Back to Journals » Cancer Management and Research » Volume 10
Calcium channel α2δ1 subunit (CACNA2D1) enhances radioresistance in cancer stem-like cells in non-small cell lung cancer cell lines
Authors Sui X, Geng JH, Li YH ![]() , Zhu GY, Wang WH
, Zhu GY, Wang WH
Received 7 June 2018
Accepted for publication 16 August 2018
Published 26 October 2018 Volume 2018:10 Pages 5009—5018
DOI https://doi.org/10.2147/CMAR.S176084
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Raphael Catane
Xin Sui,1,* Jian-Hao Geng,1,* Yong-Heng Li,1 Guang-Ying Zhu,2 Wei-Hu Wang1
1Key Laboratory of Carcinogenesis and Translational Research (Ministry of Education/Beijing), Department of Radiation Oncology, Peking University Cancer Hospital and Institute, Beijing, China; 2Department of Radiation Oncology, National Clinical Research Center for Respiratory Disease, Center for Respiratory Disease, Lung Cancer Center, China-Japan Friendship Hospital, Peking University Health Science Center, Beijing, China
*These authors contributed equally to this work
Purpose: Radiotherapy is a major treatment method for patients with non-small cell lung cancer (NSCLC). However, the presence of radioresistant cancer stem cells (CSCs) may be associated with disease relapse or a poor outcome after radiotherapy. Voltage-gated calcium channel α2δ1 subunit (encoded by the gene CACNA2D1) isoform 5 is a marker of CSCs in hepatocellular carcinoma. This study aimed to investigate the radiosensitivity of α2δ1-high cells in NSCLC cell lines.
Materials and methods: NSCLC cell lines A549, H1975, H1299, and PC9 were used. CACNA2D1-knockdown and CACNA2D1-overexpressing cell lines were established by lentiviral infection. Colony formation assay was performed to determine radiosensitivity. Sphere formation assay in serum-free medium was performed to evaluate self-renewal capacity. Proteins associated with DNA damage repair were analyzed by immunofluorescence or Western blot. The monoclonal antibody of α2δ1 was applied alone or in combination with radiation either in vitro or in vivo to determine the anti-tumor effect of the antibody.
Results: α2δ1-high cells showed greater sphere-forming efficiency than α2δ1-low cells and were relatively resistant to radiation. CACNA2D1 knockdown in A549 cells enhanced radiosensitivity, whereas CACNA2D1 overexpression in PC9 and H1975 cells reduced radiosensitivity, suggesting that α2δ1 imparted radioresistance to NSCLC cells. Analysis of proteins involved in DNA damage repair suggested that α2δ1 enhanced the efficiency of DNA damage repair. The monoclonal antibody of α2δ1 had a synergistic effect with that of radiation to block the self-renewal of α2δ1-high cells and enhanced the radiosensitivity of α2δ1-positive cells in colony formation assays. The combination of the α2δ1 antibody with radiation repressed A549 xenograft growth in vivo.
Conclusion: α2δ1 enhances radioresistance in cancer stem-like cells in NSCLC. The α2δ1 monoclonal antibody sensitizes α2δ1-high cells to radiation, suggesting that the antibody may be used to improve the treatment outcome when combined with radiation in NSCLC.
Keywords: radioresistance, cancer stem cell, calcium channel, DNA damage repair
Introduction
Radiotherapy is a major treatment method for patients with non-small cell lung cancer (NSCLC). However, a certain proportion of tumors do not respond effectively to radiation or relapse after radiotherapy, which may be due to the existence of radioresistant cancer stem cells (CSCs).1,2 CSCs are typified by their self-renewal capacity and ability to generate heterogenous cancer cells that constitute the bulk of the tumor.3,4 Radioresistance of CSCs has been reported in several types of malignant tumors. In glioma, CD133-positive CSCs activate the DNA damage checkpoint more effectively, leading to more rapid DNA damage repair.5 Inhibitor of CHK1 and CHK2 checkpoint kinases can abrogate the radioresistance of CD133-positive cells.5,6 A study on breast cancer has shown that CD24-negative/low CD49f-positive Lin-negative CSCs exhibit higher levels of free-radical scavenging systems, resulting in lower levels of reactive oxygen species (ROS) and resistance to radiation. Moreover, ROS scavenger inhibitors radiosensitize breast CSCs.7 In lung cancer cell lines, the upregulation of CSC markers, such as CD44, CD24, and aldehyde dehydrogenase (ALDH)1 after radiation has been reported, suggesting the radioresistance of CSCs in lung cancer.8,9 Therefore, drugs specifically targeting CSCs could be promising approaches to avoid radioresistance without increasing normal tissue damage.
