Back to Journals » OncoTargets and Therapy » Volume 12
CD4+ CD25+ regulatory T cells promote hepatocellular carcinoma invasion via TGF-β1-induced epithelial–mesenchymal transition
Authors Shi C, Chen Y, Chen Y, Yang Y, Bing W, Qi J
Received 27 April 2018
Accepted for publication 18 September 2018
Published 28 December 2018 Volume 2019:12 Pages 279—289
DOI https://doi.org/10.2147/OTT.S172417
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr William C. Cho
Chunying Shi,1 Ying Chen,1 Yaodong Chen,1 Yuchuan Yang,1 Wang Bing,1 Jiping Qi2
1Department of Radiology, The First Affiliated Hospital of Harbin Medical University, Heilongjiang, Harbin 150001, China; 2Department of Pathology, The First Affiliated Hospital of Harbin Medical University, Heilongjiang, Harbin 150001, China
Background: CD4+ CD25+ regulatory T cells (Tregs), a crucial component of the infiltration of immune cells in tumor microenvironment, are associated with progression and metastasis of hepatocellular carcinoma (HCC).
Methods: The mechanism of Tregs in the invasion and metastasis of HCC was investigated in vivo and in vitro using immunohistochemical analysis, western blot, and quantitative reverse transcription-PCR (qRT-PCR).
Results: Analysis of 78 clinical HCC samples indicated that high expression of Tregs was strongly associated with poor cancer-free survival and overall survival of patients. The reduced expression of E-cadherin and enhanced expression of Vimentin and transforming growth factor-beta 1 (TGF-β1) were found in HCC tissue compared with normal liver tissue. The HCC Hepa1-6 cells were treated with the supernatant of Tregs-conditioned medium (Tregs-CM) to investigate the epithelial-mesenchymal transition (EMT) and TGF-β1. Western blot and qRT-PCR also showed that down-regulated E-cadherin and up-regulated Vimentin and TGF-β1 were found in Tregs-CM-treated Hepa1-6 cells. An experiment of tumorigenicity in C57 mice showed larger and heavier tumors in Tregs-CM-treated group than in the control group. Tregs produced higher TGF-β1 compared with Tregs treated with FOXP3 shRNA. TGF-β1 with neutralizing antibodies was used to deplete TGF-β1 in Tregs-CM, which enhanced expression of E-cadherin, reduced expression of Vimentin and TGF-β1, and decreased migratory and invasive capacity of Hepa1-6 cells.
Conclusion: Tregs could promote the invasion and migration of Hepa1-6 cells, which are possibly maintained by TGF-β1-induced EMT. This study showed that the development of therapeutic strategies against TGF-β1 pathway is valuable in HCC therapy.
Keywords: hepatocellular carcinoma, CD4+ CD25+ regulatory T cells, epithelial to mesenchymal transition, TGF-β1
Introduction
Hepatocellular carcinoma (HCC) is one of the most common and aggressive human malignancies in the world.1 It is the third most common cause of cancer-related death in adults.2 In total, 750,000 new cases are generated globally each year. In China, new cases every year account for half of all global cases in total.3 Hepatic resection, liver transplantation, and various minimally invasive therapies are widely used to improve the outcomes in patients with HCC. Nevertheless, due to recurrence and metastasis, the 5-year survival of patients undergoing therapy remains disappointingly low.4,5 Recent studies have found that two mechanisms of metastasis are particularly important for HCC, including the tumor microenvironment and the epithelial–mesenchymal transition (EMT).6–10
The tumor microenvironment is a systematic concept that defines the behavior of cancer, not only by the genetics of the tumor cells but also by the surrounding milieu, which plays an important role in the development of tumors.11,12 Recent studies unambiguously show that the cross talk between tumor cells and their surrounding microenvironment is necessary for cell survival, growth, proliferation, EMT, and metastasis.13–17 The tumor microenvironment is mainly composed of tumor cells and a variety of tumor stromal cells, cytokines and chemokines, immune cells, and their mediators presenting in the tumor microenvironment of most tumors. CD4+ CD25+ regulatory T cells (Tregs) are T-cell subsets with immunosuppressive functions. At present, many studies have supported the ability of Tregs to promote HCC recurrence and metastasis by suppressing immune surveillance, which is important for the tumor microenvironment and responsible for poor prognosis.18–21 Nevertheless, the mechanism of Tregs in the development of HCC is not clear enough.
EMT is a process by which epithelial tumor cells lose epithelial features and gain mesenchymal phenotypes.22 It is considered a key step in tumor cells gaining higher invasive and metastatic ability. TGF-β1 is one of the most important members of the TGF-β family. It is a potent inducer of epithelial plasticity leading to EMT in cancer cells.23,24 Increasing numbers of studies have demonstrated the importance of TGF-β1 in cancer progression and metastasis. The release of TGF-β1 in the tumor microenvironment can promote the growth and migration of tumor cells, which can induce epithelial interstitial transformation, inhibit the expression of antiapoptotic genes, help the tumor escape immune surveillance, and promote the production of genes for invasion and angiogenesis.25
In this study, the role of Tregs in HCC progression was investigated with the focus on TGF-β1. It was found that the expression of Tregs was significantly increased in HCC and was associated with tumor metastasis and poor prognosis. In HCC samples, the expression and transformation of epithelial mesenchymal markers were correlated with Tregs. After being treated with the supernatant from cultured Tregs, HCC cells showed higher invasive activity. The expression of EMT markers, the invasive ability, and tumorigenicity of these cells were all increased. Neutralization of TGF-β1 inhibited the invasive activity by limiting TGF-β1-induced EMT in hepatoma cells. It was demonstrated that Tregs promote migration and invasion by TGF-β1-induced EMT in hepatoma cells.
