Back to Journals » Journal of Multidisciplinary Healthcare » Volume 18
Causal Effects of Gut Microbiome on Tinnitus: A Mendelian Randomization Study
Authors Zeng Q, Xie W, Dong H, Liu X, Shao S, Chen L, Zhang W
Received 28 February 2025
Accepted for publication 16 June 2025
Published 22 July 2025 Volume 2025:18 Pages 4159—4172
DOI https://doi.org/10.2147/JMDH.S525502
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Scott Fraser
Qiang Zeng,1 Wei Xie,2 Huaqian Dong,2 Xing Liu,1 Shuai Shao,1 Li Chen,1 Wenwen Zhang1
1School of Nursing, Guizhou University of Traditional Chinese Medicine, Guiyang, 550002, People’s Republic of China; 2Department of Nursing, the First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine, Guiyang, 550001, People’s Republic of China
Correspondence: Wei Xie, Department of Nursing, the First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine, Guiyang, 550001, People’s Republic of China, Email [email protected]
Background: Previous studies have identified a clinical association between gut microbiota (GM) and tinnitus; however, the potential causal relationship remains unclear. Therefore, we conducted a Mendelian randomization(MR) study to evaluate the potential causal effects of 412 gut microbiome features on tinnitus risk.
Methods: In the study, we used pooled data from a genome-wide association study(GWAS) of gut microbes from a Dutch population. Summary statistics of tinnitus were drawn from the FinnGen R10 database. Inverse variance weighted, weighted median and MR-Egger were used to evaluate the potential causal link between GM and tinnitus.
Results: The MR analysis revealed a negative association between the abundance of Parabacteroides distasonis and the risk of tinnitus (OR = 0.84, 95% CI: 0.74– 0.94, p = 0.003), suggesting a potential protective role. Conversely, positive associations were observed for Eggerthella (OR = 1.11, 95% CI: 1.01– 1.22, p = 0.032), Alistipes onderdonkii, and Lachnospiraceae bacterium 5-1-63FAA, all linked to increased tinnitus risk.
Conclusion: Our findings suggest potential causal effects of specific gut microbial taxa and pathways on tinnitus, highlighting promising targets for prevention and treatment. However, further research is needed to validate these associations and clarify underlying mechanisms.
Keywords: causal relationship, gut microbiota, Mendelian randomization, tinnitus
Introduction
Tinnitus, defined as the perception of sound in the absence of external auditory stimuli, is a prevalent symptom affecting approximately 14.4% of adults worldwide (95% CI: 12.6%–16.5%).1 Among them, nearly 120 million individuals experience severe symptoms that profoundly impair quality of life, often manifesting as sleep disturbances, anxiety, and depression.2–4 This condition poses considerable challenges to both individuals and society by increasing healthcare utilization and reducing productivity, thereby imposing a substantial economic burden. Although current treatment —including sound therapy, cognitive behavioral therapy, and pharmacological interventions—offer symptom relief, their efficacy remains inconsistent and does not address the underlying pathophysiology.5 Recent advances have proposed the concept of a “gut–brain–ear axis” suggesting that gut microbiota (GM) may influence the onset and severity of chronic tinnitus through modulation of metabolic pathways and systemic inflammation.6–9
The GM, composed mainly of bacteria along with viruses, fungi, and other microorganisms,10,11 plays a fundamental role in regulating host metabolism, immunity, and neurological function.12 Growing evidence suggests that GM plays a pivotal role in the pathophysiology of various systemic and neuropsychiatric conditions, such as cardiovascular disease, metabolic disorders (including diabetes and obesity), sleep disturbances, and mood disorders like depression. Recent studies have also identified distinct microbial alterations in individuals with tinnitus, raising the possibility that GM imbalances may influence auditory processing or systemic inflammatory pathways. However, the precise causal relationship remains unclear.13,14 Several theories have been proposed regarding the underlying mechanisms of tinnitus, but none have been definitively proven. These gaps highlight the urgent need for mechanistic studies to clarify GM’s role in tinnitus pathogenesis and to support the development of more effective, targeted therapies.
Mendelian randomization (MR) is a statistical approach used to infer causal relationships between exposures and outcomes by leveraging genetic variations—typically single nucleotide polymorphisms (SNPs)—as instrumental variables (IVs).15 Because genetic variants are fixed at conception and not influenced by environmental confounders or reverse causation, MR offers significant advantages over traditional observational studies by effectively minimizing bias.16,17 The present research was conducted to explore the causal relationship between GM and its functional pathways and tinnitus from a genetic perspective.
