been shown to induce the tolerogenic phenotype in DCs.64,66–69,71 These DCs maturation inhibitors modulate the expression of stimulatory surface molecules, including CD40, CD80, CD83, CD86, and MHC-II,64,66–68,71 and cytokines such as TNF-α, IL-6, IL-12, and IFN-γ that would otherwise promote a proinflammatory response (Figure 1A).64,66–68
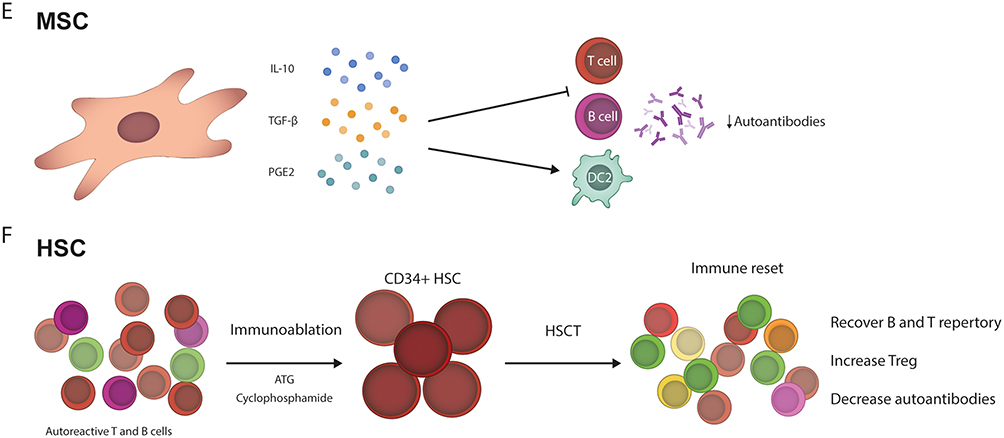
Figure 1 Continued. Figure 1 Main mechanisms of different cell-based therapy platforms. (A) Tolerogenic dendritic cells (tolDC) elicit an antinflammatory response by their low expression of MHC-II and low co-stimulatory molecules (CD80, CD40, CD83, and CD86). Additionally, tolDC produce low amounts of proinflammatory cytokines (TNF-α, IL-6, IL-12, and IFN-γ). tolDC have been reported to reduce T cell proliferation and autoantibodies production while increasing Treg cells. (B) Regulatory T cells (Treg) express regulatory surface molecules (LAG-3, CTLA-4, and TIGIT), leading to DC inhibition. Also, Tregs highly express anti-inflammatory cytokines (IL-10, IL-35, and TGF-β), which can inhibit both T and B cells, and systemic increase of IL-10 have been reported, while reduce TNF-α. (C) In this case, chimeric antigen receptor (CAR) T cells are engineered T cells to express a specific CAR for recognizing an antigen, CD19. After the recognition, the CAR T cell can kill the target cell by perforin/granzyme secretion or by inducing apoptosis by FasL signaling, diminishing B cells and then reducing autoantibodies production. (D) Regulatory CAR T cells are also engineered to overexpress FoxP3, which leads to a regulatory profile and anti-inflammatory function. When the CAR binds a CD19+ B cell, this is inhibited by the IL-10 produced from CAR Treg cells, decreasing the autoantibodies secretion. (E) Mesenchymal stem cells (MSC) are characterized by their anti-inflammatory cytokines/compounds (IL-10, TGF-β, PGE2) secretion capacity. Those immune mediators inhibit both T and B cells, diminish autoantibodies, and induce an increase in anti-inflammatories DC2. (F) The application of hematopoietic stem cells (HSC) requires the previous use of anti-thymocyte globulin and cyclophosphamide as immunoablation conditioning, depleting autoreactive T and B cells. Subsequently, the HSC transplantation (HSCT) is performed, leading to an “immune reset”, characterized by the recovery of TCR and BCR repertoire diversity, increasing of Treg and reduction of autoantibodies.
Extensive preclinical research has been conducted in murine models of autoimmunity, demonstrating the therapeutic efficacy of tolDC in a wide range of conditions, including experimental autoimmune encephalomyelitis (EAE), SLE-prone mice, non-obese diabetic (NOD) mice, and proteoglycan induced arthritis model. For instance, tolDCs have been explored in models of MS, where their administration was evaluated in preventive and therapeutic schedules, observing a reduction of clinical score in both conditions, an increase in IL-10 expression, and a Treg cell expansion.65 In NOD mice, murine type 1 diabetes model, tolDCs have delayed diabetes onset, leading to induction of splenic CD62L+ Tregs and reduced proinflammatory cytokines like TNF-α and IFN-γ.72 Similarly, tolDC in arthritis mice aims to enhance the proliferation of CD62L+ and FoxP3+ Treg subsets and reduce clinical arthritis manifestations.73 Finally, in NZM2410 SLE-prone mice, tolDCs elicit reduced anti-histone and anti-dsDNA antibodies, improved clinical scores, and reduced cutaneous lesions.66
Addressing the clinical trials findings, tolDCs have been evaluated in autoimmune diseases, including T1D, MS, and RA. In T1D, two Phase 1 clinical trials were conducted (NCT00445913 and NCT03895996). In the first one, tolDCs were generated from peripheral blood monocytes and treated with antisense oligonucleotides targeting CD40, CD80, and CD86 gene primary transcripts to induce a tolerogenic profile, and they were injected subcutaneously.74 The therapy was well tolerated, and no adverse events (AEs) reported could be related to the treatment.74 Notably, there was an increase in the frequency of peripheral B220+ CD11c− B cells and an increase of IL-4 and IL-10 compared with baseline.74 The second clinical trial in T1D consisted of three autologous intravenous administrations of DCs differentiated from monocytes and loaded with a synthetic oligopeptide of a signal peptide derived from the leader sequence of heat shock protein 60 (Hsp60sp), therapy called AVT001. The treatment was generally well tolerated, but two grade 3 AE of neutrophil count decrease (both in AVT001 and placebo group) were reported.75 Regarding the efficacy, the AVT001 treated group exhibited a lower decrease of C peptide compared to placebo. However, no apparent differences were found in hemoglobin A1c (HbA1c) or insulin-required dose.75 Regarding MS and NMO, there is only one completed clinical trial with posted results (NCT02283671). In this clinical trial, tolDCs were generated from autologous monocytes, and after differentiation, the cells were treated with dexamethasone and loaded with myelin and aquaporin peptides.76 Therapy with tolDCs was administrated in three intravenously doses every 2 weeks. It was well tolerated, without severe AE and without any therapy-related reactions. Patients remained stable clinically regarding relapses, disability, and magnetic resonance imaging and optical coherence tomography. In this case, it was reported an increase in IL-10 release from peripheral blood mononuclear cells (PBMCs) stimulated with the peptides and an increase in the frequency of a CD4+ CD10+ T cell, known as Tr1.76
Regarding RA, two clinical trials have been conducted (NCT03337165, NCT01352858). In the first one, tolDCs were generated from monocytes in the presence of dexamethasone, adding azoximer bromides as maturation stimuli.77 The single intra-articular injection of tolDCs was safe and well tolerated, and no one participant reported any AE. Additionally, the therapy improved disease activity, pain, and function for up to 6 months.77 Another trial in RA evaluated tolDCs produced from autologous CD14+ monocytes, and the tolerogenic profile was induced with dexamethasone and the active form of vitamin D3. Then, they were loaded with synovial fluid autoantigens. The administration was intra-articular, and the injections were safe and well tolerated, with no major knee flares.78 However, they did not show consistent clinical effects or sustained changes in the serum cytokines IL-8, IL-10, IL-16, IL-17, TNF-α, and IFN-γ. Despite this, two participants in the tolDC group showed symptom improvement and remained stable for 3 months, suggesting a potential beneficial effect.78
Regulatory T Cells Therapy
Regulatory T cells (Tregs) are a specialized subset of CD4+ T cells defined by the transcription factor forkhead box P3 (FoxP3) expression, which is crucial for their development and suppressive function.79 The pivotal role of this cell subset in immune homeostasis has been established by descriptions of Scurfy mice, which carry a FoxP3 mutation, conducing to the development of severe and lethal autoimmunity manifestations characterized by lymphoproliferation and multiorgan inflammation.79,80 Phenotypically, Tregs are characterized by high surface expression of CD25 (IL-2 receptor), low or absent CD127 (IL-7 receptor),38,81 and the presence of markers associated with their immunosuppressive capabilities such as cytotoxic T-lymphocyte antigen 4 (CTLA-4), lymphocyte-activation gen 3 (LAG3) and T cell immunoreceptor with Ig and ITIM domains (TIGIT).82–84
Treg cells exert their immunosuppressive and inflammation control effects through various characterized mechanisms.82 One of the most thoroughly studied involves the secretion of inhibitory cytokines, specifically IL-10, IL-35, and transforming growth factor beta (TGF-β),85–89 which play crucial roles in suppressing effector T cells,90 DCs,91 and B cells.88 However, their high expression of CD25 diminishes the availability of IL-2 cytokine, promoting a starvation environment for effector T cells (Figure 1B).92