Materials and methods
Patients and specimens
From May 2000 to February 2010, 78 patients who underwent curative liver resection and pathology-proven HCC were investigated. Tumor stage was determined according to the 2009 International Union for International Cancer Control TNM Classification of Malignant Tumors (seventh edition). The available characteristics of patients are shown in Table 1. This study was approved by the Research Ethics Committee of The First Affiliated Hospital of Harbin Medical University. The patients whose tissues were used in this research have provided the written informed consents. All specimens were obtained from the archives of formalin-fixed, paraffin tissue blocks in the Department of Pathology. Overall survival (OS) was defined as the period between the date of surgery and death, or date of last contact of living patients. Disease-free survival (DFS) was defined as the interval from the date of surgery to the first appearance of recurrence or death, which ever occurred first, or the date of the last follow-up.
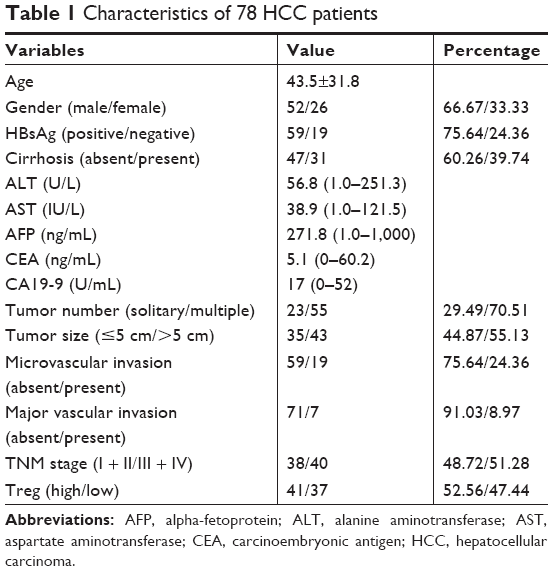 | Table 1 Characteristics of 78 HCC patients |
Conditioned medium and cell culture
The CD4+ CD25+ Regulatory T Cell Isolation Kit (ID: 130-091-041, Miltenyi, Bergisch Gladbach, Germany) was used to isolate Tregs from the spleen of mice. Tregs were cultured in RPIM-1640 (Gibco-BRL, Gaithersburg, MD, USA) with 10% FBS for 24 hours, then changed to serum-free medium for 24 hours. The supernatant of Tregs-conditioned medium (Tregs-CM) was collected by centrifugation. Tregs-CM was incubated with 1 μg/mL TGF-β1-neutralizing antibody (mouse monoclonal anti-TGF-β1, [9016] (ab64715); Abcam) for 24 hours at 4°C to obtain TGF-β1-depleted Tregs-CM. A total of 5×105 HCC Hepa1–6 cells/well (Shanghai Cell Bank of the Chinese Academy of Sciences) were precultured in 2 mL DMEM (Gibco-BRL) with 10% FBS in 6-well plates for 24 hours. Then, the culture medium was replaced with 1 mL Tregs-CM and 1 mL complete medium per well for 24 hours. The treated Hepa1–6 cells were used in the subsequent experiment.
ELISA
The procedures were completed using the methodology previously described.26 FOXP3 shRNA was first transfected into Tregs cells by ultrasound-targeted microbubble destruction and Lipofectamine 2000 for 24 hours. The supernatant was collected by centrifugation. The TGF-β1 concentration secreted by Tregs was measured by ELISA kits (BG, Shanghai, China), according to the manufacturer’s instructions.
Mice and in vivo tumorigenicity experiments
Six-week-old male C57 mice were purchased from the Experimental Animal Center of First Affiliated Hospital of Harbin Medical University. All animal experiments were conducted according to the University’s Laboratory Animal Center Care Guidelines and were approved by this committee. Twenty mice were randomized into two groups. The suspensions of cells were injected subcutaneously into the right back at 1×106 cells/injection site. Mice were sacrificed by cervical dislocation 4 weeks after the injection. Tumors were isolated for analysis of weight and size.
Colony formation assay
A total of 1×103 Hepa1–6 cells per well were seeded into a 6-well culture dish. After incubation at 37°C for 2 weeks, the cells were washed twice with PBS and stained with 0.1% crystal violet solution. The number of colonies containing P50 cells was counted under a microscope. The experiments were performed in triplicate.
Wound-healing assay
A total of 5×104 Hepa1–6 cells were seeded in 24-well plates and incubated for 24 hours, then the monolayer cells were disrupted by scratching with a 10 μL microsterile pipette tip. Photographs were taken at 0, 24, and 48 hours in a phase-contrast microscope. The assays were performed in triplicate, and four fields of each point were measured.
Transwell invasion assay
A total of 5×104 Hepa1–6 cells in 200 μL serum-free medium from different groups were seeded in the upper chamber. Five hundred microliters medium supplemented with 10% FBS was added to the lower compartment. After incubation for 24 hours, the cells that migrated to the lower surface of the membrane were fixed with 4% paraformaldehyde, stained with 0.1% crystal violet, and counted under a microscope. All these samples were plated three times.
Quantitative real-time polymerase chain reaction
Total RNA of Hepa1–6 cells was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s protocol. Expression of mRNA was determined by real-time PCR using SYBR Green Master Mix (TaKaRa, Japan). The results were normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The primer sequences are shown in Table 2.
 | Table 2 Primer sequences used in the work |
Western blot analysis
Western blot was performed as previously described.26 The protein of Hepa1–6 cells was extracted by RIPA Lysis Buffer (Beyotime, Haimen, China) with PMSF protease inhibitor (Cwbiotech, Beijing, China), according to the manufacturers’ instructions. Primary antibodies were rabbit anti-mouse polyclonal FOXP3 antibody (236A/E7, ab20034, 5 μg/mL; Abcam), E-cadherin (M168, ab76055, 1:1,000; Abcam), vimentin (EPR3776, ab92547, 1:5,000; Abcam), and SNAIL (ab53519, 1:500; Abcam), using GAPDH (1:5,000; Bioworld Technology, Nanjing, China) as a control. These primary antibodies were detected with goat polyclonal secondary antibody to rabbit IgG (1:10,000; Abcam). Proteins were detected by the enhanced chemiluminescence detection reagent.
