Back to Journals » Nature and Science of Sleep » Volume 17

Can the Sun Prevent Weekend Sleep Advance After Early Weekday Wakeups?
Authors Putilov AA ![]() , Verevkin EG
, Verevkin EG ![]()
Received 29 May 2025
Accepted for publication 26 July 2025
Published 22 August 2025 Volume 2025:17 Pages 1895—1913
DOI https://doi.org/10.2147/NSS.S543386
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Ahmed BaHammam
Arcady A Putilov,1,2,* Evgeniy G Verevkin1,*
1Independent Research Group, Biomedical Systems Math-Modeling, Berlin, Germany; 2Laboratory of Sleep/Wake Neurobiology, Institute of Higher Nervous Activity and Neurophysiology of the Russian Academy of Sciences, Moscow, Russia
*These authors contributed equally to this work
Correspondence: Arcady A Putilov, Email [email protected]
Purpose: There is sparse evidence on how circadian sleep timing is affected by 5 days on/2 days off school/work schedule. In an in silico study, we applied a model of sleep-wake regulation to highlight the difference in predictions based on two alternative explanations proposing either sun time or social time (eg, either solar midday or weekday risetime) as the major contributor to light entrainment of sleep timing. Self-reported sleep times were then used to confirm these predictions.
Methods: The difference between earlier and later weekday risers in weekend sleep timing their difference in weekday risetime were compared. This difference in weekday risetime is equal to the sum of differences in sleep phase shift on weekends and sleep loss on weekdays measured as the differences in weekend risetime and weekend-weekday gap in risetime, respectively. Three sets of samples were used for these estimations: 87 and 100 paired samples obtained before vs during lockdown and during early vs later school start time (five and three age subsets, respectively) and 1250 vs 1192 unpaired samples with weekday risetime not earlier vs later than 7 a.m. (five age subsets).
Results: In any age subset, a shift in social time (weekday risetime) caused a shift in weekend sleep phase (weekend risetime) due to a shift in the 24-h pattern of exposure to light, but this sleep phase shift was less pronounced than the shift in social time.
Conclusion: Both social time and sun time substantially contribute to the light entrainment of circadian sleep timing.
The Plain Language Summary: There is sparse evidence on how the circadian timing of sleep is affected by 5 days on/2 days off school/work schedule. We applied a model of sleep-wake regulation to highlight the difference in predictions based on two alternative explanations, proposing either sun time or social time as the major contributor to light entrainment of sleep timing. The sets of samples for the estimation of these contributions included 87 and 100 paired samples obtained before vs during lockdown and during early vs later school start time (five and three age subsets, respectively), and 1250 vs 1192 unpaired samples with weekday risetime not earlier vs later than 7 a.m. (five age subsets). A shift in social time (weekday risetime) associated with a shift in the weekend sleep phase (weekend risetime) was found in each age subset. It reflected a shift in the 24-h pattern of exposure to artificial and natural light sources. However, this shift in sleep phase was less pronounced than the shift in social time. Therefore, the shift in social time (weekday risetime) resulted in an additional loss of sleep on weekdays (weekend-weekday gap in risetime). It seems that the circadian and homeostatic regulators of the sleep-wake cycle cannot provide full adjustment to early weekday wakeups. Special interventions might be recommended to increase the contribution of advancing shift of circadian sleep timing at the expense of weekday sleep loss.
Keywords: sleep duration, sleep timing, sleep-wake regulating processes, school start time, lockdown
Introduction
Under natural environmental conditions, endogenous circadian clocks are entrained to the external light-dark cycle with 24-h period.1 These entrained clocks interact with the homeostatic drive for sleep to establish the 24-h periodicity of the sleep-wake cycle.2,3 Therefore, the circadian and homeostatic processes of sleep-wake regulation are expected to be the endogenous determinants of bed- and risetimes in our species.2 However, the bed- and risetimes in the conditions of modern post-industrial societies can be at odds with those determined by these endogenous sleep-wake regulators. These conditions often require either advancing or delaying shifts of bed- and risetimes relative to the times determined by the endogenous sleep-wake regulators. In particular, when adolescents and adults are learning/working on 5 days on/2 days off schedule, they are forced to sleep earlier and shorter during days on (eg, weekdays) compared to days off (eg, weekends). Under the unchanged 24-h pattern of light exposure, the phase of circadian clocks cannot be changed by a shift in risetime in earlier hours or by a shift in bedtime in later hours.4 Therefore, Wittmann et al5 proposed a concept of the conflicting clocks to highlight the problem of people who, instead of waking up at the time dictated by their internal (biological) clocks, are waking up earlier due to the demands of their work-rest schedule dictated by the so-called “social clocks”.5 If risetimes after ad libitum sleep on days off can be mostly determined by the biological clocks, risetimes on the preceding and following days on are determined by these “social clocks”. Consequently, biological clocks fail to compensate for a shift in weekday risetimes on an earlier hour by a similarly large advancing shift in bedtimes. As a result, sleep duration on weekdays is reduced.5
There is sparse evidence on how the circadian timing of sleep is affected by the common 5 days on 2 days off school/work schedule. It remains to be elucidated whether social schedules (eg, early school/work start times) are capable to shift the phase of biological clocks and sleep timing. Two alternative explanations are proposed. They agreed that light is the primary stimulus that entrains human circadian clocks and sets bed- and risetimes by interacting with the homeostatic drive for sleep. However, these explanations differ in postulating that either sun time (eg, the sun’s position in the sky) or social time (eg, early school/work start times) plays a crucial role in light entrainment of the circadian clock governing the sleep-wake cycle.6 Wittmann et al5 (the authors of the concept of conflicting clocks) postulated that the natural light-dark cycle is the major stimulus that entrains human circadian rhythms and that the phase relationship between sun time (eg, solar midday) and circadian time (eg, the phase of a rhythm-marker of the circadian clock) is relatively stable throughout the week. Because the sun’s position in the sky remains unaffected during weekend-weekday shifts of rise- and bedtimes, the sleep-wake cycle becomes unlocked with the circadian clocks on weekdays to return under the control of these clocks on free days. Consequently, Wittmann et al5 and Roenneberg et al7,8 postulated that sleep timing on free days reflects the phase of entrainment of the circadian clock to sun time and it is not affected by early risetimes on the preceding weekdays.
Although the circadian clocks phase under the unchanged 24-h pattern of light exposure cannot be changed by an advancing shift in risetime,4 such shifts can inevitably cause shifts in this pattern because people living in modern post-industrial societies have access to artificial light sources at any time of the day. Therefore, an alternative explanation suggests that although the sun’s position in the sky remains unaffected during weekend-weekday shifts in sleep times, social time can influence sleep timing by inducing shifts in the 24-h pattern of light exposure, which, in turn, leads to shifts in the circadian phase and sleep timing.9,10 Consequently, the circadian sleep timing can be adjusted to earlier school/work schedule through the changes in light self-exposure leading to the changes of the circadian clocks phase.6,9,10
It is of practical importance to answer to the question of which of these two alternative explanations can be supported by data on sleep duration and timing in real-life situations. The first of these two alternative explanations postulates the possibility of temporal disorganization of the mechanism of the circadian adjustment to early weekday wakeups and remarkable loss of sleep on weekdays,4,6,7 while the second explanation postulates the ability of the circadian and homeostatic regulators of the sleep-wake cycle to rapidly adapt to early weekday wakeups and, therefore, to avoid sleep loss due to the compensating advance of the circadian sleep timing.5,9,10
In the present study, we tried to find out whether the internal body clocks can adjust to early weekday wakeups, or their response to these wakeups leads to the circadian misalignment and sleep loss. First, we showed in in silico study (ie, a study that is using a computational model rather than empirical data) that the remarkable differences in predictions based on the two alternative explanations can be revealed by applying a model of sleep-wake regulation for description of the response of circadian and homeostatic regulators to earlier and later weekday wakeups. Second, we demonstrated that these predictions can be fully or partially supported by a comparison of sleep times reported by earlier and later weekday risers in real-life situations. Recently, we proposed a methodology for estimating the contribution of social time to the difference in weekend sleep timing on the example of sleep times reported by 4940 university students and lecturers.11 Therefore, in the present in silico study, we applied one of the versions of the two processes model of sleep-wake regulation12 to highlight the difference in predictions based on the two alternative explanations of the response of the sleep-wake cycle to early wakeups. In the following empirical study, we determined which of the predictions came true by applying the currently proposed methodology for estimation of the contribution of social time to the shifts of weekend sleep timing11 to the analysis of three datasets of sleep times reported by earlier and later weekday risers from 13 age subsets.
The following hypotheses concerning the contribution of sun time and social time to sleep timing were tested by comparing the predictions based on the results of the in silico study with the results of analysis of reported sleep times:
Hypothesis 1. Sun time was the major contributor. Hypothesis 2. Social time was the major contributor. Hypothesis 3. Both sun time and social time were substantial contributors.
Materials and Methods
Table 1 illustrates which sleep times can be additional calculated using bed- and risetimes reported on weekdays and weekends by earlier and later weekday risers. Moreover, the notes in this table explain our recently proposed methodology for estimating the contribution of social time to sleep timing.11 In particular, Table 1 illustrates that the difference between earlier and later risers in social time (measured as the difference in weekday risetime) is equal to the sum of differences between these risers in sleep loss on weekdays (measured as the difference in weekend-weekday gap in risetime) and sleep phase shift on weekends (measured as the difference in weekend risetime).
