Back to Journals » Infection and Drug Resistance » Volume 15
Cefmetazole Resistance Mechanism for Escherichia Coli Including ESBL-Producing Strains
Authors Ito R, Kawamura M ![]() , Sato T
, Sato T ![]() , Fujimura S
, Fujimura S
Received 12 July 2022
Accepted for publication 20 September 2022
Published 10 October 2022 Volume 2022:15 Pages 5867—5878
DOI https://doi.org/10.2147/IDR.S382142
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Ryota Ito, Masato Kawamura, Takumi Sato, Shigeru Fujimura
Tohoku Medical and Pharmaceutical University, Division of Clinical Infectious Diseases & Chemotherapy, Sendai, Miyagi, Japan
Correspondence: Ryota Ito, Tohoku Medical and Pharmaceutical University, Division of Clinical Infectious Diseases & Chemotherapy, 4-4-1 Komatsushima, Aoba-ku, Sendai, Miyagi, 981-8558, Japan, Tel +81-22-727-0169, Fax +81-22-727-0176, Email [email protected]
Purpose: Cefmetazole (CMZ), a cephamycin antibiotic, is primarily used as a definitive therapy for Extended Spectrum β-Lactamase (ESBL)-producing Escherichia coli infections. However, the mechanism of CMZ resistance in E. coli is still unknown. To elucidate the resistance mechanism and to determine combined drugs for prevention of resistance acquisition.
Methods: Clinical isolates of 14 ESBL-producing E. coli and non-producing 12 isolates were used in in vitro testing of CMZ resistance acquisition. After 10-day of CMZ exposure (1st subculture), these strains were incubated in an antibacterial-free medium for 14-day. These strains were again exposed to CMZ for 10-day (2nd subculture) and confirmed for changes in MIC. For each strain detected after 1st subculture, each mRNA expression level of porin, chromosomal ampC, and drug-efflux pump was measured using real-time RT-PCR. Relebactam (REL) has the potency to recover antimicrobial activity against carbapenem-resistant Enterobacterales that has porin deficiency. REL was added to the CMZ dilution series, and MIC changes and those of porin were confirmed.
Results: Of these 26 strains, 15 strains (57.7%) acquired resistance after 1st subculture, but after passage culture on the antibacterial-free medium, 11 strains recovered susceptibility. These 11 strains showed resistance after 2nd subculture. The expression levels of ompF and ompC were significantly decreased in these strains (P< 0.05). When REL was added, all strains suppressed resistance acquisition after 1st subculture. The mechanism was the activation of ompF.
Conclusion: Our results showed that the mRNA expression levels of genes encoding porin were decreased in the strains that acquired resistance due to CMZ exposure, and that ompF and ompC in particular were thought to be involved in the acquisition of resistance. The CMZ acquisition of resistance was also suppressed by the concomitant use of REL and actually suppressed the decrease in mRNA expression in ompF. It was confirmed that porin reactivated by REL.
Keywords: cefmetazole, porin, relebactam, extended spectrum β-lactamase, ESBL, Escherichia coli
A Letter to the Editor has been published for this article.
Introduction
Escherichia coli is the main causative organism of urinary tract infection.1 The intractable cases due to this pathogen increase; and two reasons are suggested recently. Firstly, biofilm formation attracts attention, and biofilms have 3-dimensional structures which formed on solid surfaces such as indwelling urinary catheters and other devices. Eradication of the biofilm-formed strain is extremely difficult because they function to protect the bacterial cell from antimicrobial agents.2 Conventionally, though the sterilization by the silver catalyst has been investigated as coping, most of enough effects are not shown.3,4 The sterilization technology using the new metal catalyst was reported recently and the clinical application is expected.5
The second is the emergence of various antimicrobial resistance E. coli. Extended-spectrum-lactamase (ESBL)-producing strain in particular rages worldwide, and is generally resistant to most penicillin and cephalosporin.6 In the United Kingdom, the strain which further acquired resistance in ciprofloxacin and trimethoprim/sulfamethoxazole was isolated at high frequency.7
ESBL-producing strain is detected in the world recently, the strain in E. coli of urinary tract origin was reported to be 17.9% in Europe in 2018, and 24% in Japan in 2015.8,9 Such ESBL-producing E. coli has drawn attention as a cause for not only nosocomial infections but also community-acquired infection.10,11
The Infectious Diseases Society of America (IDSA) guidelines for resistant bacteria treatment recommend such as carbapenem, quinolone, and trimethoprim/sulfamethoxazole as antibiotics to treat infections by ESBL-producing bacteria.12 Meanwhile in Japan, aside from carbapenem, β-lactam/β-lactamase inhibitor combinations such as tazobactam/piperacillin and tazobactam/ceftolozane are available for use, but the cephamycin antibiotic, cefmetazole (CMZ), is mainly used. Generally, the first-line drug in serious infections such as sepsis, is carbapenem, while CMZ is often selected as the initial drug of choice in moderate or mild infections in Japan.13 However, cephamycin has been reported to acquire resistance during use for some ESBL-producing bacteria.14 The acquisition of resistance due to mutation in outer membrane protein is suggested as a cause for such failure but details in CMZ remain unknown. In this study, we elucidate the resistance mechanism and determine combined drugs for the prevention of resistance acquisition.
