Back to Journals » Journal of Inflammation Research » Volume 19
Brain-Region-Mediated Neuroimmune Modulation in Sepsis: Research Advances and Therapeutic Prospects
Authors Zhou X ![]() , Tang Y, Chen H, Huang W, Qiu H
, Tang Y, Chen H, Huang W, Qiu H
Received 9 October 2025
Accepted for publication 24 January 2026
Published 10 February 2026 Volume 2026:19 570541
DOI https://doi.org/10.2147/JIR.S570541
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Anh Ngo
Xuechun Zhou, Ying Tang, Hui Chen, Wei Huang, Haibo Qiu
Jiangsu Provincial Key Laboratory of Critical Care Medicine, Department of Critical Care Medicine, Zhongda Hospital, School of Medicine, Southeast University, Nanjing, Jiangsu, 210009, People’s Republic of China
Correspondence: Wei Huang, Jiangsu Provincial Key Laboratory of Critical Care Medicine, Department of Critical Care Medicine, Zhongda Hospital, School of Medicine, Southeast University, Nanjing, Jiangsu, 210009, People’s Republic of China, Email [email protected] Haibo Qiu, Jiangsu Provincial Key Laboratory of Critical Care Medicine, Department of Critical Care Medicine, Zhongda Hospital, School of Medicine, Southeast University, Nanjing, Jiangsu, 210009, People’s Republic of China, Email [email protected]
Abstract: Sepsis, a life-threatening organ dysfunction caused by a dysregulated host response to infection, lacks effective treatments targeting its pervasive immune dysregulation. Despite growing interest in neuroimmune interactions, there remains a fragmented understanding of how distinct brain regions integrate peripheral immune signals and orchestrate systemic immune responses in sepsis. Emerging evidence highlights the brain as a master regulator of systemic immunity in sepsis through the neuro-immune-endocrine network. This review summarizes recent advances in understanding how distinct brain regions, including the hypothalamus, brainstem, cortex and cerebellum, sense peripheral inflammation and feedback-regulate systemic immune responses via dedicated neural circuits, neurotransmitters, and autonomic outputs. We also highlight the therapeutic potential of neuromodulation techniques designed to target these central circuits and discuss their implications for developing future precision medicine strategies in sepsis management.
Keywords: sepsis, neuroimmunity, brain regions, central nervous system
Graphical Abstract:

Introduction
Sepsis is a life-threatening acute organ dysfunction syndrome triggered by an abnormal inflammatory response to infection.1 Sepsis has become a major challenge in global public health. Its pathophysiological hallmark is an uncontrolled inflammatory response triggered when pathogens breach the host’s defense barriers. This manifests as a complex state of simultaneous excessive inflammation and immunosuppression, persisting in a state of imbalance that ultimately leads to organ dysfunction and death.2
The central nervous system (CNS), serving as the core regulatory hub of the body, plays a pivotal regulatory role in the pathological process of sepsis through the neuro-immune-endocrine network. Recent studies indicate that vagus nerve stimulation (VNS) significantly reduces systemic inflammatory responses, multi-organ injury, and mortality in murine models of sepsis by activating the vagus nerve-cholinergic anti-inflammatory pathway.3 This finding suggests a critical regulatory role of neuroimmune mechanisms within the pathophysiology of sepsis.
While the central nervous system (CNS) is increasingly recognized as a key player in sepsis, current literature largely addresses the hypothalamus, brainstem, cortex and cerebellum in isolation. To bridge this gap, this review proposes an integrated framework centered on distinct brain nuclei, synthesizing these discrete pathways into a coherent neuroimmune network. This review will examine key regions including the hypothalamus (paraventricular nucleus, arcuate nucleus, supraoptic nucleus), brainstem (nucleus tractus solitarius, rostral ventrolateral medulla), cerebral cortex (insula, prefrontal cortex), cerebellum, and other brain nuclei (amygdala, hippocampus). In addition, we will explore how to develop neuroimmunotherapy strategies for specific brain areas in sepsis (such as targeted neuroregulation and receptor-specific drug intervention) based on the understanding of the regulatory mechanisms of different brain regions.
