Back to Journals » Breast Cancer: Targets and Therapy » Volume 17
CD151 Promotes Cancer Progression in Triple-Negative Breast Cancer by Inducing EMT through the MAPK Signaling Pathway
Authors Lv H, Zhang B, Weng X, Li Y, Deng C, Wang R, Shi L, Yin Y
Received 31 January 2025
Accepted for publication 21 May 2025
Published 28 May 2025 Volume 2025:17 Pages 455—470
DOI https://doi.org/10.2147/BCTT.S518760
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Pranela Rameshwar
Haishu Lv,1 Beibei Zhang,1 Xi Weng,1 Youjia Li,2 Chaoxian Deng,1 Rui Wang,2 Lei Shi,3 Yuanqin Yin1
1Cancer Institute, The First Affiliated Hospital of China Medical University, Shenyang, Liaoning Province, People’s Republic of China; 2Department of Pathology, The Fourth Affiliated Hospital of China Medical University, Shenyang, Liaoning Province, People’s Republic of China; 3Department of Ophthalmology, The Affiliated Shengjing Hospital of China Medical University, Shenyang, Liaoning Province, People’s Republic of China
Correspondence: Lei Shi, The Affiliated Shenjing Hospital of China Medical University, Shenyang, 110004, People’s Republic of China, Email [email protected] Yuanqin Yin, Department of Tumor Biotherapy and Cancer Research, the First Affiliated Hospital of China Medical University, Shenyang, 110001, People’s Republic of China, Email [email protected]
Background: Breast cancer has become one of the most prevalent malignant neoplasms among women, poses a significant threat to public health. As a member of the tetraspanin family of proteins, CD151 is implicated in tumor progression and has been shown to regulate various cellular and molecular mechanisms that drive malignancy. However, the specific functions of CD151 in triple-negative breast cancer (TNBC) remain unclear. In this study, we aimed to investigate the pro-tumorigenic role of CD151 in TNBC by focusing on its interaction with integrin α 3β 1, which often forms a complex with CD151.
Methods: Our study first evaluated CD151 expression in clinical samples from TNBC patients and TNBC cell lines by immunohistochemistry and Western blotting analysis. Through RNA interference (RNAi) and constructed overexpressed plasmids, we further validated the impact of CD151 on the migration and invasion of TNBC cells. Then the differentially expressed genes were screened by single-cell RNA sequencing, and these genes were enriched and analyzed. Co-immunoprecipitation studies demonstrated the binding of CD151 with integrin α 3β 1. Western blotting analysis was used to evaluate the expression of proteins related to epithelial-mesenchymal transition (EMT) and Mitogen-activated protein kinase (MAPK) signaling pathway.
Results: CD151 is highly expressed in TNBC tissues and cell lines. It enhanced the migration and invasive ability of TNBC cells by promoting EMT. Co-IP demonstrated the binding of CD151 and integrin α 3β 1. In addition, we found that knockdown of either integrin α 3β 1 or CD151 reduced the migration and invasion of TNBC cells in vitro. Western blot analysis revealed that the CD151-integrin α 3β 1 complex could activate the MAPK signaling pathway in TNBC cells, subsequently leading to EMT of these cells.
Conclusion: Based on our findings, we propose a novel mechanism by which CD151 mediates tumor progression through the initiation of EMT. This suggests that CD151 could be considered a potential therapeutic target for TNBC.
Keywords: CD151, integrin α 3β 1, TNBC, MAPK signaling pathway, EMT
Introduction
Breast cancer poses a significant public health challenge, with metastasis being the leading cause of mortality. While early detection of breast cancer can enhance patient survival rates, it is often diagnosed only after metastasis has occurred.1 Recent epidemiological data reveal that in the United States, breast cancer accounts for 31% of all new cancer cases in women - the highest incidence among malignancies - while constituting 15% of cancer-associated deaths, positioning it as the second most lethal neoplasm.2 Triple negative breast cancer (TNBC) is characterized by the absence of estrogen receptors (ER), progesterone receptors (PR), and human epidermal growth factor receptor 2 (HER2), accounting for approximately 10–15% of all breast cancer cases. TNBC is particularly aggressive, grows more rapidly, and has a higher risk of metastasis and recurrence compared to other breast cancer types, contributing to its elevated morbidity. The underlying mechanisms of this aggressiveness are still not fully understood.3 Chemotherapy remains a vital conventional treatment for TNBC following surgery. Therefore, identifying drivers and predictors of its aggression and metastasis is crucial for gaining insights into the progression of the disease.
Tetraspanins form complexes through interactions with a range of transmembrane and cytosolic proteins, which are essential for their function. These include integrins, growth factor receptors, G-protein-coupled receptors and their associated intracellular heterotrimeric G-proteins, several peptidases, transmembrane proteins linked to tumor progression, immunoglobulin superfamily members and cytosolic signal transduction molecules.4 Tumor progression depends on the interaction between the tumor and its microenvironment. In the tumor immune microenvironment, Tetraspanin-24 (CD151) is highly expressed in both tumor cells and fibroblasts.5 CD151 regulates the post-adhesion events, ie, cell spreading, migration, and invasion.6 The molecular mechanism of CD151 in cancer, as with other tetraspanins, primarily involves meditating interactions with proteins such as: laminin-binding integrins (α3β1, α6β1, etc), growth factor receptors such as hepatocyte growth factor, epidermal growth factor receptor (EGFR), and transforming growth factor-β, and matrix metalloproteinases (MMP-2, MMP-7, and MMP-9). This highlights its significance in tumor development.7,8
Integrins are heterodimeric cell surface adhesion receptors composed of α- and β-subunits that bridge the extracellular matrix to the cytoskeleton. Binding of extracellular matrix ligands to integrins activates adhesion-dependent intracellular signaling pathways which are essential for tumor cell survival and invasion.9,10 Integrin-regulated cell migration is primarily mediated by signaling pathways involving members of the focal adhesion kinase (FAK)-SRC family of kinases.11 Activation of the FAK/SRC complex facilitates a multitude of vital signaling cascades associated with regulating cell motility.12,13 Moreover, integrins facilitate cell survival through additional environment-dependent pathways, including the activation of P53, the upregulation of pro-survival molecules such as BCL-2 and FLIP, and the activation of the Mitogen-activated protein kinase (MAPK) pathways, phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) pathway, the c-Jun N-terminal kinase (JNK), and nuclear factor-κB (NF-κB) pathways, among others.14
MAPK pathways are distinguished by a highly conserved three-tier kinase module and regulate essential cellular processes in response to stress and growth factors.15 In a mouse model of breast cancer bone metastasis, ononin Inhibits tumor bone metastasis and osteoclastogenesis, suggested that ononin has therapeutic potential as a BCBM treatment by targeting the MAPK pathway.16 The SAPK/JNK cascade and the MEK/ERK cascade are two well-characterized MAPK pathways. The interaction between these two pathways gives rise to a comprehensive and interconnected signaling network that plays a pivotal role in regulating cell proliferation, migration, and invasion in breast cancer.17–21 It has been reported that the simultaneous elevation of the activity of both pathways has been observed in several cases of breast cancer.22–25
Previous studies in murine models have indicated that integrin α3β1-mediated signaling pathways are vital in the initiation and/or growth of breast tumors.26 The aim of this study was to investigate the role of the complex formed by CD151 and integrin α3β1 in the progression of TNBC. To this end, we verified in vitro the pro-tumorigenic role of CD151 in TNBC cell migration and invasion, as well as its impact in TNBC progression after forming a complex with integrin α3β1. Our results suggest that the CD151-integrin α3β1 complex plays a significant role in breast cancer tumor progression by contributing to epithelial-mesenchymal transition (EMT) through MAPK pathway.