A specific surface antibody targeting CSCs, namely 1B50-1, was identified using subtractive immunization by a pair of hepatocellular carcinoma (HCC) cell lines, Hep-11 and Hep-12. These two cell lines were established from primary and recurrent HCC tissue respectively, from the same patient. 1B50-1 specifically targeted the Hep-12 cells, which showed tumor-initiating capacity. By immunoprecipitation and mass spectrometry analysis, the antigen of 1B50-1 was identified as calcium channel α2δ1 subunit (encoded by the gene CACNA2D1) isoform 5.10 α2δ1-positive HCC cells express higher levels of stem cell-associated genes and show high tumorigenicity. When combined with a chemotherapy regimen, 1B50-1 exhibits a therapeutic effect on HCC engraftment by eliminating CSCs. Notably, α2δ1 isoform 5 is specifically expressed in HCC stem cells and is undetectable in most normal tissues. This expression pattern highlights α2δ1 isoform 5 as a promising candidate for targeting CSC.10
In this study, we observed that the radioresistance of several NSCLC cell lines was correlated with their α2δ1 expression levels. Therefore, we investigated the radiosensitivity of α2δ1-high cells among the NSCLC cell lines and evaluated the effect of the α2δ1 monoclonal antibody on the radiosensitivity of α2δ1-high NSCLC cells.
Materials and methods
Cell culture
A549, H1299, and H1975 were purchased from American Type Culture Collection, and PC9 was a gift from Dr Yuanyuan Ma in th Department of Thoracic Surgery II, Peking University Cancer Hospital and Institute. The identity of PC9 was confirmed by Short Tandem Repeat profiling. The research was approved by the ethics committee of Peking University Cancer Hospital and Institute. Cells were cultured in RPMI-1640 (Invitrogen, Waltham, MA, USA) supplemented with 10% FBS (Invitrogen). To establish CACNA2D1-overexpressing or CACNA2D1-knockdown cell lines, lentivirus was generated according to the protocol of Zhao et al10 Cells were transfected with lentivirus and selected for blasticidin resistance according to the protocol of Lan et al11
Colony formation assay
Colony formation assay was performed to determine the radiosensitivity of the cells. Cells were seeded at densities of 100–1,000 cells per well in 12-well plates 1 day before irradiation. Culture medium was added to a height of 1 cm to ensure dose build-up. Cell irradiation was performed with a 600-C/D linear accelerator (Varian Medical Systems, Inc., Palo Alto, CA, USA) with 6-MV X-ray. Culture medium was replaced after irradiation. After 7–10 days, the cells were fixed with 4% formaldehyde and stained with crystal violet. Colonies with >50 cells were counted. The colony formation efficiency was calculated as the number of colonies divided by the number of cells seeded. The survival fraction was calculated as the colony formation efficiency at a certain irradiation dose divided by the colony formation efficiency at 0 Gy.
Sphere formation assay
Sphere formation assay was performed to determine the self-renewal capacity of the cells. The cells were seeded at a density of 200 cells per well in 96-well ultralow-attachment plates (Corning, Tewksbury, MA, USA) in DMEM/F12 containing 1% methylcellulose (Sigma-Aldrich, St. Louis, MO, USA), 25 ng/mL EGF, 25 ng/mL basic fibroblast growth factor, and 2% B27. After 10-–14 days of seeding, the number of spheres was counted under a microscope. Reagents were purchased from Invitrogen if not otherwise indicated.
Flow cytometry
1B50-1 was conjugated with fluorescein with the Lightning-Link Fluorescein kit (Innova Biosciences Ltd., Cambridge, UK). The stocking concentration of the fluorescein isothiocyanate-conjugated α2δ1 antibody was 1.5 mg/mL. The antibody was titrated and the optimal working concentration was 7.5 µg/mL. Cells were digested with trypsin, labeled with 1B50-1-fluorescein at room temperature for 40 minutes, and washed three times with PBS. Flow cytometric analysis was performed on an Accuri C6 flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). The median fluorescence intensity (MFI) was calculated by FlowJo. Cell sorting was performed on a FACSAria flow cytometer (BD Biosciences).
Western blot
SDS-PAGE was performed, and protein was transferred to a polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA). The membrane was blocked with 5% milk (BD Biosciences) in tris-buffered saline-Tween 20 (TBST) for 1 hour and incubated with primary antibody for 3 hours at room temperature. Then the membrane was washed with TBST three times and incubated with secondary antibody for 1 hour at room temperature. The Immobilon western chemiluminescent substrate (Millipore) was applied to the membrane, and signals were captured with the MiniChemi luminescent imaging and analysis system (Sage Creation Science Co, Ltd., Beijing, China).
The primary antibodies used were as follows: mouse anti-α2δ1 (Abcam, Cambridge, UK, 1:2,500), rabbit anti-γH2AX (Cell Signaling Technologies, Danvers, MA, USA, 1:1,000), rabbit anti-ataxia telangiectasia mutated (ATM) (Cell Signaling Technologies, 1:1,000), rabbit anti-phospho-ATM (Ser1981) (Cell Signaling Technologies, 1:1,000), rabbit anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Biodragon, Beijing, China, 1:10,000), and rabbit anti-β-actin (Biodragon, 1:10,000). The secondary antibodies used were as follows: goat anti-mouse horseradish peroxidase (HRP)-conjugated antibody (Zhongshan, Beijing, China, 1: 100,000) and goat anti-rabbit HRP-conjugated antibody (Zhongshan, 1:100,000). All antibodies were diluted in TBST containing 5% milk.