Methods
Study Design
We assessed causal effects of GM pathways on tinnitus risk using two-sample MR. The design of this study is outlined in Figure 1. Framework of this study is shown in Figure 2. SNPs significantly associated with GM were selected as IVs, which were required to meet the following three core assumptions: (a) IVs should be strongly associated with the exposure; (b) IVs should influence the outcome exclusively through their effect on GM; and (c) IVs should be independent of any confounding factors unrelated to GM.18
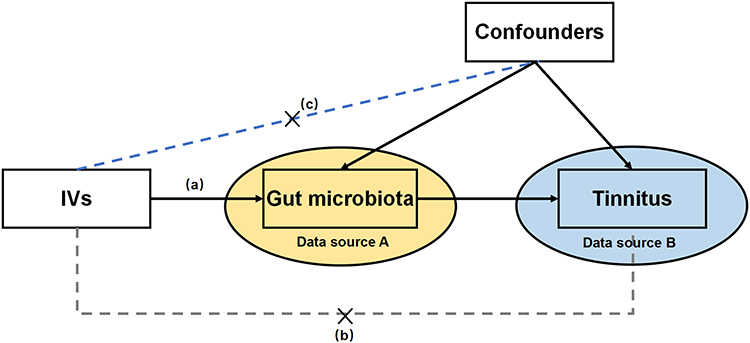 |
Figure 1 The design of this study. IVs: instrumental variables. |
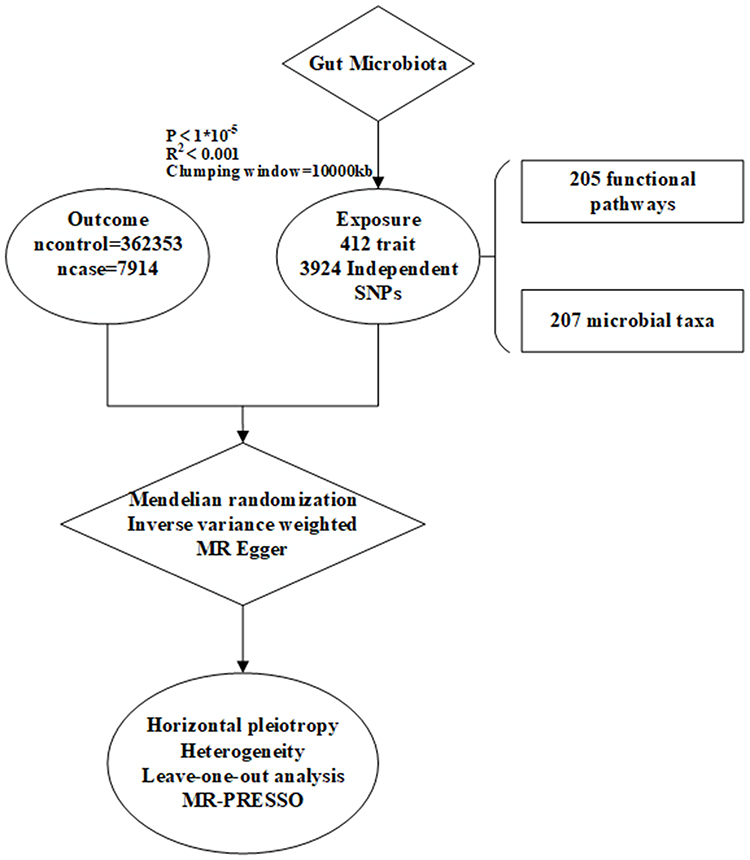 |
Figure 2 The flowchart of this study. |
Data Sources
In this study, genome-wide association study (GWAS) statistics—used to identify genetic variants linked to specific traits—were obtained from a Dutch population-based study of gut microbes. The dataset included 207 microbial taxa (5 phyla, 10 classes, 13 orders, 26 families, 48 genera, and 105 species) and 205 functional pathways. Summary statistics for microbiota metabolism pathways were extracted from the Dutch Microbiome Project (DMP) in 7738 European ancestry participants, in which the metabolism pathways were determined by shotgun metagenomic sequencing.19
The GWAS pooled data for tinnitus were derived from the pooled data (GWAS ID: H8_TINNITUS) published in the Finnish database R10 (https://www.finngen.fi/en) for individual of European origin. The study population consisted of 7914 tinnitus cases and 362353 control subjects. The pooled data used in this article were obtained from published databases, which are freely available for download and have received ethical approval.
Instruments Variables Selection
To identify potential associations between exposure and outcome, stringent criteria were employed for the selection of SNPs as IVs. The genome-wide significance threshold was initially set at P < 5 × 10–8. However, due to the limited number of SNPs meeting this criterion, the threshold was relaxed to P < 1 × 10–5 to allow for the inclusion of more IVs and ensure more robust and comprehensive results.20 This relaxed threshold has also been widely applied in previous MR studies. Additionally, the threshold for linkage disequilibrium(LD) was set at r2 < 0.001, with the search distance for LD values limited to 10,000 kb.21
To address potential bias from weak IVs, the F-statistic of each IV was calculated, applying a threshold of F-statistic > 10 to identify and exclude weak instruments.22 This approach was intended to enhance the robustness of our findings by ensuring the exclusion of instruments that could contribute to bias. Furthermore, ambiguous and palindromic SNPs were removed during the dataset refinement process to minimize potential sources of error.23
Statistical Analysis
In this MR study, we employed MR-Egger regression, weighted median, and IVW methods to investigate the causal relationship between GM composition and the risk of developing tinnitus. A circular heatmap was constructed to visually represent the relationships between the gut microbiome taxa and the risk of tinnitus, providing an intuitive depiction of the association strengths and directions. IVW served as the primary method, combines the Wald ratios of IVs in a meta-analytic framework, with its accuracy contingent upon the validity of all IVs and the absence of directional horizontal pleiotropy. While the IVW method offers high statistical power, its estimates may be biased if core MR assumptions are violated; therefore, to enhance the robustness of our findings, we employed MR-Egger regression and the weighted median method as complementary sensitivity analyses.
We used Cochran’s Q test to assess heterogeneity, considering p < 0.05 as evidence of significant heterogeneity.21,24 Moreover, the MR Pleiotropy Residual Sum and Outlier (MR-PRESSO) methodology were applied to scrutinize the presence of pleiotropic bias.25 Additionally, MR-Egger regression was conducted to assess horizontal pleiotropy, with a p-value < 0.05 suggesting potential violations of the assumptions inherent to MR analysis.26 Finally, to confirm the robustness of our findings, a leave-one-out sensitivity analysis was performed, wherein each SNP was systematically excluded to ensure that the conclusions were not unduly influenced by any individual variant.
R version 4.3.1 was utilized to conduct all the statistical analyses. Two sample MR and MR-PRESSO packages were used.