Methods
We conducted a comprehensive narrative review of the literature on brain-region-mediated neuroimmune modulation in sepsis. A systematic search was performed in PubMed and Web of Science databases from inception to December 2025. The search strategy combined the following key terms: “sepsis”, “neuroimmunity”, “brain region”, “central nervous system”, “hypothalamus”, “brainstem”, “cortex”, “cerebellum” and specific nuclei names. We included preclinical studies, human neuroimaging studies, and clinical trials that investigated the role of specific brain regions or neural circuits in regulating immune responses during sepsis. Studies were excluded if they (1) were not published in English, (2) did not directly address brain-immune crosstalk. The evidence was synthesized to provide a mechanistic overview and to discuss emerging therapeutic strategies.
Anatomical Localization and Function of Brain Regions Associated with Sepsis
As illustrated in Figure 1, distinct brain regions, by virtue of their specific anatomical positioning, neural circuit connectivity, and functional specialization, play indispensable and diverse roles in the pathophysiology of sepsis, collectively forming a central network within the CNS that regulates systemic immunity.
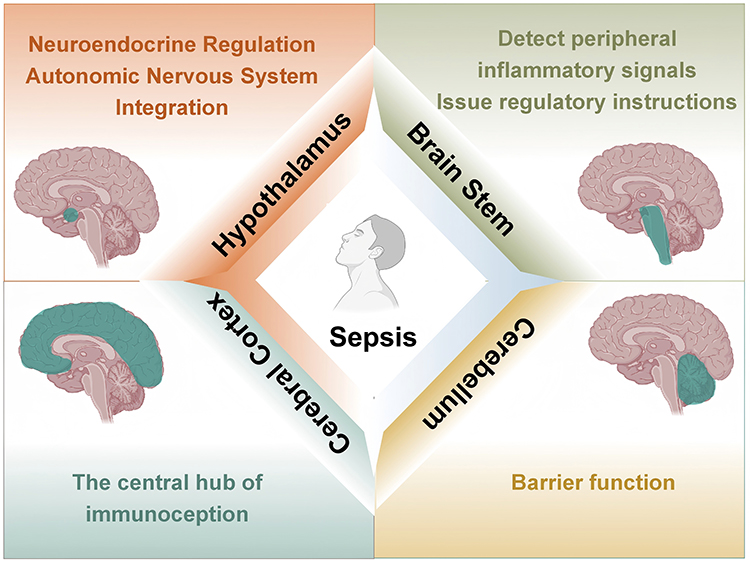 |
Figure 1 Schematic overview of the anatomical locations and proposed functional roles of key brain regions in the neuroimmune axis of sepsis. Created with BioRender.com. |
Hypothalamus
The hypothalamus functions as the central hub for neuroendocrine regulation and the integration of autonomic nervous system functions. The hypothalamic-pituitary axis integrates neural and humoral regulation.4,5 The hypothalamus is located on the ventral surface of the brain, beneath the thalamus. At least 70 distinct types of neurons exist within the hypothalamic region,6 including corticotropin-releasing hormone neurons,7 growth hormone-releasing hormone neurons,8 oxytocin neurons,9 and somatostatin neurons,10 among others. Increased adrenocorticotropic hormone (ACTH) leads to sustained activation of the hypothalamic-pituitary-adrenal (HPA) axis, impairing immune function.11 The hypothalamic nuclei most closely associated with sepsis include the paraventricular nucleus (PVN), arcuate nucleus (ARC), and supraoptic nucleus (SON).
Brainstem
Unlike the hypothalamus, which coordinates systemic response through the neuroendocrine axis, the brain stem, as the core relay and integration centre of visceral sensation and autonomous movement instructions, can convert peripheral immune information into a rapid and direct neurological response, thus instantly regulating inflammation and cardiovascular function.
The brainstem contains dense clusters of neural nuclei and nerve tracts, serving as the center for transmission and reflexes.12 Located in the anterior part of the posterior cranial fossa, it acts as the hub connecting the cerebrum, cerebellum, and spinal cord. The brainstem receives peripheral inflammatory signals,13 integrates information, and issues regulatory commands through neural and endocrine pathways.14–16 Among its regions most relevant to sepsis are the nucleus tractus solitarius (NTS) and the rostral ventrolateral medulla (RVLM).
Cortex
In contrast to the subcortical centers governing autonomic and endocrine outputs, the cerebral cortex provides higher-order regulation of the immune system. Cortical regions process the subjective and cognitive dimensions of sickness, integrate multisensory signals, and can modulate peripheral inflammation through descending pathways to hypothalamic and brainstem nuclei.