Immunofluorescence
Cells were fixed with 4% formaldehyde for 5 minutes at room temperature and permeabilized with 0.5% Triton-X100 in PBS for 30 minutes. Cells were blocked in PBS with 5% goat serum (Zhongshan) for 1 hour and incubated with γH2AX antibody (Cell Signaling Technologies, 1:400) in PBS containing 5% goat serum at 4°C overnight. Cells were then washed with PBS three times, incubated with goat anti-rabbit rhodamine-conjugated secondary antibody (Zhongshan, 1:200) at room temperature for 1 hour, and washed with PBS three times. Nuclei were stained with DAPI. Cells were examined under a Leica SP5 confocal microscope (Leica, Wetzlar, Germany).
Quantitative PCR (qPCR)
RNA was extracted with TRIzol (Invitrogen) and reverse transcribed with Moloney murine leukemia virus reverse transcriptase (Invitrogen). qPCR was performed with the SybrGreen dye (Toyoba, Osaka, Japan) on an ABI 7500-Fast real-time PCR system (Applied Biosystems, Waltham, MA, USA). Relative expression was calculated as 2−ΔΔCt.
The primer sequences were as follows: CACNA2D1-F, CAGTTGAGATGGAGGATGATG; CACNA2D1-R, TTGTATGAGCAGTCGTGTGTC; NANOG-F, TGCCTCACACGGAGACTGTC; NANOG-R, TGCTATTCTTCGGCCAGTTG; OCT4-F, GACAACAATGAAAATCTTCAGGAGA; OCT4-R, CTGGCGCCGGTTACAGAACCA; RAD51-F, CAGTGATGTCCTGGATAATGTAGC; RAD51-R, TTACCACTGCTACACCAAACTCAT; BRCA1-F, AGCAGAATGGTCAACTGATGAATA; BRCA1-R, ACTGCTGCTTATAGGTTCAGCTTT; CHEK2-F, TGATCAGTCAGTTTATCCTAAGGC; CHEK2-R, TCGAAAGCCAGCTTTACCTC; MDC1-F, CAGTCCAGAGCATGGAGGAT; MDC1-R, AATGGCTGTGTAGCCAGGAC; GAPDH-F, GTCGGAGTCAACGGATTTGG; and GAPDH-R, AAAAGCAGCCCTGGTGACC.
Mouse xenograft model
Six-week-old female BALB/c nude mice were purchased from Vital River Laboratory Animal Technology Co. (Beijing, China). A total of 2×106 A549 cells were injected subcutaneously into the right leg of each mouse. When the tumor volume reached 200–250 mm3, mice were randomly assigned to four groups (n=5): control, antibody, radiation, and radiation plus antibody. For radiation, the mice were anesthetized by intraperitoneal injection of pentobarbital sodium. The right leg bearing the tumor was positioned in the radiation field. A tissue equivalent bolus of 1.0 cm thickness was placed on top of the tumor to provide dose build-up. A single dose of 10 Gy X-ray or sham irradiation was delivered by a 600-C/D linear accelerator (Varian Medical Systems, Inc., Palo Alto, CA, USA). A total of 800 µg of 1B50-1 or its isotype IgG3 was administered every other day by intraperitoneal injection. The tumor size was measured with a digital caliper, and the tumor volume was calculated with the following formula: V(mm3)=larger diameter×(smaller diameter)2×0.5. All animal experiments were performed in accordance with protocols approved by the ethics committee of Peking University Cancer Hospital and Institute.
Statistics
GraphPad Prism was used to analyze the data. The unpaired two-sided Student’s t-test was performed to evaluate the significance between groups. P<0.05 was considered to be statistically significant. Data are represented as the mean and SD if not otherwise indicated. Representative data from at least three biologically independent experiments with similar results are presented.
Results
α2δ1-positive NSCLC cells are relatively radioresistant
The radiosensitivity of several NSCLC cell lines was evaluated by colony formation assay. H1299 and A549 were relatively resistant to radiation, whereas H1975 was sensitive to radiation (Figure 1A). To determine the correlation between radiosensitivity and candidate CSC marker α2δ1, we analyzed the α2δ1-positive proportion by flow cytometry. The results showed that H1299 and A549 expressed relatively high level of α2δ1, PC9 expressed low level of α2δ1, and H1975 was negative for α2δ1 (Figure 1B). This pattern led to the hypothesis that α2δ1 expression was related to NSCLC cell radioresistance. Therefore, α2δ1-high and α2δ1-low A549 and H1299 cells were sorted. Colony formation analysis showed that compared with the α2δ1-low cells, the α2δ1-high cells were relatively resistant to ionizing radiation (Figure 1C–E).