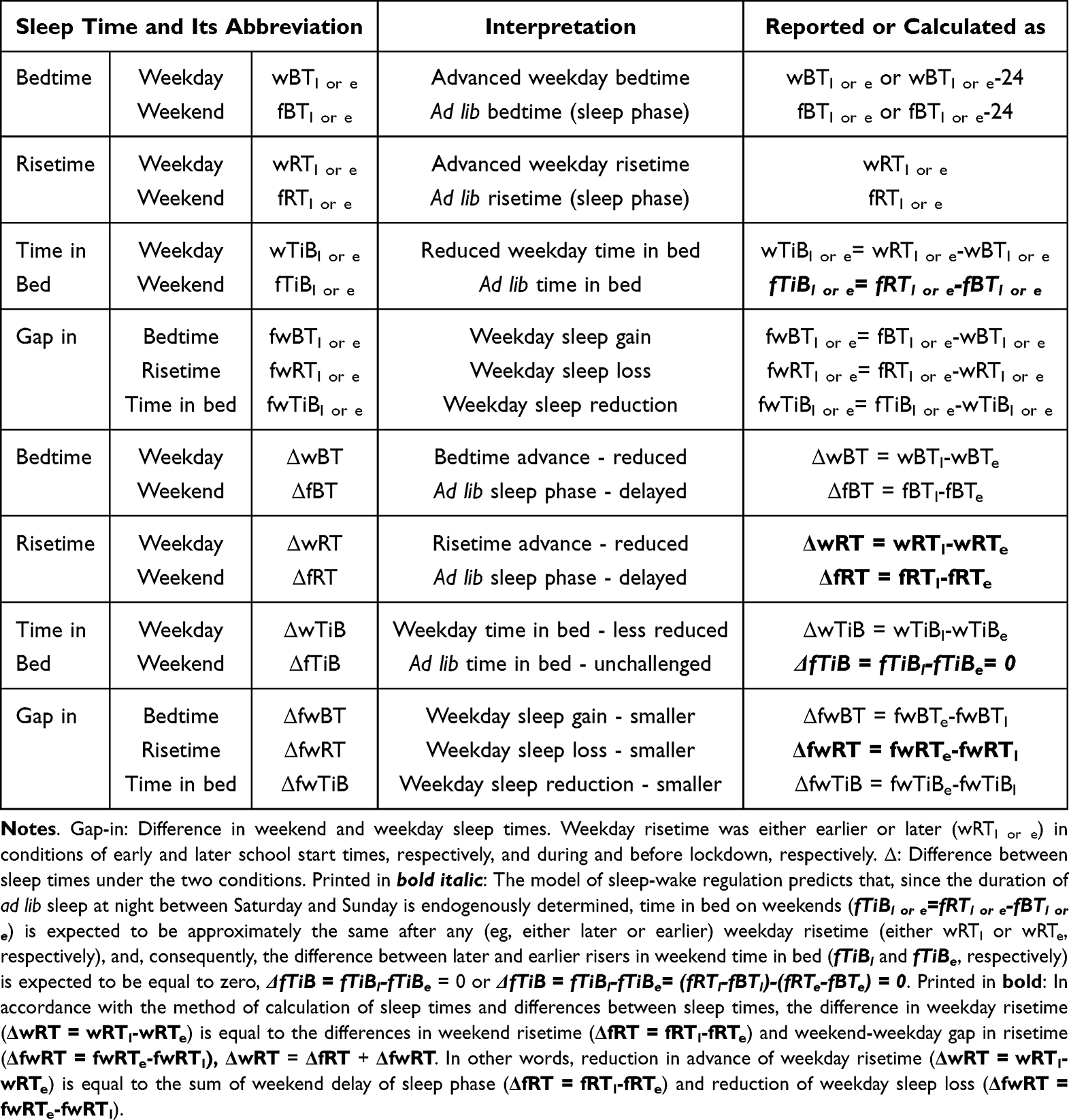 |
Table 1 Differences in Sleep Times Reported by Earlier and Later Weekday Risers |
Our in silico study was designed to demonstrate how the proposed methodology provides the possibility to reveal the difference in predictions based on the alternative explanations of the major role of either sun time or social time in circadian sleep timing. To highlight such differences in predictions based on these explanations, one of the variants of the two-process model of sleep-wake regulation12 was applied. This variant proposes a modulation of all parameters of the homeostatic process described in the classical variant of the two-process model2 by the circadian clocks, that is, the circadian process in the classical variant of the model.2 In the present in silico calculations and previous simulations,12–15 this modulation was incorporated into the model as a simple periodic (sine) function with circadian period. If 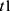 and
and 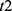 are the initial times for the buildup and decay phases of this process of sleep-wake regulation
are the initial times for the buildup and decay phases of this process of sleep-wake regulation 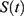 , the mechanism of the circadian and homeostatic regulation of the sleep-wake cycle can be described as follows:
, the mechanism of the circadian and homeostatic regulation of the sleep-wake cycle can be described as follows:
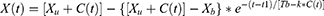
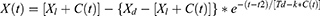
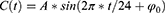
is a sine function with a circadian period. Thus, function (2) accounts for the modulation of homeostatic processes by the circadian clocks. In the present calculations and previous simulations,12–15 this period was assigned to 24.0 h because it is reasonable to assume that the circadian clocks in daytime learner/workers always remain under control of (ie, are entrained to) the external light-dark cycle with the 24.0-h period.
The parameters of this model were initially derived by Putilov12 from data on 1) the duration of recovery sleep after six gradually increasing intervals of extended wakefulness16 and 2) the levels of SWA in 10 naps17 and two recovery sleep episodes.18,19 The simulation of such experimental data provides the possibility of using measurements of relative slow-wave activity (rSWA) for calculations of the time course of the process of sleep-wake regulation, 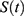 (1,2). These initial parameters were slightly modified in previously performed simulations13–15 and in the present calculations (Table 2, Figures 1 and 2) to account for the difference between the initially simulated experimental sleep durations16 and times in bed calculated from self-reported bed- and risetimes on weekdays and weekends.13–15
(1,2). These initial parameters were slightly modified in previously performed simulations13–15 and in the present calculations (Table 2, Figures 1 and 2) to account for the difference between the initially simulated experimental sleep durations16 and times in bed calculated from self-reported bed- and risetimes on weekdays and weekends.13–15
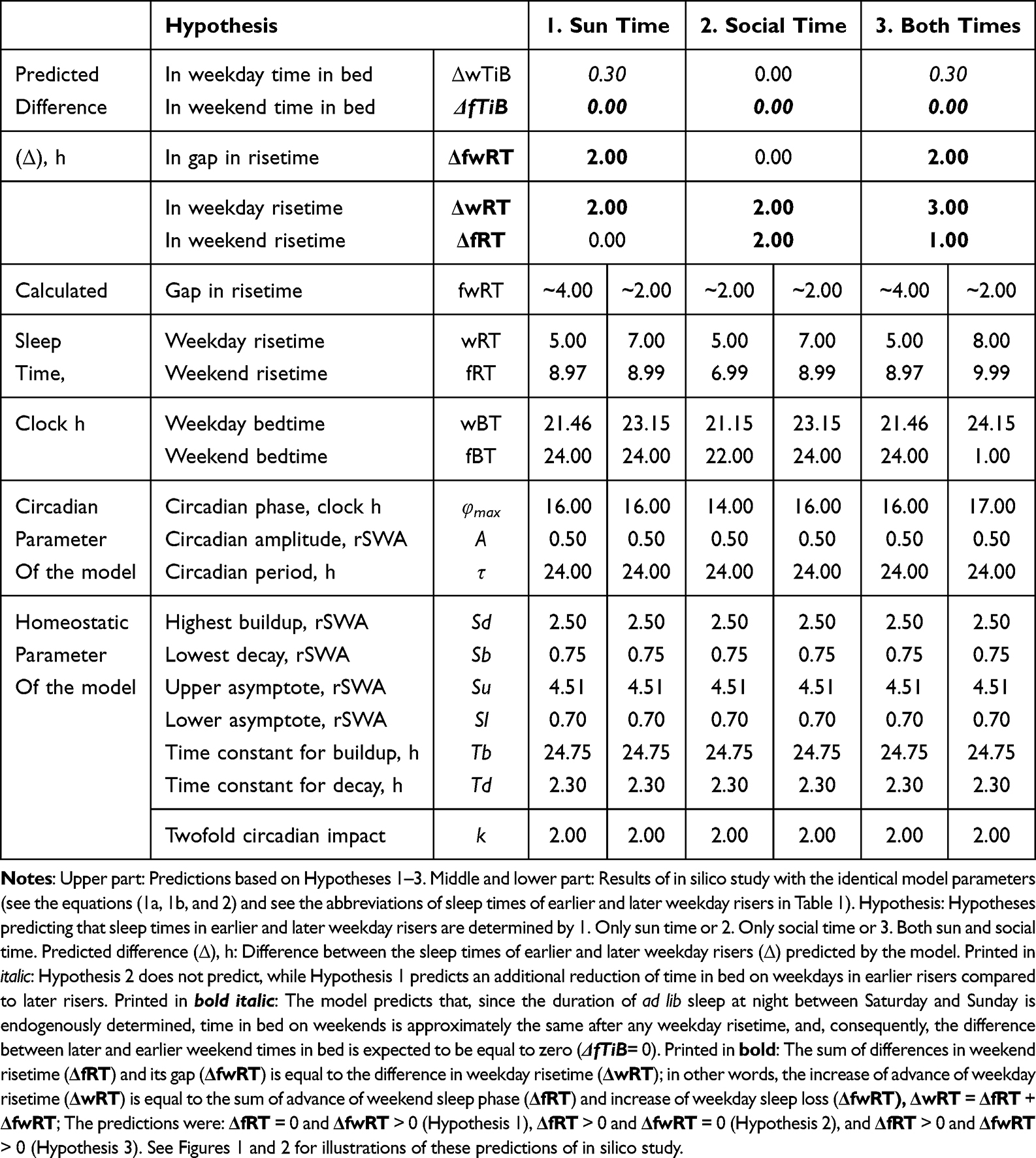 |
Table 2 Comparison of Predictions of the Model Based on Three Hypotheses |
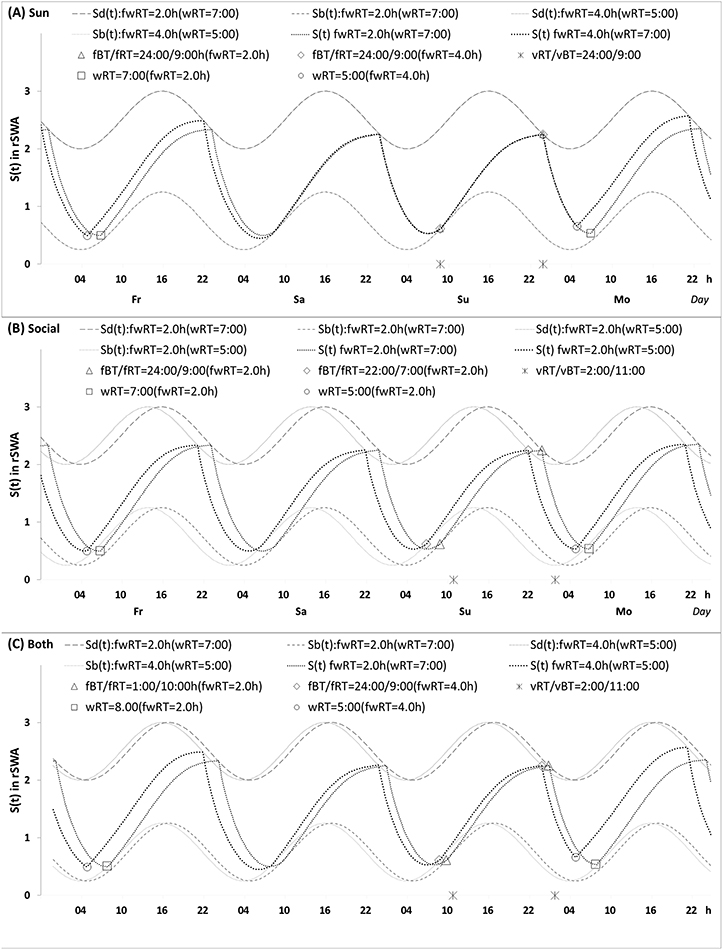 |
Figure 1 Predictions of three hypotheses for four sleep sleep-wake cycles. Two (wake and sleep) phases of the process of sleep-wake regulation |
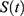
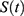
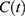
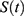
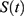
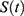
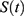
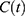
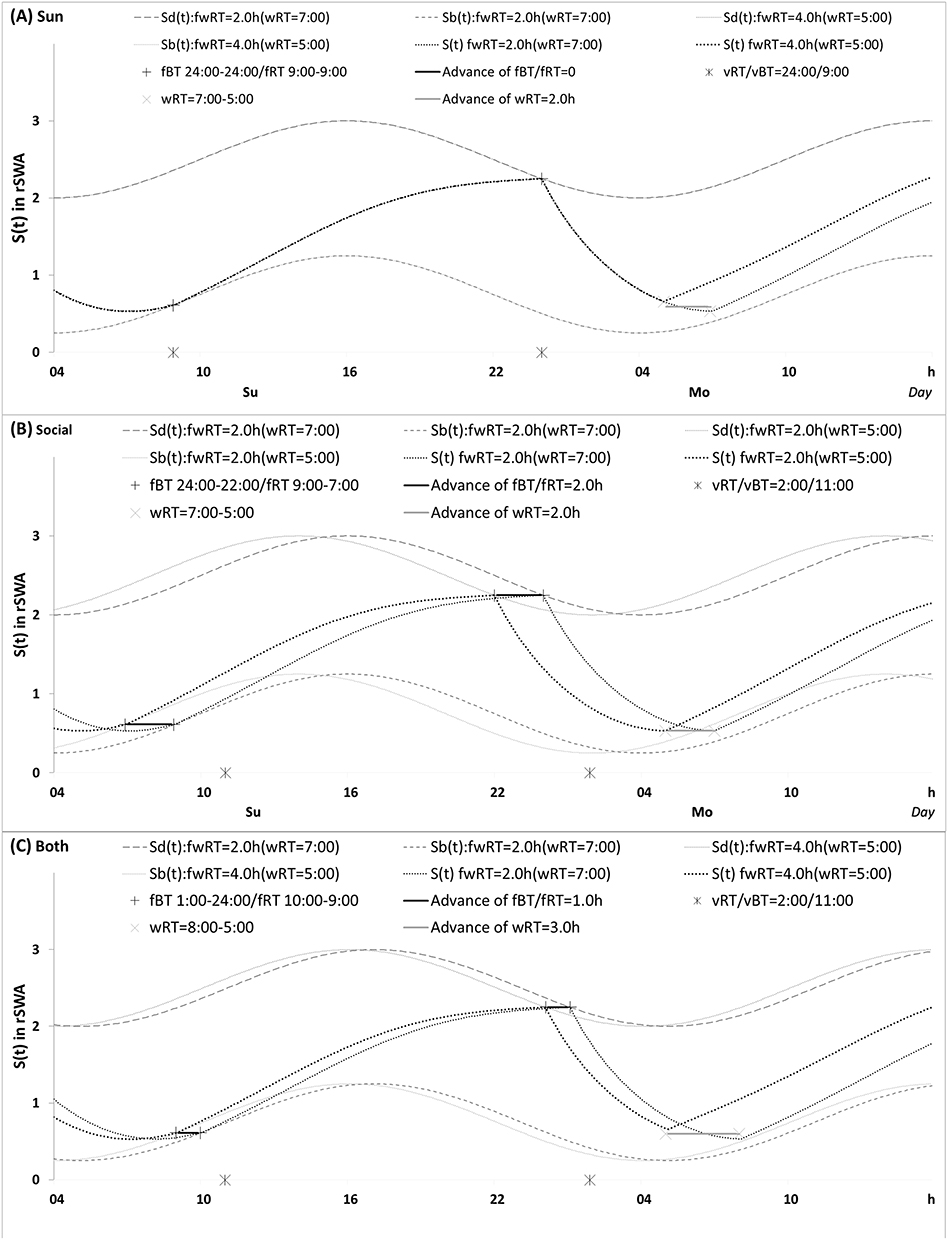 |
Figure 2 Predictions of three hypotheses for two sleep sleep-wake cycles. Only two of the four sleep cycles (time courses of S(t) for Sunday-Monday, So-Mo) from Figure 1 illustrate the differences among the predictions of the three hypotheses. (A) Zero advance of weekend sleep times (fRT and fBT) despite the 2.0-h advance of weekday risetime (wRT). (B) The advance of weekend sleep times (fRT and fBT) is equal to the 2.0-h advance of weekday risetime (wRT). (C) Advance of weekend sleep times (fRT and fBT) is equal to 1.0 h because it is equal to the 3.0-h difference in advance of weekday risetime (wRT) minus the 2.0-h difference in gap between weekend and weekday in risetime (fwRT). See the model’s parameters in Table 2. |