Materials and Methods
Bacterial Strains and Drugs Used
A total of 63 strains were used: 62 strains of E. coli that were clinically isolated from 6 medical institutions in the Tohoku area of Japan, and a standard strain of E. coli ATCC 25922. Specimens for these strains originated from urine (55%), vaginal secretion (37%), and other sources (blood, stool) (8%).
The antimicrobial agent used was cefmetazole (CMZ: Sigma-Aldrich, Tokyo).
The following antimicrobial-containing KB disks (Eiken, Tokyo) were used to screen for ESBLs: ceftazidime (CAZ), cefotaxime (CTX), ceftazidime/clavulanate (CAZ/CVA), and cefotaxime/clavulanate (CTX/CVA).
Screening of ESBL-Producing Strains and Gene Analysis
The disk diffusion method that conforms to the Clinical & Laboratory Standards Institute (CLSI) was used to check whether ESBL was produced. Namely, the inhibition circle diameters of CTX and CAZ, the discs for drug susceptibility testing, were compared to those of the fixed-dose combination with CVA (CTX/CVA and CAZ/CVA). If the inhibition ring of the compound with CVA extended 5 mm or more than that of a single agent of CTX or CAZ, it was assessed as a producer of ESBL.15
To ascertain a genotype of ESBL, the ESBL-producing colony was suspended in 500 µL of milli-Q water and then centrifuged (12,000 rpm, 2 minutes). Centrifugal pellets were spiked with 100 µL of InstaGene Matrix (Bio-Rad, Hercules, CA), stirred, and incubated for 30 minutes at 56℃. Following heat block (100℃, 8 minutes), supernatant obtained from centrifugation (12,000 rpm, 3 minutes) was used as the template DNA. Each ESBL gene of the CTX-M-1 group, CTX-M-2 group, CTX-M-8 group, CTX-M-9 group, CTX-M-25 group, CTX-M chimera, GES, TEM, and SHV was determined by the Cica Geneus® ESBL Genotype Detection KIT 2 (Kanto Chemical Co., Inc., Tokyo). As for the PCR reaction conditions, one cycle (reaction for 2 minutes at 94℃ is followed by 15 seconds at 94℃, 15 seconds at 63℃, and 40 seconds at 72℃) was repeated for a total of 30 cycles. The PCR product was added to 2% agarose gel and electrophoresis was performed at 100V for 30 minutes to examine whether a band is present.
In vitro Testing of CMZ Resistance Acquisition
The broth microdilution method was used to determine the minimum inhibitory concentration (MIC) of CMZ against all 63 E. coli strains. E. coli was cultured using Muller-Hinton Agar (MHA: Eiken) at 37℃ for 24 hours. Phosphate buffered saline (PBS) was used to adjust the bacterial solution to McFarland 0.5, and 5 µL of the adjusted bacterial solution was added to 1.5 mL of the Muller-Hinton broth (MHB: Becton Dickinson, NJ). The obtained solution was inoculated with 100 µL into a dilution series of CMZ prepared in the range of 0.125–256 µg/mL, and incubated at 37℃ for 16–20 hours. The drug susceptibility test results were compliant with CLSI, and it was deemed resistant when the MIC of CMZ was ≥64 µg/mL.15
In this study, 14 E. coli strains confirmed to be ESBL-producing were used, along with 12 non-ESBL-producing strains (including ATCC25922 strains) as the control. These 26 strains were divided into CMZ’s MIC: ≤1 µg/mL group, and 2–4 µg/mL group to assess in vitro resistance acquisition. As for CMZ exposure. To a dilution series (2 MIC-1/4 MIC) of CMZ prepared in 96-well microplates, the bacterial solution was added as described above and this was incubated at 37℃ for 16–20 hours. Surviving strains in 1/2 MIC of CMZ were inoculated into another dilution series and after similar incubation, the MIC was determined. This operation was repeated for a total of 10 days.
Ten days later, surviving strains in 1/2 MIC were sub-cultured every day into antibacterial-free MHB for 14 days. Finally, these strains were exposed to CMZ for 10 days, similar to the method described above, to confirm changes in MIC.
Changes in Expression Levels of Porin, Chromosomal ampC, and Drug Efflux Pump
For each strain detected after 10 days of CMZ exposure, each mRNA expression level of porin (ompF, ompC, phoE), chromosomal ampC, and drug-efflux pump (acrA, yhiV, mdfA) was measured using real-time RT-PCR.