Materials and Methods
Clinical Specimens
We obtained 64 pairs of paraffin-embedded tissues from invasive TNBC, consisting of 48 cases of ductal carcinoma and 16 cases of lobular carcinoma, as well as 4 cases of normal breast tissue from the fourth affiliated hospital of China Medical University between January 2021 and December 2022. The pathological tumor stage was assessed according to the American Joint Committee on Cancer. The histological grade was determined according to the Bloom – Richardson classification scheme. All patients were initially treated in our hospital and had not received surgery, radiotherapy, chemotherapy, immunotherapy, molecular targeted therapy, etc. in the past, all cases had detailed information and well documented follow-up records, and carcinoma was approved by pathological diagnostic tools by two expert pathologists who resided in the hospital. This project was confirmed by the Ethics Committee of the fourth affiliated hospital of China Medical University.
Cell Culture
All cell lines were purchased from Procell Life Science & Technology Co., Ltd (Wuhan, China). The human breast cancer cell lines HS578T, BT549, MDA-MB-453 and MDA-MB-468 were seeded and grown in dulbecco’s modified eagle medium (DMEM) (Procell, Wuhan, China), supplemented with 10% fetal bovine serum (Procell, Wuhan, China) and 1% antibiotics (Procell, Wuhan, China) at 37 °C in a humidified atmosphere with 5% CO2.
RNA Extraction and Quantitative Real-Time qPCR Analysis
Total RNA was extracted from cells using the SteadyPure Quick RNA Extraction Kit (Accurate Biotechnology, Hunan, China) in accordance with the manufacturer’s instructions.27 cDNA was synthesized using the Evo M-MLV RT Premix for qPCR (Accurate Biotechnology, Hunan, China).28 The primer sequences utilized for mRNA detection are as follows: CD151: 5′-CGAGACCATGCCTCCAACAT-3′(forward), 5′-CATGCCAAAGACCIGCACAC-3′(reverse); GAPDH: 5′-GCACCGTCAAGGCTGAGAAC-3′(forward), 5′-TGGTGAAGACGCCAGTGGA-3′(reverse). Real-Time PCR analysis was conducted using the SYBR Green Premix Pro Taq HS qPCR Kit (Accurate Biotechnology, Hunan, China)29 and the QuantStudioTM 3 Real-Time PCR system (Thermo Fisher Scientific, China). CT values of CD151 mRNA were equalized to the internal control, GAPDH. The relative expression was calculated by using the ΔΔCt method.
Transfection
HS578T cells were inoculated in T25 cell culture flasks. When the cells reached 70–80% confluence, we performed transfection using lipofectamine 3000 reagent (Thermo Fisher) according to the manufacturer’s instructions. Cells were collected 24–48h after transfection for further experiments. CD151 siRNA, CD151 overexpression plasmid and corresponding controls were purchased from OriGene Technologies (Wuxi, China), and integrin α3 (ITGA3) and integrin β1 (ITGB1) siRNAs and corresponding controls were purchased from Starvio Biotechnology (Shanghai, China). The target siRNA sequences were as follows: CD151 siRNA: 5′-GCUGUUUACCUACAAUUGCUGCUTC −3′; ITGA3 siRNA: 5′-GGAUGAACAUCACAGUGAATT-3′; ITGB1 siRNA: 5′-GCAGCACAGAUGAAGUUAATT-3′.
Immunohistochemistry
Sections were incubated overnight at 4 °C with CD151 antibody (dilution ratio 1:200, Proteintech, 66567-1-Ig) followed by incubation with the corresponding biotinylated secondary antibody. Reactions were performed with Elivision TM plus Polyer HRP IHC Kit (KIT-9902, MXB biotechnologies, Fuzhou, China), and sections were counterstained with hematoxylin. CD151 expression was independently reviewed and scored by two clinical pathologists. Differing cases were re-examined until a consensus was reached. CD151 expression was assessed semi-quantitatively using the IRS score, which is derived from the intensity score multiplied by the percentage of positive cell score. 6 points and more were judged to be IRS positive. Immunostaining intensity was scored on a scale of 0–3: negative, 0; weak, 1; moderate, 2; and strong, 3. The percentage of positive cells was scored on a scale of 0–4: less than 1%, 0; 1–10%, 1; 11–50%, 2; 50–80%, 3; 81 −100%, 4.30,31
Western Blot Assay
Cells were lysed by 1xRIPA buffer with protease inhibitor cocktail (Seven Biotech, Beijing, China) and phosphatase inhibitors (MCE) on ice for 30 min, and centrifuged at 4 °C, 12,000g for 15 min. The extracted proteins’ concentration was detected using the Ready-to-Use BCA Protein Concentration Assay Kit (Seven Biotech, Beijing, China), then separated using a 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis and subsequently transferred to PVDF membranes (Millipore, Billerica, MA, USA). Antibodies using in the analysis were as follows: anti-CD151 (66567-1-Ig), anti-Integrin α3 (21992-1-AP), anti-Integrin β1 (12594-1-AP), anti-GAPDH (60004-1-Ig), anti-JNK (24164-1-AP), anti-p-JNK (80024-1-RR), anti-ERK1/2 (11257-1-AP), anti-p-ERK1/2 (28733-1-AP) and anti-mouse (SA00001-1) or anti-rabbit (SA00001-2) secondary antibodies (Proteintech, Wuhan, China); anti-N-cadherin (13116), anti-Vimentin (5741), anti-Snail (3879) and were all purchased from Cell Signaling Technology (Danvers, MA, USA).
Co-Immunoprecipitation (Co-Ip) Assay
Co-ip experiments were performed according to the instructions of the immunoprecipitation kit (IK-1004, Biolinkedin, Shanghai, China).32 Briefly, HS578T breast cancer cells were cultured in 100mm plates to 90–100% confluence. The cells were washed twice by the pre-cooled phosphate buffer saline (PBS) and collected by scraping. Then lysed with 1mL of IP buffer containing PMSF and phosphatase inhibitor cocktail for 20 min. Collected cell lysates by centrifugation at 14000×g at 4 °C for 10 min. In a centrifuge tube, 2 µg of Mouse anti-CD151 (66567-1-Ig, Proteintech, Wuhan, China) or Mouse IgG (B900620, Proteintech, Wuhan, China) and 25 µL of Protein A/G magnetic beads, previously washed with pre-cooled PBS, should be mixed gently and then rotated overnight in a refrigerator set at 4 °C. The magnetic beads were collected using a magnetic rack and washed three times with IP lysis. Subsequently, 100 µL SDS-PAGE Sample Loading Buffer (1x) was added to the centrifuge tube, and the sample was heated in a water bath at 60 °C for 15 minutes. The magnetic beads were then separated using a magnetic rack, and the supernatant was retained. Finally, 20 µL of the sample was loaded onto an SDS-PAGE gel for further Western blot analysis.
Single Cell RNA Sequencing
Breast cancer patients were recruited from the Fourth Affiliated Hospital of China Medical University, and their breast cancer tissue and adjacent tissue samples were collected for preparing single-cell suspensions. The preparation of the scRNA-seq libraries was conducted using the SeekOne DD Single Cell 5′ Library Preparation Kit (SeekGene). The purified libraries were subsequently subjected to sequencing using an Illumina NovaSeq 6000 with a read length of PE150. The raw sequencing data were then processed by the SeekOne tools (V.1.2.0) software suite. Low-quality cells were excluded if the number of expressed genes was less than 200 or greater than 6,000, or if the number of unique molecular identifiers was less than 2,200, or if the proportion of mitochondrial genes was greater than 5%.