In addition to cytokine-mediated suppression, Tregs engage in immunosuppression via direct cell-to-cell interactions. A key molecule in Treg-mediated suppression is CTLA-4, which interacts with CD80/CD86 on antigen-presenting cells, competing with the stimulatory coreceptor CD28 and ultimately reducing T cell activation.84,93 Another cell-to-cell mechanism is caused by the lymphocyte-activation gene 3 (LAG3) expression on Treg cells, and this molecule interacts with MHC-II molecules from DCs and elicits a higher antigen presentation, decreasing their antigen uptake, collaborating to induce a tolerogenic profile.94–96 TIGIT is another critical molecule highly expressed in Tregs that has emerged as part of cell-cell mechanisms controlling Treg function.97,98 TIGIT binds to CD155 on DCs, promoting the development of a mature immunoregulatory profile characterized by the expression of CD80, CD83, and CD86 but a reduced IL-12 production and enhanced IL-10 secretion.98 Moreover, TIGIT+ Tregs exert direct suppressive effects on effector T cells, particularly Th1 and Th17 cells, by secretion of soluble fibrinogen-like protein 2 (Fgl2).99
The mechanisms attributed to this subset of T cells have highlighted their potential as a therapeutic tool for autoimmune disorders. Therefore, several researchers have investigated the effects of adoptive Treg transfer on the development of autoimmunity across various animal models, including EAE,88 lupus-prone mice,100 and collagen-induced arthritis (CIA) mice.101 In EAE mice, adoptive transfer of Tregs resulted in reduced pain signs, delayed disease progression, and promoted IL-10 expression in T and B cells, attributing these effects to IL-35 production by Tregs.88 Treg transfer has been evaluated in lupus-prone NZB/NZW mice, leading to decreased renal damage due to reduced immune complex accumulation, lower proteinuria levels, and increased survival. However, no reduction in anti-dsDNA antibodies was observed.100 Finally, in the CIA model, a decrease in disease score and reduced TNF-α expression in T cells were noted.101
Moreover, the use of Tregs targeting a specific SLE antigen has been proposed, such as an overexpressing FoxP3 and a TCR specific for a peptide from Smith (Sm) antigen,102 a relevant antigen targeted in SLE. In this case, in-vitro evaluation of Sm-TCR Treg cells in co-culture with PBMCs showed IL-10 to increase, while IFN-γ and IL-17A decreased. Additionally, in-vivo studies in a humanized mouse model showed that the transfer of Sm-TCR Treg cells significantly reduced proteinuria and improved renal injury histological markers.102
Chimeric Antigen Receptor T (CAR T) Cells and CAR Treg
Chimeric antigen receptor T cells are genetically engineered ex-vivo and then readministered to patients. This cell engineering strategy enables generating autologous cells with specificity for a target antigen and delivers highly targeted cytotoxic effects against cells bearing that antigen.103 In the past few years, the therapeutic use of CAR T cells, specifically anti-CD19 or anti-B-cell maturation antigen (BCMA), has primarily targeted hematologic cancers such as non-Hodgkin lymphoma, acute leukemia, and multiple myeloma. Due to their effectiveness in these settings, CAR T cell therapies have gained recognition as the standard of care in some instances of refractory or relapsed hematologic cancers.104 CAR T cells’ primary mechanism of action is their cytotoxic activity,105 primarily mediated through the secretion of perforin, granzyme B, and granzyme A, which induce target cell lysis. Additionally, CAR T cells express Fas ligand (FasL), triggering apoptosis in Fas-expressing target cells (Figure 1C).105,106
The ability of CAR T cells to target and deplete CD19+ cells in the context of hematological cancers has increased interest in their applicability in autoimmune diseases, where dysregulated B cells and autoantibodies contribute significantly to disease pathogenesis.107 Accordingly, anti-CD19 CAR T cells have been tested in autoimmune murine models,108 such as SLE,109,110 and EAE,111 showing effectiveness in reducing clinical disease progression, diminishing proinflammatory cytokines, autoantibodies production, and elevating Treg cells.
In addition to conventional CAR T cell therapies, other CAR construct alternatives have been developed to generate therapeutic options, such as CAR Tregs.39 This type of therapy has been investigated as a therapeutic approach in an induced SLE model, overexpressing FoxP3 CAR anti-CD19 cells were generated and showed to reduce inflammation by increasing IL-10 production, which led to serum anti-dsDNA antibodies reduction and reduced histological kidney damage (Figure 1D).112
More than 120 clinical trials are currently registered for evaluating CAR T cell therapy in different autoimmune diseases. Besides, recent clinical evidence of anti-CD19 CAR T cells has been published. For example, a case series of SLE, IIM, and SS patients, refractories to immunosuppressive therapies, were treated with anti-CD19 CAR T cells.113 The therapy was considered safe, as no participants showed grade 3 or 4 cytokine release syndrome (CRS). One patient developed grade 4 neutropenia, which was resolved with clinical management. In the long term, another patient developed pneumonia, which resolved with antibiotic treatment.113 A transient depletion of B cells after therapy infusion was observed, with their reappearance after a mean of 112 days.113 Interestingly, in SLE patients, relevant autoantibodies against dsDNA, single-stranded DNA, secondary necrotic cells, nucleosomes, and Smith protein become negative and remain for at least one year of follow-up.113 All the patients had remission or significant clinical improvement and stopped their immunosuppressive therapy at the last follow-up.113 Similar results were reported in a phase 1 clinical trial (NCT04162353 and NCT05474885) where it was evaluated an anti-BCMA-CD19 compound CAR T cell for SLE patients with lupus nephritis.114 In this study, patients did not present severe CRS, and a seroconversion of autoantibodies with a disease activity reduction was reported in 9 out of 13 patients, achieving a symptom medication-free remission with a significant improvement in renal function.114
While CAR T therapies hold promise as a potential tool for treating autoimmune diseases, numerous challenges remain to allow their scaling up and achieve their use as a standard of care. A significant challenge in autologous cell therapy is batch-to-batch variability, which arises from the reliance on patient-derived cells. Each cell product partially depends on the initial material’s quality (apheresis product), including cell viability and T cell quantity.115 A proposed solution uses allogeneic or “universal” CAR T cells, which utilize healthy donor cells as starting material.116 This approach enables the production of larger cell quantities, allowing multiple doses to be obtained from a single batch. Additionally, it reduces costs associated with individual product processing and enhances manufacturing efficiency by providing ready-to-use products.116 Moreover, allogeneic CAR T cells present the risk of immune rejection.117 To mitigate this, gene engineering techniques have been applied to disrupt the expression of TCR and MHC, preventing these undesirable reactions.117 Another key challenge is therapy safety, and current clinical trials have demonstrated good tolerance, as well as an absence of severe AEs related to therapy. However, further clinical studies, including different patient cohorts and larger sample sizes, are necessary to confirm both safety and efficacy robustly.