For RNA extraction of each strain, TRI REAGENTTM LS (Molecular Research Center, Inc. OH) was used. The mRNA expression level was determined by using iTaq Universal SYBR Green One-Step Kit (Bio-Rad) in the CFX ConnectTM Real-time System (Bio-Rad). The sequences of primers used in the study are summarized in Table 1. Each mRNA expression level was calculated by using the mRNA expression of each wild type as 1 and indicated by fold difference. In this study, the expression level of each gene was defined as 2-fold or more up-regulated and 0.5-fold or less down-regulated.16–19
 |
Table 1 Primers Used in This Study |
Changes in CMZ Susceptibility at the Time of Addition of Relebactam
A total of 10 strains (5 ESBL-producing strains, and 5 non-EBSL-producing strains) were selected: 5 strains that showed CMZ resistance on Day 5 of in vitro testing of resistance acquisition; and 5 strains that retained susceptibility.
In the in vitro study, relebactam (REL) has effects to recover carbapenem antibacterial activity against carbapenem-resistant Enterobacterales that has porin deficiency in the outer membrane.20,21 Thus, REL was used in this test. REL was added to the previously described CMZ dilution series, and the broth microdilution method was used to measure MIC. This operation was continued for 10 days and changes in MIC were confirmed. The concentration of REL in each well was set at 4 µg/mL, 8 µg/mL, and 16 µg/mL, and changes by concentration were examined.20 The mRNA expression of porin (ompF, ompC, phoE) was measured using real-time RT-PCR for those strains on Day 5 of CMZ exposure at the time of REL addition (16 µg/mL).
Statistical Analysis
Statistical analysis was performed to examine the presence or absence of resistance between two groups using the Chi-square test. The difference in each mRNA expression level was analyzed using the Student’s t-test. If the difference was P<0.05, it was determined as “significantly different.”
Results
CMZ Susceptibility and Genotype of ESBL-Producing Strain
The MIC range of CMZ against 62 test strains (wild type) was 0.5–4 µg/mL, and all strains showed susceptibility. Of the 62 strains, ESBL-producing strains accounted for 14 strains (22.6%). Of the 14, 13 strains showed CTX-M type ESBL-producing genes, and 5 of the 13 strains simultaneously held TEM type genes. The remaining one strain only showed TEM type genes.
Of the 14 ESBL-producing strains, 6 strains were in the MIC: ≤1 µg/mL group of CMZ for wild type, and 8 strains were in the 2–4 µg/mL group. Of the 12 non-ESBL-producing strains selected as the control, 7 strains were in the MIC: ≤1 µg/mL group and 5 strains were in the 2–4 µg/mL group. Of these 26 strains, 15 strains (57.7%) acquired resistance after the 10-day CMZ exposure (Figure 1A). The breakdown was that 10 of 13 strains (76.9%) in the MIC: 2–4 µg/mL group showed resistance, and this resistance rate was significantly higher than that (38.4%) in the MIC: ≤1 µg/mL group (P<0.05) (Figure 1A). Figure 1B shows the results in ESBL-producing strains. In the MIC: 2–4 µg/mL group (wild type), resistant strains began to develop on Day 4 of the exposure, and increased time-dependently. In the MIC: ≤1 µg/mL group, resistant strains began to develop on Day 6 of the exposure, and increased time-dependently as in the case of the MIC: 2–4 µg/mL group. On Day 10, resistance rates exceeded 60% in both groups. In the case of non-ESBL-producing strains, resistant strains began to develop on Day 5 of the exposure in the MIC: 2–4 µg/mL group, but only one strain increased resistance on Day 10 of exposure in the MIC: ≤1 µg/mL group (Figure 1C). In other words, on Day 10 of exposure, significant resistance was acquired in the MIC: 2–4 µg/mL group (P<0.05).
 |
Figure 1 Resistance to in vitro cefmetazole exposure, by MIC group, in E. coli total (A), ESBL-producing E. coli (B), and non-ESBL E. coli (C). The data shows the cumulative resistance rate and 95% CI due to everyday cefmetazole exposure. Note: *P < 0.05. |
Changes in MIC of CMZ in in vitro Loading Models
Of a total of 26 strains, 15 acquired resistance after 10 days of CMZ exposure (1st subculture), but after passage culture for 14 days on the antibacterial-free medium, 11 strains recovered susceptibility. The remaining 4 strains (ES-1, −4, −7, −11) maintained resistance and all of these were ESBL-producing strains (Table 2). All of the 11 strains that recovered susceptibility showed resistance after re-exposure to CMZ for 10 days (2nd subculture).
 |
Table 2 MIC of Cefmetazole Against Escherichia Coli |
After the second 10-day CMZ exposure, resistance appeared in 12 of 14 ESBL-producing strains (85.7%), and 9 of 12 non-ESBL-producing strains (75%). Compared with the first 10-day CMZ exposure, 19 of 26 strains (73%) showed an increase in MIC by a factor of 2–32 after the second 10-day CMZ exposure. On the other hand, as for 11 strains that retained susceptibility after the first 10-day CMZ exposure, 6 strains (54.5%) acquired resistance after the second 10-day CMZ exposure.