Immunohistochemical analysis
The staining was performed on formalin-fixed, paraffin tissue blocks of HCC. Primary antibodies were rabbit anti-mouse FOXP3 polyclonal antibody (236A/E7, ab20034, 5 μg/mL; Abcam), E-cadherin (M168, ab76055, 5 μg/mL; Abcam), vimentin (EPR3776, ab92547, 1:200; Abcam), SNAIL (ab53519, 1:1,000; Abcam), and horseradish peroxidase-conjugated streptavidin (Zhongshan Jingqiao Company, Beijing, China).
Statistical analysis
Statistical analysis was performed with SPSS 20.0. All data are shown as mean ± standard error of the mean. The differences between two groups were analyzed by unpaired Student’s t-test. For clinical samples, analysis of Kaplan–Meier curves was performed to determine survival. P<0.05 was considered statistically significant.
Results
Expression of Tregs in HCC patient tissue and analysis of prognosis
Immunohistochemical analysis showed that the expression of Tregs in the tissue of HCC patients was significantly higher than in normal liver tissue, with statistical significance (14.76±6.23 vs 3.85±2.58 a/HP, P<0.05, Figure 1A). FOXP3 is a specific molecular marker of Tregs, which plays an important role in the development of Tregs and their inhibitory functions. According to the results of Western blot and quantitative reverse transcription-PCR (qRT-PCR), the FOXP3 protein expression and the FOXP3 mRNA expression in HCC tumor tissue were both higher than in normal hepatic tissue (Figure 1B and C). To evaluate the prognostic value of Tregs in HCC patients, tissue samples from 78 patients with HCC were collected. Positive expression of Tregs was detected in 41 of 78 HCC cases (52.56%), showing predominant staining of interstitial lymphocytes. It showed that the positive expression of Tregs was an independent prognostic factor for both OS and DFS. The DFS and OS rates for patients with low expression of Tregs were significantly higher than those with high expression of Tregs (Figure 1D and E). These results demonstrated that high density of Tregs could be an important risk factor for HCC.
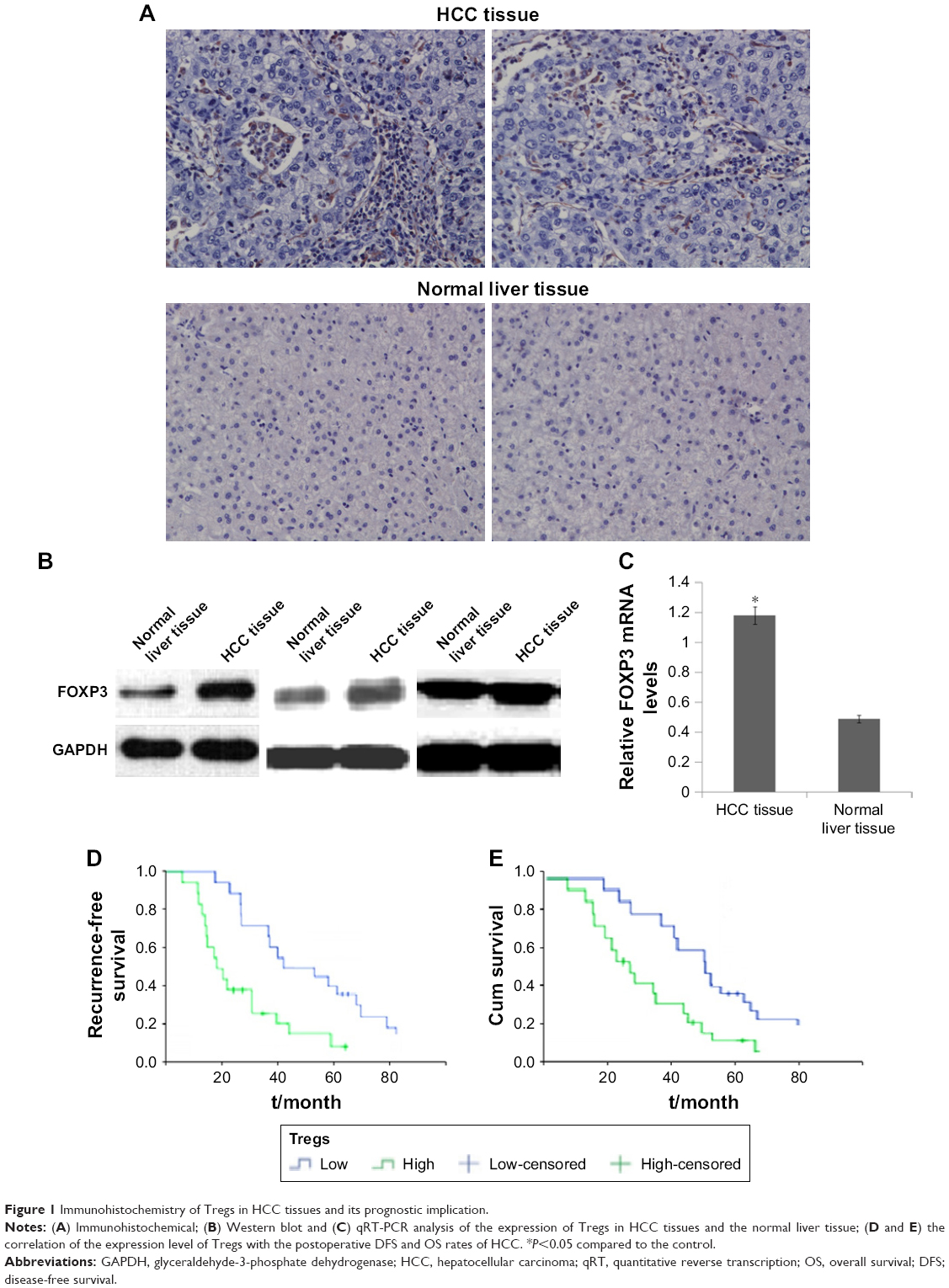 | Figure 1 Immunohistochemistry of Tregs in HCC tissues and its prognostic implication. |
Expression of EMT markers and TGF-β1 in HCC tissue and relationship with Tregs
Immunohistochemical analysis showed that the expression of epithelial marker of E-cadherin in the HCC cell membrane and cytoplasm was reduced, while the expression of the mesenchymal marker of vimentin in the cytoplasm and the expression of SNAIL (EMT-associated TGF-β1) in the cytoplasm or nucleus were increased (Figure 2A). Western blot and qRT-PCR results also indicated reduced expression of E-cadherin and enhanced expression of vimentin and SNAIL in HCC tissue compared with those in normal liver tissue (Figure 2B and C). Spearman correlation analysis showed that E-cadherin was negatively correlated with vimentin and SNAIL (vimentin: r=−0.557, P<0.05; SNAIL: r=−0.612, P<0.05). Tregs were mainly expressed in the stroma, with numbers negatively correlated with E-cadherin (r=−0.513, P<0.05) and positively correlated with vimentin and SNAIL (vimentin: r=0.607, P<0.01; SNAIL: r=0.518, P<0.05).