Stem Cells: Mesenchymal Stem Cells, Hematopoietic Stem Cells
Mesenchymal Stem Cells
Mesenchymal stem cells (MSCs) are multipotent stromal precursor cells originating from the embryonic mesoderm. These cells are well characterized, and their established hallmarks include self-renewal capacity, multipotency, transplantability, plasticity, maintenance of genome integrity, and dependence on niche signals.118 They are harvested from various sources, including bone marrow, adipose tissue, menstrual blood, umbilical cord, placenta, and other tissues.119 Their differentiation potential spans osteogenic, adipogenic, chondrogenic lineages, and other cell types under specific conditions.120 Phenotypically, MSCs express markers like CD13, CD29, CD44, CD73, CD90, and CD105, while lacking markers such as CD14, CD19, CD20, CD34, CD45, CD79α and HLA-DR.121 The criteria for MSC characterization include their adherence to plastic surfaces, expression of the specific surface markers CD73, CD90, and CD105, and being negative for CD14, CD19, CD34, CD45, and HLA-DR, and capacity for tri-lineage differentiation into adipocytes, osteoblasts, and chondrocytes.122,123
In terms of mechanisms of action (Figure 1E), MSCs are known for their immunomodulatory properties through the secretion of various factors. They produce mainly TGF-β, IL-10, and prostaglandin E2 (PGE2), but also include IL-1 receptor antagonist (IL-1RA) hepatocyte growth factor (HGF), nitric oxide (NO), among others, which collectively limit immune responses. MSCs also influence immune cell populations by increasing regulatory T cells, anti-inflammatory Th2 cells, and type 2 DCs (DC2) while reducing pro-inflammatory Th1 cells, DC1 cells, and natural killer (NK) cells. Additionally, MSCs induce a shift in macrophage phenotype from the M1 (pro-inflammatory) to the M2 (anti-inflammatory), reducing IgG production in B cells.120 On the other hand, MSCs can promote tissue regeneration, and is well known their capacity to secrete paracrine trophic factors, which enhance injury regeneration and aim cell migration to damaged tissues such as HGF, stromal-derived factor-1 (SDF-1), insulin-like growth factor (IGF-1), epithelial growth factor (EGF), nerve growth factor (NGF), transforming growth factor-alpha (TGF-α), and tissue angiogenesis vascular endothelial growth factor (VEGF).124,125
In preclinical studies of lupus using different murine disease models, MSCs have shown significant therapeutic potential. In a murine model of SLE, MSCs have been observed to induce apoptosis in CD4+ T cells, thereby reducing kidney lesions associated with lupus.126 Additionally, human embryonic MSCs have demonstrated efficacy in alleviating pathological changes in MRL/Faslpr mice, which is attributed to their regulation of Th17 cell differentiation. These effects include enhanced survival, reduced anti-dsDNA antibody levels, and decreased renal damage, indicating the capacity of MSCs to mitigate both immune dysregulation and end-organ damage in lupus.127
Hematopoietic Stem Cells
Hematopoietic stem cells (HSCs) are multipotent cells essential for maintaining the lifelong production of all blood cell lineages, including erythrocytes, leukocytes, and platelets, through hematopoiesis.128 HSCs can be sourced from umbilical cord blood, collected from bone marrow, or mobilized to peripheral blood using recombinant G-CSF.129–131 These cells exhibit similar hallmark properties as described before for MSCs, such as self-renewal, multipotency, plasticity, maintenance of genome integrity, and dependence on niche signal.118 HSCs express key phenotypic markers, highlighting CD34 but also including CD33, CD133, CD90, and CD117, which allow their identification and characterization.132 Their potential multipotency enables them to have regenerative capacity, making them indispensable in hematopoietic reconstitution for treating malignancies and non-malignant diseases like leukemia or aplastic anemia, respectively.133,134
Hematopoietic stem cell transplantation (HSCT) has represented a transformative approach for hematologic malignancies and resetting the immune system in autoimmune diseases.42,131,135 Mechanistically, HSCT exerts its effects through profound immunological reprogramming, achieving a state of immune self-tolerance.42,135 This is primarily mediated by two pivotal mechanisms: First, the elimination of autoreactive immune cells through immunoablation, and second, the regeneration of a naive immune repertoire, often referred to as an “immune reset”.42,135–137 Immunoablation using anti-thymocyte globulin and cyclophosphamide eradicates autoreactive memory cells, evidenced by the disappearance of pathogenic autoantibodies and clonal T-cell populations post-transplantation.136 Simultaneously, HSCT stimulates functional renewal of Tregs, bolstering immune tolerance (Figure 1F).136 Thymic reactivation following transplantation further replenishes the immune system, generating a repertoire of naive T and B cells akin to those observed in young individuals.136–138
mRNA-Based Therapies
Messenger RNA is a single-stranded RNA (ssRNA) that participates in protein synthesis.139 Conventional mRNA-based vaccines code for a protein of interest, containing 5′ and 3′ untranslated regions (UTRs), flanking the coding region.139,140
mRNA vaccines are distributed in the body according to their formulation and route of administration, activating specific receptors that trigger a local inflammatory response.141 This allows the recruitment of antigen-presenting cells and the generation of a robust adaptive immune response, with T cell activation and long-lasting antibody production.140,141
mRNA technology, known primarily for its use in vaccines to induce antibody production, could be applied innovatively to regulate antibody production in autoimmune diseases.142 This strategy would take advantage of natural regulatory mechanisms of the immune system, such as feedback inhibition, where high-affinity antibodies can prevent B cell activation.141,142 mRNA could encode molecules that enhance this regulation or induce selective apoptosis of B and plasma cells, reducing the production of harmful autoantibodies.141 The mRNA could be engineered to produce molecules that interfere with key receptors such as BCRs and TLRs or immune checkpoints regulating B cell activation and function.142 It could also be used to generate proteins that neutralize autoantibodies or modulate cytokine responses and intracellular signaling pathways, affecting B cell proliferation and activation.142,143 Furthermore, it could enhance the function of regulatory T cells, which suppress autoimmune responses, or interfere with plasma cell survival, reducing antibody production under autoimmune conditions.142
Recently, a study revealed that vaccines based on mRNA modified with 1-methyl pseudouridine (m1Ψ) have the potential to induce antigen-specific immune tolerance in murine models of MS.144 These vaccines, formulated in liposomes without adjuvant capacity (m1Ψ-LPX), achieved prolonged and non-inflammatory antigen expression by avoiding the activation of TLR7 receptors, thus reducing the release of inflammatory cytokines and the activation of immune cells.144,145
In an EAE model, vaccination with m1Ψ-LPX encoding the MOG35-55 antigen completely protected mice from developing the disease when administered preventively.144 In cases of established disease, the vaccine stopped the progression and reversed the pathology in several cases. This effect was associated with the induction of regulatory T cells (FoxP3+) and elevated expression of immune exhaustion markers such as programmed cell death protein 1 (PD-1) and CTLA-4 on antigen-specific CD4+ T cells.144 Furthermore, the vaccine specifically suppressed Th1, Th17, and Th1/Th17 inflammatory cells without eliminating them through immune checkpoint-mediated mechanisms since inhibition of PD-1 or CTLA-4 abrogated their protective effect.144
Notably, this strategy was also effective in EAE models with different antigenic epitopes (PLP139-151), demonstrating cross-tolerance but maintaining intact immune responses to unrelated antigens.144 This approach’s flexibility, rapidity, and low cost allow for the customization of vaccines and their adaptation to multiple autoantigens, positioning them as a promising and highly innovative tool for treating autoimmune diseases.144,145
Future Perspectives and Challenges
In recent years, advances in cell therapy have led the field of biomedical research, offering new alternatives in regenerative medicine and oncology. These therapeutic strategies have also introduced innovative options for treating autoimmune diseases. Due to the versatility and range of underlying biological mechanisms of various cell therapy platforms (Figure 1), it is possible to apply these cutting-edge strategies to different autoimmune pathologies.