Changes in mRNA Expression Levels in Porin, Chromosomal ampC, and Drug Efflux Pump
In 15 strains that acquired resistance after the first 10-day CMZ exposure (MIC: ≥64 µg/mL), the expression levels of all porin genes (ompF, ompC, phoE) significantly decreased as compared with the wild type (P<0.05) (Figure 2). Also, in 11 non-resistant strains, the expression level of phoE significantly decreased (P<0.05). In contrast with porin, the expression levels of acrA, yhiV, and mdfA, which code chromosomal ampC and the drug efflux pump, showed no changes (Figure 3).
 |
Figure 2 mRNA expression levels of ompF, ompC, and phoE in resistant and non-resistant strains after 10 days of cefmetazole exposure. Abbreviations: R, cefmetazole resistant group; non-R, cefmetazole non-resistant group. Note: *P < 0.05. |
 |
Figure 3 mRNA expression levels of chromosomal ampC, acrA, yhiV, mdfA genes in this study cefmetazole exposure assay. Abbreviations: R, cefmetazole resistant group; non-R, cefmetazole non-resistant group. Notes: The expression levels of ampC, acrA, yhiV, mdfA were expressed with each wild type as 1. |
Effect of Relebactam Addition to the Susceptibility of Cefmetazole
Five strains (3 strains of the ESBL production and 2 non-production) which acquired resistance after CMZ exposure in an early stage (five days later) were investigated. The addition of 8 µg/mL REL resulted in the suppression of resistance acquisition in 3 of 5 strains (60%) after the 10-day CMZ exposure (Figure 4). However, when 16 µg/mL REL was combined, all 5 strains (100%) suppressed resistance acquisition after the 10-day CMZ exposure. Thus, the resistance rate was significantly lower than that after the exposure to the CMZ only (P<0.05).
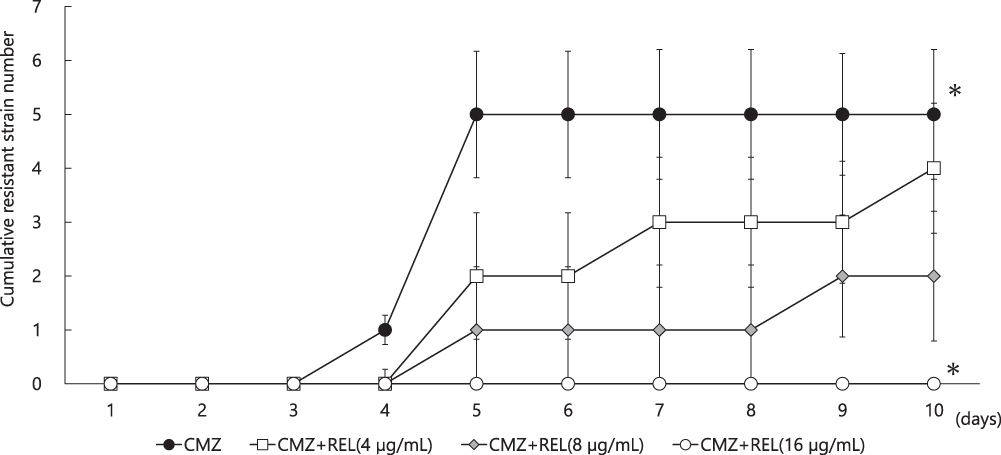 |
Figure 4 Resistance to in vitro cefmetazole exposure at the time of addition of relebactam. Abbreviations: CMZ, cefmetazole; REL, relebactam. Note: *P < 0.05. |
In in vitro testing of resistance acquisition, those strains that increased resistance to CMZ showed susceptibility after REL was added, and the mRNA expression levels of ompF, ompC, or phoE also increased significantly (Table 3). On the other hand, those strains that retained susceptibility showed a significant increase only in ompF after the addition of REL.