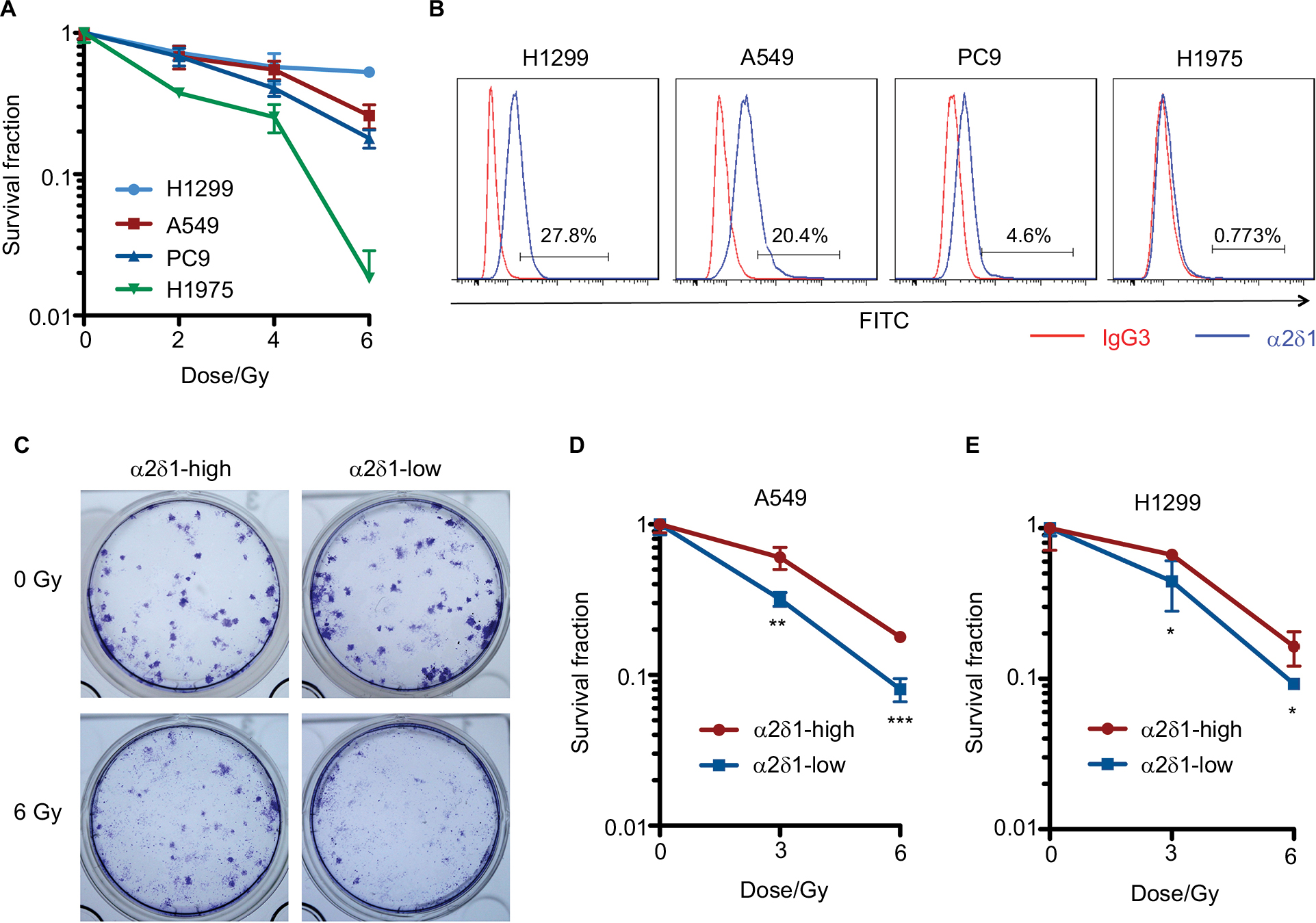 | Figure 1 α2δ1 expression is associated with radioresistance in NSCLC cell lines. Notes: (A) Survival curves of NSCLC cell lines. (B) Flow cytometric analysis of the α2δ1 proportion of each NSCLC cell line. IgG3 is the isotype control. The ratio of MFI-α2δ1 over MFI-isotype was 3.46 in H1299, 3.22 in A549, 1.76 in PC9, and 1.13 in H1975. (C) Representative images of colonies formed by α2δ1-high and α2δ1-low A549 cells upon radiation. (D, E) Survival curves of α2δ1-high and α2δ1-low A549 (D) and H1299 (E) cells. The numbers of seeded cells were 200 cells per well for 0 Gy, 400 cells for 3 Gy, and 800 cells for 6 Gy, respectively. *P<0.05, **P<0.01, ***P<0.001. Abbreviations: FITC, fluorescein isothiocyanate; MFI, median fluorescence intensity; NSCLC, non-small cell lung cancer. |
α2δ1 marks CSCs in HCC.10 To investigate the feasibility of α2δ1 as a CSC marker in NSCLC, we sorted the α2δ1-high and α2δ1-low cells and evaluated the self-renewal capacity by sphere formation analysis. The α2δ1-high cells had a higher sphere-forming capacity than their α2δ1-low counterparts, and the sphere formation efficiency of α2δ1-high cells increased in subsequent serial propagation (Figure 2A, B). We used lentivirus to overexpress CACNA2D1 in the α2δ1-negative H1975 and α2δ1-low PC9 cell lines. CACNA2D1 overexpression increased the sphere formation efficiency (Figure 2C–F). Conversely, CACNA2D1 knockdown in A549 cells resulted in a reduction in the sphere formation efficiency (Figure 2G, H). These results indicated that the α2δ1-positive cells had high self-renewal capacity, which was a major characteristic of CSCs.