To achieve the clinical implementation of these innovative therapies reviewed in this article, it is essential to advance clinical trials to evaluate their safety, feasibility, and efficacy in humans. Although several therapies have already completed clinical studies and published their results, it is not uncommon for some trials to be suspended or their outcomes to remain unpublished. Nonetheless, the large number of registered clinical trials currently in recruitment and active status suggests promising progress in this field in the coming years.
Another significant challenge to the broad adoption of novel cell therapy alternatives is their high production cost, representing an access barrier. This problem is exemplified by the CAR T cell therapies currently approved for patient use.146,147
Finally, further research into cell-based therapies may improve our understanding of the immunological mechanisms driving autoimmune diseases, dysregulated inflammatory response conditions, and loss of immune homeostasis. Lasting, providing insights could lead to developing safer, more effective, personalized therapies tailored to each patient’s disease characteristics.
Conclusion
The emergence of novel and innovative cell-based therapeutic alternatives offers significant opportunities for patients suffering from autoimmune diseases. The various cell therapy platforms discussed above exhibit distinct mechanisms of action that enable treating, mitigating, and repairing the damage caused by autoimmunity. While these proposed approaches are highly promising, further clinical studies are needed, along with the completion of several ongoing trials and thorough reporting of their results and findings. Additionally, enhancing academic collaboration with industry is crucial to improve scalability and offset the high costs associated with manufacturing biological products. These therapies require Good Manufacturing Practice (GMP) facilities, clinically compatible reagents, specialized equipment, and highly trained technical, scientific, and clinical personnel. Numerous challenges remain in developing cell therapies for autoimmune diseases, yet preliminary results are encouraging, and their continued progress could redefine the standard of care for these conditions.
This article highlights the importance of these innovations and their potential impact on treating autoimmune diseases, providing a comprehensive analysis of the challenges and opportunities that will define the future of cell therapy. With a deeper understanding of these approaches and increased investment in research and development, these strategies could revolutionize the standard of autoimmune treatments, offering more effective and personalized treatments for patients.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Millennium Institute on Immunology and Immunotherapy ANID ACE 210015 (CN09_016 / ICN 2021_045; former P09/016-F, AMK); ANID/FONDECYT Regular grants #11900830, #1231851 (AMK); Biomedical Research Consortium CTU06 (AMK); FONDEF ID22I10252 (AMK); Fundación COPEC-UC 2019.R.1169 (AMK); ANID/FONDECYT Postdoctoral grant #3240624 (MAF).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Ahuja N, Singh J, Minz RW, Anand S, Das A, Taneja S. HLA and Non-HLA gene polymorphisms in autoimmune hepatitis patients of North Indian adults. Front Immunol. 2023;13. doi:10.3389/fimmu.2022.984083
2. Elfishawi S, Elfishawi M. Human leukocyte antigen and autoimmunity. Trans Autoimmun. 2022;255–263. doi:10.1016/B978-0-12-822564-6.00015-X
3. Ishigaki K, Lagattuta KA, Luo Y, James EA, Buckner JH, Raychaudhuri S. HLA autoimmune risk alleles restrict the hypervariable region of T cell receptors. Nat Genet. 2022;54(4):393–402. doi:10.1038/s41588-022-01032-z
4. Mazzone R, Zwergel C, Artico M, et al. The emerging role of epigenetics in human autoimmune disorders. Clin Epigenet. 2019;11(1):34. doi:10.1186/s13148-019-0632-2
5. Funes SC, Fernández-Fierro A, Rebolledo-Zelada D, Mackern-Oberti JP, Kalergis AM. Contribution of dysregulated DNA methylation to autoimmunity. Int J mol Sci. 2021;22(21):11892. doi:10.3390/ijms222111892
6. Funes SC, Rios M, Fernández-Fierro A, Di Genaro MS, Kalergis AM. Trained immunity contribution to autoimmune and inflammatory disorders. Front Immunol. 2022;13. doi:10.3389/fimmu.2022.868343
7. Ellis JA, Kemp AS, Ponsonby AL. Gene–environment interaction in autoimmune disease. Expert Rev Mol Med. 2014;16:e4. doi:10.1017/erm.2014.5
8. Touil H, Mounts K, De Jager PL. Differential impact of environmental factors on systemic and localized autoimmunity. Front Immunol. 2023;14. doi:10.3389/fimmu.2023.1147447
9. Albornoz EA, Carreño LJ, Cortes CM, et al. Gestational hypothyroidism increases the severity of experimental autoimmune encephalomyelitis in adult offspring. Thyroid. 2013;23(12):1627–1637. doi:10.1089/thy.2012.0401
10. Opazo MC, Haensgen H, Bohmwald K, et al. Imprinting of maternal thyroid hormones in the offspring. Int Rev Immunol. 2017;36(4):240–255. doi:10.1080/08830185.2016.1277216
11. Recalde G, Moreno-Sosa T, Yúdica F, et al. Contribution of sex steroids and prolactin to the modulation of T and B cells during autoimmunity. Autoimmun Rev. 2018;17(5):504–512. doi:10.1016/j.autrev.2018.03.006
12. Herrada AA, Campino C, Amador CA, Michea LF, Fardella CE, Kalergis AM. Aldosterone as a modulator of immunity. J Hypertens. 2011;29(9):1684–1692. doi:10.1097/HJH.0b013e32834a4c75
13. Muñoz-Durango N, Vecchiola A, Gonzalez-Gomez LM, et al. Modulation of immunity and inflammation by the mineralocorticoid receptor and aldosterone. Biomed Res Int. 2015;2015:1–14. doi:10.1155/2015/652738
14. Barber MRW, Falasinnu T, Ramsey-Goldman R, Clarke AE. The global epidemiology of SLE: narrowing the knowledge gaps. Rheumatology. 2023;62(Supplement_1):i4–i9. doi:10.1093/rheumatology/keac610
15. Goonesekera SD, Dey S, Thakur S, Davila EP. Racial/ethnic differences in autoimmune disease prevalence in US claims/EHR data. Am J Manag Care. 2024;30(1):e4–e10. doi:10.37765/ajmc.2024.89488
16. Wise A, Boring MA, Odom EL, et al. Racial and ethnic differences in the prevalence of patients with arthritis and severe joint pain and who received provider counseling about physical activity for arthritis among adults aged 18 years or older—United States, 2019. Arthritis Care Res. 2024;76(7):1028–1036. doi:10.1002/acr.25316
17. Langer-Gould AM, Gonzales EG, Smith JB, Li BH, Nelson LM. Racial and ethnic disparities in multiple sclerosis prevalence. Neurology. 2022;98(18). doi:10.1212/WNL.0000000000200151