 |
Table 3 Changes in ompF, ompC and phoE mRNA Expression in Combination with Relebactam and Cefmetazole |
Discussion
The resistance of β-lactams to E. coli is triggered by overexpression of β-lactamase and downregulation of porin.22 E. coli is frequently the causative bacteria in urinary tract and intra-abdominal infections, and the prevalence of E. coli having ESBL-producing genes has increased in recent years.23 Overuse of carbapenem antibiotics as the standard treatment of ESBL-producing bacterial infections may lead to an increase in strains resistant to carbapenem. For this reason, with a view to reducing the use of carbapenem, CMZ, which is effective against ESBL-producing strains in vitro is often used as a substitute. Some reports indicate results where the therapeutic effects of non-carbapenem antibiotics can be expected depending on the infected sites.24 In fact, the use of CMZ increased by 21% in the period from 2004 to 2016 in Japan.25
The MIC range of CMZ against ESBL-producing E. coli is reported to be 0.5–4 µg/mL in Japan,23 and the same results were obtained also in this study. After the 10-day CMZ exposure (1st subculture), 57.7% increased resistance in the study. Particularly against E. coli where MIC of the wild type shows 2–4 µg/mL, significant resistance was acquired (Figure 1). The MIC50 of CMZ against ESBL-producing E. coli (CTX-M-14, −15, −27 type) is ≤1 µg/mL, and MIC90 of CMZ is reported to be 8 µg/mL, 4 µg/mL, and 4 µg/mL, respectively, showing variation in the MIC distributions.23 In both groups, more than 60% of ESBL-producing strains showed resistance after 1st subculture. Thus, it was considered that ESBL-producing strains increased resistance after CMZ exposure regardless of the wild type’s MIC. By passage culture in the antibacterial-free medium thereafter, 11 of 26 strains (including 6 ESBL-producing strains) recovered susceptibility, but showed resistance again after 2nd subculture. This suggests that even if the first administration of CMZ was effective, it was expected to acquire resistance and ineffective when administered again.
To elucidate the mechanism of acquiring resistance to CMZ, we examined changes in porin, AmpC β-lactamase, and the drug efflux pump. On the outer membrane of gram-negative bacteria, a protein, called porin, exists, functioning as a pathway for essential nutrients and antimicrobials to enter the bacteria and is also involved in the acquisition of resistance in gram-negative bacteria.26 OmpF, OmpC, and PhoE are major porins in E. coli.27 OmpF is the outer membrane permeation pathway for many antibacterials, including β-lactams and quinolone, while OmpC has an affinity with positive ions, and PhoE has an affinity with negative ions and inorganic phosphate.27 The expression status of PhoE is usually inferior to that of OmpF or OmpC, but the pore diameter of OmpF is 1.2 nm,28 and that of PhoE is 1.0 nm.29 All of these are involved in the uptake of β-lactams into the bacterial cells.30 Decrease or loss of these porins causes changes in drug susceptibility.31 In cefoxitin, ie, cephamycin antibiotic similar to CMZ, there have been reports that a decrease in porin is involved in the acquisition of resistance to E. coli or Klebsiella pneumoniae.32 In the group that acquired resistance after CMZ exposure, decreases in mRNA expression levels were confirmed in ompF, ompC, and phoE (Figure 2), suggesting the involvement of porins in resistance acquisition. PhoE is mainly selected when OmpF and OmpC are deficient in E. coli,28 but in this study, the mRNA expression level of phoE in the non-resistant group significantly decreased. It is known that E. coli decrease phoE expression to skirt the accumulation of secondary metabolic products such as organic acid.33 Despite this phenomenon also being observed in our experiment, its decreased levels were similar in both groups. Therefore, it was considered that phoE decrease is little contributed to CMZ-resistance acquisition. In contrast, E. coli upregulate ompF to intake metallic ions.34 However, ompF was decreased in CMZ-resistance acquired group; therefore, it was suggested that this mechanism was strongly associated with CMZ resistance.
In E. coli, mutation of ompF and possession of ESBL (TEM type of SHV type) contributes to the 4 times raising of MIC value,30 but in strains that acquired resistance after CMZ exposure, the resistance was not affected by the difference in genotype. Nevertheless, of the strains that increased resistance to CMZ, 4 strains (ES-1, −4, −7, −11) maintained resistance even after passage culture without CMZ exposure, and all of them were ESBL-producing strains.
One of the resistance mechanisms for β-lactams is the involvement of AmpC β-lactamase.35 Cephamycin is not usually hydrolyzed at ESBL, but it is decomposed by AmpC β-lactamase.36
Normally, E. coli produces little AmpC, but when the promoter of the ampC gene is mutated,37,38 1–2% of E. coli produce large amounts of AmpC.22 In this study, we confirmed changes in chromosomal ampC mRNA expression due to CMZ exposure, but these changes were scant.
Similarly, the impact of the drug efflux pump is a mechanism to acquire resistance. The main efflux pump is AcrAB-TolC in gram-negative bacteria, followed by YhiV, and MdfA. The involvement of overexpression of genes that code the drug efflux pump is suggested in the resistance acquisition of cephamycins.39 In this study, however, we confirmed little change in the mRNA expression level of the drug efflux pump in the strains resistant to CMZ as compared with the mRNA expression level in each wild type, and confirmed no relation with the resistance acquisition.