Results
Causal Association Between the Gut Microbiota Composition and the Risk of Developing Tinnitus
We performed MR analyses on 412 microbiome features and screened 3924 SNPs as IVs. The circular plot displays the results of the MR analysis for the IVs, with the IVW method as the primary approach used in this analysis (Figure 3). In our MR analysis, 4 GM abundances and 10 gut bacterial pathway abundances initially showed suggestive evidence of causal associations with the risk of developing tinnitus (Figure 4). The species Parabacteroides distasonis was nominally associated with a reduced risk of tinnitus, with an odds ratio (OR) of 0.84 and a 95% confidence interval (CI) of 0.74–0.94 (p = 0.003), indicating a potential protective effect. In contrast, Eggerthella (OR = 1.11, 95% CI: 1.01–1.22, p = 0.032), Alistipes onderdonkii(OR = 1.16, 95% CI: 1.01–1.34, p = 0.041), and Lachnospiraceae bacterium 5-1-63FAA(OR = 1.07, 95% CI: 1.00–1.15, p = 0.042) were positively associated with tinnitus risk. However, these associations did not remain statistically significant after correction for multiple testing using the false discovery rate (FDR), and therefore should be interpreted with caution as they may represent false-positive findings.
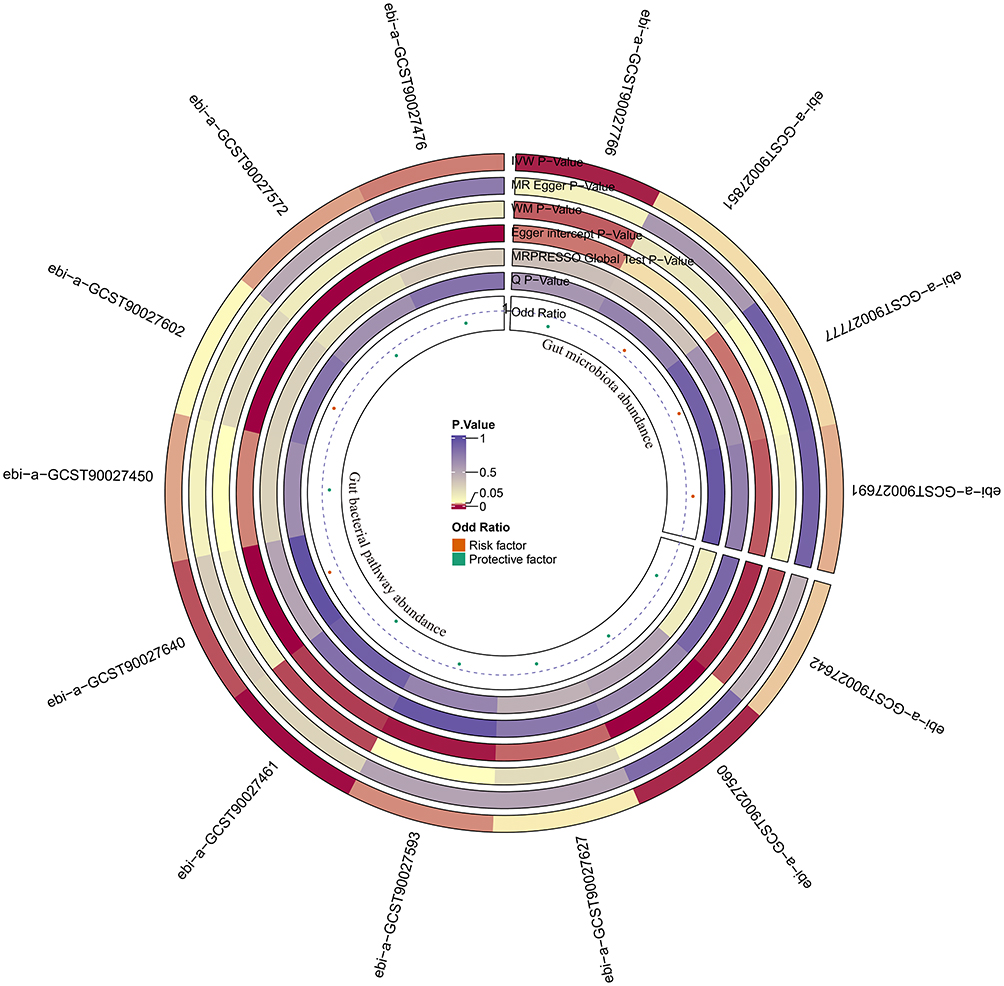 |
Figure 3 The circular plot showing the MR results for the GM. |
 |
Figure 4 Forest plot of the associations between gut microbiota and Pathway with the risk of tinnitus. |
We also identified 10 gut microbial functional pathways: chorismite biosynthesis I, dTDP-L-rhamnose biosynthesis I, glycogen biosynthesis I(from ADP-D Glucose), superpathway of menaquinol 6 biosynthesis I, S-adenosyl L methionine cycle I, cinnamate and 3 hydroxycinnamate degradation to 2 oxopent 4 enoate, mannan degradation, and superpathway of L-serine and glycine biosynthesis I are associated with a reduced risk of developing tinnitus (p<0.05). Conversely, thiamin salvage II and L-rhamnose degradation I, which may have a causal relationship with increased tinnitus risk (p<0.05). Unfortunately, none of the aforementioned causal effects remained significant after correction for the FDR (p > 0.05).The causal estimates in the MR analysis for the above results are presented in Table 1. Scatter plots are presented in Figure 5.