The single-cell RNA sequencing (scRNA-seq) data consists of one luminal B breast cancer sample (C1), two luminal A breast cancer samples (C2 and C4), one adjacent tissue sample of C2 (N2), and one TNBC sample (C3). ScRNA-seq data analysis and visualization were performed by SeekSoul Online (https://seeksoul.online/index.html#/login).
Cell Proliferation Analysis (CCK8)
Cell proliferation was determined using Cell Counting Kit-8 (CCK-8) (Seven Biotech, Beijing, China). Well-grown HS578T cells were inoculated into 96-well plates at an amount of 2×103 per well and cultured under normal culture conditions for 24, 48 and 72 h, respectively, and assayed according to the instructions. The experiment was repeated three times.
Migration and Invasion Assays
Transwell inserts with 8.0 μm pore (TCS003024, Biofil, Guangzhou, China) were utilized for the performance of cell migration and invasion assays. To perform the migration assay, 500 µL of DMEM medium with 20% FBS was added to each lower chamber of a transwell insert. 3×105 stable cells with medium containing 1% FBS were seeded into the upper chamber. The cells were then incubated at 37 °C for 24 h in a humidified atmosphere with 5% CO2. Subsequently, the cells that had migrated onto the lower surface of the insert were fixed with 4% paraformaldehyde for 20 minutes, air-dried for 10 minutes, stained with 0.1% crystal violet for 30 minutes, and washed with PBS third times, air-dried for 10 minutes. Subsequently, photographed and counted the cells. For the invasion assay, the inserts were pre-coated with Matrix-Gel™ Basement Membrane Matrix (Beyotime Biotechnology, Shanghai, China) diluted with serum-free medium at a ratio of 1:8, then incubated at 37 °C for 3 h. The remaining procedures were performed in accordance with the migration assay. Each experiment was repeated three times.
Wound Healing Assay
Inoculate HS578T stable cells at a certain density into 12-well tissue culture plates, they should then reach about 70–80% confluence. Use a new sterile 10µL pipette tip to gently and slowly scrape the monolayer through the center of the wells, aiming for the resulting gap distance to be equal to the outer diameter of the end of the tip. Subsequently, the detached cells were removed via two gentle washes in PBS. The well was refilled with fresh DMEM medium with 1% FBS, and the cells were cultured for an additional 24 hours. The cells were observed and imaged under a microscope (EVOS FL Auto 2 Cell Imaging System, Thermo Fisher Scientific) at the same magnification and settings. The width of the gap was subsequently quantified using ImageJ.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 10.1.2 (GraphPad, San Diego, CA, USA). Student’s t-test and ANOVA were used depending on experimental conditions. All data presented as means ± SD, P < 0.05 was considered statistically significant.
Results
Patient Characteristics and CD151 Expression
To evaluate CD151 expression patterns in TNBC clinical samples and cell lines and assess its clinical relevance. In 64 cases, the median age at diagnosis was 53 years, with a range from 30 to 86 years. The characteristics of patients are summarized in Table 1. Notably, overexpression of CD151 was observed more frequently in TNBC cases, accounting for 82.8% of the instances. There were no significant differences found in lymph node involvement (P=0.601) or the different stages of cancer (P=0.526), nor between ductal and lobular TNBC (P=0.169). Additionally, age did not appear to influence CD151 expression (P=0.248) (Table 1 and Figure 1A). In normal breast tissue, CD151 was expressed in only a few cells. In the adjacent tissue, CD151 was expressed in the basal-myoepithelial cell layer of some cells. However, in invasive cancers, the expression of CD151 was primarily localized to the membrane, with some cells also showing expression in the cytoplasm (Figure 1B).
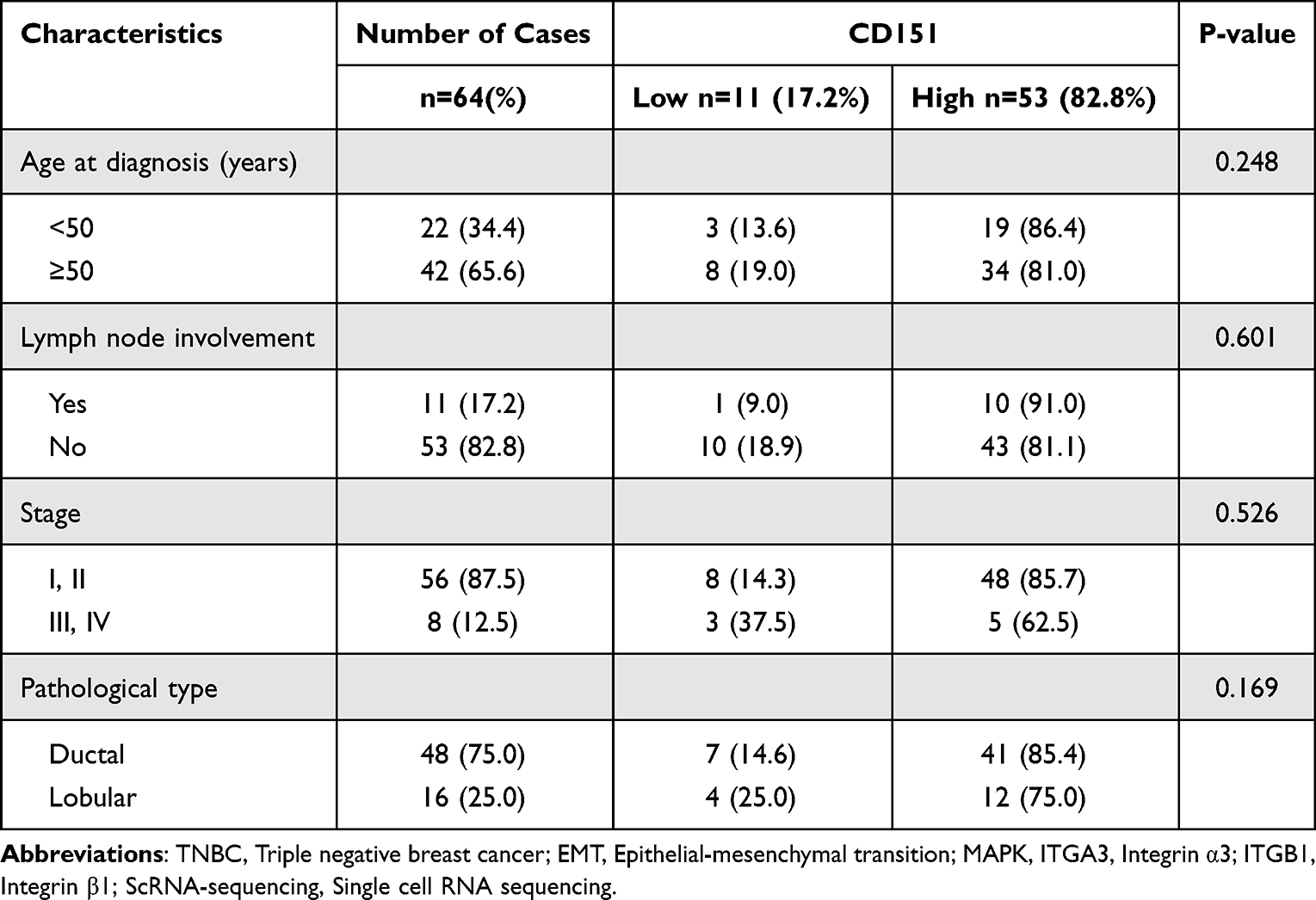 |
Table 1 Characteristics of Patients with Invasive Breast Cancer Based on CD151 Expression |
 |
Figure 1 CD151 expression is up-regulated in TNBC tissues and cell lines. (A), Characteristics of patients with invasive breast cancer based on CD151 expression. Data are presented in percent stacted column chart. There were no significant differences found in lymph node involvement or the different stages of cancer, nor between ductal and lobular TNBC. Additionally, age did not appear to influence CD151 expression. Comparisons were made within groups. (B), TNBC samples were immunostained with anti-CD151 antibody. The figures illustrate representative samples of TNBC, adjacent tissue, and normal breast tissue. In normal breast tissue, CD151 was expressed in only a few cells. In the adjacent tissue, CD151 was expressed in the basal-myoepithelial cell layer of some cells. In invasive cancers, CD151 expression was primarily localized to the membrane, with some cells also showing expression in the cytoplasm. (C), CD151 mRNA and protein expression level in different breast cancer cell lines, data from the Proteinatlas database (https://www.proteinatlas.org/ENSG00000177697-CD151/cell+line). (D), CD151 mRNA expression in human breast cancer cell lines were analyzed by RT-qPCR, GAPDH as an internal reference (n=3). (E) CD151 protein expression in human breast cancer cell lines were analyzed by Western blot, GAPDH as an internal reference (n=3). Abbreviation: ns, no significance. |
Immunohistochemical analysis revealed that the expression of CD151 was significantly higher in TNBC tissues and adjacent tissues compared to normal tissues. However, data from the Proteinatlas database (https://www.proteinatlas.org/ENSG00000177697-CD151/cell+line) indicated varying expression levels in different breast cancer cell lines (Figure 1C). Our findings further confirmed these variations in the HER2-positive cell line MDA-MB-453, as well as in TNBC cell lines MDA-MB-468, BT549, and HS578T (Figure 1D and E). Notably, mRNA and protein levels of CD151 were highest in the HS578T cell line. These findings established CD151 as a biomarker overexpressed in aggressive TNBC, correlating with metastatic potential but independent of traditional prognostic factors like lymph node status.