 | Figure 2 α2δ1 marks the radioresistant cancer stem-like cells. Notes: (A) Morphology of the spheres formed by the sorted α2δ1-high and α2δ1-low A549 cells (bar=200 µm). (B) Sphere formation efficiency of α2δ1-high and α2δ1-low A549 cells. (C) Western blot of α2δ1 expression in the control and CACNA2D1-overexpressing H1975 cells. (D) Sphere formation efficiency of the control and CACNA2D1-overexpressing H1975 cells. (E, F) Western blot and sphere formation efficiency of the control and CACNA2D1-overexpressing PC9 cells. (G, H) Western blot and sphere formation efficiency of the control and CACNA2D1-knockdown A549 cells. (I) Flow cytometric analysis of the α2δ1 expression of the A549 cells 48 hours after irradiation. The MFI of was as follows: isotype control (IgG3)–7688, 0 Gy-24790, 4 Gy-32177, 8 Gy-38865. *P<0.05, **P<0.01, ***P<0.001. Abbreviations: CTRL, control; FITC, fluorescein isothiocyanate; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; MFI, median fluorescence intensity; OE, overexpression. |
Studies on radioresistant CSCs have shown that the percentage of CSCs increases after radiation.5,7,12 We found that the α2δ1-positive proportion was enhanced after radiation in a dose-dependent manner (Figure 2I). These data demonstrated that the α2δ1-high NSCLC stem-like cells were relatively radioresistant.
α2δ1 imparts radioresistance to NSCLC cells
To determine whether α2δ1 was just a surface marker for radioresistant NSCLC stem cells or it played a role in the acquisition of radioresistance, we performed a colony formation analysis of CACNA2D1-overexpressing H1975 and PC9 cells and CACNA2D1-knockdown A549 cells. The survival curves showed that radioresistance was acquired in CACNA2D1-overexpressing H1975 and PC9 cells (Figure 3A, B). In contrast, CACNA2D1 knockdown by shRNA sensitized A549 cell line to radiation (Figure 3C). The changes in radiosensitivity induced by the overexpression or knockdown of CACNA2D1 suggested that α2δ1 imparted radioresistance to the NSCLC cells.
 | Figure 3 α2δ1 imparts radioresistance to NSCLC cells. Notes: Representative images of the colonies and survival curves of the control and CACNA2D1-overexpressing H1975 (A) and PC9 (B) cells and the CACNA2D1-knockdown A549 cells (C). The numbers of seeded cells were as follows: H1975 – 200 cells per well for 0 Gy, 200 cells for 2 Gy, 400 cells for 4 Gy, and 800 cells for 6 Gy; PC9 – 100 cells per well for 0 Gy, 100 cells for 2 Gy, 200 cells for 4 Gy, 400 cells for 6 Gy, and 800 cells for 8 Gy; A549 – 100 cells per well for 0 Gy, 200 cells for 4 Gy, and 800 cells for 8 Gy. *P<0.05, **P<0.01, ***P<0.001. Abbreviations: CTRL, control; OE, overexpression; NSCLC, non-small cell lung cancer. |
Radiosensitivity is associated with the capacity of DNA damage repair. We examined the kinetics of γH2AX, which marks the DNA double-strand break sites. Immunofluorescence staining showed that the number of γH2AX foci increased after radiation and that the number of foci decreased more rapidly in the CACNA2D1-overexpressing cells than in the control group, indicating a more efficient capacity of DNA damage repair in the CACNA2D1-overexpressing H1975 cells (Figure 4A). The Western blot results also showed faster downregulation of γH2AX in the CACNA2D1-overexpressing group than in the control one (Figure 4B). We also examined the level of phosphorylated ATM, which initiates the DNA damage repair machinery.13 In the sorted A549 cells, the α2δ1-high cells showed higher levels of activated ATM upon radiation, which represented a more efficient capacity of DNA damage repair (Figure 4C). These data collectively suggested that α2δ1 imparted the radioresistance to NSCLC cells partially by enhancing the efficiency DNA damage repair in response to radiation.