The cerebral cortex, as the higher neural center, exerts precise regulation over the immune system through distinct functional subregions.17 As the outermost layer of the brain composed of gray matter,18 different cortical areas influence the inflammatory response and immune status in sepsis via neuroendocrine, autonomic, and neuroimmune pathways.19–21 Among these, the insular cortex and prefrontal cortex (PFC) are particularly relevant to sepsis.
Cerebellum
Beyond the classical roles in motor coordination, the cerebellum is increasingly recognized as an immunomodulatory interface in sepsis. Its unique vascular architecture and connectivity allow it to sense circulating inflammatory mediators and influence neuroimmune crosstalk, often manifesting as non-motor neurological complications.
In sepsis, increased permeability of the cerebellar blood-brain barrier (BBB) permits the entry of circulating immune cells into the brain, which triggers or exacerbates glial cell activation.22,23 Wang J et al found elevated activation levels of microglia and astrocytes in the cerebellum of sepsis model mice. Protein interaction network analysis indicates that interleukin-1β (IL-1β) is a central factor in sepsis-related brain alterations.24 As a key immune signaling molecule, IL-1β may induce fever,25 promote thrombosis,26 and facilitate inflammatory cell extravasation27,28 during sepsis. Several drugs have demonstrated potential for treating sepsis-induced cerebellar injury: pregabalin partially ameliorates LPS-induced cerebellar damage in sepsis,29 while agomelatine prevents LPS-induced brain and cerebellar injury in rats.30
Immunoregulatory Mechanisms of Sepsis Involving Different Brain Nuclei
Figure 2 illustrates how the PVN, ARC, and SON, as core hypothalamic nuclei, play crucial roles in the pathological progression of sepsis through distinct neuroimmune mechanisms.
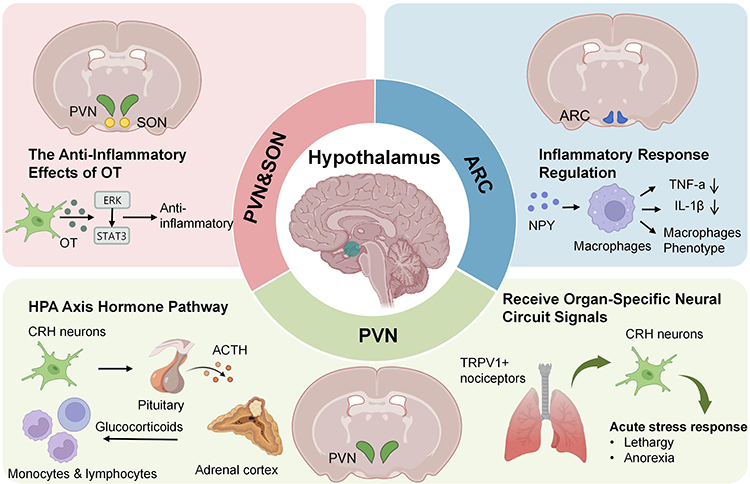 |
Figure 2 Mechanisms of hypothalamic nuclei in sepsis. The diagram summarizes the pathways mediated by the paraventricular nucleus (PVN), arcuate nucleus (ARC), and supraoptic nucleus (SON). Created with BioRender.com. Abbreviations: ACTH, Adrenocorticotropic Hormone; ARC, Arcuate Nucleus; CRH, Corticotropin-Releasing Hormone; ERK, Extracellular Regulated Protein Kinases; HPA, Hypothalamic-Pituitary-Adrenal; IL-1β, Interleukin-1β; NPY, Neuropeptide Y; OT, Oxytocin; PVN, Paraventricular Nucleus; SON, Supraoptic Nucleus; STAT3, Signal Transducer and Activator of Transcription 3; TNF-α, Tumor Necrosis Factor-α; TRPV1, Transient Receptor Potential Vanilloid1. |
PVN
As a key hypothalamic nucleus, the PVN31 regulates immune and stress responses in sepsis not only through the classical HPA axis but also via direct neural circuitry from peripheral tissues. Corticotropin-releasing hormone (CRH) neurons constitute the primary component of the parvocellular neurons in the PVN.32 These neurons regulate the body’s stress response and immune response by promoting the secretion of ACTH33 and cortisol.34 Clinical evidence indicates that plasma ACTH levels are often suppressed in patients with sepsis and are inversely correlated with disease severity.35 ACTH reaches the adrenal cortex via the bloodstream, stimulating it to release mineralocorticoids and glucocorticoids (GC).36 In the early stages of sepsis, plasma levels of ACTH and cortisol increase rapidly,37 accompanied by disruption of the body’s hormonal secretion rhythms38 and the onset of immune dysregulation.39 This suggests that CRH may be involved in the development of sepsis, and the mechanism by which CRH mediates sepsis remains to be explored.