CD151 Promotes the Migration and Invasive Ability of TNBC Cells but Inhibits Proliferation
Knockdown of CD151 in TNBC Cells
To elucidate the functions of CD151 in TNBC, we knockdown the CD151 in the TNBC cell line HS578T. Following transfection with siRNA, a significant reduction in CD151 expression was observed at the mRNA levels (Figure 2A). Subsequently, the relative proliferation of HS578T cells was determined by CCK-8 assay, which demonstrated that knockdown of CD151 enhanced HS578T cell proliferation (Figure 2B). The role of CD151 in regulating cell migration and invasion was further evaluated. Results of the transwell assay showed that both the migration and invasion abilities of HS578T cells were inhibited after the knockdown of CD151 (Figure 2C). The results of the wound healing assay confirmed that the migration of HS578T cells was inhibited following the knockdown of CD151 (Figure 2D). To elucidate further the mechanisms of CD151 promoting the migration and invasion ability of HS578T cells, we examined the expression of EMT biomarkers by Western blot. The results indicated that the expression of EMT biomarkers decreased after the knockdown of CD151 (Figure 2E–I).
 |
Figure 2 Knockdown of CD151 inhibits breast cancer cells migration and invasion but promotes proliferation in vitro via Epithelial–mesenchymal transition. (A), CD151 mRNA levels in HS578T cells which transfected with negative control (si-NC) and CD151 siRNA (si-CD151), GAPDH as an internal reference (n=3). (B), CCK-8 assay of cell relative proliferation in HS578T cells (n=5). (C), Representative images of transwell assay results of cell migration and invasion of HS578T cells (si-CD151 vs si-NC) (n=3). (D), Wound healing assay was performed to observe the role of CD151 in HS578T cells migration. (E), Western blot assay showed changes in EMT biomarkers’ expression in HS578T cells after CD151 knockdown. (F-I), Results of statistical analysis of protein expression of CD151, N-cadherin, Vimentin, and Snail. GAPDH was used as an internal reference for homogenization (si-CD151 vs si-NC) (n=3). Data are represented as the mean ± SD. * P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. |
CD151 Over-Expression in TNBC Cells
In addition to knocking down CD151, we evaluated the role of CD151 overexpression in regulating the proliferation, migration, and invasion of TNBC cells. CD151 expression has been significantly up-regulated, as confirmed by RT-qPCR and Western blot assay (Figure 3A). We found that CD151 overexpression inhibited cell proliferation by CCK-8 assay (Figure 3B). In addition, the results of the transwell assay and wound healing assay showed that CD151 overexpression enhanced cell migration and invasion (Figure 3C and D). Similarly, the expression of EMT biomarkers was up-regulated after overexpression of CD151 (Figure 3E–I). These results indicated that CD151 exhibits a dual role—enhancing metastatic capacity while paradoxically suppressing proliferation.
 |
Figure 3 Overexpression of CD151 promotes breast cancer cells migration and invasion but inhibits proliferation in vitro via Epithelial–mesenchymal transition. (A), CD151 mRNA and protein levels in CD151-overexpressed HS578T cells, GAPDH as an internal reference (n=3). (B), CCK-8 assay of cell relative proliferation in HS578T cells (n=5). (C), Representative images of transwell assay results of cell migration and invasion of HS578T cells (CD151-overexpressed vs Vector) (n=3). (D), Wound healing assay was performed to observe the role of CD151 in HS578T cells migration. (E), Western blot assay showed changes in EMT biomarkers expression in HS578T cells after CD151 overexpressed. (F-I), Results of statistical analysis of protein expression of CD151, N-cadherin, Vimentin, and Snail. GAPDH was used as an internal reference for homogenization (CD151-overexpressed vs Vector) (n=3). Data are represented as the mean ± SD. * P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001. |
CD151 Promotes EMT by Activating MAPK Signaling Pathways
To elucidate the mechanism of CD151-mediated EMT, we performed differential gene enrichment analyses by scRNA-seq for the TNBC sample (C3, n=1), luminal A samples (C2, n=1 and C4, n=1), luminal B sample (C1, n=1) and luminal A breast cancer adjacent tissue sample (N2, n=1). After integrating the transcriptional data from all acquired cells, we first applied low‐resolution Uniform Manifold Approximation and Projection plot (UMAP), clustering and generated 2D graphs with 6 clusters in 5 samples (Figure 4A). To identify the main cell types of this atlas, we annotated each cluster with their marker gene expression. All cells were divided into five cell types, including epithelial cells (COX6C, KRT19, TFF3, SCGB2A2, MUCL1), endothelial cells (PTPRC, FYN, CD52, IL7R, IGKC), fibroblast cells (DDCN, LUM, COL1A1, APOD), myeloid cells (CCL3, CCL3L1, CXCL8, CCL4L2, SPP1) and pericytes cells (MYL9, TAGLN, ACTA2, MYH11, RGS5) (Figure 4B). Considering that breast cancer cells originate from epithelial cells, we subsequently analyzed the differential gene enrichment in epithelial cells from C3 versus C1 and C2 breast cancer sample on Seeksoul Online. The results showed that the differential genes between TNBC and other types of breast cancer were mainly enriched in the MAPK signaling pathway, integrin binding, EMT, and cell migration (Figure 4C and D).