18. Fanouriakis A, Kostopoulou M, Andersen J, et al. EULAR recommendations for the management of systemic lupus erythematosus: 2023 update. Ann Rheum Dis. 2024;83(1):15–29. doi:10.1136/ard-2023-224762
19. Smolen JS, Landewé RBM, Bergstra SA, et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2022 update. Ann Rheum Dis. 2023;82(1):3–18. doi:10.1136/ard-2022-223356
20. Apostolopoulos D, Morand EF. It hasn’t gone away: the problem of glucocorticoid use in lupus remains. Rheumatology. 2016;kew406. doi:10.1093/rheumatology/kew406
21. Ponticelli C, Glassock RJ. Prevention of complications from use of conventional immunosuppressants: a critical review. J Nephrol. 2019;32(6):851–870. doi:10.1007/s40620-019-00602-5
22. Gordon C, Amissah-Arthur MB, Gayed M, et al. The British society for rheumatology guideline for the management of systemic lupus erythematosus in adults. Rheumatology. 2018;57(1):e1–e45. doi:10.1093/rheumatology/kex286
23. NEGREI C, Bojinca V, BALANESCU A, et al. Management of rheumatoid arthritis: impact and risks of various therapeutic approaches. Exp Ther Med. 2016;11(4):1177–1183. doi:10.3892/etm.2016.3045
24. Rosenblum MD, Gratz IK, Paw JS, Abbas AK. Treating Human Autoimmunity: current Practice and Future Prospects. Sci Transl Med. 2012;4(125). doi:10.1126/scitranslmed.3003504
25. Randall KL. Rituximab in autoimmune diseases. Aust Prescr. 2016;39(4):131–134. doi:10.18773/austprescr.2016.053
26. Leone GM, Mangano K, Petralia MC, Nicoletti F, Past FP. Present and (Foreseeable) future of biological Anti-TNF alpha therapy. J Clin Med. 2023;12(4):1630. doi:10.3390/jcm12041630
27. So A, De Meulemeester M, Pikhlak A, et al. Canakinumab for the treatment of acute flares in difficult‐to‐treat gouty arthritis: results of a multicenter, Phase II, dose‐ranging study. Arthritis Rheum. 2010;62(10):3064–3076. doi:10.1002/art.27600
28. Levi M, Grange S, Frey N. Exposure‐exposure relationship of tocilizumab, an anti–IL‐6 receptor monoclonal antibody, in a large population of patients with rheumatoid arthritis. J Clin Pharmacol. 2013;53(2):151–159. doi:10.1177/0091270012437585
29. Ogata A, Tanaka T. Tocilizumab for the treatment of rheumatoid arthritis and other systemic autoimmune diseases: current perspectives and future directions. Int J Rheumatol. 2012;2012:1–14. doi:10.1155/2012/946048
30. Mendoza-Pinto C, Etchegaray-Morales I, Ugarte-Gil MF. Improving access to SLE therapies in low and middle-income countries. Rheumatology. 2023;62(Supplement_1):i30–i35. doi:10.1093/rheumatology/keac530
31. Patel JP, Konanur Srinivasa NK, Gande A, Anusha M, Dar H, Baji DB. The role of biologics in rheumatoid arthritis: a narrative review. Cureus. 2023. doi:10.7759/cureus.33293
32. Petrosyan A, Martins PN, Solez K, Uygun BE, Gorantla VS, Orlando G. Regenerative medicine applications: an overview of clinical trials. Front Bioeng Biotechnol. 2022;10. doi:10.3389/fbioe.2022.942750
33. Miliotou AN, Papadopoulou LC. CAR T-cell therapy: a new era in cancer immunotherapy. Curr Pharm Biotechnol. 2018;19(1):5–18. doi:10.2174/1389201019666180418095526
34. Jogalekar MP, Rajendran RL, Khan F, Dmello C, Gangadaran P, Ahn BC. CAR T-cell-based gene therapy for cancers: new perspectives, challenges, and clinical developments. Front Immunol. 2022;13. doi:10.3389/fimmu.2022.925985
35. Garg AD, Coulie PG, Van den Eynde BJ, Agostinis P. Integrating next-generation dendritic cell vaccines into the current cancer immunotherapy landscape. Trends Immunol. 2017;38(8):577–593. doi:10.1016/j.it.2017.05.006
36. Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, Sancho D. Dendritic cells in cancer immunology and immunotherapy. Nat Rev Immunol. 2020;20(1):7–24. doi:10.1038/s41577-019-0210-z
37. Passeri L, Marta F, Bassi V, Gregori S. Tolerogenic dendritic cell-based approaches in autoimmunity. Int J mol Sci. 2021;22(16):8415. doi:10.3390/ijms22168415
38. Shevyrev D, Tereshchenko V, Rhys HI. Treg heterogeneity, function, and homeostasis. Front Immunol. 2020;11:10. doi:10.3389/fimmu.2019.03100
39. Arjomandnejad M, Kopec AL, Keeler AM. CAR-T regulatory (CAR-Treg) cells: engineering and applications. Biomedicines. 2022;10(2):287. doi:10.3390/biomedicines10020287
40. Rampotas A, Richter J, Isenberg D, Roddie C. CAR-T cell therapy embarks on autoimmune disease. Bone Marrow Transplant. 2025;60(1):6–9. doi:10.1038/s41409-024-02429-6
41. Dazzi F, Krampera M. Mesenchymal stem cells and autoimmune diseases. Best Pract Res Clin Haematol. 2011;24(1):49–57. doi:10.1016/j.beha.2011.01.002
42. Alexander T, Greco R. Hematopoietic stem cell transplantation and cellular therapies for autoimmune diseases: overview and future considerations from the Autoimmune Diseases Working Party (ADWP) of the European Society for Blood and Marrow Transplantation (EBMT). Bone Marrow Transplant. 2022;57(7):1055–1062. doi:10.1038/s41409-022-01702-w
43. Sung SSJ. Monocyte-Derived Dendritic Cells as Antigen-Presenting Cells in T-Cell Proliferation and Cytokine Production. In: Allergy methods and protocols. 2019:131–141. doi:10.1007/978-1-4939-9591-2_9
44. Théry C, Amigorena S. The cell biology of antigen presentation in dendritic cells. Curr Opin Immunol. 2001;13(1):45–51. doi:10.1016/S0952-7915(00)00180-1
45. Hilligan KL, Ronchese F. Antigen presentation by dendritic cells and their instruction of CD4+ T helper cell responses. Cell mol Immunol. 2020;17(6):587–599. doi:10.1038/s41423-020-0465-0
46. Schraml BU, e Sousa C. Defining dendritic cells. Curr Opin Immunol. 2015;32:13–20. doi:10.1016/j.coi.2014.11.001
47. Tiberio L, Del Prete A, Schioppa T, Sozio F, Bosisio D, Sozzani S. Chemokine and chemotactic signals in dendritic cell migration. Cell mol Immunol. 2018;15(4):346–352. doi:10.1038/s41423-018-0005-3
48. Heuzé ML, Vargas P, Chabaud M, et al. Migration of dendritic cells: physical principles, molecular mechanisms, and functional implications. Immunol Rev. 2013;256(1):240–254. doi:10.1111/imr.12108
49. Yin X, Chen S, Eisenbarth SC. Dendritic cell regulation of T helper cells. Annu Rev Immunol. 2021;39(1):759–790. doi:10.1146/annurev-immunol-101819-025146
50. Gardner A, de Mingo Pulido Á, Ruffell B, Qiu L, Zhu L. Dendritic cells and their role in immunotherapy. Front Immunol. 2020;11:11. doi:10.3389/fimmu.2020.00924
51. Takenaka MC, Quintana FJ. Tolerogenic dendritic cells. Semin Immunopathol. 2017;39(2):113–120. doi:10.1007/s00281-016-0587-8