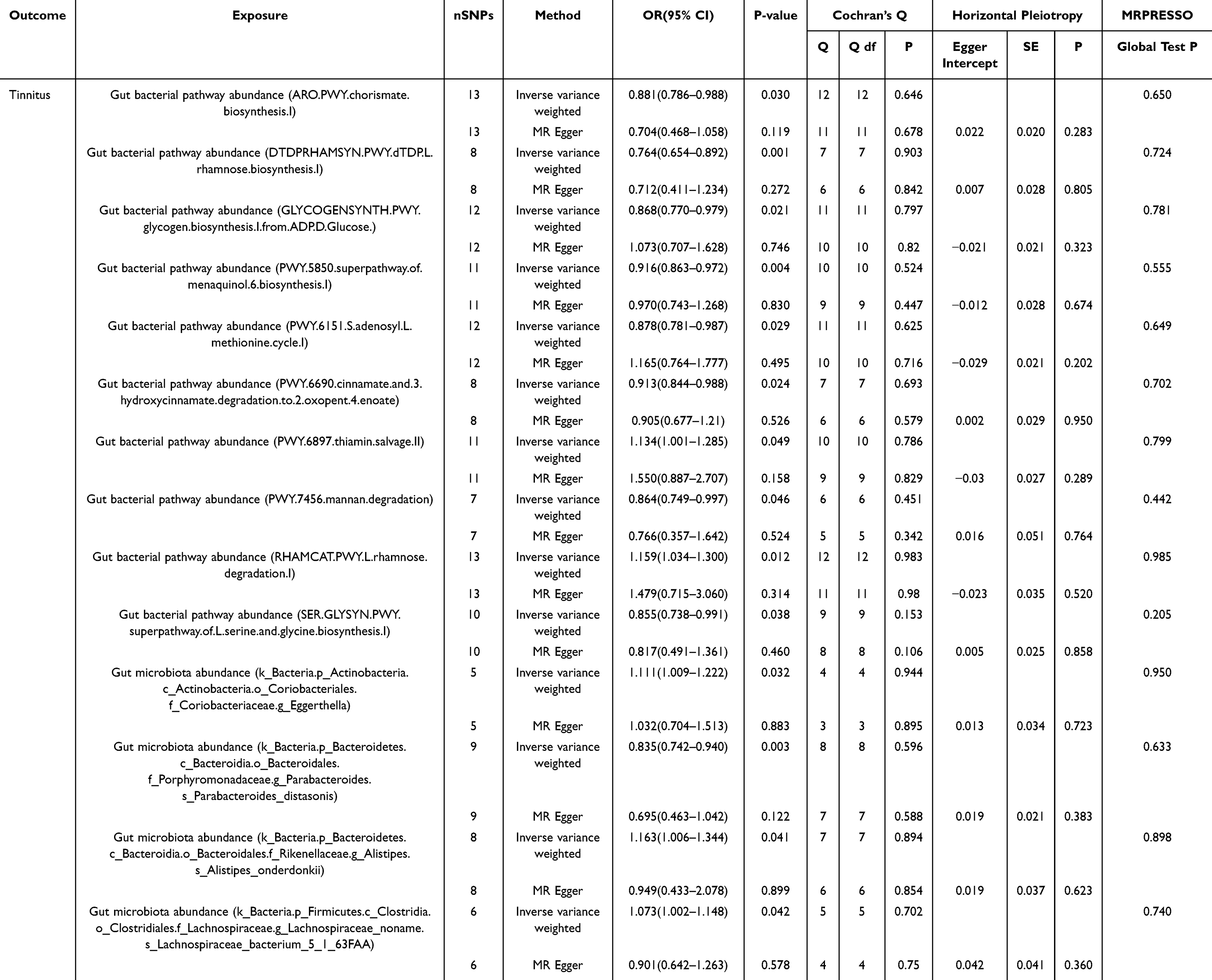 |
Table 1 MR Estimates and Sensitivity Analyses of the Causal Effects in the Gut Microbiota on Tinnitus |
 |
Figure 5 Scatter plots of MR results of gut microbiota on tinnitus, including (A) chorismate biosynthesis I, (B) dTDP-L-rhamnose biosynthesis I, (C) glycogen biosynthesis I (from ADP-D-glucose), (D) superpathway of menaquinol-6 biosynthesis I, (E) S-adenosyl-L-methionine cycle I, (F) cinnamate and 3-hydroxycinnamate degradation to 2-oxopent-4-enoate, (G) thiamin salvage II, (H) mannan degradation, (I) L-rhamnose degradation I, (J) superpathway of L-serine and glycine biosynthesis I, (K) genus Eggerthella, (L) species Parabacteroides distasonis, (M) species Alistipes onderdonkii, and (N) species Lachnospiraceae bacterium 5-1-63FAA. |
Sensitivity Analysis
The SNPs used in the MR analysis all exhibited F-statistics greater than 10, which helps mitigate the potential bias from weak IVs. To assess the reliability of the remaining SNPs, a leave-one-out approach was applied, systematically removing one SNP at a time. The results clearly demonstrated that the effects of the remaining SNPs remained largely unchanged upon the exclusion of any individual SNP. Both Cochran’s Q test and the MR-Egger intercept yielded p-values greater than 0.05, indicating no significant heterogeneity among the IVs and no evidence of directional pleiotropy (Figure 4). Additionally, MR-PRESSO and leave-one-out analyses detected no influential outliers, and no evidence of heterogeneity or horizontal pleiotropy was observed, supporting the overall robustness of the IVW results (Figure 6). The funnel plot indicates the absence of publication bias in this study (Figure 7).