 |
Figure 4 CD151 is associated with MAPK and integrin signaling pathway and abnormal expression of CD151 disrupts MAPK signaling. (A), UMAP plot of 54836 cells colored by major cell type in this study. (B), Bubble plots of marker genes expressed in major cell types. Dot colors reflect expression levels and dot sizes represent the percentage of cells expressing marker genes in different cell types. (C), Pathway analysis was used to identify the significant pathway of the differential genes according to the KEGG, HALLMARK and GO database between C3 (TNBC) and C1 (Luminal (B) samples. (D), Pathway analysis was used to identify the significant pathway of the differential genes according to the KEGG, HALLMARK and GO database between C3 (TNBC) and C2 (Luminal (A) samples. We used the Wilcoxon test to select the significant pathways, and the threshold of significance was defined by p.adjust. (E), Western blot of CD151, JNK, p-JNK, ERK, p-ERK expression in CD151 knockdown or overexpressed cells when compared to control cells. GAPDH was used as an internal control. (F-J), Results of statistical analysis of protein expression of JNK, p-JNK, ERK, p-ERK. GAPDH was used as an internal reference for homogenization (si-CD151 vs si-NC) (CD151-overexpressed vs Vector) (n=3). Data are represented as the mean ± SD. * P < 0.05; **P < 0.01; ***P < 0.001. Abbreviation: ns, no significance. |
Therefore, we verified the changes in phosphorylation levels of key molecules on the MAPK signaling pathway by Western blot. Western blot analysis indicated that the levels of JNK and ERK phosphorylation were decreased when CD151 was knocked down and increased when CD151 was overexpressed in TNBC cells. Moreover, protein expressions of JNK and ERK remained unaltered (Figure 4E–J). Therefore, these findings demonstrated that CD151 could promote EMT by activating MAPK signaling pathways, especially JNK and ERK.
CD151 Activates the MAPK Signaling Pathway by Forming a Complex with Integrin α3β1
To further investigate the complex of CD151 with integrins, Co-ip experiments were performed. The results of Co-ip demonstrated that CD151 could bind to integrin α3 and integrin β1 in HS578T cells and form CD151-integrin α3β1 complex (Figure 5A). Besides, the results of Western blot analysis demonstrated that the expression of integrin α3 and β1 was not affected by the expression levels of CD151 (Figure 5B and C).
 |
Figure 5 CD151 can form a complex with integrin α3β1. (A), Co-immunoprecipitation of CD151 and integrins is shown. (B), Western blot of effect of CD151 expression levels on IGTA3 and IGTB1 expression levels, respectively. (C), Results of statistical analysis of protein expression of IGTA3 and IGTB1. GAPDH was used as an internal reference for homogenization (si-CD151 vs si-NC) (CD151-overexpressed vs Vector) (n=3). Data are represented as the mean ± SD. Significant differences compared with the control. Abbreviation: ns, no significance. |
Based on the findings, we used specific siRNAs to disrupt the expression of integrin α3 and β1 in CD151-overexpression cells. The results of the transwell and wound healing assay showed that the migration and invasion capacity of HS578T cells was also suppressed following the knockdown of integrin α3 and β1. Nevertheless, CD151 overexpression was not observed to reverse the diminished cell migration and invasion capacity (Figure 6A and B). The results of Western blot revealed that the phosphorylation levels of JNK and ERK were elevated in CD151 over-expression TNBC cells. However, when integrin was interfered with specific siRNAs, the phosphorylation levels of JNK and ERK decreased (Figure 6C–I). Therefore, these results indicated that CD151-integrin α3β1 complex formation is essential for MAPK activation, revealing a non-canonical integrin signaling mechanism independent of α3β1 expression modulation.
 |
Figure 6 Impact of integrin α3 and β1 expression in CD151 overexpression HS578T cells. (A), CD151 overexpression HS578T cells were seeded into 6-well plates and then treated with specific siRNA for transwell assay, respectively. (B), CD151 overexpression HS578T cells were seeded into 6-well plates and then treated with specific siRNA for wound healing assay, respectively. (C), Western blot assay showed the effect of integrin α3 or β1 knockdown on the MAPK signaling pathway in CD151 overexpression cells. (D-I), Results of statistical analysis of protein expression of IGTA3, IGTB1, JNK, p-JNK, ERK, p-ERK. GAPDH was used as an internal reference for homogenization (Vector vs Vector+si-IGA3/si-IGB1) (CD151-overexpressed vs CD151-overexpresse+ si-IGA3/si-IGB1) (n=3). Data are represented as the mean ± SD. * P < 0.05; **P < 0.01; ***P < 0.001. Abbreviation: ns, no significance. |
CD151-Integrin α3β1 Complex Promotes EMT via MAPK Signaling Pathway
Combined with the results we obtained; we present a schematic representation of the role of the CD151-integrin α3β1 complex in TNBC (Figure 7). In conclusion, the present study has demonstrated the pro-migratory and invasive ability of CD151 in TNBC cells. The pro-tumorigenic effects of CD151 are exerted through its binding to integrin α3β1, rather than through the promotion of integrin expression. The CD151-integrin α3β1 complex can activate a variety of signaling pathways, which in our study are the SAPK/JNK cascade and the MEK/ERK cascade. The activated MAPK signaling pathway subsequently promoted EMT in TNBC cells.
 |
Figure 7 Schematic representation of the functional role of the CD151-integrin α3β1 complex in TNBC. In TNBC, CD151 forms a complex with integrin α3β1. Then, the complex activates the MAPK pathway, which promotes EMT and enhances the migration and invasion of TNBC cells. |
In summary, these findings suggest that CD151 contributes to EMT by forming a complex with integrin α3β1, thereby activating the MAPK pathway.
Discussion
As a member of the tetraspanin family of proteins, CD151 expression is typically higher in highly invasive cancer cells and cancer-derived exosomes.33–35 CD151 plays a regulatory role in interactions between stromal and tumor cells. This is achieved, in part, through binding and stimulation of pro-MMPs on cell membranes, which in turn promotes the degradation of ECM components, including collagen and laminin. This process facilitates tumor cell invasion and metastasis.8,36–42 Our results are consistent with past studies, as shown in Figure 1, CD151 expression was higher in TNBC tissues and cell lines. The results of the transwell and wound healing assays indicate that CD151 promotes migration and invasion in TNBC cells by promoting EMT. Interestingly, clinical samples revealed a high expression of CD151 in TNBC; however, this expression did not correlate with lymph node involvement or cancer stage.
CD151 is pivotal in regulating cell adhesion through binding to laminin integrins. The CD151-integrin complex is essential for signaling and is a prerequisite for CD151 to exert several biological effects.43–45 In breast cancer cells, CD151 activates RhoA via integrin α3β1, which controls ErbB2 dimerization and thus promotes breast cancer motility and metastasis.46 In hepatocellular carcinoma cells, CD151 interacts with integrin α3β1 and integrin α6β1 to form a complex, which further affects the biological function of hepatocellular carcinoma.47 Additionally, in non-small cell lung cancer, CD151 has been demonstrated to facilitate the development and progression of NSCLC by forming a complex with integrin α3β1, which in turn regulates EGFR/ErbB2 and its downstream signaling pathways.48 These studies prompted us to investigate a potential association between integrin α3β1 and CD151 in TNBC. Our findings suggest that CD151 formed a complex with integrin α3β1 in TNBC cells; however, unlike in non-small cell lung cancer, the expression level of CD151 does not affect the expression level of integrin α3 or integrin β1.