REL used in this study is a β-lactamase inhibitor that blocks β-lactamase, belonging to Ambler class A and class C.40 REL itself has no antibacterial activity.41 However, REL may have a potency to recover antibacterial activity for carbapenem-resistant Enterobacterales due to the porin deficiency. Resistance to CMZ confirmed in the study was suppressed in combined use with REL. Remarkably, all strains did not acquire resistance when REL was added at 16 µg/mL. This may be due to mRNA deregulation especially in ompF being suppressed and REL having effects to activate the inflow of antibacterial into bacterial cells by activating ompF. The combined use of REL may be expected to suppress CMZ’s acquisition of resistance to E. coli. The agent with the porin activator action is not known at present except REL. However, it was reported that titanium dioxide was effective for sterilization of the drug-resistant gram-negative bacteria recently.42,43 Though the titanium dioxide is not medical supplies, we may expect it as a new porin activating agent.
Conclusion
The strain of 57.7% of E. coli acquired resistance by in vitro exposure for 10 days of CMZ which was one of the therapeutic drugs for the ESBL-producing gram-negative bacteria infection. As mRNA expression of ompF decreased 50% in these acquisitions of resistant strains in comparison with that of wild type, this was supposed with a resistant mechanism of CMZ. Also, this acquisition of resistance was suppressed by the addition of REL. Therefore, the combination of CMZ and REL may inhibit resistance for E. coli including the ESBL-producing bacteria. Therefore, further clinical trials are necessary.
Ethical Approval
Because clinical isolates collected in Japanese research groups are used in this study, I hereby declare that the Japanese Ministry of Health, Labour and Welfare has indicated in its guidelines that this study is not subject to ethical review.
Acknowledgments
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Funding
This study was funded internally.
Disclosure
The authors declared that they have no conflicts of interest in this work.
References
1. Paitan Y. Current trends in antimicrobial resistance of Escherichia coli. Curr Top Microbiol Immunol. 2018;416:181–211. PMID: 30088148. doi:10.1007/82_2018_110
2. Trautner BW, Darouiche RO. Role of biofilm in catheter-associated urinary tract infection. Am J Infect Control. 2004;32(3):177–183. PMID: 15153930; PMCID: PMC2963581. doi:10.1016/j.ajic.2003.08.005
3. Riley DK, Classen DC, Stevens LE, Burke JP. A large randomized clinical trial of a silver-impregnated urinary catheter: lack of efficacy and staphylococcal superinfection. Am J Med. 1995;98(4):349–356. PMID: 7709947. doi:10.1016/S0002-9343(99)80313-1
4. Pickard R, Lam T, Maclennan G, et al. Types of urethral catheter for reducing symptomatic urinary tract infections in hospitalised adults requiring short-term catheterisation: multicentre randomised controlled trial and economic evaluation of antimicrobial- and antiseptic-impregnated urethral catheters (the CATHETER trial). Health Technol Assess. 2012;16(47):1–197. PMID: 23199586. doi:10.3310/hta16470
5. Yougbaré S, Mutalik C, Okoro G, et al. Emerging trends in nanomaterials for antibacterial applications. Int J Nanomedicine. 2021;16:5831–5867. PMID: 34475754; PMCID: PMC8405884. doi:10.2147/IJN.S328767
6. Doi Y, Iovleva A, Bonomo RA. The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world. J Travel Med. 2017;24(suppl_1):S44–S51. doi:10.1093/jtm/taw102
7. Toner L, Papa N, Aliyu SH, Dev H, Lawrentschuk N, Al-Hayek S. Extended-spectrum beta-lactamase-producing Enterobacteriaceae in hospital urinary tract infections: incidence and antibiotic susceptibility profile over 9 years. World J Urol. 2016;34(7):1031–1037. PMID: 26511749. doi:10.1007/s00345-015-1718-x
8. Critchley IA, Cotroneo N, Pucci MJ, Jain A, Mendes RE. Resistance among urinary tract pathogens collected in Europe during 2018. J Glob Antimicrob Resist. 2020;23:439–444. PMID: 33212286. doi:10.1016/j.jgar.2020.10.020
9. Kobayashi K, Yamamoto S, Takahashi S, et al. The third national Japanese antimicrobial susceptibility pattern surveillance program: bacterial isolates from complicated urinary tract infection patients. J Infect Chemother. 2020;26(5):418–428. PMID: 32081647. doi:10.1016/j.jiac.2020.01.004
10. Chong Y, Shimoda S, Yakushiji H, et al. Community spread of extended-spectrum β-lactamase-producing Escherichia coli, Klebsiella pneumoniae and Proteus mirabilis: a long-term study in Japan. J Med Microbiol. 2013;62(Pt7):1038–1043. PMID: 23538565. doi:10.1099/jmm.0.059279-0
11. Doi Y, Park YS, Rivera JI, et al. Community-associated extended-spectrum β-lactamase-producing Escherichia coli infection in the United States. Clin Infect Dis. 2013;56(5):641–648. PMID: 23150211; PMCID: PMC3563390. doi:10.1093/cid/cis942
12. Tamma PD, Aitken SL, Bonomo RA, Mathers AJ, van Duin D, Clancy CJ. Infectious Diseases Society of America guidance on the treatment of extended-spectrum β-lactamase producing Enterobacterales (ESBL-E), carbapenem-resistant Enterobacterales (CRE), and Pseudomonas aeruginosa with difficult-to-treat resistance (DTR-P. aeruginosa). Clin Infect Dis. 2021;72(7):e169–e183. PMID: 33106864. doi:10.1093/cid/ciaa1478
13. Vardakas KZ, Tansarli GS, Rafailidis PI, Falagas ME. Carbapenems versus alternative antibiotics for the treatment of bacteraemia due to Enterobacteriaceae producing extended-spectrum β-lactamases: a systematic review and meta-analysis. J Antimicrob Chemother. 2012;67(12):2793–2803. PMID: 22915465. doi:10.1093/jac/dks301
14. Pangon B, Bizet C, Buré A, et al. In vivo selection of a cephamycin-resistant, porin-deficient mutant of Klebsiella pneumoniae producing a TEM-3 beta-lactamase. J Infect Dis. 1989;159(5):1005–1006. PMID: 2651531. doi:10.1093/infdis/159.5.1005
15. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing.