 |
Figure 6 Leave-one-out sensitivity analyses for the causal effects of gut microbiota on the risk of tinnitus, including (A) chorismate biosynthesis I, (B) dTDP-L-rhamnose biosynthesis I, (C) glycogen biosynthesis I (from ADP-D-glucose), (D) superpathway of menaquinol-6 biosynthesis I, (E) S-adenosyl-L-methionine cycle I, (F) cinnamate and 3-hydroxycinnamate degradation to 2-oxopent-4-enoate, (G) thiamin salvage II, (H) mannan degradation, (I) L-rhamnose degradation I, (J) superpathway of L-serine and glycine biosynthesis I, (K) genus Eggerthella, (L) species Parabacteroides distasonis, (M) species Alistipes onderdonkii, and (N) species Lachnospiraceae bacterium 5-1-63FAA. |
 |
Figure 7 Funnel plots for the causal effects of gut microbiota on the risk of tinnitus, including (A) chorismate biosynthesis I, (B) dTDP-L-rhamnose biosynthesis I, (C) glycogen biosynthesis I (from ADP-D-glucose), (D) superpathway of menaquinol-6 biosynthesis I, (E) S-adenosyl-L-methionine cycle I, (F) cinnamate and 3-hydroxycinnamate degradation to 2-oxopent-4-enoate, (G) thiamin salvage II, (b) mannan degradation, (I) L-rhamnose degradation I, (J) superpathway of L-serine and glycine biosynthesis I, (K) genus Eggerthella, (L) species Parabacteroides distasonis, (M) species Alistipes onderdonkii, and (N) species Lachnospiraceae bacterium 5-1-63FAA. |
Discussion
The etiology of tinnitus is multifactorial, involving auditory, neurological, and psychological components. Increasing evidence suggests that the GM may influence distant organ systems through immune modulation, metabolic pathways, and the gut–brain axis, potentially affecting neurological functions relevant to auditory processing.27–29 Building on these insights, our study employed a two-sample MR framework to explore whether alterations in GM composition may causally influence tinnitus risk.30–33
The results of the present study suggest that Parabacteroides distasonis may have a protective role in preventing tinnitus development, and the underlying mechanisms may be manifested through the multilayered regulation of the gut-brain axis.34 As a beneficial symbiotic bacterium, Parabacteroides distasonis may promote intestinal barrier function and mucosal immune homeostasis through the production of metabolites such as short-chain fatty acids, and influence the inflammatory microenvironment and neuronal excitability in the central nervous system via vagal signaling and alteration of blood-brain barrier permeability.35,36 In addition, the flora may alleviate persistent abnormal discharges and neural hyperexcitability in the auditory center by modulating the levels of immune mediators and neurotransmitters GABA between the gut and the central nervous system.37,38 This multidimensional immune-metabolic-neuromodulatory effect helps to alleviate tinnitus-associated central inflammation and increased neural excitability, thus exerting a neuroprotective effect.39
However, changes in the composition of gut flora such as Eggerthella, Alistipes onderdonkii, and Lachnospiraceae bacterium 5-1-63FAA may contribute to tinnitus by adversely affecting neuroinflammation and metabolic homeostasis in the auditory center via the gut-brain axis. Bacteria of the genus Eggerthella are often associated with inflammatory intestinal environments and may place central auditory pathways in a susceptible environment by increasing systemic low-grade inflammation through elevated intestinal epithelial permeability. Metabolites of Alistipes onderdonkii have been associated with negative mood and depressive symptoms in established studies, and may indirectly affect auditory pathways by interfering with neurotransmitter synthesis and metabolic pathways, such as 5-HT and GABA.40 Anabolic pathways and indirectly affecting the central modulation process of auditory signals. Specific strains of Lachnospiraceae, on the other hand, may play complex roles in short-chain fatty acid production, bile acid metabolism, and inflammatory regulation, which, when disrupted, may exacerbate neuroinflammatory responses and neuronal hyperexcitability.41,42 Together, these changes contribute to the onset and persistence of tinnitus by placing the auditory centers in a state of hyperexcitability and abnormal signaling.43
In terms of metabolic pathways, the GM may modulate central nervous system (CNS) function and inflammatory homeostasis through diverse metabolic and signaling mechanisms, thereby contributing to the prevention of tinnitus.8 In particular, the chorismite biosynthesis pathway is involved in the production of aromatic amino acids and their downstream metabolites, which are critically important for maintaining the balance of central neurotransmitters GABA and the stability of the neuroinflammatory milieu.44 Simultaneously, the dTDP-L-rhamnose biosynthesis pathway influences bacterial cell wall structure and immune regulation, indirectly affecting gut-brain axis communication and the neural microenvironment.45 At the level of energy metabolism, glycogen biosynthesis provides a stable energy reservoir for symbiotic bacteria and supports the generation of beneficial metabolites.46,47 This metabolic advantage may reduce excessive neuronal excitability within central auditory pathways by modulating short-chain fatty acid production and the permeability of the blood-brain barrier. Furthermore, vitamin K2 produced via the menaquinol 6 biosynthesis pathway may exert neuroprotective and anti-inflammatory effects, buffering hyperexcitable auditory circuits in the CNS.48
It’s worth noting that the maintenance of a balanced S-adenosyl-L-methionine cycle helps prevent maladaptive neuroplastic changes in central auditory pathways.49 Concurrently, the degradation of cinnamate and related compounds may alleviate the harmful impact of potentially neurotoxic metabolites.50 Additionally, enhancing mannan degradation and optimizing L-serine and glycine biosynthesis pathways could improve host amino acid and polysaccharide metabolism, thereby augmenting the availability of central inhibitory neurotransmitters glycine and preserving excitatory-inhibitory balance within auditory networks.51 Multiple studies have supported this concept, indicating that modulating GM and their metabolic products may provide a novel avenue for mitigating tinnitus and other chronic neurological symptoms.52–54
In addition, thiamine is essential for maintaining normal energy metabolism and neurotransmitter synthesis in the CNS. Alterations in the thiamine recycling II pathway may lead to abnormal thiamine utilization and conversion by the intestinal flora, thus indirectly affecting the host’s vitamin B₁ status.55,56 The thiamine salvage II pathway (nominally associated, p = 0.049) may plausibly influence tinnitus via neuronal energy metabolism, though our MR did not directly test this pathway.57 This pathway may plausibly influence tinnitus through its role in neuronal energy metabolism, as thiamine deficiency or dysregulated thiamine metabolism has been shown to impair CNS energy supply, potentially leading to neuronal hyperexcitability and abnormal firing—factors that have been implicated in the onset and persistence of tinnitus.58,59 Meanwhile, abnormalities in the L-rhamnose degradation I pathway may involve changes in cell wall structure and metabolite release in specific bacterial colonies. Since L-rhamnose is an important component of certain bacterial cell wall polysaccharides, disruption of this degradation pathway may alter microbial composition and adhesion properties, weaken gut barrier function and impair gut-brain axis signaling.60 These changes may contribute to the pathophysiological process of tinnitus by enhancing CNS inflammatory responses through pro-inflammatory signaling and metabolic mediators and increasing the state of hyperexcitability of auditory pathways. For example, Mohajeri emphasized the bidirectional effects of microbial metabolites on CNS health and disease, and Cryan elucidated the importance of microbial diversity in maintaining neurochemical homeostasis through the gut-brain axis.61,62 In addition, Zeng showed that specific beneficial strains can modulate the host’s immune and metabolic environment through metabolic pathways, thereby influencing the pathologic process of CNS.63 Overall, identifying and targeting specific bacterial metabolic pathways may provide new insights into understanding and intervening in the central pathogenesis of tinnitus.