To illustrate the difference in sleep time predicted on the basis of three hypotheses, we calculated hypothetical weekday and weekend sleep cycles using model parameters (Table 2, Figures 1 and 2) resembling the parameters obtained in previous simulations of bed- and risetimes on weekdays and weekends.15 Weekend time in bed was rounded to 9.00 h, and weekend bedtime was set at midnight (Table 2). The following predictions based on three hypotheses were tested by comparing these predictions with the results of analysis of empirical data:
Predictions based on Hypothesis 1. Despite the weekend-weekday changes in risetime (ie, social time), the shifts in the circadian phase of weekend sleep are not expected due to the absence of changes in the sun’s position in the sky (ie, sun time).
Predictions based on Hypothesis 2. Despite the absence of changes in the sun’s position in the sky (ie, sun time), these shifts in the circadian phase of weekend sleep are expected and these shifts are equal to the weekend-weekday changes in risetime (ie, social time).
Predictions based on Hypothesis 3 combining Hypotheses 1 and 2. The shifts in the circadian phase of weekend sleep are expected after changes in risetime (ie, social time), but these shifts are smaller than these changes in risetime.
To test the plausibility of each of three hypothesis, we compared the predictions based on each hypothesis with the results of analysis of three sets of samples with reported sample-averaged bed- and risetimes on weekdays and weekends (Table 3). The whole dataset consisting of 2442 samples and subsamples with sleep times on weekdays and weekends (see Supplementary Tables 1–3) was obtained by extension of the dataset previously collected from the literature and used for model-based simulations of sleep times in several previous studies.13–15 The publications13–15 include more details on the process of the collection of sleep times from the literature for this whole dataset of unpaired samples/subsamples and two datasets of paired samples/subsamples. The content of journal papers with more or less relevant titles was inspected to determine whether weekday and weekend sleep times were reported (usually in a table). In the case of publication of sleep times for both controls and patients or for both baseline and experimental conditions, only a subsample with control (healthy) study participants or a subsample for control condition, respectively, was included. There were no other exclusion criteria for including a sample/subsample in the dataset. Both sleep times reported for the whole sample and for its two subsamples (three in total) were included when sleep times were reported separately for male and female study participants and for morning and evening types. Moreover, sleep times reported only for subsamples were included when these sleep times were reported for early and later school start times (paired subsamples), before and during lockdown (paired subsamples), and different ages (unpaired subsamples). For the present analysis, we did not collect any information on such additional characteristics of samples/subsamples as heterogeneity of age, season, timing of indoor and outdoor light exposure, using artificial lighting devices, cultural and social contexts, sleep and data quality, etc. Additional information in Supplementary Tables 1–3 is limited to the country of data collection and authors and year of paper publication.
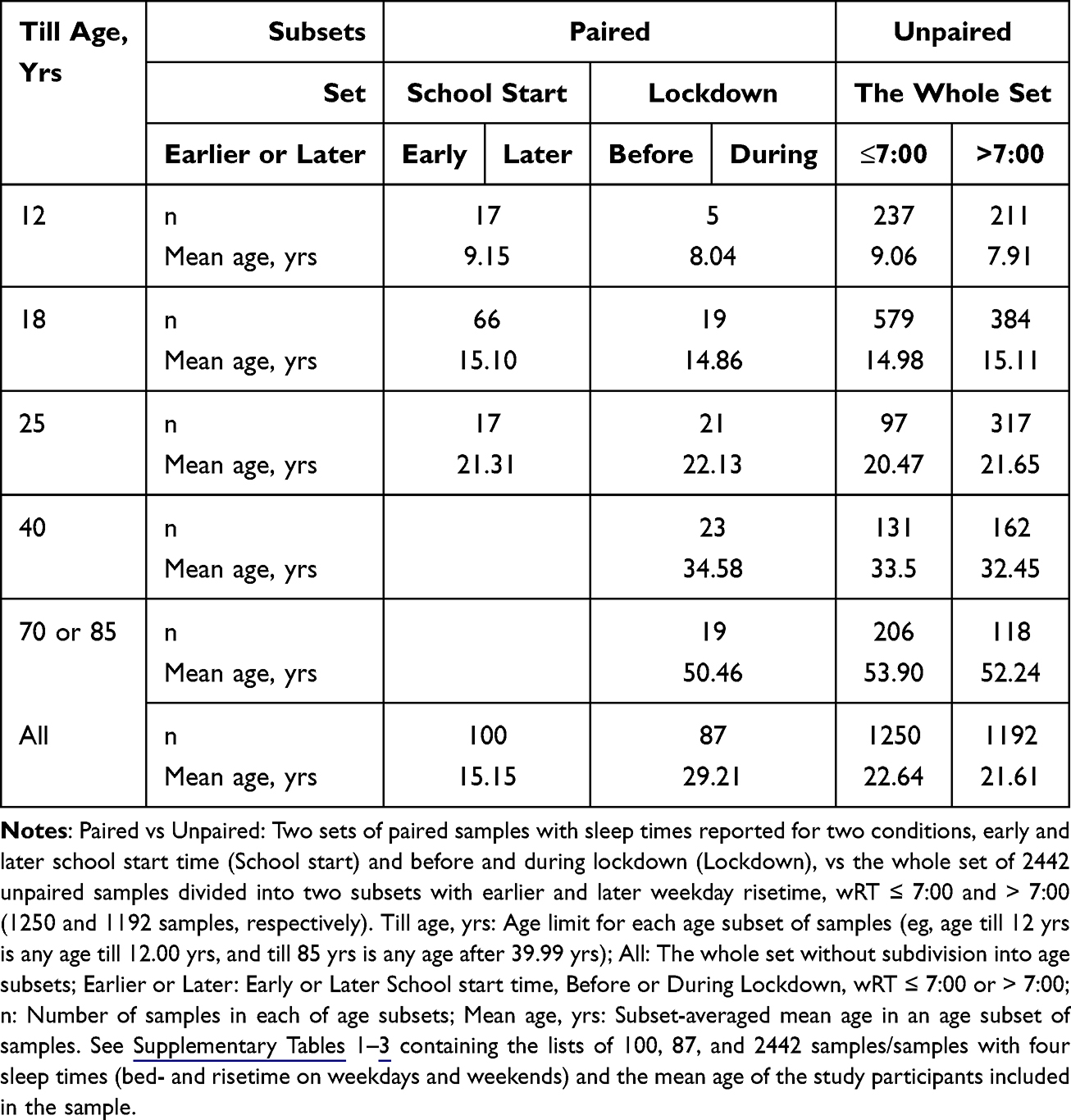 |
Table 3 Subdivision of Three Sets of Samples with Reported Sleep Times into 13 Age Subsets |
To demonstrate that the results of testing the study hypotheses are replicable in any age group, mean age reported for a sample/subsample was used to subdivide each of three sets of samples/subsamples into age subsets. The number of samples/subsamples per each of these age subsets is given in Table 3. In total, two sets of 100 and 87 paired samples were included in the subsets with sleep times before vs during lockdown and during early vs later school start time (five and three age subsets, respectively). The whole set included 1250 vs 1192 unpaired samples with weekday risetime not earlier vs later than 7 a.m. (five age subsets).