The two principal functions of integrins are mechanical attachment to the extracellular matrix (ECM) and the activation of signal transduction pathways that regulate a range of cellular processes essential for the initiation, progression, and metastasis of solid tumors.49 Integrins are capable of bidirectional signaling, and various cytoplasmic interactions control the activation of integrins, which in turn alters their affinity for extracellular ligands that transmit “inside-out” signals.50,51 Activated integrins increase their affinity for ECM ligands.52 Consequently, integrin-ECM interactions result in the recruitment of signaling molecules to the cytoplasmic structural domains of integrins, thereby facilitating the assembly of macromolecular complexes, which are also known as focal adhesions.53 It can be reasonably deduced that integrin-controlled cell migration is primarily mediated by a signaling pathway involving members of the adhesion patch kinase (FAK)-SRC family of kinases, which is highly dependent on integrin-specific mechanisms. In summary, integrin-ligand adhesion results in an increase in FAK tyrosine (Tyr) 397 phosphorylation, which generates binding sites for SRC kinase structural domains, namely SRC homology 2 (SH2) and SH3.54,55 The activated FAK/SRC complex facilitates various key signaling cascades that regulate cell motility.12 Integrins also play a role in cancer cell proliferation and survival, which, as previously described, is promoted through several complex and context-dependent pathways. In addition to P53 activation, integrin ligation has been demonstrated to trigger the up-regulation of BCL-2 and FLIP pro-survival molecules, as well as the mitogen-activated protein kinase (MAPK)/extracellularly regulated kinase (ERK) pathway,56–58 the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) pathway,59–61 the c-Jun N-terminal kinase (JNK),62–64 and the stress-activated MAP kinase (SAPK) or activation of nuclear factor-κB (NF-κB) signaling.65–67
To elucidate the mechanism by which the CD151-integrin α3β1 complex promotes EMT, we conducted a comprehensive analysis of differential gene enrichment utilizing single-cell RNA sequencing in a series of breast cancer samples, including TNBC and luminal A and B subtypes. Our findings revealed that the differentially expressed genes were mainly enriched in the MAPK signaling pathway, EMT and integrin binding (Figure 4A). This is consistent with our prediction and past findings that activation of the MAPK pathway is often involved in the progression of TNBC.68–70 As mentioned earlier, the SAPK/JNK cascade and the MEK/ERK cascade are two well-characterized MAPK pathways; the interactions between these two pathways form a comprehensive, interconnected signaling network that plays a key role in regulating the proliferation, migration, and invasion of breast cancer cells.17–20,25 Combining previous studies and our results, we conclude that binding of CD151 to integrins is required in TNBC progression. In TNBC, CD151 exerts a pro-tumor progression function mainly by activating various signaling pathways after forming a complex with integrin α3β1. In our study, the MAPK signaling pathway was activated, which in turn contributed to the promotion of EMT. Considering the practical limitations associated with CD151 gene deletion, the most common approach to targeting CD151 involves using a monoclonal antibody. Previous studies have shown that monoclonal antibodies directed against CD151 and integrin α6β1 complex binding inhibit HCC cell growth and invasion as well as tumor-induced angiogenesis through cytoplasmic and membrane-bound CD151 antigen responses.71 Our findings highlight the need for further investigation into the potential role of CD151 in the clinical management of TNBC.
While our current findings have shed light on the pro-tumorigenic role of CD151 in TNBC progression, several mechanistic aspects remain to be fully elucidated. Specifically, the precise cellular mechanisms by which CD151 promote EMT require deeper investigation at the molecular level, the pathophysiological relevance of these findings needs validation through appropriate in vivo models. To address these gaps, our future work will focus on employing CRISPR-based gene editing and proteomic approaches to map CD151’s interactome and downstream signaling networks in TNBC cells and establishing patient-derived tumor xenograft (PDX) and conditional knockout mouse models.
Conclusion
Our study establishes a novel link between the complex formed by CD151 with integrin α3β1 and the MAPK signaling pathway. Given the role of CD151 in TNBC progression, it suggests that CD151 could be a potential target for clinical treatment of this condition. A deeper understanding of these interactions and their effects on tumor progression may help to identify new prognostic markers for TNBC and new strategies to prevent cancer metastasis. Considering the practical limitations associated with CD151 gene deletion, the most common approach to targeting CD151 involves using a monoclonal antibody. This treatment effectively disrupts the binding between CD151 and integrin α6β1. In conclusion, our findings highlight the need for further investigation into the potential role of CD151 in the clinical management of TNBC.
Data Sharing Statement
The data presented in this study are available on request from the corresponding author.
Ethics Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Ethics Committee at the Fourth Affiliated Hospital of China Medical University (EC-2021-KS-041, 2021/04/29). Due to the non-interventional nature of the study, the fact that the waiver of informed consent would not adversely affect the health and rights of the subjects, and that the subjects’ privacy and personally identifiable information would be protected, the waiver of informed consent was granted.
Acknowledgments
The authors gratefully acknowledge all the patients and their families. We also acknowledge the Central Laboratory at the First Affiliated Hospital of China Medical University.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by National Natural Science Foundation of China (Grant No. 82072936).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Thakur C, Qiu Y, Pawar A, Chen F. Epigenetic regulation of breast cancer metastasis. Cancer Metastasis Rev. 2024;43(2):597–619. doi:10.1007/s10555-023-10146-7
2. Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA. 2024;74(1):12–49. doi:10.3322/caac.21820
3. James N, Owusu E, Rivera G, Bandyopadhyay D. Small molecule therapeutics in the pipeline targeting for triple-negative breast cancer: origin, challenges, opportunities, and mechanisms of action. Int J Mol Sci. 2024;25(11):6285. doi:10.3390/ijms25116285