Our study employs a two-sample MR approach to rigorously examine the causal relationship between GM and tinnitus, leveraging the principles of Mendelian genetics to minimize confounding and reverse causality inherent in observational studies.64 By utilizing publicly available GWAS datasets, we efficiently identify genetic instruments without incurring additional financial costs, thereby enhancing the robustness of our findings. The integration of genetic perspectives into the GM and tinnitus association offers significant public health implications, potentially informing preventive strategies and emphasizing the importance of early screening and diagnosis. However, our analysis is constrained by the limited sample size of the GWAS datasets, necessitating larger studies to secure more reliable IVs. Additionally, the use of a less stringent SNP selection threshold (p <1×10–5) may introduce imprecise proxies, and the predominantly European ancestry of the study population limits the generalizability of our results. Concerns regarding pleiotropy and the biological functions of the genetic instruments further highlight the need for adherence to MR assumptions and underscore the necessity for experimental validation to elucidate the specific pathways through which GM influences tinnitus. Future research should address these limitations to reinforce the causal inferences and expand the applicability of our findings across diverse populations.
Conclusion
In summary, our two-sample MR study identified several nominal associations between gut microbial taxa, metabolic pathways, and the risk of tinnitus. Although these findings did not remain statistically significant after FDR correction, they offer preliminary insights into potential microbiota-related mechanisms underlying tinnitus. Notably, taxa such as Parabacteroides distasonis may be particularly interesting for future investigation. Given the limitations of the current evidence, robust validation through larger studies and randomized controlled trials is warranted to confirm these associations and elucidate the underlying biological pathways.
Ethics Approval
This research is based on publicly available abstract-level data obtained from comprehensive genome-wide association studies (GWAS). According to the national legislation guidelines, such as item 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects dated February 18, 2023, China, this study is exempt from ethical approval.
Acknowledgments
We would like to thank the researchers who collected and organized the shared IEU OpenGWAS data and FinnGen dataset.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work is supported by the National Natural Science Foundation of China (NSFC, No. 82060832) and the Traditional Chinese Medicine Nursing (Ethnomedicine Nursing) Inheritance and Innovation Talent Team (GZYYFY2025006).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Jarach CM, Lugo A, Scala M, et al. Global prevalence and incidence of tinnitus: a systematic review and meta-analysis. JAMA Neurol. 2022;79(9):888–900. doi:10.1001/jamaneurol.2022.2189
2. Langguth B, Landgrebe M, Kleinjung T, Sand GP, Hajak G. Tinnitus and depression. World J Biol Psychiatry. 2011;12(7):489–500. doi:10.3109/15622975.2011.575178
3. Gupta J, Gupta R. a scoping new revolution for tinnitus management: a neuromonics narrative review. Curr Drug Targets. 2023;24(3):274–286. doi:10.2174/1389450124666221220165137
4. Molnár A, Mavrogeni P, Tamás L, Maihoub S. correlation between tinnitus handicap and depression and anxiety scores. Ear Nose Throat J. 2022;1455613221139211.
5. Chari DA, Limb CJ. Tinnitus. Med Clin North Am. 2018;102(6):1081–1093. doi:10.1016/j.mcna.2018.06.014
6. Maihoub S, Mavrogeni P, Molnár V, Molnár A. Tinnitus and its comorbidities: a comprehensive analysis of their relationships. J Clin Med. 2025;14(4):1285. doi:10.3390/jcm14041285
7. Vitale G, Dicitore A, Barrea L, et al. From microbiota toward gastro-enteropancreatic neuroendocrine neoplasms: are we on the highway to hell? Rev Endocr Metab Disord. 2021;22(3):511–525. doi:10.1007/s11154-020-09589-y
8. Wang J, Xiang JH, Peng XY, et al. Characteristic alterations of gut microbiota and serum metabolites in patients with chronic tinnitus: a multi-omics analysis. Microbiol Spectr. 2024;e0187824.
9. Zhuang M, Zhang X, Cai J. Microbiota-gut-brain axis: interplay between microbiota, barrier function and lymphatic system. Gut Microbes. 2024;16(1):2387800. doi:10.1080/19490976.2024.2387800
10. Blaut M. Ecology and physiology of the intestinal tract. Curr Top Microbiol Immunol. 2013;358:247–272. doi:10.1007/82_2011_192
11. Voreades N, Kozil A, Weir TL. Diet and the development of the human intestinal microbiome. Front Microbiol. 2014;5:494. doi:10.3389/fmicb.2014.00494
12. Tobaldini E, Costantino G, Solbiati M, et al. Sleep, sleep deprivation, autonomic nervous system and cardiovascular diseases. Neurosci Biobehav Rev. 2017;74(Pt B):321–329. doi:10.1016/j.neubiorev.2016.07.004
13. Lee SY, Nam DW, Koo JW, De Ridder D, Vanneste S, Song JJ. No auditory experience, no tinnitus: lessons from subjects with congenital- and acquired single-sided deafness. Hear Res. 2017;354:9–15. doi:10.1016/j.heares.2017.08.002
14. Lee JM, Kim Y, Ji JY, Koo JW, Song JJ. Auditory experience, for a certain duration, is a prerequisite for tinnitus: lessons from subjects with unilateral tinnitus in the better-hearing ear. Prog Brain Res. 2021;260:223–233.