16. Chetri S, Singha M, Bhowmik D, et al. Transcriptional response of OmpC and OmpF in Escherichia coli against differential gradient of carbapenem stress. BMC Res Notes. 2019;12(1):138. PMID: 30871640; PMCID: PMC6419367. doi:10.1186/s13104-019-4177-4
17. Haldorsen B, Aasnaes B, Dahl KH, et al. The AmpC phenotype in Norwegian clinical isolates of Escherichia coli is associated with an acquired ISEcp1-like ampC element or hyperproduction of the endogenous AmpC. J Antimicrob Chemother. 2008;62(4):694–702. PMID: 18583329. doi:10.1093/jac/dkn257
18. Paltansing S, Tengeler AC, Kraakman ME, Claas EC, Bernards AT. Exploring the contribution of efflux on the resistance to fluoroquinolones in clinical isolates of Escherichia coli. Microb Drug Resist. 2013;19(6):469–476. PMID: 23909485. doi:10.1089/mdr.2013.0058
19. Tomás M, Doumith M, Warner M, et al. Efflux pumps, OprD porin, AmpC beta-lactamase, and multiresistance in Pseudomonas aeruginosa isolates from cystic fibrosis patients. Antimicrob Agents Chemother. 2010;54(5):2219–2224. PMID: 20194693; PMCID: PMC2863613. doi:10.1128/AAC.00816-09
20. Livermore DM, Warner M, Mushtaq S. Activity of MK-7655 combined with imipenem against Enterobacteriaceae and Pseudomonas aeruginosa. J Antimicrob Chemother. 2013;68(10):2286–2290. PMID: 23696619. doi:10.1093/jac/dkt178
21. Young K, Painter RE, Raghoobar SL, et al. In vitro studies evaluating the activity of imipenem in combination with relebactam against Pseudomonas aeruginosa. BMC Microbiol. 2019;19(1):150. PMID: 31272373; PMCID: PMC6610938. doi:10.1186/s12866-019-1522-7
22. Livermore DM. Beta-lactamases in laboratory and clinical resistance. Clin Microbiol Rev. 1995;8(4):557–584. PMID: 8665470; PMCID: PMC172876. doi:10.1128/CMR.8.4.557
23. Matsumura Y, Yamamoto M, Nagao M, Tanaka M, Takakura S, Ichiyama S. In vitro activities and detection performances of cefmetazole and flomoxef for extended-spectrum β-lactamase and plasmid-mediated AmpC β-lactamase-producing Enterobacteriaceae. Diagn Microbiol Infect Dis. 2016;84(4):322–327. PMID: 26782634. doi:10.1016/j.diagmicrobio.2015.12.001
24. Gutiérrez-Gutiérrez B, Pérez-Galera S, Salamanca E, et al. A multinational, preregistered cohort study of β-Lactam/β-Lactamase inhibitor combinations for treatment of bloodstream infections due to extended-spectrum-β-lactamase-producing Enterobacteriaceae. Antimicrob Agents Chemother. 2016;60(7):4159–4169. PMID: 27139473; PMCID: PMC4914653. doi:10.1128/AAC.00365-16
25. Tsutsui A, Yahara K, Shibayama K. Trends and patterns of national antimicrobial consumption in Japan from 2004 to 2016. J Infect Chemother. 2018;24(6):414–421. PMID: 29428566. doi:10.1016/j.jiac.2018.01.003
26. Ghai I, Ghai S. Understanding antibiotic resistance via outer membrane permeability. Infect Drug Resist. 2018;11:523–530. PMID: 29695921; PMCID: PMC5903844. doi:10.2147/IDR.S156995
27. Nikaido H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol Mol Biol Rev. 2003;67(4):593–656. PMID: 14665678; PMCID: PMC309051. doi:10.1128/MMBR.67.4.593-656.2003
28. Nikaido H, Rosenberg EY, Foulds J. Porin channels in Escherichia coli: studies with beta-lactams in intact cells. J Bacteriol. 1983;153(1):232–240. PMID: 6294048; PMCID: PMC217361. doi:10.1128/jb.153.1.232-240.1983