52. Iberg CA, Hawiger D. Natural and induced tolerogenic dendritic cells. J Immunol. 2020;204(4):733–744. doi:10.4049/jimmunol.1901121
53. Mackern‐Oberti JP, Llanos C, Riedel CA, Bueno SM, Kalergis AM. Contribution of dendritic cells to the autoimmune pathology of systemic lupus erythematosus. Immunology. 2015;146(4):497–507. doi:10.1111/imm.12504
54. Mackern-Oberti JP, Llanos C, Vega F, et al. Role of dendritic cells in the initiation, progress and modulation of systemic autoimmune diseases. Autoimmun Rev. 2015;14(2):127–139. doi:10.1016/j.autrev.2014.10.010
55. Iruretagoyena M, Wiesendanger M, Kalergis A. The dendritic cell-T cell synapse as a determinant of autoimmune pathogenesis. Curr Pharm Des. 2006;12(2):131–147. doi:10.2174/138161206775193145
56. Mackern-Oberti JP, Jara EL, Riedel CA, Kalergis AM. Hormonal modulation of dendritic cells differentiation, maturation and function: implications for the initiation and progress of systemic autoimmunity. Arch Immunol Ther Exp. 2017;65(2):123–136. doi:10.1007/s00005-016-0418-6
57. Llanos C, Carreño LJ, Kalergis AM. Contribution of dendritic cell/T cell interactions to triggering and maintaining autoimmunity. Biol Res. 2011;44(1):53–61. doi:10.4067/S0716-97602011000100007
58. Bourque J, Hawiger D. Life and death of tolerogenic dendritic cells. Trends Immunol. 2023;44(2):110–118. doi:10.1016/j.it.2022.12.006
59. Carreño LJ, González PA, Bueno SM, Riedel CA, Kalergis AM. Modulation of the dendritic cell–T-cell synapse to promote pathogen immunity and prevent autoimmunity. Immunotherapy. 2011;3(sup4):6–11. doi:10.2217/imt.11.38
60. Marín E, Cuturi MC, Moreau A, Wu M. Tolerogenic dendritic cells in solid organ transplantation: where do we stand? Front Immunol. 2018;9:9. doi:10.3389/fimmu.2018.00274
61. Sun W, Wei J, Li H, Wei F, Li J, Wen W. Adoptive cell therapy of tolerogenic dendritic cells as inducer of regulatory T cells in allergic rhinitis. Int Forum Allergy Rhinol. 2018;8(11):1291–1299. doi:10.1002/alr.22217
62. Mackern-Oberti J, Vega F, Llanos C, Bueno S, Kalergis A. Targeting dendritic cell function during systemic autoimmunity to restore tolerance. Int J mol Sci. 2014;15(9):16381–16417. doi:10.3390/ijms150916381
63. Llanos C, Mackern-Oberti JP, Vega F, Jacobelli SH, Kalergis AM. Tolerogenic dendritic cells as a therapy for treating lupus. Clin Immunol. 2013;148(2):237–245. doi:10.1016/j.clim.2013.04.017
64. Harry RA, Anderson AE, Isaacs JD, Hilkens CMU. Generation and characterisation of therapeutic tolerogenic dendritic cells for rheumatoid arthritis. Ann Rheum Dis. 2010;69(11):2042–2050. doi:10.1136/ard.2009.126383
65. Mansilla MJ, Sellès‐Moreno C, Fàbregas‐Puig S, et al. Beneficial effect of tolerogenic dendritic cells pulsed with <scp>MOG</scp> autoantigen in experimental autoimmune encephalomyelitis. CNS Neurosci Ther. 2015;21(3):222–230. doi:10.1111/cns.12342
66. Funes SC, Ríos M, Gómez‐Santander F, et al. Tolerogenic dendritic cell transfer ameliorates systemic lupus erythematosus in mice. Immunology. 2019;158(4):322–339. doi:10.1111/imm.13119
67. Zhao Y, Sun X, Yang X, et al. Tolerogenic dendritic cells generated by BAFF silencing ameliorate collagen-induced arthritis by modulating the Th17/regulatory T cell balance. J Immunol. 2020;204(3):518–530. doi:10.4049/jimmunol.1900552
68. Obreque J, Vega F, Torres A, et al. Autologous tolerogenic dendritic cells derived from monocytes of systemic lupus erythematosus patients and healthy donors show a stable and immunosuppressive phenotype. Immunology. 2017;152(4):648–659. doi:10.1111/imm.12806
69. Anderson AE, Swan DJ, Wong OY, et al. Tolerogenic dendritic cells generated with dexamethasone and vitamin D3 regulate rheumatoid arthritis CD4+ T cells partly via transforming growth factor- β 1. Clin Exp Immunol. 2016;187(1):113–123. doi:10.1111/cei.12870
70. Carreño LJ, Riedel CA, Kalergis AM. Induction of tolerogenic dendritic Cells by NF-κB blockade and Fcγ receptor modulation. In: Suppression and Regulation of Immune Responses: Methods and Protocols. 2010:339–353. doi:10.1007/978-1-60761-869-0_22
71. Xie Z, Chen J, Zheng C, et al. 1,25‐dihydroxyvitamin D 3 ‐induced dendritic cells suppress experimental autoimmune encephalomyelitis by increasing proportions of the regulatory lymphocytes and reducing T helper type 1 and type 17 cells. Immunology. 2017;152(3):414–424. doi:10.1111/imm.12776
72. Machen J, Harnaha J, Lakomy R, Styche A, Trucco M, Giannoukakis N. Antisense oligonucleotides down-regulating costimulation confer diabetes-preventive properties to nonobese diabetic mouse dendritic cells. J Immunol. 2004;173(7):4331–4341. doi:10.4049/jimmunol.173.7.4331
73. Jansen MAA, Spiering R, Ludwig IS, van Eden W, Hilkens CMU, Broere F. Matured tolerogenic dendritic cells effectively inhibit autoantigen specific CD4+ T cells in a murine arthritis model. Front Immunol. 2019;10:10. doi:10.3389/fimmu.2019.02068
74. Giannoukakis N, Phillips B, Finegold D, Harnaha J, Trucco M. Phase I (Safety) study of autologous tolerogenic dendritic cells in type 1 diabetic patients. Diabetes Care. 2011;34(9):2026–2032. doi:10.2337/dc11-0472
75. Gaglia JL, Daley HL, Bryant NK, et al. Novel autologous dendritic cell therapy AVT001 for type 1 diabetes. NEJM Evidence. 2024;3(7). doi:10.1056/EVIDoa2300238
76. Zubizarreta I, Flórez-Grau G, Vila G, et al. Immune tolerance in multiple sclerosis and neuromyelitis optica with peptide-loaded tolerogenic dendritic cells in a phase 1b trial. Proc Natl Acad Sci. 2019;116(17):8463–8470. doi:10.1073/pnas.1820039116
77. Kurochkina Y, Tyrinova T, Leplina O, et al. THU0178 THE EFFECT OF TOLEROGENIC DENDRITIC CELLS IN PATIENTS WITH RHEUMATOID ARTHRITIS: a SMALL PILOT TRIAL. Ann Rheum Dis. 2019;78:364. doi:10.1136/annrheumdis-2019-eular.2965
78. Bell GM, Anderson AE, Diboll J, et al. Autologous tolerogenic dendritic cells for rheumatoid and inflammatory arthritis. Ann Rheum Dis. 2017;76(1):227–234. doi:10.1136/annrheumdis-2015-208456
79. Brunkow ME, Jeffery EW, Hjerrild KA, et al. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat Genet. 2001;27(1):68–73. doi:10.1038/83784
80. Hadaschik EN, Wei X, Leiss H, et al. Regulatory T cell-deficient scurfy mice develop systemic autoimmune features resembling lupus-like disease. Arthritis Res Ther. 2015;17(1):35. doi:10.1186/s13075-015-0538-0
81. Sjaastad LE, Owen DL, Tracy SI, Farrar MA. Phenotypic and functional diversity in regulatory T cells. Front Cell Dev Biol. 2021;9. doi:10.3389/fcell.2021.715901