The SPSS26.0 statistical software package (IBM, Armonk, NY, USA) was applied for statistical analyses of the differences between earlier and later risers included in 13 age subsets. Paired t-tests were used to analyze the differences between paired samples of earlier and later risers (Table 4). One-way ANOVAs with the independent factor “Weekday risetime” (either not later than or later than 7:00) were run to test the significance of this factor and its size effect (Table 4, left columns).
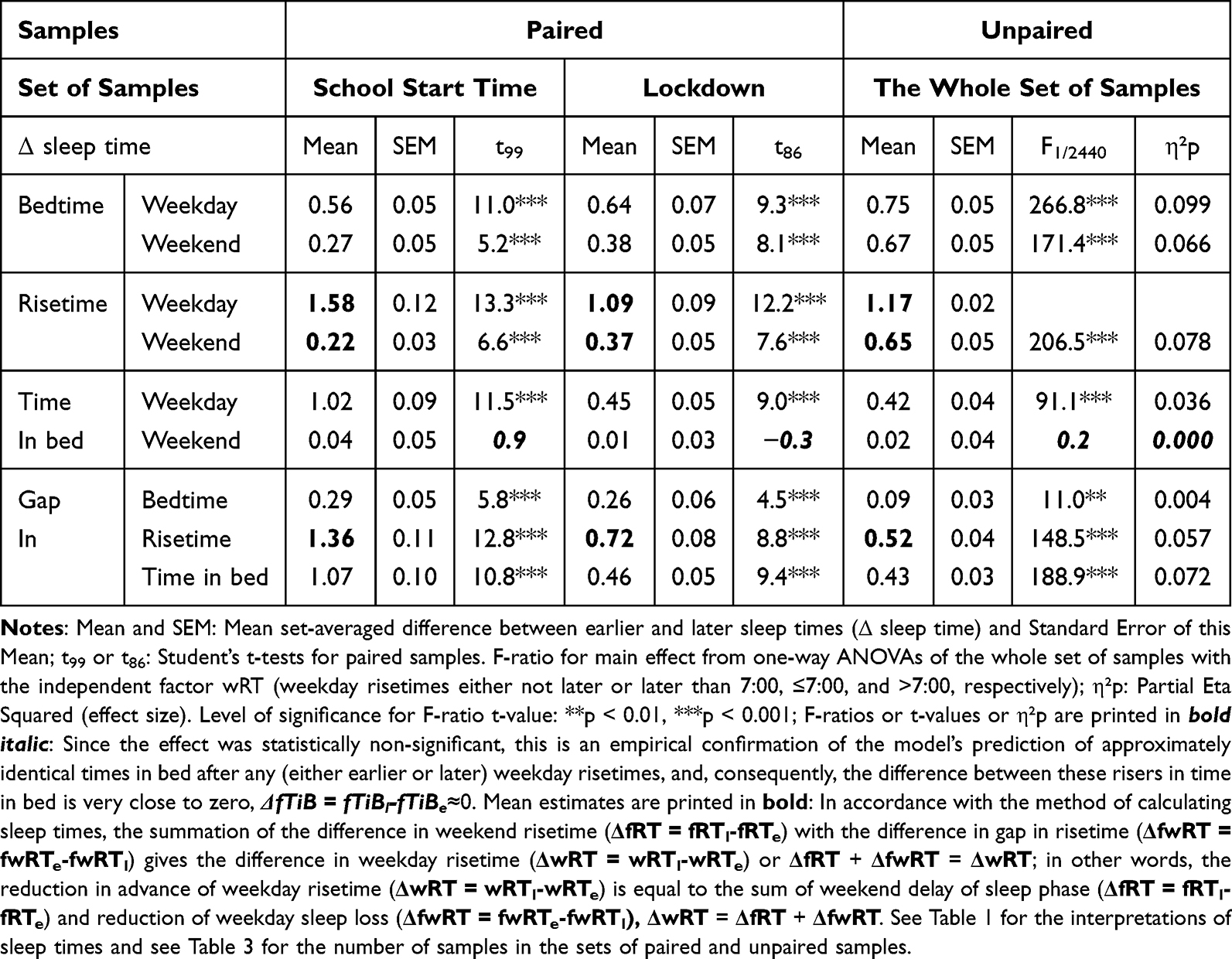 |
Table 4 Paired t-Test for Paired Samples and Main Effect of Weekday Risetime for Unpaired Samples |
The comparison of the model-based predictions with reported sleep times included the testing whether significant weekday sleep shortening and/or significant weekend sleep phase advancing predicted by the hypotheses 1–3 occurred in natural settings as the response to weekday risetime advancing. As shown in Table 2 (upper part), Hypothesis 1 predicts a zero difference between earlier and later weekday risers in weekend sleep timing (as a result, 0 + weekday sleep shortening = weekday risetime advancing). In contrast, Hypothesis 2 predicts a zero difference between these risers in weekday sleep shortening (as a result, weekend sleep phase advancing + 0 = weekday risetime advancing). Hypothesis 3 predicts a non-zero difference between these risers in both weekend sleep timing and weekday sleep shortening (as a result, weekday sleep shortening + weekend risetimes advancing = weekday risetimes advancing; Table 2, upper part). The additional comparisons included the testing of predictions of 1) a zero difference between earlier and later risers in weekday time in bed (the 2nd, but not the 1st and 3rd hypotheses) and 2) a zero difference between these risers in weekend time in bed (any of the hypotheses; Table 2, upper part).
Results
Figure 3 illustrates sleep times calculated for 13 age subsets of earlier and later weekday risers, and Figure 4 illustrates the difference between earlier and later risers in these sleep times. In particular, it is shown in Figure 4 that the difference between earlier and later risers in weekday risetime (social time, ΔwRT) is equal to the sum of differences between these risers in sleep loss on weekdays (measured as the difference in weekend-weekday gap in risetime, ΔfwRT) and sleep phase shift on weekends (measured as the difference in weekend risetime, ΔfRT).
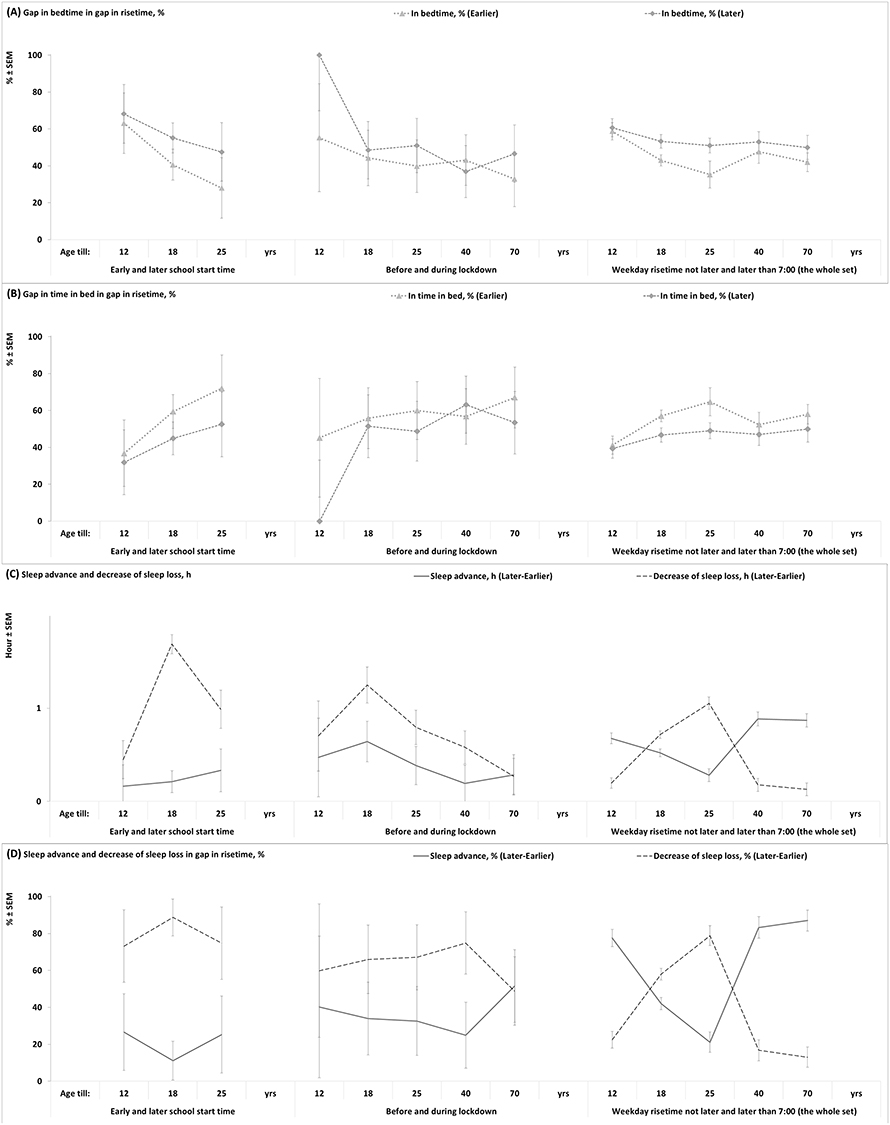 |
Figure 4 Delay of weekend sleep phase as a consequence of delay of weekday risetime. (A and B) The weekend-weekday gap in risetime is equal to the sum of weekend-weekday gaps in bedtime and time in bed, as illustrated by the percentages of these gaps in a one-hour gap in risetime. Gaps in time in bed, risetime, and bedtime were interpreted as sleep reduction, sleep loss, and sleep gain, respectively (Table 2). When the time in bed is shorter on weekends than on weekdays (wTiBe or wTiBl < fTiBe or fTiBl, respectively), the reduction in time in bed on weekdays (fwTiBe and fwTiBl, respectively) is a consequence of a larger weekday sleep loss (fwRTl or fwRTe) compared to weekday sleep gain (fwBTl and fwBTe, respectively), that is, fwTiBl= fwRTl-wBTl or fwTiBe= fwRTe-wBTe, respectively (Table 2). In accord with the model-based interpretation of these gaps, the delay of weekday risetime on one h (ΔwRT = wRTl-wRTe= 1.0 h), leads to less than one-h decrease of sleep gain (ΔfwBT = fwBTe-fwBTl< 1 but always >0) and less than one-h decrease in sleep reduction (ΔfwTiB = fwTiBe-fwTiBl< 1 but always >0), ie, ΔwRT = ΔfwBT + ΔfwTiB = 1.0 h. (C) The difference between later and earlier weekend risetimes (ΔwRT = wRTl-wRTe) is equal to the sum of differences in weekday sleep loss (ΔfwRT = fwRTl-fwRTe) and weekend advance of sleep phase (ΔfRT = fRTl-fRTe). (D) Consequently, the one-h difference between later and earlier weekend risetimes (ΔwRT = wRTl-wRTe= 1.0 h) leads to less than one-h decrease of sleep loss (ΔfwRT = fwRTl-fwRTe< 1.0 h but always >0) and less than one-h advance of sleep phase (ΔfRT = fRTl-fRTe< 1.0 h but always >0), because their sum is equal to one h (ΔwRT = ΔfwRT + ΔfRT = 1.0 h). |
As shown in Table 2 (upper part), the prediction based on Hypothesis 1 suggests a zero difference between earlier and later weekday risers in weekend sleep timing. Therefore, ΔwRT = ΔfRT + ΔfwRT = 0 + 2.0 = 2.0 h, ie, the 2.0-h difference in weekday risetime (ie, social time, ΔwRT) is explained exclusively by the 2.0-h difference in gap between weekend and weekday risetime (ΔfwRT) in Figures 1A and 2A, while ΔfRT = 0. The earlier risers slept 2.0 h less on weekdays than later risers because the circadian phase of sleep was identical in these risers due to the same position of the sun in the sky (the sun time; Table 2). Instead, the results on reported sleep times shown in Table 4, Figures 3 and 4 suggested that ΔfRT > 0. Therefore, the prediction of Hypothesis 1 was not supported.