 | Figure 4 The capacity of DNA damage repair is enhanced by α2δ1. Notes: (A) Immunofluorescence of γH2AX foci in the control and CACNA2D1-overexpressing H1975 cells after 2-Gy radiation (bar=20 µm). (B) Western blot of the γH2AX levels in the control and CACNA2D1-overexpressing H1975 cells after 2-Gy radiation. (C) Western blot analysis of phosphorylated ATM at Ser1981 and total ATM 1 hour after 2-Gy radiation of the sorted α2δ1-high and α2δ1-low A549 cells. (D, E) Relative expression of stemness and DNA damage repair-related genes by quantitative PCR in the control and CACNA2D1-overexpressing H1975 cells (D) and in the control and CACNA2D1-knockdown A549 cells (E). (F) Correlation between CACNA2D1 expression and NANOG, RAD51, BRCA1, or CHEK2 expression by GEO profile analysis in data set GSE4115. *P<0.05, **P<0.01, ***P<0.001. Abbreviations: ATM, ataxia telangiectasia mutated; CACNA2D1, calcium channel subunit α2δ1; CHEK2, checkpoint kinase 2; CTRL, control; GEO, Gene Expression Omnibus; IR, irradiation; MDC1, mediator of DNA damage checkpoint 1; NS, not significant; OE, overexpression; pS1981, phosphorylated serine 1981; γH2AX, phosphorylated histone H2A member X. |
The expression levels of other DNA damage repair-related genes and stem cell markers were analyzed by qPCR. Among these genes, NANOG, a transcription factor that regulates embryonic stem cell pluripotency and is associated with CSCs and a worse clinical outcome,14 and RAD51, the recombinase catalyzing the process of homologous recombination after DNA double-strand breaks,15 were upregulated in the CACNA2D1-overexpressing H1975 cells and downregulated in the CACNA2D1-knockdown A549 cells. DNA damage repair-related genes BRCA1, CHEK2, and MDC1 were also upregulated in CACNA2D1-overexpressing H1975 in a mild manner, but not all these genes were downregulated in CACNA2D1-knockdown A549. The expression of OCT4 was not affected by α2δ1 overexpression or knockdown (Figure 4D–E). We also performed Gene Expression Omnibus (GEO) profile analysis of CACNA2D1 and DNA damage repair-related genes. In a data set of histologically normal large-airway epithelial cells from smokers with suspected lung cancer (GSE4115),16 the GEO profiles of the smokers who were ultimately diagnosed with lung cancer showed that the expression of CACNA2D1 was also positively correlated with the expression of NANOG, RAD51, BRCA1, and CHEK2 (Figure 4F). These results also implied the correlation between α2δ1 and the capacity of DNA damage repair.
1B50-1 blocks the self-renewal capacity of α2δ1-positive cells and enhances the radiosensitivity
1B50-1, the α2δ1 monoclonal antibody raised against a recurrent HCC cell line, blocks sphere formation in α2δ1-positive HCC cells and has a synergistic effect with that of chemotherapy.10 We applied this antibody to the NSCLC cell lines and found that in the sorted α2δ1-high A549 cells, the 1B50-1 treatment blocked sphere formation (Figure 5A). Moreover, the combination of 1B50-1 and ionizing radiation reduced sphere formation to a much lower level (Figure 5A). In the colony formation assay, the 1B50-1 treatment enhanced the radiosensitivity of the α2δ1-high cells (Figure 5B). Conversely, 1B50-1 had a mild effect on the α2δ1-low cells (data not shown).
 | Figure 5 The α2δ1 monoclonal antibody blocks the self-renewal capacity and enhances the radiosensitivity of α2δ1-high cells. Notes: (A) The sphere formation efficiency of α2δ1-high A549 cells treated with 25 µg/mL α2δ1 antibody 1B50-1, 2-Gy radiation or the combination of 1B50-1 and radiation. IgG3 is the isotype control. (B) Survival curves of α2δ1-high A549 cells treated with 50 µg/mL 1B50-1 or the isotype control. (C) Tumor volumes of the A549 xenografts in the nude mice receiving the indicated treatments. *P<0.05, **P<0.01, ***P<0.001. Abbreviation: IR, irradiation. |
To explore the effect of 1B50-1 on NSCLC cells in vivo, subcutaneous A549 xenografts were established in nude mice. The mice were treated with 1B50-1 or radiation, or the combination of 1B50-1 and radiation. The 1B50-1 treatment alone had little effect on tumor size, whereas when combined with a single dose of 10 Gy radiation, tumor growth was blocked effectively. The combination modality also showed an advantage over radiation alone (Figure 5C). These results suggested that α2δ1 monoclonal antibody 1B50-1 specifically blocked the self-renewal capacity of the α2δ1-high cells and enhanced their radiosensitivity. The antibody had a synergistic effect with that of radiation on repressing the growth of the high α2δ1-expressing NSCLC cells in the mouse model.
Discussion
CSCs showed the capacity of self-renewal and tumor initiation and were resistant to cytotoxic therapies. Calcium channel α2δ1 subunit isoform 5 has been identified as a CSC marker in HCC.10 In this study, we demonstrated that the α2δ1-high NSCLC cells also had self-renewal capacity and were resistant to radiation. The overexpression of CACNA2D1 imparted radioresistance to the NSCLC cells with a more efficient capacity of DNA damage repair after radiation. The α2δ1 monoclonal antibody blocked the self-renewal capacity of the α2δ1-high cells and sensitized them to radiation. Therefore, we propose α2δ1 as a target to eliminate radioresistant NSCLC stem cells.