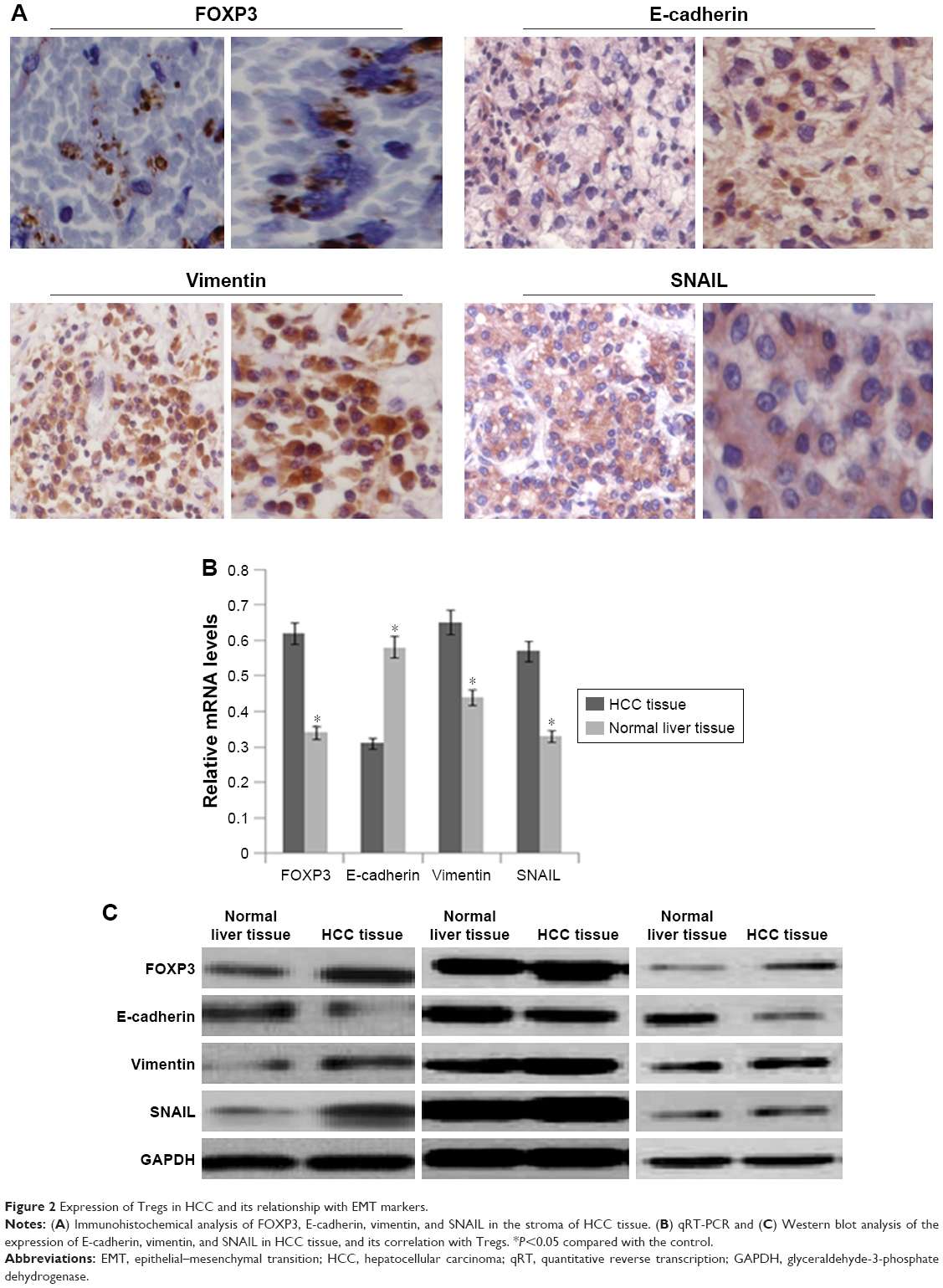 | Figure 2 Expression of Tregs in HCC and its relationship with EMT markers. |
Tregs induce EMT and promotion of invasive capability of Hepa1–6 cells
Western blot and qRT-PCR were conducted to analyze the expression of EMT markers and TGF-β1 in Tregs-CM-treated Hepa1–6 cells. As shown in Figure 3A and B, the expression of E-cadherin was reduced, while the expression of vimentin and SNAIL was upregulated in Tregs-CM-treated Hepa1–6 cells. To investigate whether Tregs-CM could promote the invasive and migratory abilities of Hepa1–6 cells, transwell migration assay and wound-healing assay were conducted. Compared with the control, Tregs-CM-treated Hepa1–6 cells showed faster closure of the wound. The result was confirmed by transwell assay (Figure 3C and D). In addition, an experiment of the tumorigenicity of C57 mice was adopted to evaluate whether Tregs could regulate hepatoma cells in vivo. Suspension of Tregs-CM-treated Hepa1–6 cells and unpretreated control cells was injected subcutaneously into the left axilla of mice to form tumors. The effects of Tregs were evaluated by the volume and weight of the tumors. As a result, the tumors of the Tregs-CM-treated group were larger and heavier than those of the control group (Figure 3E). Thus, Tregs showed a significant promotion of the growth of tumors in mice. The results indicated that Tregs promoted migratory and invasive behaviors of Hepa1–6 cells in vitro and in vivo, which could be through the mediation of EMT.