82. Dikiy S, Rudensky AY. Principles of regulatory T cell function. Immunity. 2023;56(2):240–255. doi:10.1016/j.immuni.2023.01.004
83. Grover P, Goel PN, Greene MI. Regulatory T cells: regulation of identity and function. Front Immunol. 2021;12. doi:10.3389/fimmu.2021.750542
84. Marangoni F, Zhakyp A, Corsini M, et al. Expansion of tumor-associated treg cells upon disruption of a CTLA-4-dependent feedback loop. Cell. 2021;184(15):3998–4015.e19. doi:10.1016/j.cell.2021.05.027
85. Li MO, Sanjabi S, Flavell RA. Transforming growth factor-β controls development, homeostasis, and tolerance of t cells by regulatory t cell-dependent and -independent mechanisms. Immunity. 2006;25(3):455–471. doi:10.1016/j.immuni.2006.07.011
86. Tsuji-Takayama K, Suzuki M, Yamamoto M, et al. The production of IL-10 by human regulatory T cells is enhanced by IL-2 through a STAT5-responsive intronic enhancer in the IL-10 locus. J Immunol. 2008;181(6):3897–3905. doi:10.4049/jimmunol.181.6.3897
87. Hsu P, Santner-Nanan B, Hu M, et al. IL-10 potentiates differentiation of human induced regulatory T cells via STAT3 and Foxo1. J Immunol. 2015;195(8):3665–3674. doi:10.4049/jimmunol.1402898
88. Duffy SS, Keating BA, Perera CJ, et al. Regulatory T cells and their derived cytokine, interleukin-35, reduce pain in experimental autoimmune encephalomyelitis. J Neurosci. 2019;39(12):2326–2346. doi:10.1523/JNEUROSCI.1815-18.2019
89. Schmidt A, Oberle N, Krammer PH. Molecular mechanisms of treg-mediated T cell suppression. Front Immunol. 2012;3:3. doi:10.3389/fimmu.2012.00051
90. Rubtsov YP, Rasmussen JP, Chi EY, et al. Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces. Immunity. 2008;28(4):546–558. doi:10.1016/j.immuni.2008.02.017
91. Li R, Li H, Yang X, Hu H, Liu P, Liu H. Crosstalk between dendritic cells and regulatory T cells: protective effect and therapeutic potential in multiple sclerosis. Front Immunol. 2022;13. doi:10.3389/fimmu.2022.970508
92. Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ. CD4+CD25+Foxp3+ regulatory T cells induce cytokine deprivation–mediated apoptosis of effector CD4+ T cells. Nat Immunol. 2007;8(12):1353–1362. doi:10.1038/ni1536
93. Lucca LE, Dominguez-Villar M. Modulation of regulatory T cell function and stability by co-inhibitory receptors. Nat Rev Immunol. 2020;20(11):680–693. doi:10.1038/s41577-020-0296-3
94. Huang CT, Workman CJ, Flies D, et al. Role of LAG-3 in regulatory T cells. Immunity. 2004;21(4):503–513. doi:10.1016/j.immuni.2004.08.010
95. Liang B, Workman C, Lee J, et al. Regulatory T cells inhibit dendritic cells by lymphocyte activation gene-3 engagement of MHC class II. J Immunol. 2008;180(9):5916–5926. doi:10.4049/jimmunol.180.9.5916
96. Arce-Sillas A, Álvarez-Luquín DD, Tamaya-Domínguez B, et al. Regulatory T cells: molecular actions on effector cells in immune regulation. J Immunol Res. 2016;2016:1–12. doi:10.1155/2016/1720827
97. Lee DJ. The relationship between TIGIT+ regulatory T cells and autoimmune disease. Int Immunopharmacol. 2020;83:106378. doi:10.1016/j.intimp.2020.106378
98. Yu X, Harden K, Gonzalez CL, et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat Immunol. 2009;10(1):48–57. doi:10.1038/ni.1674
99. Joller N, Lozano E, Burkett PR, et al. Treg cells expressing the coinhibitory molecule TIGIT selectively inhibit proinflammatory Th1 and Th17 cell responses. Immunity. 2014;40(4):569–581. doi:10.1016/j.immuni.2014.02.012
100. Scalapino KJ, Daikh DI, Unutmaz D. Suppression of glomerulonephritis in NZB/NZW lupus prone mice by adoptive transfer of ex vivo expanded regulatory T cells. PLoS One. 2009;4(6):e6031. doi:10.1371/journal.pone.0006031
101. Sun G, Hou Y, Gong W, et al. Adoptive induced antigen-specific treg cells reverse inflammation in collagen-induced arthritis mouse model. Inflammation. 2018;41(2):485–495. doi:10.1007/s10753-017-0704-4
102. Eggenhuizen PJ, Cheong RMY, Lo C, et al. Smith-specific regulatory T cells halt the progression of lupus nephritis. Nat Commun. 2024;15(1):899. doi:10.1038/s41467-024-45056-x
103. Moreno C, Haynie C, Cheever A, Weber K. Alternative CAR therapies: recent approaches in engineering chimeric antigen receptor immune cells to combat cancer. Biomedicines. 2022;10(7):1493. doi:10.3390/biomedicines10071493
104. Wang C, Wang J, Che S, Zhao H. CAR-T cell therapy for hematological malignancies: history, status and promise. Heliyon. 2023;9(11):e21776. doi:10.1016/j.heliyon.2023.e21776
105. Benmebarek MR, Karches CH, Cadilha BL, Lesch S, Endres S, Kobold S. Killing mechanisms of Chimeric Antigen Receptor (CAR) T cells. Int J mol Sci. 2019;20(6):1283. doi:10.3390/ijms20061283
106. Montalvo MJ, Bandey IN, Rezvan A, et al. Decoding the mechanisms of chimeric antigen receptor (CAR) T cell-mediated killing of tumors: insights from granzyme and Fas inhibition. Cell Death Dis. 2024;15(2):109. doi:10.1038/s41419-024-06461-8
107. Gómez-Bañuelos E, Fava A, Andrade F. An update on autoantibodies in systemic lupus erythematosus. Curr Opin Rheumatol. 2023;35(2):61–67. doi:10.1097/BOR.0000000000000922
108. Liu Y, Dong M, Chu Y, et al. Dawn of CAR-T cell therapy in autoimmune diseases. Chin Med J. 2024;137(10):1140–1150. doi:10.1097/CM9.0000000000003111
109. Jin X, Xu Q, Pu C, et al. Therapeutic efficacy of anti-CD19 CAR-T cells in a mouse model of systemic lupus erythematosus. Cell mol Immunol. 2021;18(8):1896–1903. doi:10.1038/s41423-020-0472-1
110. Kansal R, Richardson N, Neeli I, et al. Sustained B cell depletion by CD19-targeted CAR T cells is a highly effective treatment for murine lupus. Sci Transl Med. 2019;11(482). doi:10.1126/scitranslmed.aav1648
111. Gupta S, Simic M, Sagan SA, et al. CAR-T cell–mediated B-cell depletion in central nervous system autoimmunity. Neurol Neuroimmunol Neuroinflamm. 2023;10(2). doi:10.1212/NXI.0000000000200080
112. Doglio M, Ugolini A, Bercher-Brayer C, et al. Regulatory T cells expressing CD19-targeted chimeric antigen receptor restore homeostasis in systemic lupus erythematosus. Nat Commun. 2024;15(1):2542. doi:10.1038/s41467-024-46448-9
113. Müller F, Taubmann J, Bucci L, et al. CD19 CAR T-cell therapy in autoimmune disease — a case series with follow-up. N Engl J Med. 2024;390(8):687–700. doi:10.1056/NEJMoa2308917
114. Wang W, He S, Zhang W, et al. BCMA-CD19 compound CAR T cells for systemic lupus erythematosus: a phase 1 open-label clinical trial. Ann Rheum Dis. 2024;83(10):1304–1314. doi:10.1136/ard-2024-225785