15. Verduijn M, Siegerink B, Jager KJ, Zoccali C, Dekker FW. Mendelian randomization: use of genetics to enable causal inference in observational studies. Nephrol Dial Transplant. 2010;25(5):1394–1398. doi:10.1093/ndt/gfq098
16. Smith GD, Ebrahim S. Mendelian randomization: prospects, potentials, and limitations. Int J Epidemiol. 2004;33(1):30–42. doi:10.1093/ije/dyh132
17. Bochud M. On the use of Mendelian randomization to infer causality in observational epidemiology. Eur Heart J. 2008;29(20):2456–2457. doi:10.1093/eurheartj/ehn428
18. Larsson SC, Butterworth AS, Burgess S. Mendelian randomization for cardiovascular diseases: principles and applications. Eur Heart J. 2023;44(47):4913–4924. doi:10.1093/eurheartj/ehad736
19. Lopera-Maya EA, Kurilshikov A, van der Graaf A, et al. Effect of host genetics on the gut microbiome in 7,738 participants of the Dutch microbiome project. Nat Genet. 2022;54(2):143–151. doi:10.1038/s41588-021-00992-y
20. Kurki MI, Karjalainen J, Palta P, et al. Author Correction: finnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;615(7952):E19. doi:10.1038/s41586-023-05837-8
21. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32(5):377–389. doi:10.1007/s10654-017-0255-x
22. Burgess S, Thompson SG. Bias in causal estimates from Mendelian randomization studies with weak instruments. Stat Med. 2011;30(11):1312–1323. doi:10.1002/sim.4197
23. Rogne T, Gill D, Liew Z, et al. Mediating factors in the association of maternal educational level with pregnancy outcomes: a Mendelian randomization study. JAMA Netw Open. 2024;7(1):e2351166. doi:10.1001/jamanetworkopen.2023.51166
24. Burgess S, Small DS, Thompson SG. A review of instrumental variable estimators for Mendelian randomization. Stat Methods Med Res. 2017;26(5):2333–2355. doi:10.1177/0962280215597579
25. Verbanck M, Chen CY, Neale B, Do R. Publisher correction: detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018;50(8):1196. doi:10.1038/s41588-018-0164-2
26. Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44(2):512–525. doi:10.1093/ije/dyv080
27. Mottawea W, Yousuf B, Sultan S, et al. Multi-level analysis of gut microbiome extracellular vesicles-host interaction reveals a connection to gut-brain axis signaling. Microbiology Spectrum. 2024;13.
28. Nguyen Q-D, Fujii K, Ishibashi K, et al. Regulation of gut starvation responses through drosophila NP3253 neurons. Genes Cells. 2025;302.
29. Duan J, Li Q, Cheng Y, Zhu W, Liu H, Li F. Therapeutic potential of parabacteroides distasonis in gastrointestinal and hepatic disease. MedComm. 2024;5.
30. Mou Y, Du Y, Zhou L, et al. Gut microbiota interact with the brain through systemic chronic inflammation: implications on neuroinflammation, neurodegeneration, and aging. Front Immunol. 2022;13:796288. doi:10.3389/fimmu.2022.796288
31. Fung TC. The microbiota-immune axis as a central mediator of gut-brain communication. Neurobiol Dis. 2020;136:104714. doi:10.1016/j.nbd.2019.104714
32. Kowalski K, Mulak A. Brain-gut-microbiota axis in alzheimer’s disease. J Neurogastroenterol Motil. 2019;25(1):48–60. doi:10.5056/jnm18087
33. Rutsch A, Kantsjö JB, Ronchi F. The gut-brain axis: how microbiota and host inflammasome influence brain physiology and pathology. Front Immunol. 2020;11:604179. doi:10.3389/fimmu.2020.604179
34. Cuffaro B, Assohoun ALW, Boutillier D, et al. In vitro characterization of gut microbiota-derived commensal strains: selection of parabacteroides distasonis strains alleviating TNBS-induced colitis in mice. Cells. 2020;9(9):2104. doi:10.3390/cells9092104
35. Gaifem J, Mendes-Frias A, Wolter M, et al. Akkermansia muciniphila and Parabacteroides distasonis synergistically protect from colitis by promoting ILC3 in the gut. mBio. 2024;15(4):e0007824. doi:10.1128/mbio.00078-24
36. Koh GY, Kane AV, Wu X, Crott JW. Parabacteroides distasonis attenuates tumorigenesis, modulates inflammatory markers and promotes intestinal barrier integrity in azoxymethane-treated A/J mice. Carcinogenesis. 2020;41(7):909–917. doi:10.1093/carcin/bgaa018
37. Vaher K, Cabez MB, Parga PL, et al. The neonatal gut microbiota: a role in the encephalopathy of prematurity. Cell Rep Med. 2024;5(12):101845. doi:10.1016/j.xcrm.2024.101845
38. Garcez FB, Garcia de Alencar JC, Fernandez SSM, et al. Association between gut microbiota and delirium in acutely ill older adults. J Gerontol a Biol Sci Med Sci. 2023;78(8):1320–1327. doi:10.1093/gerona/glad074
39. Noreña AJ, Farley BJ. Tinnitus-related neural activity: theories of generation, propagation, and centralization. Hear Res. 2013;295:161–171. doi:10.1016/j.heares.2012.09.010
40. Shulman A, Wang W, Luo H, Bao S, Searchfield G, Zhang J. Neuroinflammation and Tinnitus. In: Searchfield GD, Zhang J, editors. The Behavioral Neuroscience of Tinnitus. Cham: Springer International Publishing; 2021:161–174.