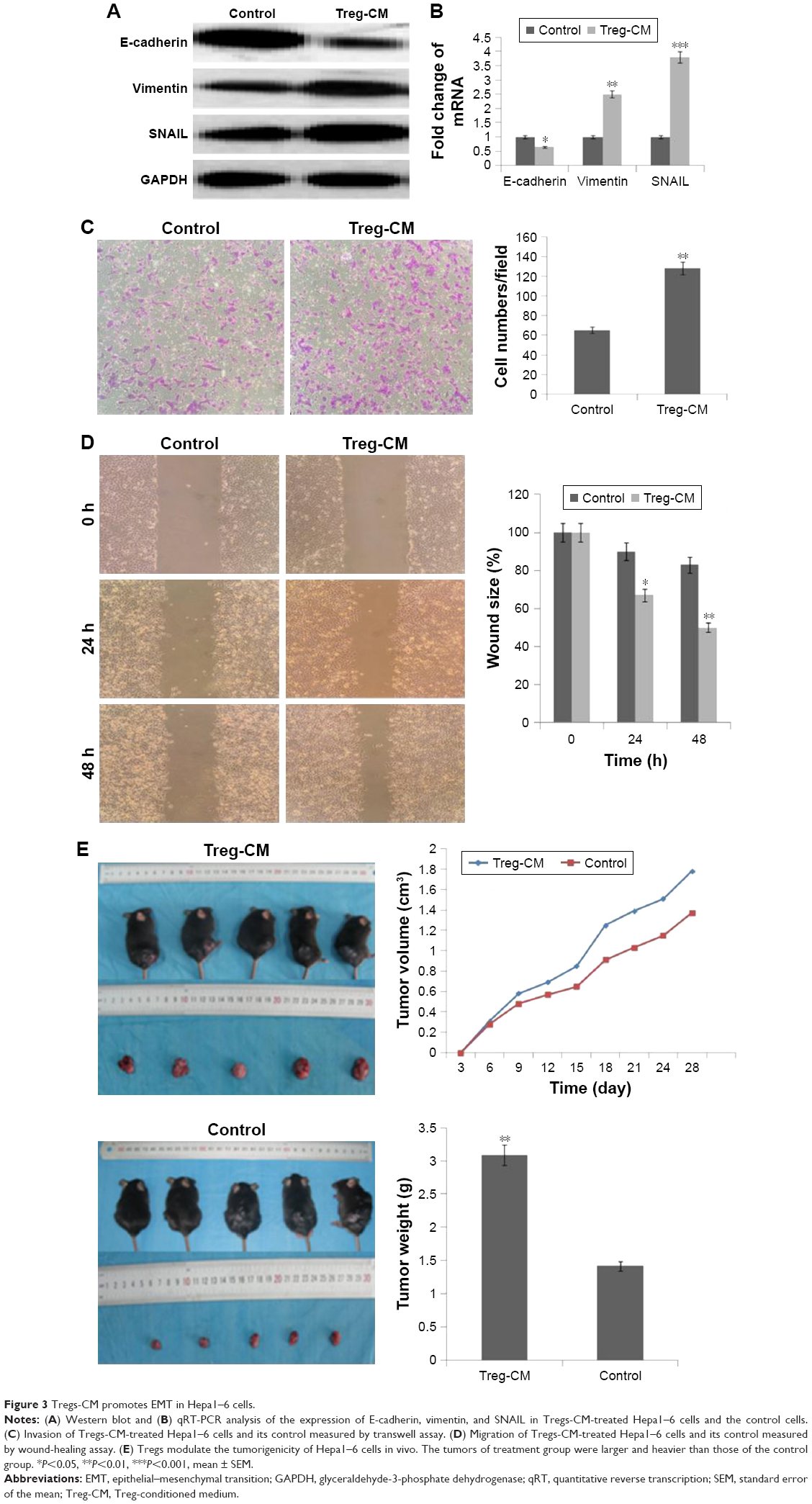 | Figure 3 Tregs-CM promotes EMT in Hepa1–6 cells. |
TGF-β1-neutralizing antibody inhibits the effect of TGF-β1-induced EMT in Hepa1–6 cells
It has been previously accepted that TGF-β1, which can be secreted by Tregs, plays a major role in tumor progression and malignancy by mediating hepatocyte plasticity, facilitating EMT, and secreting some proteins that act on the tumor microenvironment.27 To understand the mechanism by which Treg-CM promoted invasive properties of Hepa1–6 cells, TGF-β1 secreted by Tregs and its mRNA level was measured by qRT-PCR (Figure 4A) and ELISA (Figure 4B). Tregs control showed a higher production of TGF-β1 compared with Tregs treated with FOXP3 shRNA (Figure 4). The knockdown efficiency of FOXP3 shRNA was about 30%–40%. Subsequently, to determine whether TGF-β1 could have effects on Tregs-mediated EMT in hepatoma cells, TGF-β1-neutralizing antibody was used to deplete TGF-β1 in Tregs-CM. As shown in Figure 5A, after using TGF-β1-neutralizing antibody in Treg-CM, the colony formation ability of Hepa1–6 cells was significantly lower than the cells treated with Tregs-CM. According to the results of Western blot and qRT-PCR (Figure 5B and C), compared with Tregs-CM-treated cells, the upregulated E-cadherin and the downregulated vimentin and SNAIL were detected in Hepa1–6 cells cocultured with TGF-β1-depleted Tregs-CM. Transwell assay and wound-healing assay indicated that the depletion of TGF-β1 in Tregs-CM decreased the migratory and invasive capacities of Hepa1–6 cells in vitro (Figure 5D and E). The results suggested that depletion of TGF-β1 could block the effect of Tregs-induced EMT, which could be a potential reason to promote the invasive migration of Hepa1–6 cells. Nevertheless, there might be other factors that need further investigation.