Poller et al demonstrated that acute stress activates PVN neurons. Specifically, chemogenetic activation of PVN-CRH neurons not only elevated peripheral corticosterone levels but also significantly reduced circulating monocytes and lymphocytes. Notably, further animal experiments demonstrated that while this acute stress response confers protective effects in autoimmune disease models, it exacerbates the severity of SARS-CoV-2 and influenza virus infections.40 These findings in preclinical models elucidate the potential bidirectional regulatory function of PVN-CRH neurons across different immune contexts.
The PVN not only participates in sepsis immunity by secreting hormones through neurons but also directly regulates immunity by receiving signals from specific neural circuits. Research by Granton E et al elucidates the central role of the PVN of the hypothalamus in pulmonary infection: in a lipopolysaccharide (LPS)-induced pneumonia model, activation of TRPV1+ nociceptors in the lungs stimulates PVN CRH neurons through a specific neural circuit, triggering a classic acute stress response characterized by disease behaviors such as lethargy and anorexia, as well as thermoregulation dysregulation. The study specifically revealed that biofilm pathogens capable of producing extracellular polymeric substances (EPS) enhance pathogenicity by evading activation of this lung-brain neural pathway.41 This discovery may provide a new perspective for understanding the mechanisms of neuro-immune interactions in infectious diseases. Notably, the effects of PVN-CRH activation appear highly context-dependent; while it may confer protection in autoimmune settings, it can exacerbate outcomes in viral infections, underscoring the need to delineate its precise role across diverse sepsis etiologies before considering targeted interventions.
ARC
The ARC primarily contains neuropeptide Y (NPY) neurons, which participate in regulating appetite, energy metabolism, and bone metabolism. NPY is stored and released together with norepinephrine (NE).42 In sepsis, NPY expression increases,43 which is closely associated with the release of inflammatory mediators such as tumor necrosis factor alpha (TNF-α) and IL-1β.44 During the inflammatory phase, NPY promotes macrophage production of TNF-α by activating Y1 receptors (Y1R) while simultaneously inhibiting IL-1β release, thereby helping to regulate the inflammatory response and prevent tissue damage caused by excessive inflammation.45 NPY can also suppress interleukin-12 (IL-12) and TNF-α production in macrophages by activating Y1R, thereby converting macrophages to an M2 anti-inflammatory phenotype and reducing pro-inflammatory factor release.46 The precise mechanisms governing ARC-mediated bidirectional cytokine regulation remain unclear. Further investigation is required to determine whether NPY signaling shifts dynamically during the transition from the hyperinflammatory to the immunosuppressive phase of sepsis.
SON
Oxytocin (OT) secreted by magnocellular neurons in the SON and PVN exerts anti-inflammatory effects through multiple mechanisms: On one hand, oxytocin directly inhibits the release of proinflammatory cytokines and modulates immune responses via the hypothalamic-pituitary-portal system.47 On the other hand, it reduces levels of inflammatory mediators such as TNF-α, IL-6, and IL-1β by suppressing the shift of macrophages from a resting state to a proinflammatory phenotype, thereby mitigating organ damage caused by sepsis.48 In the pathophysiology of sepsis-associated encephalopathy (SAE), oxytocin activates the oxytocin receptor (OXTR), thereby modulating the ERK/STAT3 signaling pathway. This effectively suppresses abnormal activation of microglia and neuroinflammation, thus protecting synaptic plasticity in the hippocampus and improving cognitive dysfunction.49 It is noteworthy that despite basic research having clearly demonstrated oxytocin’s significant organ-protective efficacy, its translation into clinical applications still requires careful and comprehensive evaluation. Particularly in perinatal patients, strict and dynamic monitoring of fluid balance must be conducted during clinical administration to avoid adverse reactions.