41. Liu X, Zhang Y, Li W, et al. Fucoidan ameliorated dextran sulfate sodium-induced ulcerative colitis by modulating gut microbiota and bile acid metabolism. J Agric Food Chem. 2022;70(47):14864–14876. doi:10.1021/acs.jafc.2c06417
42. Seekatz AM, Theriot CM, Rao K, et al. Restoration of short chain fatty acid and bile acid metabolism following fecal microbiota transplantation in patients with recurrent Clostridium difficile infection. Anaerobe. 2018;53:64–73. doi:10.1016/j.anaerobe.2018.04.001
43. Ventura I, Chomon-García M, Tomás-Aguirre F, et al. Therapeutic and immunologic effects of short-chain fatty acids in inflammatory bowel disease: a systematic review. Int J Mol Sci. 2024;25(20):10879. doi:10.3390/ijms252010879
44. Shende VV, Bauman KD, Mosore BS. The shikimate pathway: gateway to metabolic diversity. Nat Prod Rep. 2024;41(4):604–648. doi:10.1039/D3NP00037K
45. Abdel-Haq R, Schlachetzki JCM, Glass CK, Mazmanian SK. Microbiome-microglia connections via the gut-brain axis. J Exp Med. 2019;216(1):41–59. doi:10.1084/jem.20180794
46. Chandra G, Chater KF, Bornemann S. Unexpected and widespread connections between bacterial glycogen and trehalose metabolism. Microbiology. 2011;157(Pt 6):1565–1572. doi:10.1099/mic.0.044263-0
47. Morin M, Ropers D, Cinquemani E, Portais JC, Enjalbert B, Cocaign-Bousquet M. The csr system regulates Escherichia coli fitness by controlling glycogen accumulation and energy levels. mBio. 2017;8(5). doi:10.1128/mBio.01628-17
48. Chatterjee K, Mazumder PM, Sarkar SR, et al. Neuroprotective effect of Vitamin K2 against gut dysbiosis associated cognitive decline. Physiol Behav. 2023;269:114252. doi:10.1016/j.physbeh.2023.114252
49. Sharma A, Gerbarg P, Bottiglieri T, et al. S-Adenosylmethionine (SAMe) for Neuropsychiatric Disorders. J Clin Psychiatry. 2017;78(6):e656–e667. doi:10.4088/JCP.16r11113
50. Drakontaeidi A, Pontiki E. Multi-target-directed cinnamic acid hybrids targeting alzheimer’s disease. Int J Mol Sci. 2024;25(1):582. doi:10.3390/ijms25010582
51. Zhang X, Ning Y, Chai L, Yin Y, Luo D, Xu W. Physicochemical properties and in vitro digestive behavior of astaxanthin loaded Pickering emulsion gel regulated by konjac glucomannan and κ-carrageenan. Int J Biol Macromol. 2024;278(Pt 2):134710. doi:10.1016/j.ijbiomac.2024.134710
52. De Caro C, Iannone LF, Citraro R, et al. Can we ‘seize’ the gut microbiota to treat epilepsy? Neurosci Biobehav Rev. 2019;107:750–764. doi:10.1016/j.neubiorev.2019.10.002
53. Guo R, Chen LH, Xing C, Liu T. Pain regulation by gut microbiota: molecular mechanisms and therapeutic potential. Br J Anaesth. 2019;123(5):637–654. doi:10.1016/j.bja.2019.07.026
54. Ullah H, Tovchiga O, Daglia M, Khan H. Modulating gut microbiota: an emerging approach in the prevention and treatment of multiple sclerosis. Curr Neuropharmacol. 2021;19(11):1966–1983. doi:10.2174/1570159X19666210217084827
55. Bozic I, Lavrnja I. Thiamine and benfotiamine: focus on their therapeutic potential. Heliyon. 2023;9(11):e21839. doi:10.1016/j.heliyon.2023.e21839
56. Hurt JK, Coleman JL, Fitzpatrick BJ, et al. Prostatic acid phosphatase is required for the antinociceptive effects of thiamine and benfotiamine. PLoS One. 2012;7(10):e48562. doi:10.1371/journal.pone.0048562
57. Orlova S, Nikitina E, Balashova N, et al. The role of individual neuronutrients in optimizing nutritional therapy for neurological patients. Medical Alphabet;2023. 36–47. 10.33667/2078-5631-2023-8-36-47
58. Butterworth RF. Thiamin deficiency and brain disorders. Nutr Res Rev. 2003;16(2):277–284. doi:10.1079/NRR200367
59. Ke ZJ, DeGiorgio LA, Volpe BT, Gibson GE. Reversal of thiamine deficiency-induced neurodegeneration. J Neuropathol Exp Neurol. 2003;62(2):195–207. doi:10.1093/jnen/62.2.195
60. Ulluwishewa D, Nicholls G, Henderson H, et al. Effects of bovine whey protein on exercise-induced gut permeability in healthy adults: a randomised controlled trial. Eur J Appl Physiol. 2024;124(7):2045–2056. doi:10.1007/s00421-024-05423-4
61. Keane L, Cryan JF, Gleeson JP. Exploiting the gut microbiome for brain tumour treatment. Trends Mol Med. 2024;31:213–223. doi:10.1016/j.molmed.2024.08.008
62. Tran SM, Mohajeri MH. The role of gut bacterial metabolites in brain development, aging and disease. Nutrients. 2021;13(3):732. doi:10.3390/nu13030732
63. Long J, Zhang J, Zeng X, Wang M, Wang N. Prevention and treatment of alzheimer’s disease via the regulation of the gut microbiota with traditional Chinese medicine. CNS Neurosci Ther. 2024;30(11):e70101. doi:10.1111/cns.70101
64. Zheng J, Baird D, Borges MC, et al. Recent developments in Mendelian randomization studies. Curr Epidemiol Rep. 2017;4(4):330–345. doi:10.1007/s40471-017-0128-6
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.