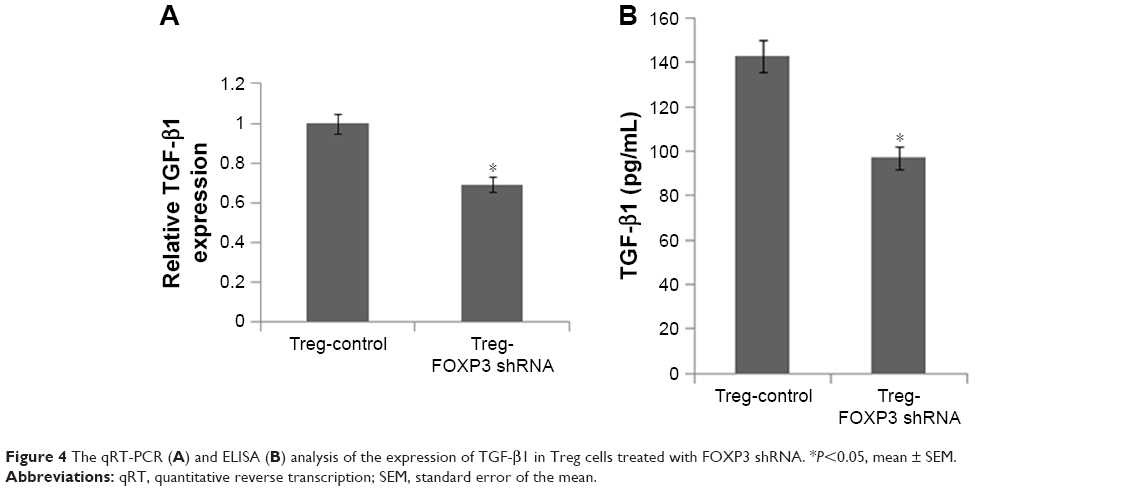 | Figure 4 The qRT-PCR (A) and ELISA (B) analysis of the expression of TGF-β1 in Treg cells treated with FOXP3 shRNA. *P<0.05, mean ± SEM. |
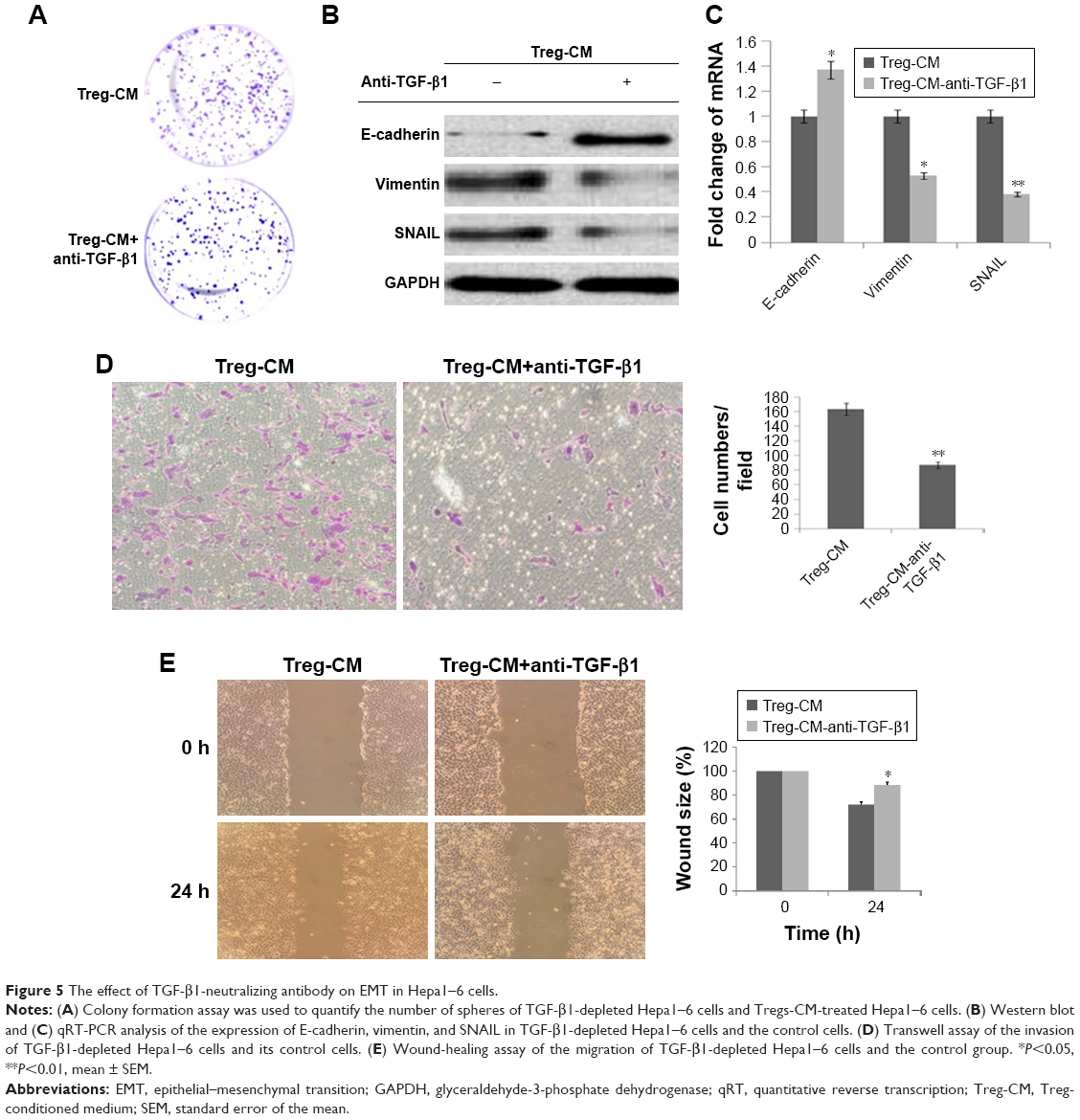 | Figure 5 The effect of TGF-β1-neutralizing antibody on EMT in Hepa1–6 cells. |
Discussion
The critical role of the tumor microenvironment in modulating tumorigenesis, EMT, tumor invasion, and metastasis has been widely accepted. The tumor microenvironment consists of stromal cells, including carcinoma-associated fibroblasts, mesenchymal cells, endothelial cells, and various immune cells.28 As a main component of tumor-infiltrating leukocytes, Tregs play a decisive role in tumor progression through the expression of cytokines, chemokines, growth factors, and matrix metalloproteases. A better understanding of the underlying mechanism of the regulation of Tregs in tumor initiation and development is needed.