The NTS and RVLM, as core nuclei regulating sepsis in the brainstem, play pivotal roles in systemic inflammation and organ injury by integrating the vagus nerve anti-inflammatory reflex and mediating sympathetic nervous system excitation, respectively, as depicted in Figure 3.
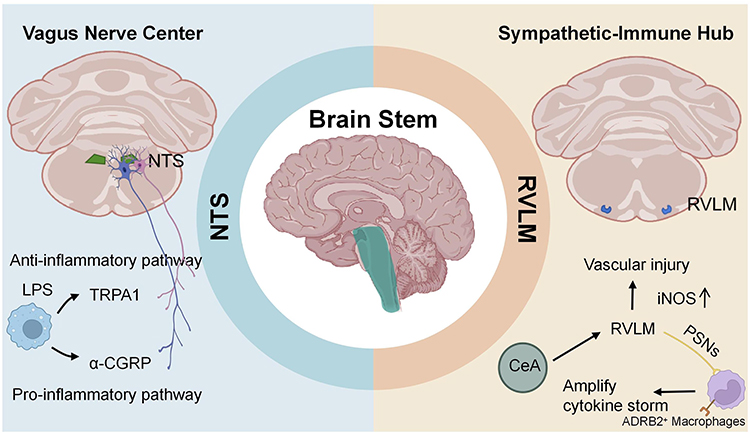 |
Figure 3 Mechanisms of brainstem nuclei in sepsis. The diagram highlights the roles of the nucleus tractus solitarius (NTS) and the rostral ventrolateral medulla (RVLM). Created with BioRender.com. Abbreviations: a-CGRP, alpha-Calcitonin Gene - Related Peptide; ADRB2, β2-adrenergic receptors; CeA, Central Amygdala; iNOS, Inducible Nitric Oxide Synthase; LPS, Lipopolysaccharide; NTS, Nucleus Tractus Solitarius; PSNs, Pulmonary Sympathetic Nerves; RVLM, Rostral Ventrolateral Medulla; TRPA1, Transient Receptor Potential Channel A1. |
NTS
The NTS serves as the primary projection target and integration center for the central axons of the vagus nerve.50 Following intraperitoneal injection of LPS in mice, cytokines activate distinct vagal sensory neuron populations to transmit inflammatory status to the brain. These signals travel along two distinct pathways: one carries anti-inflammatory signals by activating the transient receptor potential channel A1 (TRPA1) and acts on NTS neurons to enhance the anti-inflammatory response. Conversely, another pathway responds to proinflammatory signals by activating calcitonin gene-related peptide alpha (α-CGRP) and contributes to reducing proinflammatory responses. Both pathways activate distinct populations of vagal ganglion neurons, ultimately converging in the NTS.51 In sepsis, the number of activated microglia in the NTS increases, and elevated levels of interleukin-1β induce hypotension and tachycardia, ultimately leading to vascular injury.52 In a cecal ligation and puncture (CLP) model, activation of central adenosine A1 receptors (A1AR) in the NTS promotes cholinergic antagonism of cardiac and neuroinflammatory responses associated with sepsis and related cardiomyopathies and neuropathologies.53
RVLM
The RVLM is a key nucleus in sepsis-associated pneumonia and vascular injury.54 Numerous studies indicate that the upregulation of inducible nitric oxide synthase (iNOS) in the RVLM during sepsis is a key contributor to vascular injury.55,56 Dexmedetomidine may improve sepsis by inhibiting sympathetic activation,57 whereas morphine exacerbates sepsis-induced cardiovascular and autonomic dysfunction through μ-opioid receptor-mediated mechanisms, while also amplifying central inflammatory, chemotactic, and oxidative signaling.58 Research by Li W et al has revealed the critical regulatory role of the RVLM in pneumonia-associated cytokine storms. Specifically, their work demonstrates that GABAergic neurons in the central amygdala (CeA) activate β2-adrenergic receptors (ADRB2) on interstitial macrophages (Mac2) via the CeA-RVLM-pulmonary sympathetic nerves (PSNs) pathway, thereby amplifying the cytokine storm.59 Targeted inhibition of this pathway has been shown to improve prognosis in animal models, suggesting a potential therapeutic strategy for pneumonia. However, the significance of this pathway in human polymicrobial sepsis remains to be established, and therapeutic targeting of the RVLM requires balancing its dual role in driving harmful inflammation and maintaining essential sympathetic vascular tone.