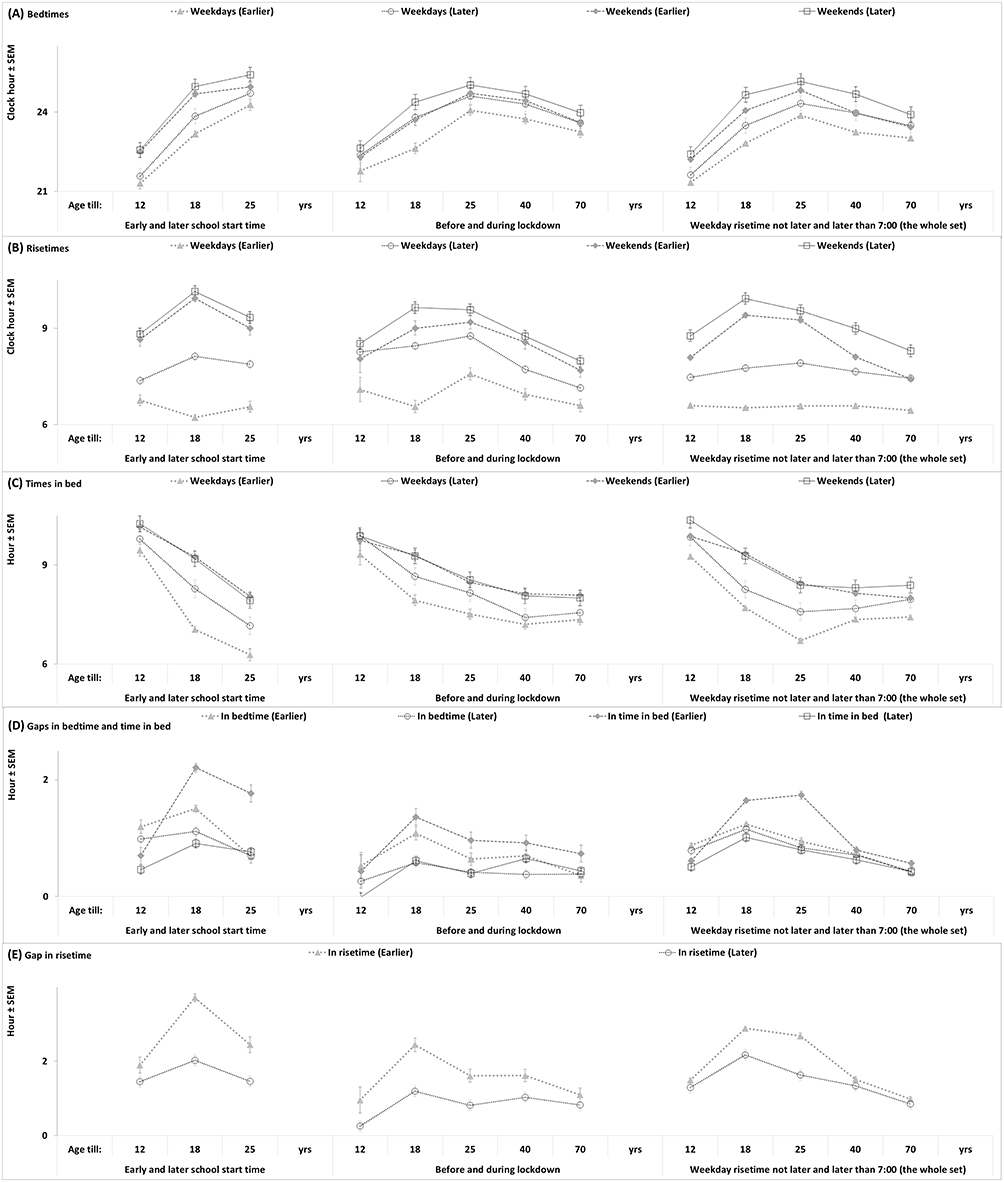 |
Figure 3 Sleep times in age subsets of paired and unpaired samples. (A) Bedtimes on weekdays and weekends. (B) Risetimes on weekdays and weekends. (C) Times in bed on weekdays and weekends. (D) Weekend-weekday gaps in bedtimes and times in bed. (E) Weekend-weekday gap in risetimes. |
In contrast, the prediction based on Hypothesis 2 suggests a zero difference between the earlier and later weekday risers in weekday sleep loss, ΔwRT = ΔfRT + ΔfwRT = 2.0 + 0 = 2.0 h, ie, the 2.0-h difference in weekday risetime (ie, social time, ΔwRT) is explained exclusively in Figures 1B and 2B by the 2.0-h difference in weekend risetime (ΔfRT), while ΔfwRT = 0. The duration of sleep on weekdays was identical in the earlier and later risers because the 2.0-h shift in the circadian phase of sleep compensated for the 2.0-h shift in weekday risetime (Table 2). Instead, the results on reported sleep times shown in Table 4, Figures 3 and 4 suggested that, ΔfRT > 0. Therefore, the prediction of Hypothesis 2 was not supported.
The predictions based on Hypothesis 3 combine the predictions of two previous hypotheses. Instead of predicting a zero difference between the earlier and later weekday risers in weekday risetime (ΔfRT) or the gap in risetime (ΔfwRT), the predictions suggest non-zero differences in weekday sleep loss and non-zero differences in weekend sleep timing, ΔwRT = ΔfRT + ΔfwRT = 1.0 + 2.0 = 3.0 h, in Figures 1C and 2C. Thus, the predictions explain the 3.0-h difference in weekday risetime (ie, social time, ΔwRT) by the 2.0-h difference in weekday sleep loss (ΔfwRT) combined with the 1.0-h difference in weekend risetime (ΔfRT). The circadian phase of sleep is expected to be shifted ahead in earlier risers relative to that of later risers, thus partly compensating for the 3.0-h shift in weekday risetime (Figures 1C and 2C). The earlier risers slept 2.0 h less on weekdays than later risers despite the more prominent (3.0-h) shift of weekday risetime, ΔwRT (Figures 1C and 2C). Thus, these predictions suggest that the same position of the sun in the sky above the heads of the earlier and later risers prevents a larger advancing shift of sleep timing. This leads to a more profound weekday sleep loss in earlier risers than in later risers, ΔfwRT = ΔwRT-ΔfRT = 3.0–1.0 = 2.0 h (Table 2). The results on reported sleep times (Table 4) suggested that, indeed, ΔfRT > 0 and ΔfwRT > 0. Therefore, the predictions of Hypothesis 1 were supported. In each of 13 age subsets, the difference in weekend risetime and the difference in gap between weekends and weekdays in risetime contributed to the difference in weekday risetime, ΔfwRT = ΔwRT-ΔfRT. The advance of weekend sleep time (ΔfRT) and the increase in weekday sleep loss (ΔfwRT) in each age subset was larger than zero (Figure 4). As shown in Table 4, the results of analyses of three sets of samples indicated that the contributions of both ΔfRT and ΔfwRT to ΔwRT were statistically significant. Therefore, as suggested by Hypothesis 3, both social time and sun time significantly contributed to weekend sleep timing in any of three sets of samples.
Besides, the model predicts, irrespective of the hypothesis, a zero difference between the earlier and later risers in time in bed on weekends. Given that the duration of ad lib sleep at night between Saturday and Sunday is endogenously determined, time in bed on weekends (fTiB) is expected to be approximately the same after any (eg, either or later) weekday risetime (Table 1 and Table 2). As shown in Table 4, the statistical results supported this prediction of the model. They suggested that the difference between the earlier and later risers in time in bed on weekends was non-significant in any of three sets of samples (Table 4 and Figure 4). If this prediction was not supported by the empirical data (ΔfTiB <> 0), the difference between earlier and later risers in weekday risetime (social time, ΔwRT) cannot be accurately represented by the sum of differences between these risers in sleep loss on weekdays (measured as the difference in weekend-weekday gap in risetime, ΔfwRT) and sleep phase shift on weekends (measured as the difference in weekend risetime, ΔfRT).
In practical terms, the difference between the shift in weekday risetime (social time) and the shift in weekend risetime (circadian phase of sleep) suggested that the shift in social time increases sleep loss on weekdays (measured as the gap between weekend and weekday risetime, ΔfwRT). Figure 5 illustrates the profound differences between earlier and later risers in sleep loss and advance in weekend sleep timing. Table 5 illustrates the negative consequences of earlier risetimes for sleep duration and timing in 13 analyzed age subsets. Compared to later risers, earlier risers were more frequently included in samples characterized by sleep insufficiency (averaged time in bed < 6.0 h), sleep curtailment (averaged weekend-weekday gap in risetime > 3.0 h), and sleep displacement (in average, less than 80% overlap between weekend and weekday sleep). Such drastic difference is not predicted by Hypothesis 2, which suggests that sleep timing can be adjusted to an earlier school/work schedule through changes in light self-exposure, leading to changes in the circadian phase of sleep. However, the difference would be even larger in accord with the prediction of Hypothesis 1. In support to the prediction of Hypothesis 3, Table 4 highlights that an advance in sleep timing partly compensated this sleep loss. Namely, the additional sleep loss was equal to 1.36 h after 1.58-h advance of weekday risetime during early school start time relative to later time, because 0.22 h were compensated by advance of sleep timing. The additional sleep loss was equal to 0.72 h after 1.09-h advance of weekday risetime before lockdown relative to the period of lockdown due to the 0.37-h compensating advance of sleep timing. The additional sleep loss was equal to 0.52 h after 1.17-h advance of weekday risetime relative to later weekday risetime (after 7:00) due to the 0.67-h compensating advance of sleep timing.