EMT is a key process for dissemination and metastasis of cancer cells.29 Moreover, cells that have undergone an EMT are acting in many respects like cancer stem cells.30 Vimentin is one of the type III intermediate filament proteins that are normally found in mesenchymal cells, which can induce the mesenchymal phenotype.31 The loss of epithelial E-cadherin and the gain of mesenchymal vimentin is a major hallmark of EMT. In the work, immunohistochemical analysis, Western blot, and qRT-PCR results indicated the reduced expression of E-cadherin and enhanced expression of vimentin in HCC tissue compared with normal liver tissue. Moreover, Western blot and qRT-PCR also showed that the expression of E-cadherin was reduced, while the expression of vimentin was upregulated in Tregs-CM-treated Hepa1–6 cells. Thus, it was speculated that Hepa1–6 cells treated with Tregs-CM underwent EMT, which increased the migratory and invasive abilities (Figure 3C and D). As demonstrated by the tumorigenicity experiment in C57 mice, larger and heavier tumors were found in the Tregs-CM-treated group than those in the control group. Thus, Tregs might promote invasive migration of HCC by EMT.
EMT is modulated by several transcription factors, such as EMT-associated TGF-β1 of SNAIL.32–34 TGF-β1 serves as a central regulator in HCC and inflammation.35 The accumulated evidence about cancer demonstrates that TGF-β1 may contribute to the impaired differentiation of stem cells and allow for the development of cancer. It is known that TGF-β1 is a potent EMT inducer in cancer.32–35 Many studies have shown that TGF-β1 can induce the occurrence of EMT in a wide variety of cancer cells.36–39 In the present work, EMT-associated TGF-β1 of SNAIL was upregulated in HCC tissues and Tregs-CM-treated Hepa1–6 cells. Compared with Tregs treated with FOXP3 shRNA, Tregs showed a higher production of TGF-β1. Tregs might promote the invasive capability of Hepa1–6 cells by TGF-β1-induced EMT. To determine whether TGF-β1 affected the Tregs-mediated EMT in hepatoma cells, TGF-β1-neutralizing antibody was used to deplete TGF-β1 in Tregs-CM. The colony formation ability of Hepa1–6 cells was significantly lower than in cells treated with Tregs-CM. The upregulated E-cadherin, the downregulated vimentin, and SNAIL were detected in Hepa1–6 cells cocultured with TGF-β1-depleted Tregs-CM compared with Tregs-CM-treated cells. Moreover, depletion of TGF-β1 in Tregs-CM also decreased the migratory and invasive capacities of Hepa1–6 cells in vitro. It was concluded that depletion of TGF-β1 could block the effect of Tregs-induced EMT and promote the invasive migration of Hepa1–6 cells.
In the present study, the positive expression of Tregs was associated with poor survival in patients after resection of primary tumors. Among these samples, the high expression of Tregs indicated a worse outcome for patients than those with low expression of Tregs. According to current in vitro and in vivo studies, Tregs might promote the invasion and migration of Hepa1–6 cells that were possibly maintained by TGF-β1-induced EMT. Therefore, it was proposed that high density of Tregs could be an important risk factor for HCC.
Conclusion
In summary, this work demonstrates that Tregs regulate the essential characteristic of HCC through the TGF-β1-signaling pathway. Tregs are important factors that have a negative function in regulating the body’s immune system, and Tregs play an important role in the antitumor response. To explore effective methods for inhibiting tumor-infiltrating Tregs and improving the outcomes of HCC, developing therapeutic strategies against the TGF-β1 pathway will provide great values for patients with HCC.
Acknowledgments
This study was supported by General Projects of Heilongjiang Province Natural Science Foundation of China (grant number: H2017026); Project of research foundation of the talent of scientific and technical innovation of Harbin City (grant number: 2016RAQXJ148); and Scientific Research Innovation Fund of The First Affiliated Hospital of Harbin Medical University (grant number: 2017B011).
Disclosure
The authors report no conflicts of interest in this work.