As summarized in Figure 4, the insula acts as the hub of “immunoception”, and the prefrontal cortex (PFC) is the primary target for cognitive impairment, reflecting their critical roles within the cerebral cortex during sepsis.
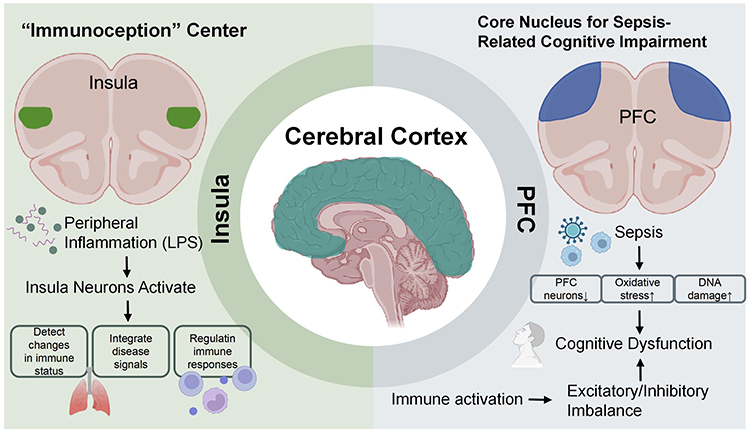 |
Figure 4 Mechanisms of cerebral cortex nuclei in sepsis, focusing on the insula and prefrontal cortex (PFC). Created with BioRender.com. Abbreviations: LPS, Lipopolysaccharide; PFC, Prefrontal Cortex. |
Insula
As the central hub of “immunoception”, the insula participates in regulating neuroinflammation and systemic immune homeostasis during sepsis by detecting changes in immune status, integrating disease signals, and modulating peripheral immune responses. Asya Rolls et al found that neurons distributed in the mouse insula activate during inflammatory states, thereby regulating peripheral immune responses.60 MRI scans of the brains of sepsis patients reveal hyperactivation of the insular cortex. During LPS-induced inflammatory responses, the insula participates in the perception and integration of disease symptoms such as physical discomfort.61 Although direct studies on the role of the insula in neuroimmunoregulation during sepsis are currently limited, existing neuroimmunological research and neuroinflammatory mechanisms associated with sepsis suggest that the insula may influence the immune response to sepsis by regulating “immunoreception” and neuroinflammatory pathways. It remains unclear whether insular activity primarily serves as an afferent function for inflammation perception in sepsis or also directly participates in efferent commands for immune regulation. Future research should integrate functional neuroimaging with immune profiling analysis to elucidate this critical mechanism in clinical patients.
PFC
The PFC experiences a disruption in excitatory/inhibitory balance due to neuroinflammation during sepsis, leading to cognitive dysfunction. Research indicates that sepsis can lead to a reduction in the number of PFC neurons, increased oxidative stress, and oxidative DNA damage.62 Additionally, sepsis causes a significant increase in proinflammatory factors (such as TNF-α and IL-1β) in the PFC, which are key contributors to neuroinflammation and cognitive impairment.63 Research by Mittli et al indicates that systemic immune challenges affect the function of PFC neurons through multiple molecular and cellular mechanisms, including increased intrinsic excitability of pyramidal cells, while inhibitory interneurons show no significant changes. Additionally, immune activation affects the synaptic networks of the PFC, leading to increased excitatory synaptic currents and decreased inhibitory synaptic currents, thereby triggering an excitatory/inhibitory imbalance in the PFC. This imbalance may correlate with the activation of microglia, subsequently further exacerbating neuroinflammation.64 Multiple drugs and therapies have been shown to modulate the neuroimmune response in the PFC. For example, pleiotrophin (PTN) has been demonstrated to regulate glial cell-mediated neuroinflammation;65 Berberine inhibits NLRP3 inflammasome-mediated neuroinflammation in mice.66 While these findings illuminate potential mechanisms of cognitive dysfunction, it remains unclear whether the observed excitatory/inhibitory imbalance is a cause or a consequence of sustained neuroinflammation. Moreover, the efficacy of preclinical interventions in reversing long-term cognitive deficits in human sepsis survivors represents a major translational gap that future clinical trials need to address.