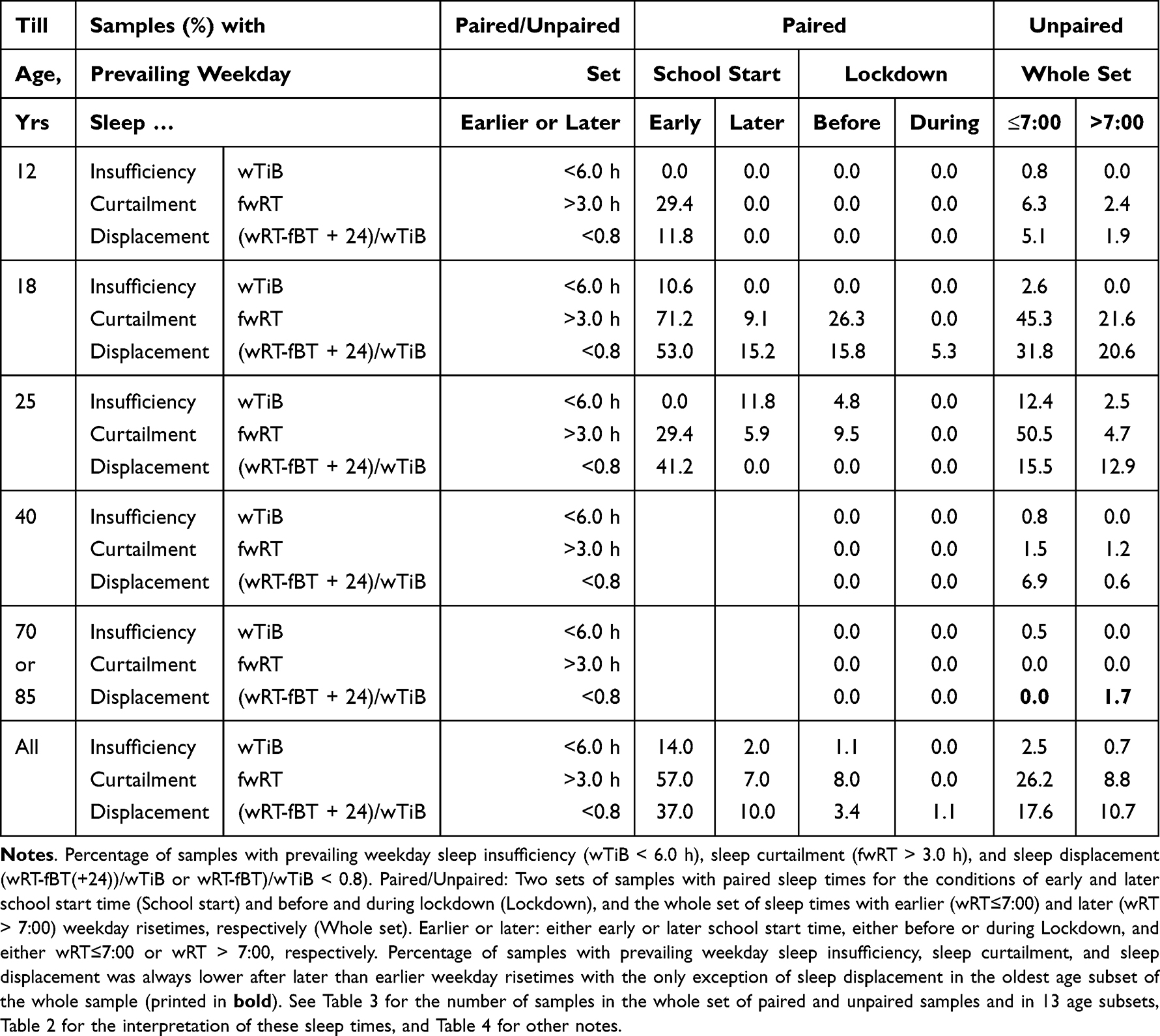 |
Table 5 Prevalence of Weekday Sleep Insufficiency, Curtailment, and Displacement |
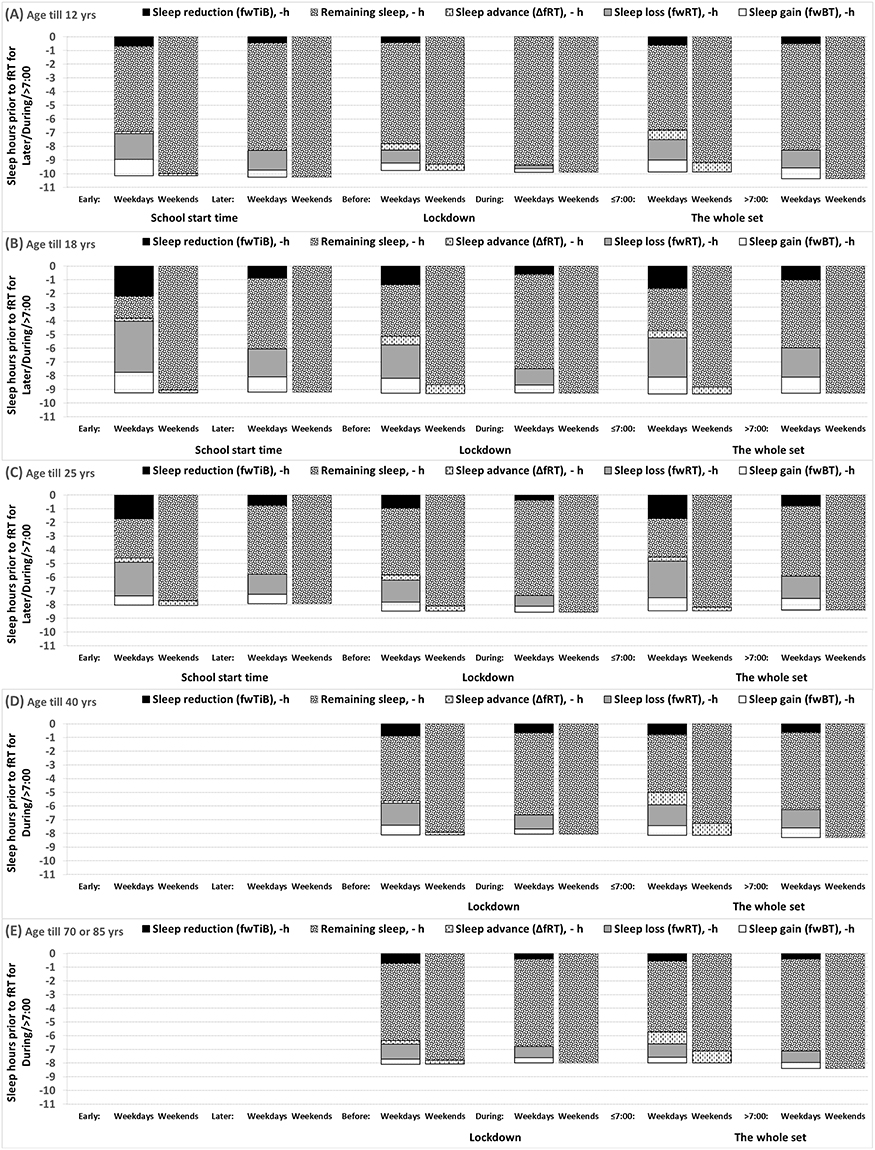 |
Figure 5 Fractions of time in bed in 13 age subsets of paired and unpaired samples. (A–E) Comparison of fractions of time in bed for age subsets shown in Figure 1. The fractions are shown relative to weekend risetime after later weekday risetime (wRTl). Given a model prediction of the almost identical risetime after ad lib sleep following any (earlier on weekdays and later on weekends) weekday bedtime (wRTl or e), the weekend-weekday gaps in risetime (fwRTl or e), bedtime (fwBTl or e), and in time in bed (fwTiBl or e) were termed “sleep loss”, “sleep gain”, and “sleep reduction”, respectively, fwTiBl or e = fwRTl or e - fwBTl or e (Table 2). The gap in risetime (fwRTl or e) rather than the gap in time in bed (fwTiBl or e) was named “sleep loss” because the model predicts that, despite earlier bedtime on weekdays compared to weekends, ad lib sleep after weekday bedtime (eg, between Friday and Saturday) is terminated at the same time point as ad lib sleep after weekend bedtime (ie, the reason for this is the circadian influence on sleep duration that is longer when sleep starts at an earlier circadian phase compared to sleep started at later circadian phase, eg, on weekday and weekend, respectively). In accord with a model’s prediction supported by the results of analysis of reported sleep times (see Table 4), the difference between weekend time in bed (fTiBl or e) after any (earlier or later) weekday risetime (wRTl or e) is almost identical, and the difference between these wRTl and wRTe (ΔwRT = wRTl-wRTe) is the sum of the difference in sleep loss (ΔfwRT = fwRTe-fwRTl) and the delay of weekend sleep phase (ΔfRT = fRTl-fRTe). In other words, after a shift of weekday risetime from wRTe to wRTl, sleep loss is decreased but the decrease is smaller than ΔwRT = wRTl-wRTe, because weekend sleep phase is delayed (ΔfRT = fRTl-fRTe), and, consequently, ΔfwRT = ΔwRT - ΔfRT (Table 2). |
Discussion
There are conflicting views on the ability of circadian clocks to adjust sleep timing to early wakeups on weekdays. In the present in silico study, we applied the sleep-wake regulation model to demonstrate the difference between sleep times of earlier and later weekday risers predicted by two alternative explanations of the roles of sun time or social time in the light-induced entrainment of the circadian sleep phase. To test the predictions of the model, we analyzed the sleep times reported for paired samples from the studies of sleep before and during lockdown and early and later school start time, and for unpaired samples with weekday risetimes not earlier and later than 7 a.m. We calculated the difference between earlier and later risers in weekday risetime (social time) that is equal to the sum of differences between these risers in weekday sleep loss and weekend sleep phase measured as the difference in the weekend-weekday gap in risetime and the difference in weekend risetime, respectively. The results of comparison of predicted and reported sleep times support the combined explanation that both sun time and social time contribute to light-induced entrainment of the circadian phase and sleep timing.
Namely, the difference between earlier and later weekday risers was explained in any of 13 analyzed age subsets by the non-zero difference in sleep loss and the non-zero difference in sleep phase shift. Therefore, the shift in social time leads to a shift in the 24-h pattern of exposure to artificial and natural light sources, thus causing a shift in circadian and sleep phases. However, this shift was insufficient to fully compensate for differences between earlier and later weekday in sleep loss. Therefore, earlier risers were more frequently than later risers included in samples characterized by sleep insufficiency, sleep curtailment, and sleep displacement.