The presence of CSCs in NSCLC has been reported, and CSCs have been selected based on CD133, CD166, CD44 positivity or ALDH activity17–20, or with serum-free self-renewal sphere culture medium.21 We also examined the expression of CD166 in our experiments. CD166 expression was broad in A549, PC9, and H1975, and was about 50% in H1299, partially overlapping with that of α2δ1 (data not shown). The expression pattern of CD166 is not correlated with radioresistance, whereas the correlation with radioresistance is observed in α2δ1 expression. Therefore, we mainly focused on how α2δ1 regulates the radiosensitivity in NSCLC cell lines. In this study, the α2δ1-positive cells showed a higher sphere formation capacity in serum-free self-renewal medium than the α2δ1-negative cells, suggesting the feasibility of α2δ1 as a CSC marker. α2δ1 expression has also been reported to be associated with poor overall survival and progression-free survival in epithelial ovarian cancer.22 Additionally, downregulation of CACNA2D1 by miR-107 promotes erythroid differentiation of chronic myeloid leukemia cells.23 These studies also supported α2δ1 as a marker for CSCs.
CSCs are relatively resistant to conventional treatments, including radiotherapy. Our results showed that the α2δ1-high NSCLC cells were resistant to radiation compared with the α2δ1-low cells. CACNA2D1 overexpression in the α2δ1-negative or α2δ1-low cell lines resulted in radioresistance, whereas CACNA2D1 knockdown in the α2δ1-high cell line enhanced radiosensitivity. These data suggested that α2δ1 was more than just a surface marker of CSCs and radioresistance; it participated in the acquisition of intrinsic resistance to radiation. Ion channels play complicated roles in tumorigenesis and the sensitivity of tumors to treatment. Several ion channels have been reported to confer radioresistance via multiple mechanisms. Potassium channel activity is induced upon radiation in response to oxidative stress.24 Palme et al have demonstrated that radiation activates the Kv3.4 voltage-gated potassium channel and enhances calcium entry, with subsequent activation of CaMKII and inactivation of cdc25B and cdc2. Kv3.4 channel inhibition with drugs or siRNA enhances radiosensitivity.25 It has also been reported that radiation-stimulated Ca2+ entry and CaMKII activiation participated in G2/M cell cycle arrest and survival of irradiated chronic myeloid leukemia cells.26 α2δ1 is a voltage-gated calcium channel. Previous study on HCC has shown that α2δ1 regulates calcium influx and maintains calcium oscillation in the tumor-initiating cells of HCC and that the intracellular calcium concentration is elevated upon CACNA2D1 overexpression.10 Moreover, phosphorylated ERK1/2 was upregulated by CACNA2D1 overexpression. This mechanism might be involved in the maintenance of CSC property and resistance to chemotherapy.10,27 In the NSCLC cell lines in this study, the increased phosphorylation level of ATM, rapid decrease in γH2AX after radiation and upregulation of DNA damage repair gene RAD51 in the CACNA2D1-overexpressing cell line or α2δ1-high cells suggests that α2δ1 might enhance radioresistance by imparting a more efficient capacity to repair DNA damage.
α2δ1 isoform 5 is the antigen of 1B50-1, which is the monoclonal antibody raised against recurrent HCC. 1B50-1 blocks the self-renewal of HCC stem cells.10 In this study, our results showed that 1B50-1 had a synergistic effect with that of radiation toward suppressing sphere and colony formations among α2δ1-high NSCLC cells. Conversely, 1B50-1 had little effect on α2δ1-low cells, which were relatively more sensitive to radiation and had a much lower capacity of self-renewal, suggesting the specificity of 1B50-1 for targeting α2δ1-high cells. The targeting of CSCs has been proposed to repress tumor recurrence after radiation in preclinical mouse model. Li et al have reported that a CD44s antibody targets pancreatic tumor-initiating cells and represses the growth, metastasis, and recurrence of pancreatic cancer xenografts after radiotherapy.28 In our study, 1B50-1 had a synergistic effect with that of radiation toward repressing xenograft growth in vivo. Considering the specificity of 1B50-1 on α2δ1-high NSCLC stem cells, 1B50-1 has the potential to eliminate CSCs when combined with radiotherapy in the treatment of NSCLC.
Conclusion
Our results suggest that α2δ1-high NSCLC cells are resistant to radiation and that α2δ1 imparts radioresistance partially by enhancing the efficiency for repair of DNA damage. Targeting α2δ1-high cells with the monoclonal antibody increases the radiosensitivity of α2δ1-high cells, suggesting the potential of the antibody to improve the treatment outcome when combined with radiation.
Acknowledgment
This work was supported by National Natural Science Foundation of China (81402535).
Disclosure
The authors report no conflicts of interest in this work.