29. Phoenix DA. On the targeting and membrane assembly of the Escherichia coli outer membrane porin, PhoE. FEMS Immunol Med Microbiol. 1996;16(2):77–82. PMID: 8988389. doi:10.1111/j.1574-695X.1996.tb00125.x
30. Jacoby GA, Carreras I. Activities of beta-lactam antibiotics against Escherichia coli strains producing extended-spectrum beta-lactamases. Antimicrob Agents Chemother. 1990;34(5):858–862. doi:10.1128/AAC.34.5.858
31. Delcour AH. Outer membrane permeability and antibiotic resistance. Biochim Biophys Acta. 2009;1794(5):808–816. PMID: 19100346; PMCID: PMC2696358. doi:10.1016/j.bbapap.2008.11.005
32. Ananthan S, Subha A. Cefoxitin resistance mediated by loss of a porin in clinical strains of Klebsiella pneumoniae and Escherichia coli. Indian J Med Microbiol. 2005;23(1):20–23. PMID: 15928416. doi:10.4103/0255-0857.13867
33. Rowbury RJ, Goodson M, Wallace AD. The PhoE porin and transmission of the chemical stimulus for induction of acid resistance (acid habituation) in Escherichia coli. J Appl Bacteriol. 1992;72:233–243. doi:10.1111/j.1365-2672.1992.tb01829.x
34. Berrier C, Coulombe A, Houssin C, Ghazi A. Voltage-dependent cationic channel of Escherichia coli. J Membr Biol. 1993;133:119–127. doi:10.1007/BF00233793
35. Rodríguez-Baño J, Gutiérrez-Gutiérrez B, Machuca I, Pascual A. Treatment of infections caused by extended-spectrum-beta-lactamase-, AmpC-, and carbapenemase-producing Enterobacteriaceae. Clin Microbiol Rev. 2018;31(2):e00079–17. doi:10.1128/CMR.00079-17
36. Jacoby GA. AmpC beta-lactamases. Clin Microbiol Rev. 2009;22:161–182. doi:10.1128/CMR.00036-08
37. Nelson EC, Elisha BG. Molecular basis of AmpC hyperproduction in clinical isolates of Escherichia coli. Antimicrob Agents Chemother. 1999;43(4):957–959. PMID: 10103209; PMCID: PMC89235. doi:10.1128/AAC.43.4.957
38. Caroff N, Espaze E, Gautreau D, Richet H, Reynaud A. Analysis of the effects of −42 and −32 ampC promoter mutations in clinical isolates of Escherichia coli hyperproducing ampC. J Antimicrob Chemother. 2000;45(6):783–788. PMID: 10837430. doi:10.1093/jac/45.6.783
39. Bialek-Davenet S, Marcon E, Leflon-Guibout V, et al. In vitro selection of ramR and soxR mutants overexpressing efflux systems by fluoroquinolones as well as cefoxitin in Klebsiella pneumoniae. Antimicrob Agents Chemother. 2011;55(6):2795–2802. PMID: 21464248; PMCID: PMC3101381. doi:10.1128/AAC.00156-11
40. Shlaes DM. New β-lactam-β-lactamase inhibitor combinations in clinical development. Ann N Y Acad Sci. 2013;1277:105–114. PMID: 23346860. doi:10.1111/nyas.12010
41. Goldstein EJC, Citron DM, Tyrrell KL, Leoncio E, Merriam CV. Comparative in vitro activities of relebactam, imipenem, the combination of the two, and six comparator antimicrobial agents against 432 strains of anaerobic organisms, including imipenem-resistant strains. Antimicrob Agents Chemother. 2018;62(2):e01992–17. PMID: 29158284; PMCID: PMC5786751. doi:10.1128/AAC.01992-17
42. Yin H, Guiying L, Chen X, et al. Accelerated evolution of bacterial antibiotic resistance through early emerged stress responses driven by photocatalytic oxidation. Appl Catal B. 2020;269:118829. doi:10.1016/j.apcatb.2020.118829
43. Chen M, Cai Y, Guiying L, Zhao H, Taicheng AN. The stress response mechanisms of biofilm formation under sub-lethal photocatalysis. Appl Catal B. 2022;307:121200. doi:10.1016/j.apcatb.2022.121200
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.