References
Saran U, Humar B, Kolly P, Dufour JF. Hepatocellular carcinoma and lifestyles. J Hepatol. 2016;64(1):203–214. | ||
Forner A, Llovet JM, Bruix J. Hepatocellular carcinoma. Lancet. 2012;379(9822):1245–1255. | ||
Nishikawa H, Osaki Y. Non-B, non-C hepatocellular carcinoma. Int J Oncol. 2013;3(43):1333–1342. | ||
Maluccio M, Covey A. Recent progress in understanding, diagnosing, and treating hepatocellular carcinoma. CA Cancer J Clin. 2012;62(6):394–399. | ||
Liu L, Zhu XD, Wang WQ, et al. Activation of beta-catenin by hypoxia in hepatocellular carcinoma contributes to enhanced metastatic potential and poor prognosis. Clin Cancer Res. 2010;16(10):2740–2750. | ||
Kudo-Saito C, Shirako H, Takeuchi T, Kawakami Y. Cancer metastasis is accelerated through immunosuppression during Snail-induced EMT of cancer cells. Cancer Cell. 2009;15(3):195–206. | ||
Wu Y, Zhou BP. Inflammation: a driving force speeds cancer metastasis. Cell Cycle. 2009;8(20):3267–3273. | ||
Mendoza M, Khanna C. Revisiting the seed and soil in cancer metastasis. Int J Biochem Cell Biol. 2009;41(7):1452–1462. | ||
Melnikova VO, Bar-Eli M. Inflammation and melanoma metastasis. Pigment Cell Melanoma Res. 2009;22(3):257–267. | ||
Erler JT, Weaver VM. Three-dimensional context regulation of metastasis. Clin Exp Metastasis. 2009;26(1):35–49. | ||
Wang H, Chen L. Tumor microenviroment and hepatocellular carcinoma metastasis. J Gastroenterol Hepatol. 2013;28 Suppl 1:43–48. | ||
Mbeunkui F, Johann DJ. Cancer and the tumor microenvironment: a review of an essential relationship. Cancer Chemother Pharmacol. 2009;63(4):571–582. | ||
Yang JD, Nakamura I, Roberts LR. The tumor microenvironment in hepatocellular carcinoma: current status and therapeutic targets. Semin Cancer Biol. 2011;21(1):35–43. | ||
Leonardi GC, Candido S, Cervello M, et al. The tumor microenvironment in hepatocellular carcinoma (review). Int J Oncol. 2012;40(6):1733–1747. | ||
Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454(7203):436–444. | ||
Linehan DC, Goedegebuure PS. CD25+ CD4+ regulatory T-cells in cancer. Immunol Res. 2005;32(1–3):155–168. | ||
Chen KJ, Lin SZ, Zhou L, et al. Selective recruitment of regulatory T cell through CCR6-CCL20 in hepatocellular carcinoma fosters tumor progression and predicts poor prognosis. PLoS One. 2011;6(9):e24671. | ||
Gallimore AM, Simon AK. Positive and negative influences of regulatory T cells on tumour immunity. Oncogene. 2008;27(45):5886–5893. | ||
Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139(5):871–890. | ||
Katsuno Y, Lamouille S, Derynck R. TGF-β signaling and epithelial-mesenchymal transition in cancer progression. Curr Opin Oncol. 2013;25(1):76–84. | ||
Zhang H, Liu L, Wang Y, et al. KLF8 involves in TGF-beta-induced EMT and promotes invasion and migration in gastric cancer cells. J Cancer Res Clin Oncol. 2013;139(6):1033–1042. | ||
Majumdar A, Curley SA, Wu X, et al. Hepatic stem cells and transforming growth factor β in hepatocellular carcinoma. Nat Rev Gastroenterol Hepatol. 2012;9(9):530–538. | ||
Shi C, Zhang Y, Yang H, et al. Combined effect of ultrasound/SonoVue microbubble on CD4(+)CD25(+) regulatory T cells viability and optimized parameters for its transfection. Ultrasonics. 2015;62:97–102. | ||
Massagué J. TGFbeta in cancer. Cell. 2008;134(2):215–230. | ||
Schiavoni G, Gabriele L, Mattei F, Giovanna S, Lucia G, Fabrizio M. The tumor microenvironment: a pitch for multiple players. Front Oncol. 2013;3(3):90. | ||
Brabletz T. EMT and MET in metastasis: where are the cancer stem cells? Cancer Cell. 2012;22(6):699–701. | ||
Mani SA, Guo W, Liao MJ, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133(4):704–715. | ||
Satelli A, Li S. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell Mol Life Sci. 2011;68(18):3033–3046. | ||
Kang Y, Massagué J. Epithelial-mesenchymal transitions: twist in development and metastasis. Cell. 2004;118(3):277–279. | ||
Lin Y, Dong C, Zhou BP. Epigenetic regulation of EMT: the Snail story. Curr Pharm Des. 2014;20(11):1698–1705. | ||
Yilmaz M, Christofori G. EMT, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev. 2009;28(1–2):15–33. | ||
Dooley S, ten Dijke P. TGF-β in progression of liver disease. Cell Tissue Res. 2012;347(1):245–256. | ||
Bhowmick NA, Ghiassi M, Bakin A, et al. Transforming growth factor-beta1 mediates epithelial to mesenchymal transdifferentiation through a RhoA-dependent mechanism. Mol Biol Cell. 2001;12(1):27–36. | ||
Vincent T, Neve EP, Johnson JR, et al. A SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transition. Nat Cell Biol. 2009;11(8):943–950. | ||
Bakin AV, Tomlinson AK, Bhowmick NA, Moses HL, Arteaga CL. Phosphatidylinositol 3-kinase function is required for transforming growth factor beta-mediated epithelial to mesenchymal transition and cell migration. J Biol Chem. 2000;275(47):36803–36810. | ||
Cho HJ, Baek KE, Saika S, Jeong MJ, Yoo J. Snail is required for transforming growth factor-beta-induced epithelial-mesenchymal transition by activating PI3 kinase/Akt signal pathway. Biochem Biophys Res Commun. 2007;353(2):337–343. | ||
Janda E, Lehmann K, Killisch I, et al. Ras and TGF[beta] cooperatively regulate epithelial cell plasticity and metastasis: dissection of Ras signaling pathways. J Cell Biol. 2002;156(2):299–313. | ||
Prunier C, Howe PH. Disabled-2 (Dab2) is required for transforming growth factor beta-induced epithelial to mesenchymal transition (EMT). J Biol Chem. 2005;280(17):17540–17548. | ||
Shintani Y, Maeda M, Chaika N, Johnson KR, Wheelock MJ. Collagen I promotes epithelial-to-mesenchymal transition in lung cancer cells via transforming growth factor-beta signaling. Am J Respir Cell Mol Biol. 2008;38(1):95–104. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.