Paradoxically, although the artificial light sources can be blamed for the delay of circadian phase observed in people living in modern postindustrial societies,20,21 they, on the other hand, can help to partially reduce this delay by advancing the 24-h pattern of light exposure after earlier weekday wakeups. Experimental human studies have confirmed the substantial influence of wakeup time, likely via associated morning light exposure, on the timing of the human circadian clock.22,23 The findings of these experimental studies were additionally supported by the results of natural experiments on weekday shifts of risetimes relative to weekend risetimes.24–28
The results on reported sleep times supported the model-based prediction that, since the duration of ad lib weekend sleep is endogenously determined, the time in bed on weekends is approximately the same after any (eg, either later or earlier) weekday risetime. The present study has practical implications because the confirmation of this prediction of the model allows the quantitative estimation of the severity of weekday sleep loss. Using sleep times in 13 age subsets as an example of such estimation, we showed that weekday sleep curtailment was dramatic in many samples. It is well-known that sleep curtailment has non-negligible consequences on a wide range of outcomes, including performance, wellbeing and health. For instance, insufficient sleep is associated with diminished productivity,29,30 reduced cognitive performance,31–34 impaired memory and attention,35–38 increased absenteeism39,40 and risk of accident and work injuries,41–43 mental disorders,44–46 social withdrawal47 and suicide,48,49 development of cancer,50–52 cardiovascular53–56 and metabolic diseases,57–60 compromised immune function,61–63 and, in overall, serious health complications and higher mortality.64–67 Such negative effects of early weekday wakeups are not predicted by Hypothesis 1 suggesting that sleep timing can be adjusted to an earlier school/work schedule through advancing light self-exposure leading to advancing the circadian phase of sleep. Although these negative effects are predicted by Hypothesis 2 suggesting that the natural light-dark cycle is the major stimulus that entrains human circadian rhythms, the effects suggested by the present analysis of reported sleep times are not as strong as predicted by this hypothesis. In agreements with predictions of Hypothesis 3, the shift in the circadian phase of sleep partly compensates the loss of weekday sleep, but it fails to fully prevent this loss after advancing weekday wakeups. Therefore, the previously approved light interventions (eg, 68–70) can be recommended to change the ratio between the two terms of the equation of advancing shift of weekday risetime. Since this shift is equal to the advancing shift of circadian sleep phase and the increase in weekday sleep loss, an appropriately timed exposure to artificial and natural bright light sources can increase the contribution of the advancing shift of circadian sleep phase at the expense of weekday sleep loss. For instance, earlier and more extensive morning light exposure combined with earlier termination and less intensive evening light exposure might be recommended for reduction of weekday sleep loss.
In the present study, we did not evaluate the effect of chronotype on the two components of the response to the advancing shift of weekday risetimes, weekday sleep shortening and weekend sleep phase advancing. However, our previous comparisons of sleep times reported by morning and evening types suggested the following absolute and relative differences in their response. Since the advancing shift of weekday risetimes is larger in evening than morning types in absolute terms, both weekday sleep shortening and weekend sleep phase advancing are lager in absolute terms in evening types.71 However, the comparison of those who, being either morning or evening types, wakeup earlier and later suggested that the responses of these types appear to be similar, ie, they did not differ in the fractions of weekday sleep shortening and weekend sleep phase advancing in response to the advancing shift of weekday risetimes.11
This study had several limitations. The application of a cross-sectional and non-repeated measures design was one of the limitations of the results of analysis of the whole set of samples (ie, an earliest riser and a late weekday riser were not the same person in this dataset). On the other hand, the sizes of two sets of paired (lockdown and school) samples were relatively small. However, since the predictions of Hypothesis 3 were supported by the results of analysis of any of 13 age subsets, the replicability and generalizability of the present results across different ages cannot be questioned. It remains to be elucidated whether this result can be also generalized across different cultures and socioeconomic conditions. Since weekday and weekend bed- and risetimes sleep were self-reported, they may not be accurate and they might not inform about the profound day-to-day variation in sleep timing and duration. Moreover, sleep timing does not indicate the phase positions of markers of the circadian phase. Therefore, it does not inform about the phase angle between the circadian and sleep phases and about possible change of this angle after advancing weekday wakeups. However, the lack of information about the phase angle cannot challenge the conclusions of our study (eg, that the predictions of Hypothesis 3 were reliably supported). The absence of information on socioeconomic status, timing and variability in light exposure, cultural and social contexts, etc. was also among the limitations of the present study. Future studies can be aimed on longitudinal evaluation of objectively measured sleep-wake cyclicity with parallel measurements of the hormonal and physiological indexes of circadian phase position and with accounting for the contribution of multiple external factors including the contribution of the 24-h light-dark cycle to the response to early weekday wakeups.
Conclusion
In the present in silico study, the model of sleep-wake regulation was applied to highlight the difference in predictions based on two alternative explanations of the major contributors to light entrainment of sleep timing, either sun time or social time. Moreover, we applied the previously proposed method of estimation of the shift in sleep phase on weekends and the loss of sleep on weekdays in response to the shift in weekday wakeups (social time). In order to determine which of the predictions came true, three sets of samples were used to provide comparison of the predictions based on the alternative hypotheses with the reported sleep times. The hypotheses proposing the major contribution of either sun time or social time were not supported. This implies that the difference in weekday risetime (social time) was not the only contributor to the light entrainment of the circadian sleep timing. Both social and sun times significantly contributed to this entrainment. Since the shifts in circadian time for sleep were less prominent than the shifts in social time, earlier weekday wakeups were inevitably associated with additional losses of weekday sleep. It seems that the circadian and homeostatic regulators of the sleep-wake cycle cannot provide the full adjustment to early weekday wakeups due to the insufficient advancing shift of the 24-h light-dark cycle. Therefore, special interventions can be recommended to decrease weekday sleep loss at the expense of compensating increase of contribution of advancing shift of circadian and sleep phases.
Abbreviations
Bedtime weekday wBT, weekend fBT; Risetime weekday wRT, weekend fRT; Time in bed weekday wTiB, weekend fTiB; Gap in bedtime fwBT, risetime fwRT, time in bed fwTiB, Difference in: Bedtime weekday ΔwBT, weekend ΔfBT; Risetime weekday ΔwRT, weekend ΔfRT, Time in bed weekday ΔwTiB, weekend ΔfTiB; Gap in bedtime ΔfwBT, risetime ΔfwRT, time in bed ΔfwTiB.
Data Sharing Statement
The datasets generated and analyzed during the current study are included in the Supplementary Tables 1–3.
Statement on the Use of Generative AI
During the preparation of this work, the author(s) did not use AI.
Author Contributions
A.A.P. Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Project administration, Supervision; Validation; Visualization; Writing (original draft); Writing (review and editing). E.G.V.: Methodology; Software; Visualization; Writing (original draft); Writing (review and editing). All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
No funding was received for the current study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pittendrigh CS, Daan S. A functional analysis of circadian pacemakers in nocturnal rodents. IV, entrainment: pacemaker as clock. J Comp Physiol. 1976;106:291–331. doi:10.1007/BF01417859
2. Daan S, Beersma DGM, Borbély AA. Timing of human sleep: recovery process gated by a circadian pacemaker. Am J Physiol Regul Integr Comp Physiol. 1984;246:R161–R178. doi:10.1152/ajpregu.1984.246.2.R161
3. Borbély AA, Daan S, Wirz-Justice A, Deboer T. The two-process model of sleep regulation: a reappraisal. J Sleep Res. 2016;25(2):131–143. doi:10.1111/jsr.12371
4. Czeisler CA, Gooley JJ. Sleep and circadian rhythms in humans. Cold Spring Harb. Symp Quant Biol. 2007;72:579–597. doi:10.1101/sqb.2007.72.064
5. Wittmann M, Dinich J, Merrow M, Roenneberg T. Social jetlag: misalignment of biological and social time. Chronobiol Int. 2006;23:497–509. doi:10.1080/07420520500545979
6. Charoenthammanon RS, Gooley JJ. The adolescent circadian clock entrains to social time rather than sun time. Curr Biol. 2025;35(9):2100–2111.e5. doi:10.1016/j.cub.2025.03.059
7. Roenneberg T, Kumar CJ, Merrow M. The human circadian clock entrains to sun time. Curr Biol. 2007;17(2):R44–5. doi:10.1016/j.cub.2006.12.011
8. Roenneberg T, Merrow M. Entrainment of the human circadian clock. Cold Spring Harb Symp Quant Biol. 2007;72:293–299. doi:10.1101/sqb.2007.72.043
9. Shochat T, Santhi N, Herer P, Flavell SA, Skeldon AC, Dijk DJ. Sleep timing in late autumn and late spring associates with light exposure rather than sun time in college students. Front Neurosci. 2019;13:882. doi:10.3389/fnins.2019.00882
10. Skeldon AC, Dijk DJ. Weekly and seasonal variation in the circadian melatonin rhythm in humans: entrained to local clock time, social time, light exposure or sun time? J Pineal Res. 2021;71(1):e12746. doi:10.1111/jpi.12746
11. Putilov AA, Verevkin EG, Sveshnikov DS, et al. Estimation of sleep shortening and sleep phase advancing in response to advancing risetimes on weekdays. Chronobiol Int. 2025;42(6):770–783. doi:10.1080/07420528.2025.2509629
12. Putilov AA. The timing of sleep modelling: circadian modulation of the homeostatic process. Biol. Rhythm Res. 1995;26(1):1–19. doi:10.1080/09291019509360320
13. Putilov AA. Sleep during “lockdown” highlighted the need to rethink the concept of weekend catch-up sleep. Sleep Breath. 2022;26(4):2001–2007. doi:10.1007/s11325-021-02492-z
14. Putilov AA. Weekend sleep after early and later school start times confirmed a model-predicted failure to catch up sleep missed on weekdays. Sleep Breath. 2023a;27(2):709–719. doi:10.1007/s11325-022-02648-5
15. Putilov AA. Reaction of the endogenous regulatory mechanisms to early weekday wakeups: a review of its popular explanations in light of model-based simulations. Front Netw Physiol. 2023b;3:1285658. doi:10.3389/fnetp.2023.1285658
16. Åkerstedt T, Gillberg M. The circadian variation of experimentally displaced sleep. Sleep. 1981;4:159–169.
17. Dijk D-J, Beersma DGM, Daan S. EEG power density during nap sleep: reflection of an hourglass measuring the duration of prior wakefulness. J Biol Rhythms. 1987;2:207–220. doi:10.1177/074873048700200304
18. Dijk D-J, Brunner DP, Borbély AA. Time course of EEG power density during long sleep in humans. Am J Physiol Regulatory Integrative Comp Physiol. 1990;258:R650–R661. doi:10.1152/ajpregu.1990.258.3.R650
19. Dijk D-J, Brunner DP, Borbély AA. EEG power density during recovery sleep in the morning. Electroencephalogr Clin Neurophysiol. 1991;78:203–214. doi:10.1016/0013-4694(91)90034-2
20. Stothard ER, McHill AW, Depner CM, et al. Circadian entrainment to the natural light-dark cycle across seasons and the weekend. Curr Biol. 2017;27(4):508–513. doi:10.1016/j.cub.2016.12.041
21. Wright KP Jr, McHill AW, Birks BR, Griffin BR, Rusterholz T, Chinoy ED. Entrainment of the human circadian clock to the natural light-dark cycle. Curr Biol. 2013;23(16):1554–1558. doi:10.1016/j.cub.2013.06.039
22. Burgess HJ, Eastman CI. A late wake time phase delays the human dim light melatonin rhythm. Neurosci Lett. 2006;395(3):191–195. doi:10.1016/j.neulet.2005.10.082
23. Crowley SJ, Carskadon MA. Modifications to weekend recovery sleep delay circadian phase in older adolescents. Chronobiol Int. 2010;27(7):1469–1492. doi:10.3109/07420528.2010.503293
24. Crowley SJ, Acebo C, Fallone G, Carskadon MA. Estimating dim light melatonin onset (DLMO) phase in adolescents using summer or school-year sleep/wake schedules. Sleep. 2006;29(12):1632–1641. doi:10.1093/sleep/29.12.1632
25. Eto T, Kitamura S, Nishimura K, et al. Circadian phase advances in children during camping life according to the natural light-dark cycle. J Physiol Anthropol. 2022;41(1):42. doi:10.1186/s40101-022-00316-x
26. Hasler BP, Oryshkewych N, Wallace ML, Clark DB, Siegle GJ, Buysse DL. Circadian phase in high school students: weekday-weekend shifts and relationships to other sleep/circadian characteristics. Sleep. 2025;48:zsaf031. doi:10.1093/sleep/zsaf031
27. Zerbini G, Winnebeck EC, Merrow M. Weekly, seasonal, and chronotype-dependent variation of dim-light melatonin onset. J Pineal Res. 2021;70(3):e12723. doi:10.1111/jpi.12723
28. Zerbini G, Merrow M, Winnebeck EC. Weekly and seasonal variation in the circadian melatonin rhythm in humans: a response. J Pineal Res. 2022;72(1):e12777. doi:10.1111/jpi.12777
29. Rosekind MR, Gregory KB, Mallis MM, Brandt SL, Seal B, Lerner D. The cost of poor sleep: workplace productivity loss and associated costs. J Occup Environ Med. 2010;52(1):91–98. doi:10.1097/JOM.0b013e3181c78c30
30. Hafner M, Stepanek M, Taylor J, Troxel WM, van Stolk C. Why sleep matters-the economic costs of insufficient sleep: a cross-country comparative analysis. Rand Health Q. 2017;6(4):11.