In addition to the nuclei within the aforementioned brain regions, the amygdala and hippocampus are also two key nuclei in the neuroimmunoregulation of sepsis. Their functions as the “two-way control hub” for neuroinflammation and the primary target site for cognitive impairment are shown in Figure 5.
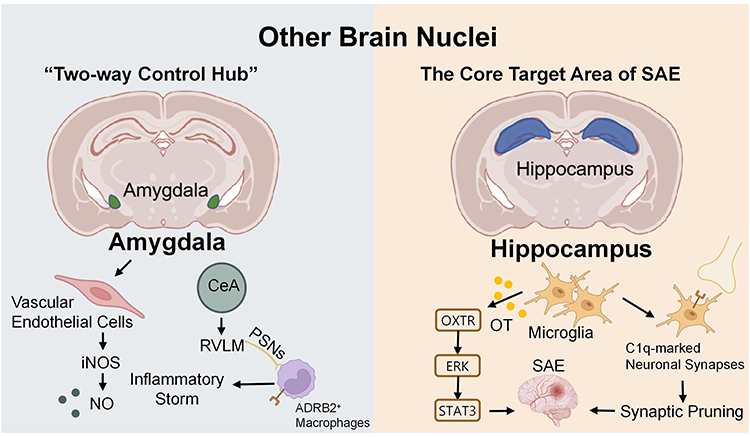 |
Figure 5 Mechanisms of the amygdala and hippocampus in sepsis. Created with BioRender.com. Abbreviations: ADRB2, β2-adrenergic receptors; CeA, Central Amygdala; ERK, Extracellular Regulated Protein Kinases; iNOS, Inducible Nitric Oxide Synthase; NO, Nitric Oxide; OT, Oxytocin; OXTR, Oxytocin Activates The Oxytocin Receptor; PSNs, Pulmonary Sympathetic Nerves; RVLM, Rostral Ventrolateral Medulla; SAE, Sepsis-Associated Encephalopathy; STAT3, Signal Transducer and Activator of Transcription 3. |
Amygdala
The amygdala, as a core brain region for emotional regulation and stress responses, serves as a bidirectional regulatory hub in the neuroimmune dysregulation triggered by sepsis. It is activated by sepsis-related immune signals while simultaneously modulating peripheral and central immune responses through neural pathways, ultimately influencing the progression and prognosis of sepsis. In septic shock, there is minimal activation of microglia and glial expression of TNF-α within the amygdala, while vascular endothelial cells exhibit a significant increase in inducible nitric oxide synthase (iNOS) expression.67 Subsequent studies have revealed that nitric oxide (NO) produced by iNOS in the amygdala participates in the central nervous system’s central response to immune stimuli during sepsis.68 The amygdala also plays a key role in driving peripheral inflammatory responses through neural circuits. As detailed in the RVLM section above, inhibitory GABAergic neurons in the central amygdala (CeA) can initiate a CeA→RVLM→pulmonary sympathetic nerve pathway that ultimately exacerbates lung inflammation via adrenergic signaling to macrophages.59 This highlights the amygdala’s role as a higher-order regulator capable of amplifying peripheral cytokine storms through well-defined brainstem and sympathetic efferents.
Hippocampus
The hippocampus is the core target region of SAE.69 SAE is one of the most common and severe complications of sepsis,70 while neuroinflammation represents the most critical pathological basis for SAE-related cognitive impairment.71 Microglia play a crucial role in the development and progression of neuroinflammation.72 Oxytocin in microglia exerts anti-inflammatory and neuroprotective effects, improving SAE outcomes through the OXTR-ERK-STAT3 signaling pathway.49 Microglia can also phagocytose C1q-marked neuronal synapses. Intrahippocampal injection of C1q-blocking antibodies reduces synaptic pruning following the acute phase of sepsis-associated encephalopathy, thereby preventing the development of neurocognitive deficits.73
Neuroimmunotherapeutic Strategies Targeting Different Brain Regions in Sepsis
Various drugs have been demonstrated to effectively intervene in the pathological progression of sepsis by targeting neuroimmune regulatory mechanisms in different brain regions (Table 1).