4. Zöller M. Tetraspanins: push and pull in suppressing and promoting metastasis. Nat Rev Cancer. 2009;9(1):40–55. doi:10.1038/nrc2543
5. Malla R, Marni R, Chakraborty A. Exploring the role of CD151 in the tumor immune microenvironment: therapeutic and clinical perspectives. Biochim Biophys Acta Rev Cancer. 2023;1878(3):188898. doi:10.1016/j.bbcan.2023.188898
6. Zijlstra A, Lewis J, Degryse B, Stuhlmann H, Quigley JP. The inhibition of tumor cell intravasation and subsequent metastasis via regulation of in vivo tumor cell motility by the tetraspanin CD151. Cancer Cell. 2008;13(3):221–234. doi:10.1016/j.ccr.2008.01.031
7. Sadej R, Grudowska A, Turczyk L, Kordek R, Romanska HM. CD151 in cancer progression and metastasis: a complex scenario. Lab Investigat. 2014;94(1):41–51. doi:10.1038/labinvest.2013.136
8. Sugiura T, Berditchevski F. Function of alpha3beta1-tetraspanin protein complexes in tumor cell invasion. Evidence for the role of the complexes in production of matrix metalloproteinase 2 (MMP-2). J Cell Biol. 1999;146(6):1375–1389. doi:10.1083/jcb.146.6.1375
9. Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110(6):673–687. doi:10.1016/S0092-8674(02)00971-6
10. Huveneers S, Danen EH. Adhesion signaling - crosstalk between integrins, Src and Rho. J Cell Sci. 2009;122(Pt 8):1059–1069. doi:10.1242/jcs.039446
11. Brunton VG, Frame MC. Src and focal adhesion kinase as therapeutic targets in cancer. Curr Opin Pharmacol. 2008;8(4):427–432. doi:10.1016/j.coph.2008.06.012
12. Tuguzbaeva G, Yue E, Chen X, et al. PEP06 polypeptide 30 is a novel cluster-dissociating agent inhibiting α v integrin/FAK/Src signaling in oral squamous cell carcinoma cells. Acta pharmaceutica Sinica B. 2019;9(6):1163–1173. doi:10.1016/j.apsb.2019.10.005
13. Fournier AK, Campbell LE, Castagnino P, et al. Rac-dependent cyclin D1 gene expression regulated by cadherin- and integrin-mediated adhesion. J Cell Sci. 2008;121(Pt 2):226–233. doi:10.1242/jcs.017012
14. Li M, Wang Y, Li M, Wu X, Setrerrahmane S, Xu H. Integrins as attractive targets for cancer therapeutics. Acta pharmaceutica Sinica B. 2021;11(9):2726–2737. doi:10.1016/j.apsb.2021.01.004
15. Arthur JS, Ley SC. Mitogen-activated protein kinases in innate immunity. Nat Rev Immunol. 2013;13(9):679–692. doi:10.1038/nri3495
16. Ganesan K, Xu C, Wu S, et al. Ononin inhibits tumor bone metastasis and osteoclastogenesis by targeting mitogen-activated protein kinase pathway in breast cancer. Research. 2024;7:0553. doi:10.34133/research.0553
17. Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002;12(1):9–18. doi:10.1038/sj.cr.7290105
18. Shen YH, Godlewski J, Zhu J, et al. Cross-talk between JNK/SAPK and ERK/MAPK pathways: sustained activation of JNK blocks ERK activation by mitogenic factors. J Biol Chem. 2003;278(29):26715–26721. doi:10.1074/jbc.M303264200
19. Mingo-Sion AM, Marietta PM, Koller E, Wolf DM, Van Den Berg CL. Inhibition of JNK reduces G2/M transit independent of p53, leading to endoreduplication, decreased proliferation, and apoptosis in breast cancer cells. Oncogene. 2004;23(2):596–604. doi:10.1038/sj.onc.1207147
20. Fujii S, Tokita K, Wada N, et al. MEK-ERK pathway regulates EZH2 overexpression in association with aggressive breast cancer subtypes. Oncogene. 2011;30(39):4118–4128. doi:10.1038/onc.2011.118
21. Wu L, Huang S, Tian W, et al. PIWI-interacting RNA-YBX1 inhibits proliferation and metastasis by the MAPK signaling pathway via YBX1 in triple-negative breast cancer. Cell Death Discovery. 2024;10(1):7. doi:10.1038/s41420-023-01771-w
22. Lopez-Bergami P, Huang C, Goydos JS, et al. Rewired ERK-JNK signaling pathways in melanoma. Cancer Cell. 2007;11(5):447–460. doi:10.1016/j.ccr.2007.03.009
23. Ramírez A, Boulaiz H, Morata-Tarifa C, et al. HER2-signaling pathway, JNK and ERKs kinases, and cancer stem-like cells are targets of Bozepinib small compound. Oncotarget. 2014;5(11):3590–3606. doi:10.18632/oncotarget.1962
24. Grijelmo C, Rodrigue C, Svrcek M, et al. Proinvasive activity of BMP-7 through SMAD4/src-independent and ERK/Rac/JNK-dependent signaling pathways in colon cancer cells. Cell Signal. 2007;19(8):1722–1732. doi:10.1016/j.cellsig.2007.03.008
25. Jiang T, Pan CQ, Low BC. BPGAP1 spatially integrates JNK/ERK signaling crosstalk in oncogenesis. Oncogene. 2017;36(22):3178–3192. doi:10.1038/onc.2016.466
26. Cagnet S, Faraldo MM, Kreft M, Sonnenberg A, Raymond K, Glukhova MA. Signaling events mediated by α3β1 integrin are essential for mammary tumorigenesis. Oncogene. 2014;33(34):4286–4295. doi:10.1038/onc.2013.391
27. SteadyPure quick RNA extraction kit [package insert]. Changsha, China: ACCURATE BIOTECHNOLOGY(HUNAN) CO.LTD; 2024.
28. Evo M-MLV RT kit with gDNA clean for qPCR [package insert]. Changsha, China: ACCURATE BIOTECHNOLOGY(HUNAN) CO.LTD; 2024.
29. SYBR® Green Premix Pro Taq HS qPCR Kit II (Rox Plus) [package insert]. Changsha, China: ACCURATE BIOTECHNOLOGY(HUNAN) CO.LTD; 2024.
30. Wang J, Qiu Q, Lian N, et al. An immunosuppressive scoring system to predict recurrence and assist in decision regarding postoperative adjuvant treatment in gastric cancer. Am J Cancer Res. 2022;12(5):2050–2067.
31. Xiong H, Liao M, Zhang H, et al. DARS expression in BCR/ABL1-negative myeloproliferative neoplasms and its association with the immune microenvironment. Sci Rep. 2024;14(1):16711. doi:10.1038/s41598-024-67067-w
32. Protein A+G magnetic beads [package insert]. Shanghai, China: Biolinkedin; 2024.
33. Li D, Lai W, Wang Q, et al. CD151 enrichment in exosomes of luminal androgen receptor breast cancer cell line contributes to cell invasion. Biochimie. 2021;189:65–75. doi:10.1016/j.biochi.2021.06.007
34. Ke AW, Shi GM, Zhou J, et al. CD151 amplifies signaling by integrin α6β1 to PI3K and induces the epithelial-mesenchymal transition in HCC cells. Gastroenterology. 2011;140(5):1629–1641.e1615. doi:10.1053/j.gastro.2011.02.008
35. Kwon MJ, Park S, Choi JY, et al. Clinical significance of CD151 overexpression in subtypes of invasive breast cancer. Br J Cancer. 2012;106(5):923–930. doi:10.1038/bjc.2012.11
36. Fullár A, Dudás J, Oláh L, et al. Remodeling of extracellular matrix by normal and tumor-associated fibroblasts promotes cervical cancer progression. BMC Cancer. 2015;15:256. doi:10.1186/s12885-015-1272-3
37. Berditchevski F. Complexes of tetraspanins with integrins: more than meets the eye. J Cell Sci. 2001;114(Pt 23):4143–4151. doi:10.1242/jcs.114.23.4143
38. Lazo PA. Functional implications of tetraspanin proteins in cancer biology. Cancer Science. 2007;98(11):1666–1677. doi:10.1111/j.1349-7006.2007.00584.x
39. Testa JE, Brooks PC, Lin JM, Quigley JP. Eukaryotic expression cloning with an antimetastatic monoclonal antibody identifies a tetraspanin (PETA-3/CD151) as an effector of human tumor cell migration and metastasis. Cancer Res. 1999;59(15):3812–3820.