31. Pilcher JJ, Huffcutt AI. Effects of sleep deprivation on performance: a meta-analysis. Sleep. 1996;19(4):318–326. doi:10.1093/sleep/19.4.318
32. Philibert I. Sleep loss and performance in residents and nonphysicians: a meta-analytic examination. Sleep. 2005;28(11):1392–1402. doi:10.1093/sleep/28.11.1392
33. Killgore WD. Effects of sleep deprivation on cognition. Prog Brain Res. 2010;185:105–129. doi:10.1016/B978-0-444-53702-7.00007-5
34. Leong RLF, Chee MWL. Understanding the Need for Sleep to Improve Cognition. Annu Rev Psychol. 2023;74:27–57. doi:10.1146/annurev-psych-032620-034127
35. Yoo SS, Hu PT, Gujar N, Jolesz FA, Walker MP. A deficit in the ability to form new human memories without sleep. Nat Neurosci. 2007;10(3):385–392. doi:10.1038/nn1851
36. Waters F, Bucks RS. Neuropsychological effects of sleep loss: implication for neuropsychologists. J Int Neuropsychol Soc. 2011;17(4):571–586. doi:10.1017/S1355617711000610
37. Lowe CJ, Safati A, Hall PA. The neurocognitive consequences of sleep restriction: a meta-analytic review. Neurosci Biobehav Rev. 2017;80:586–604. doi:10.1016/j.neubiorev.2017.07.010
38. Paller KA, Creery JD, Schechtman E. Memory and sleep: how sleep cognition can change the waking mind for the better. Annu Rev Psychol. 2021;72:123–150. doi:10.1146/annurev-psych-010419-050815
39. Barnes CM, Watson NF. Why healthy sleep is good for business. Sleep Med Rev. 2019;47:112–118. doi:10.1016/j.smrv.2019.07.005
40. Bacaro V, Carpentier L, Crocetti E. Sleep well, study well: a systematic review of longitudinal studies on the interplay between sleep and school experience in adolescence. Int J Environ Res Public Health. 2023;20(6):4829. doi:10.3390/ijerph20064829
41. Mitler MM, Carskadon MA, Czeisler CA, Dement WC, Dinges DF, Graeber RC. Catastrophes, sleep, and public policy: consensus report. Sleep. 1988;11(1):100–109. doi:10.1093/sleep/11.1.100
42. Uehli K, Mehta AJ, Miedinger D, et al. Sleep problems and work injuries: a systematic review and meta-analysis. Sleep Med Rev. 2014;18(1):61–73. doi:10.1016/j.smrv.2013.01.004
43. Czeisler CA, Wickwire EM, Barger LK, et al. Sleep-deprived motor vehicle operators are unfit to drive: a multidisciplinary expert consensus statement on drowsy driving. Sleep Health. 2016;2(2):94–99. doi:10.1016/j.sleh.2016.04.003
44. Goldstein AN, Walker MP. The role of sleep in emotional brain function. Annu Rev Clin Psychol. 2014;10:679–708. doi:10.1146/annurev-clinpsy-032813-153716
45. Ben Simon E, Vallat R, Barnes CM, Walker MP. Sleep loss and the socio-emotional brain. Trends Cognit Sci. 2020;24(6):435–450. doi:10.1016/j.tics.2020.02.003
46. Tomaso CC, Johnson AB, Nelson TD. The effect of sleep deprivation and restriction on mood, emotion, and emotion regulation: three meta-analyses in one. Sleep. 2021;44(6):zsaa289. doi:10.1093/sleep/zsaa289
47. Ben Simon E, Walker MP. Sleep loss causes social withdrawal and loneliness. Nat Commun. 2018;9(1):3146. doi:10.1038/s41467-018-05377-0
48. Pigeon WR, Pinquart M, Conner K. Meta-analysis of sleep disturbance and suicidal thoughts and behaviors. J Clin Psychiatry. 2012;73(9):e1160–7. doi:10.4088/JCP.11r07586
49. Bryant PA, Trinder J, Curtis N. Sick and tired: does sleep have a vital role in the immune system? Nat Rev Immunol. 2004;4(6):457–467. doi:10.1038/nri1369
50. Kakizaki M, Inoue K, Kuriyama S, et al. Ohsaki Cohort Study. Sleep duration and the risk of prostate cancer: the Ohsaki Cohort Study. Br J Cancer. 2008;99(1):176–178. doi:10.1038/sj.bjc.6604425
51. von Ruesten A, Weikert C, Fietze I, Boeing H. Association of sleep duration with chronic diseases in the European Prospective Investigation into Cancer and Nutrition (EP-IC)-Potsdam study. PLoS One. 2012;7:e30972. doi:10.1371/journal.pone.0030972
52. Gapstur SM, Diver WR, Stevens VL, Carter BD, Teras LR, Jacobs EJ. Work schedule, sleep duration, insomnia, and risk of fatal prostate cancer. Am J Prev Med. 2014;46(3 Suppl 1):S26–33. doi:10.1016/j.amepre.2013.10.033
53. Ayas NT, White DP, Manson JE, et al. A prospective study of sleep duration and coronary heart disease in women. Arch Intern Med. 2003;163(2):205–209. doi:10.1001/archinte.163.2.205
54. Knutson KL, Van Cauter E, Rathouz PJ, et al. Association between sleep and blood pressure in midlife: the CARDIA sleep study. Arch Intern Med. 2009;169(11):1055–1061. doi:10.1001/archinternmed.2009.119
55. Cappuccio FP, Cooper D, D’Elia L, Strazzullo P, Miller MA. Sleep duration predicts cardiovascular outcomes: a systematic review and meta-analysis of prospective studies. Eur Heart J. 2011;32(12):1484–1492. doi:10.1093/eurheartj/ehr007
56. Jackson CL, Redline S, Emmons KM. Sleep as a potential fundamental contributor to disparities in cardiovascular health. Annu Rev Public Health. 2015;36:417–440. doi:10.1146/annurev-publhealth-031914-122838
57. Gottlieb DJ, Punjabi NM, Newman AB, et al. Association of sleep time with diabetes mellitus and impaired glucose tolerance. Arch Intern Med. 2005;165(8):863–867. doi:10.1001/archinte.165.8.863
58. Knutson KL, Van Cauter E. Associations between sleep loss and increased risk of obesity and diabetes. Ann N Y Acad Sci. 2008;1129:287–304. doi:10.1196/annals.1417.033
59. Markwald RR, Melanson EL, Smith MR, et al. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc Natl Acad Sci U S A. 2013;110(14):5695–5700. doi:10.1073/pnas.1216951110
60. Davies SK, Ang JE, Revell VL, et al. Effect of sleep deprivation on the human metabolome. Proc Natl Acad Sci U S A. 2014;111(29):10761–10766. doi:10.1073/pnas.1402663111
61. Bernert RA, Kim JS, Iwata NG, Perlis ML. Sleep disturbances as an evidence-based suicide risk factor. Curr Psychiatry Rep. 2015;17(3):554. doi:10.1007/s11920-015-0554-4
62. Irwin MR. Why sleep is important for health: a psychoneuroimmunology perspective. Annu Rev Psychol. 2015;66:143–172. doi:10.1146/annurev-psych-010213-115205
63. Besedovsky L, Lange T, Haack M. The sleep-immune crosstalk in health and disease. Physiol Rev. 2019;99(3):1325–1380. doi:10.1152/physrev.00010.2018
64. Dew MA, Hoch CC, Buysse DJ, et al. Healthy older adults’ sleep predicts all-cause mortality at 4 to 19 years of follow-up. Psychosom Med. 2003;65(1):63–73. doi:10.1097/01.psy.0000039756.23250.7c
65. Cappuccio FP, D’Elia L, Strazzullo P, Miller MA. Sleep duration and all-cause mortality: a systematic review and meta-analysis of prospective studies. Sleep. 2010;33(5):585–592. doi:10.1093/sleep/33.5.585
66. Itani O, Jike M, Watanabe N, Kaneita Y. Short sleep duration and health outcomes: a systematic review, meta-analysis, and meta-regression. Sleep Med. 2017;32:246–256. doi:10.1016/j.sleep.2016.08.006
67. García-Perdomo HA, Zapata-Copete J, Rojas-Cerón CA. Sleep duration and risk of all-cause mortality: a systematic review and meta-analysis. Epidemiol Psychiatr Sci. 2019;28(5):578–588. doi:10.1017/S2045796018000379
68. Smith MR, Eastman CI. Shift work: health, performance and safety problems, traditional countermeasures, and innovative management strategies to reduce circadian misalignment. Nat Sci Sleep. 2012;4:111–132. doi:10.2147/NSS.S10372
69. Misiunaite I, Eastman CI, Crowley SJ. Circadian phase advances in response to weekend morning light in adolescents with short sleep and late bedtimes on school nights. Front Neurosci. 2020;14:99. doi:10.3389/fnins.2020.00099
70. Crowley SJ, Velez SL, Killen LG, Cvengros JA, Fogg LF, Eastman CI. Extending weeknight sleep of delayed adolescents using weekend morning bright light and evening time management. Sleep. 2023;46(1):zsac202. doi:10.1093/sleep/zsac202
71. Putilov AA, Sveshnikov DS, Bakaeva ZB, et al. The irrecoverable loss in sleep on weekdays of two distinct chronotypes can be equalized by permitting a >2 h difference in waking time. Appl Sci. 2022;12:8092. doi:10.3390/app12168092
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Weekday-Weekend Shifts of Light/Dark Regimen Extend Sleep and Lifespan of Drosophila melanogaster
Zakharenko LP, Petrovskii DV, Putilov AA
Nature and Science of Sleep 2025, 17:3193-3206
Published Date: 24 December 2025