References
Brunner TB, Kunz-Schughart LA, Grosse-Gehling P, Baumann M. Cancer stem cells as a predictive factor in radiotherapy. Semin Radiat Oncol. 2012;22(2):151–174. | ||
Baumann M, Krause M, Hill R. Exploring the role of cancer stem cells in radioresistance. Nat Rev Cancer. 2008;8(7):545–554. | ||
Clarke MF, Dick JE, Dirks PB, et al. Cancer stem cells: perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res. 2006;66(19):9339–9344. | ||
Boesch M, Zeimet AG, Reimer D, et al. The side population of ovarian cancer cells defines a heterogeneous compartment exhibiting stem cell characteristics. Oncotarget. 2014;5(16):7027–7039. | ||
Bao S, Wu Q, Mclendon RE, et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444(7120):756–760. | ||
Wang WJ, Wu SP, Liu JB, et al. MYC regulation of CHK1 and CHK2 promotes radioresistance in a stem cell-like population of nasopharyngeal carcinoma cells. Cancer Res. 2013;73(3):1219–1231. | ||
Diehn M, Cho RW, Lobo NA, et al. Association of reactive oxygen species levels and radioresistance in cancer stem cells. Nature. 2009;458(7239):780–783. | ||
Gomez-Casal R, Bhattacharya C, Ganesh N, et al. Non-small cell lung cancer cells survived ionizing radiation treatment display cancer stem cell and epithelial-mesenchymal transition phenotypes. Mol Cancer. 2013;12(1):94. | ||
Mihatsch J, Toulany M, Bareiss PM, et al. Selection of radioresistant tumor cells and presence of ALDH1 activity in vitro. Radiother Oncol. 2011;99(3):300–306. | ||
Zhao W, Wang L, Han H, et al. 1B50-1, a mAb raised against recurrent tumor cells, targets liver tumor-initiating cells by binding to the calcium channel α2δ1 subunit. Cancer Cell. 2013;23(4):541–556. | ||
Lan L, Han H, Zuo H, et al. Upregulation of myosin Va by Snail is involved in cancer cell migration and metastasis. Int J Cancer. 2010;126(1):53–64. | ||
Cahu J, Bustany S, Sola B. Senescence-associated secretory phenotype favors the emergence of cancer stem-like cells. Cell Death Dis. 2012;3:e446. | ||
Lavin MF, Kozlov S. DNA damage-induced signalling in ataxia-telangiectasia and related syndromes. Radiother Oncol. 2007;83(3):231–237. | ||
Jeter CR, Yang T, Wang J, Chao HP, Tang DG. Concise Review: NANOG in cancer stem cells and tumor development: an update and outstanding questions. Stem Cells. 2015;33(8):2381–2390. | ||
Gasparini P, Lovat F, Fassan M, et al. Protective role of miR-155 in breast cancer through RAD51 targeting impairs homologous recombination after irradiation. Proc Natl Acad Sci U S A. 2014;111(12):4536–4541. | ||
Spira A, Beane JE, Shah V, et al. Airway epithelial gene expression in the diagnostic evaluation of smokers with suspect lung cancer. Nat Med. 2007;13(3):361–366. | ||
Eramo A, Lotti F, Sette G, et al. Identification and expansion of the tumorigenic lung cancer stem cell population. Cell Death Differ. 2008;15(3):504–514. | ||
Zhang WC, Shyh-Chang N, Yang H, et al. Glycine decarboxylase activity drives non-small cell lung cancer tumor-initiating cells and tumorigenesis. Cell. 2012;148(1–2):259–272. | ||
Leung EL, Fiscus RR, Tung JW, et al. Non-small cell lung cancer cells expressing CD44 are enriched for stem cell-like properties. PLoS One. 2010;5(11):e14062. | ||
Sullivan JP, Spinola M, Dodge M, et al. Aldehyde dehydrogenase activity selects for lung adenocarcinoma stem cells dependent on notch signaling. Cancer Res. 2010;70(23):9937–9948. | ||
Lundholm L, Hååg P, Zong D, et al. Resistance to DNA-damaging treatment in non-small cell lung cancer tumor-initiating cells involves reduced DNA-PK/ATM activation and diminished cell cycle arrest. Cell Death Dis. 2013;4:e478. | ||
Yu D, Holm R, Goscinski MA, Trope CG, Nesland JM, Suo Z. Prognostic and clinicopathological significance of Cacna2d1 expression in epithelial ovarian cancers: a retrospective study. Am J Cancer Res. 2016;6(9):2088–2097. | ||
Ruan J, Liu X, Xiong X, et al. miR-107 promotes the erythroid differentiation of leukemia cells via the downregulation of Cacna2d1. Mol Med Rep. 2015;11(2):1334–1339. | ||
Kuo SS, Saad AH, Koong AC, Hahn GM, Giaccia AJ. Potassium-channel activation in response to low doses of gamma-irradiation involves reactive oxygen intermediates in nonexcitatory cells. Proc Natl Acad Sci U S A. 1993;90(3):908–912. | ||
Palme D, Misovic M, Schmid E, et al. Kv3.4 potassium channel-mediated electrosignaling controls cell cycle and survival of irradiated leukemia cells. Pflugers Arch. 2013;465(8):1209–1221. | ||
Heise N, Palme D, Misovic M, et al. Non-selective cation channel-mediated Ca2+-entry and activation of Ca2+/calmodulin-dependent kinase II contribute to G2/M cell cycle arrest and survival of irradiated leukemia cells. Cell Physiol Biochem. 2010;26(4–5):597–608. | ||
Yu J, Wang S, Zhao W, et al. Mechanistic exploration of cancer stem cell marker voltage-dependent calcium channel α2δ1 subunitmediated chemotherapy resistance in small-cell lung cancer. Clin Cancer Res. 2018;24(9):2148–2158. | ||
Li L, Hao X, Qin J, et al. Antibody against CD44s inhibits pancreatic tumor initiation and postradiation recurrence in mice. Gastroenterology. 2014;146(4):1108–1118. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.