40. Kohno M, Hasegawa H, Miyake M, Yamamoto T, Fujita S. CD151 enhances cell motility and metastasis of cancer cells in the presence of focal adhesion kinase. Int J Cancer. 2002;97(3):336–343. doi:10.1002/ijc.1605
41. Chien CW, Lin SC, Lai YY, et al. Regulation of CD151 by hypoxia controls cell adhesion and metastasis in colorectal cancer. ClinClin Cancer Res. 2008;14(24):8043–8051. doi:10.1158/1078-0432.CCR-08-1651
42. Yoo SH, Lee K, Chae JY, Moon KC. CD151 expression can predict cancer progression in clear cell renal cell carcinoma. Histopathology. 2011;58(2):191–197. doi:10.1111/j.1365-2559.2011.03752.x
43. Te Molder L, Juksar J, Harkes R, Wang W, Kreft M, Sonnenberg A. Tetraspanin CD151 and integrin α3β1 contribute to the stabilization of integrin α6β4-containing cell-matrix adhesions. J Cell Sci. 2019;132(19). doi:10.1242/jcs.235366
44. Zhang XA, Bontrager AL, Hemler ME. Transmembrane-4 superfamily proteins associate with activated protein kinase C (PKC) and link PKC to specific beta(1) integrins. J Biol Chem. 2001;276(27):25005–25013. doi:10.1074/jbc.M102156200
45. Zuo H, Liu Z, Liu X, et al. CD151 gene delivery after myocardial infarction promotes functional neovascularization and activates FAK signaling. Mol Med. 2009;15(9–10):307–315. doi:10.2119/molmed.2009.00025
46. Novitskaya V, Romanska H, Kordek R, et al. Integrin α3β1-CD151 complex regulates dimerization of ErbB2 via RhoA. Oncogene. 2014;33(21):2779–2789. doi:10.1038/onc.2013.231
47. Fei Y, Wang J, Liu W, et al. CD151 promotes cancer cell metastasis via integrins α3β1 and α6β1 in vitro. Mol Med Rep. 2012;6(6):1226–1230. doi:10.3892/mmr.2012.1095
48. Zhu J, Cai T, Zhou J, et al. CD151 drives cancer progression depending on integrin α3β1 through EGFR signaling in non-small cell lung cancer. J Exp Clin Cancer Res. 2021;40(1):192. doi:10.1186/s13046-021-01998-4
49. Humphries JD, Byron A, Humphries MJ. Integrin ligands at a glance. J Cell Sci. 2006;119(Pt 19):3901–3903. doi:10.1242/jcs.03098
50. Cooper J, Giancotti FG. Integrin signaling in cancer: mechanotransduction, stemness, epithelial plasticity, and therapeutic resistance. Cancer Cell. 2019;35(3):347–367. doi:10.1016/j.ccell.2019.01.007
51. Schwartz MA, Ginsberg MH. Networks and crosstalk: integrin signalling spreads. Nat Cell Biol. 2002;4(4):E65–68. doi:10.1038/ncb0402-e65
52. Legate KR, Wickström SA, Fässler R. Genetic and cell biological analysis of integrin outside-in signaling. Genes Dev. 2009;23(4):397–418. doi:10.1101/gad.1758709
53. Hamidi H, Pietilä M, Ivaska J. The complexity of integrins in cancer and new scopes for therapeutic targeting. Br J Cancer. 2016;115(9):1017–1023. doi:10.1038/bjc.2016.312
54. Caswell PT, Chan M, Lindsay AJ, McCaffrey MW, Boettiger D, Norman JC. Rab-coupling protein coordinates recycling of alpha5beta1 integrin and EGFR1 to promote cell migration in 3D microenvironments. J Cell Biol. 2008;183(1):143–155. doi:10.1083/jcb.200804140
55. Katoh K. Signal transduction mechanisms of focal adhesions: Src and FAK-mediated cell response. Front Biosci. 2024;29(11):392. doi:10.31083/j.fbl2911392
56. Chen Y, Gong Y, Shi M, et al. miR-3606-3p alleviates skin fibrosis by integratively suppressing the integrin/FAK, p-AKT/p-ERK, and TGF-β signaling cascades. J Adv Res. 2024. doi:10.1016/j.jare.2024.11.027
57. Sugiyanto RN, Metzger C, Inal A, et al. Proteomic profiling reveals CEACAM6 function in driving gallbladder cancer aggressiveness through integrin receptor, PRKCD and AKT/ERK signaling. Cell Death Dis. 2024;15(10):780. doi:10.1038/s41419-024-07171-x
58. Tan J, Lin G, Zhang R, et al. Bufotalin induces oxidative stress-mediated apoptosis by blocking the ITGB4/FAK/ERK pathway in glioblastoma. Antioxidants. 2024;13(10):1179. doi:10.3390/antiox13101179
59. Wang JF, Wang JS, Liu Y, et al. Knockdown of integrin β1 inhibits proliferation and promotes apoptosis in bladder cancer cells. BioFactors. 2024;51. doi:10.1002/biof.2150
60. Mai RY, Ye JZ, Gao X, et al. Up-regulated ITGB4 promotes hepatocellular carcinoma metastasis by activating hypoxia-mediated glycolysis and cancer-associated fibroblasts. Eur J Pharmacol. 2025;986:177102. doi:10.1016/j.ejphar.2024.177102
61. Chan XY, Chang KP, Yang CY, et al. Upregulation of ENAH by a PI3K/AKT/β-catenin cascade promotes oral cancer cell migration and growth via an ITGB5/Src axis. Cell Mol Biol Lett. 2024;29(1):136. doi:10.1186/s11658-024-00651-0
62. Zhan Z, Chen J, Zhou H, et al. Chronic alcohol consumption aggravates acute kidney injury through integrin β1/JNK signaling. Redox Biol. 2024;77:103386. doi:10.1016/j.redox.2024.103386
63. Nie J, Dang S, Zhu R, Lu T, Zhang W. ADAMTS18 deficiency associates extracellular matrix dysfunction with a higher risk of HER2-positive mammary tumorigenesis and metastasis. Breast Cancer Res. 2024;26(1):19. doi:10.1186/s13058-024-01771-3
64. Wang Y, Li X, Zhang T, et al. Neutrophils promote tumor invasion via FAM3C-mediated epithelial-to-mesenchymal transition in gastric cancer. Int J Bio Sci. 2023;19(5):1352–1368. doi:10.7150/ijbs.79022
65. Wang Y, Qin Y, Wu C, et al. OSU-T315 overcomes immunosuppression in triple-negative breast cancer by targeting the ILK/NF-κB signaling pathway to enhance immunotherapeutic efficacy. Int Immunopharmacol. 2024;143(Pt 3):113530. doi:10.1016/j.intimp.2024.113530
66. Pan C, Wang Z, Wang Q, et al. TFAP2A-activated ITGB4 promotes lung adenocarcinoma progression and inhibits CD4(+)/CD8(+) T-cell infiltrations by targeting NF-κB signaling pathway. Transl Lung Cancer Res. 2024;13(9):2116–2138. doi:10.21037/tlcr-24-50
67. Wang J, Wang Y, Zhu J, et al. Promoter hypermethylation-induced downregulation of ITGA7 promotes colorectal cancer proliferation and migration by activating the PI3K/AKT/NF-κB pathway. Biochim Biophys Acta Mol Cell Res. 2024;1871(7):119785. doi:10.1016/j.bbamcr.2024.119785
68. Yang MD, Sun Y, Zhou WJ, et al. Resveratrol enhances inhibition effects of cisplatin on cell migration and invasion and tumor growth in breast cancer MDA-MB-231 cell models in vivo and in vitro. Molecules. 2021;26(8). doi:10.3390/molecules26082204
69. Gutierrez DA, Contreras L, Villanueva PJ, et al. Identification of a potent cytotoxic pyrazole with anti-breast cancer activity that alters multiple pathways. Cells. 2022;11(2):254. doi:10.3390/cells11020254
70. Wang SC, Sun HL, Hsu YH, et al. α-Linolenic acid inhibits the migration of human triple-negative breast cancer cells by attenuating Twist1 expression and suppressing Twist1-mediated epithelial-mesenchymal transition. Biochem Pharmacol. 2020;180:114152. doi:10.1016/j.bcp.2020.114152
71. Ke AW, Zhang PF, Shen YH, et al. Generation and characterization of a tetraspanin CD151/integrin α6β1-binding domain competitively binding monoclonal antibody for inhibition of tumor progression in HCC. Oncotarget. 2016;7(5):6314–6322. doi:10.18632/oncotarget.6833
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.