115. Abou-el-Enein M, Elsallab M, Feldman SA, et al. Scalable manufacturing of CAR T cells for cancer immunotherapy. Blood Cancer Discov. 2021;2(5):408–422. doi:10.1158/2643-3230.BCD-21-0084
116. Watanabe N, Mamonkin M. Off-the-shelf chimeric antigen receptor T cells. Cancer J. 2021;27(2):176–181. doi:10.1097/PPO.0000000000000511
117. Chen S, van den Brink MRM. Allogeneic “off-the-shelf” CAR T cells: challenges and advances. Best Pract Res Clin Haematol. 2024;37(3):101566. doi:10.1016/j.beha.2024.101566
118. Beumer J, Clevers H. Hallmarks of stemness in mammalian tissues. Cell Stem Cell. 2024;31(1):7–24. doi:10.1016/j.stem.2023.12.006
119. Berebichez-Fridman R, Montero-Olvera PR. Sources and clinical applications of mesenchymal stem cells: state-of-the-art review. Sultan Qaboos Univ Med J. 2018;18(3):264. doi:10.18295/squmj.2018.18.03.002
120. Pittenger MF, Discher DE, Péault BM, Phinney DG, Hare JM, Caplan AI. Mesenchymal stem cell perspective: cell biology to clinical progress. NPJ Regen Med. 2019;4(1):22. doi:10.1038/s41536-019-0083-6
121. Walter S, Randau T, Hilgers C, et al. Molecular and functional phenotypes of human bone marrow-derived mesenchymal stromal cells depend on harvesting techniques. Int J mol Sci. 2020;21(12):4382. doi:10.3390/ijms21124382
122. Viswanathan S, Shi Y, Galipeau J, et al. Mesenchymal stem versus stromal cells: international Society for Cell & Gene Therapy (ISCT®) mesenchymal stromal cell committee position statement on nomenclature. Cytotherapy. 2019;21(10):1019–1024. doi:10.1016/j.jcyt.2019.08.002
123. Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy. 2006;8(4):315–317. doi:10.1080/14653240600855905
124. González-González A, García-Sánchez D, Dotta M, Rodríguez-Rey JC, Pérez-Campo FM. Mesenchymal stem cells secretome: the cornerstone of cell-free regenerative medicine. World J Stem Cells. 2020;12(12):1529–1552. doi:10.4252/wjsc.v12.i12.1529
125. Fu X, Liu G, Halim A, Ju Y, Luo Q, Song G. Mesenchymal stem cell migration and tissue repair. Cells. 2019;8(8):784. doi:10.3390/cells8080784
126. Huang S, Wu S, Zhang Z, et al. Mesenchymal stem cells induced CD4+ T cell apoptosis in treatment of lupus mice. Biochem Biophys Res Commun. 2018;507(1–4):30–35. doi:10.1016/j.bbrc.2018.10.133
127. Yuan L, Xiao ZT, Huang XZ, Wu MJ, Shi H, Liu AF. Human embryonic mesenchymal stem cells alleviate pathologic changes of MRL/Lpr mice by regulating Th7 cell differentiation. Ren Fail. 2016;38(9):1432–1440. doi:10.3109/0886022X.2015.1136894
128. Chander V, Gangenahalli G. Emerging strategies for enhancing the homing of hematopoietic stem cells to the bone marrow after transplantation. Exp Cell Res. 2020;390(1):111954. doi:10.1016/j.yexcr.2020.111954
129. Bazinet A, Popradi G. A general practitioner’s guide to hematopoietic stem-cell transplantation. Current Oncol. 2019;26(3):187–191. doi:10.3747/co.26.5033
130. Amouzegar A, Dey BR, Spitzer TR. Peripheral blood or bone marrow stem cells? Practical considerations in hematopoietic stem cell transplantation. Transfus Med Rev. 2019;33(1):43–50. doi:10.1016/j.tmrv.2018.11.003
131. Swart JF, Delemarre EM, van Wijk F, et al. Haematopoietic stem cell transplantation for autoimmune diseases. Nat Rev Rheumatol. 2017;13(4):244–256. doi:10.1038/nrrheum.2017.7
132. Anjos-Afonso F, Bonnet D. Human CD34+ hematopoietic stem cell hierarchy: how far are we with its delineation at the most primitive level? Blood. 2023;142(6):509–518. doi:10.1182/blood.2022018071
133. Pollyea DA, Altman JK, Assi R, et al. Acute myeloid leukemia, version 3.2023, NCCN clinical practice guidelines in oncology. J National Compr Cancer Network. 2023;21(5):503–513. doi:10.6004/jnccn.2023.0025
134. Kulasekararaj A, Cavenagh J, Dokal I, et al. Guidelines for the diagnosis and management of adult aplastic anaemia: a British society for haematology guideline. Br J Haematol. 2024;204(3):784–804. doi:10.1111/bjh.19236
135. Thiel A, Alexander T, Schmidt CA, et al. Direct assessment of thymic reactivation after autologous stem cell transplantation. Acta Haematol. 2008;119(1):22–27. doi:10.1159/000117824
136. Alexander T, Thiel A, Rosen O, et al. Depletion of autoreactive immunologic memory followed by autologous hematopoietic stem cell transplantation in patients with refractory SLE induces long-term remission through de novo generation of a juvenile and tolerant immune system. Blood. 2009;113(1):214–223. doi:10.1182/blood-2008-07-168286
137. Głowala-Kosińska M, Chwieduk A, Smagur A, et al. Thymic activity and T cell repertoire recovery after autologous hematopoietic stem cell transplantation preceded by myeloablative radiotherapy or chemotherapy. Biol Blood Marrow Transplant. 2016;22(5):834–842. doi:10.1016/j.bbmt.2016.01.014
138. van der Maas NG, Berghuis D, van der Burg M, Lankester AC. B cell reconstitution and influencing factors after hematopoietic stem cell transplantation in children. Front Immunol. 2019;10:10. doi:10.3389/fimmu.2019.00782
139. Pardi N, Hogan MJ, Weissman D. Recent advances in mRNA vaccine technology. Curr Opin Immunol. 2020;65:14–20. doi:10.1016/j.coi.2020.01.008
140. Troncoso‐Bravo T, Ramírez MA, Loaiza RA, et al. Advancement in the development of mRNA‐based vaccines for respiratory viruses. Immunology. 2024;173(3):481–496. doi:10.1111/imm.13844
141. Verbeke R, Hogan MJ, Loré K, Pardi N. Innate immune mechanisms of mRNA vaccines. Immunity. 2022;55(11):1993–2005. doi:10.1016/j.immuni.2022.10.014
142. Niazi SK. Anti-idiotypic mRNA vaccine to treat autoimmune disorders. Vaccines. 2023;12(1):9. doi:10.3390/vaccines12010009
143. Lee DSW, Rojas OL, Gommerman JL. B cell depletion therapies in autoimmune disease: advances and mechanistic insights. Nat Rev Drug Discov. 2021;20(3):179–199. doi:10.1038/s41573-020-00092-2
144. Krienke C, Kolb L, Diken E, et al. A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science. 2021;371(6525):145–153. doi:10.1126/science.aay3638
145. Wardell CM, Levings MK. mRNA vaccines take on immune tolerance. Nat Biotechnol. 2021;39(4):419–421. doi:10.1038/s41587-021-00880-0
146. Cliff ERS, Kelkar AH, Russler-Germain DA, et al. High cost of chimeric antigen receptor T-cells: challenges and solutions. Am Soc Clin Oncol Educat Book. 2023. doi:10.1200/EDBK_397912
147. Odstrcil MS, Lee CJ, Sobieski C, Weisdorf D, Couriel D. Access to CAR T-cell therapy: focus on diversity, equity and inclusion. Blood Rev. 2024;63:101136. doi:10.1016/j.blre.2023.101136
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.