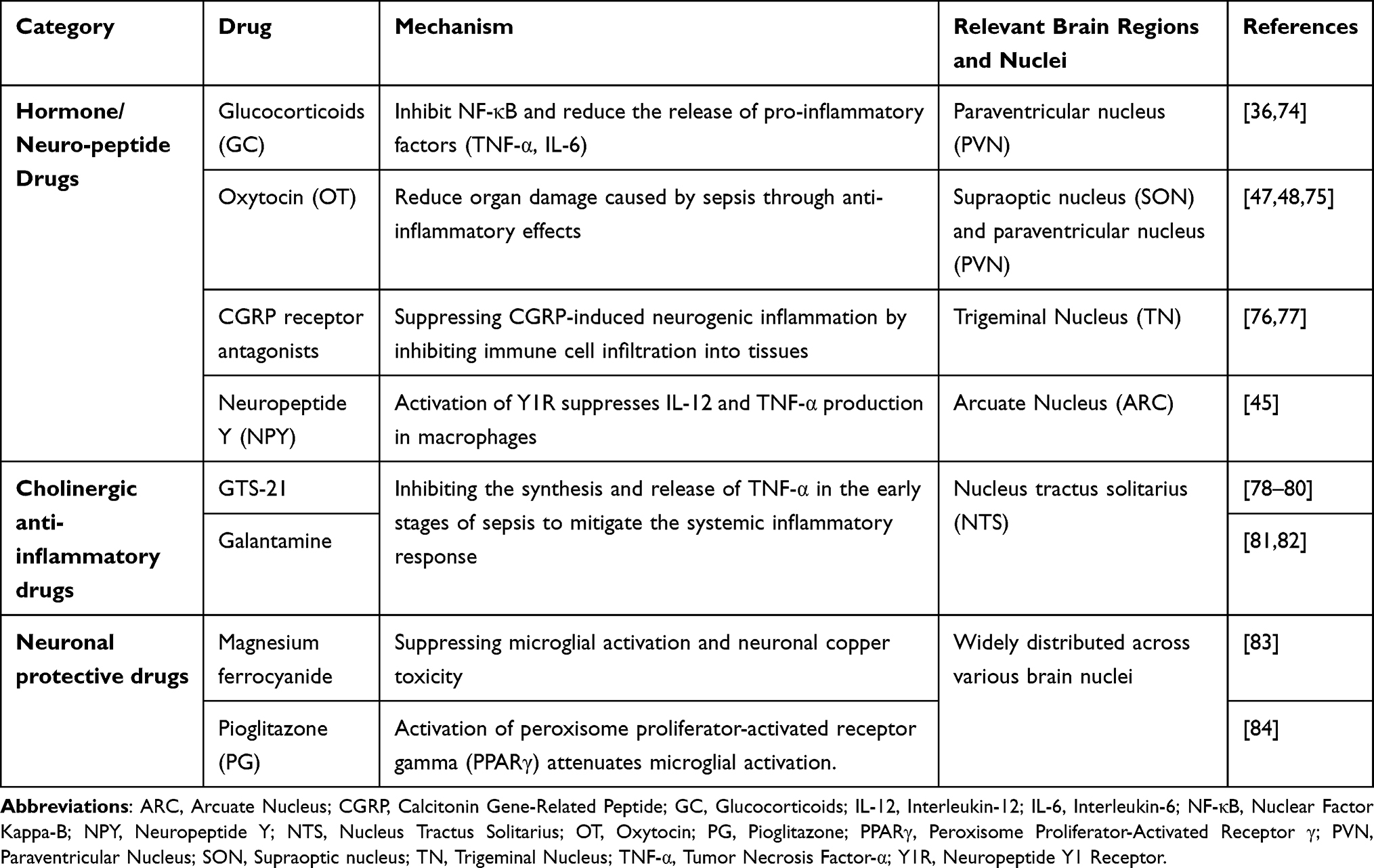 |
Table 1 Mechanisms of Neuroimmunomodulatory Drugs in Sepsis and Associated Brain Nuclei |
Hormone/Neuropeptide Drugs
The hypothalamus profoundly influences the imbalance between systemic inflammatory response syndrome (SIRS)85 and compensatory anti-inflammatory response syndrome (CARS)86 in sepsis by regulating the HPA axis and multiple neuroendocrine pathways. Under septic stress, the PVN releases CRH.68 Subsequently, the HPA axis cascade reaction is initiated: CRH stimulates the pituitary gland to secrete ACTH, ultimately triggering the adrenal glands to release GC. According to the 2024 updated SCCM guidelines on GC application in sepsis, short-term administration can suppress Nuclear Factor Kappa-B (NF-κB) and reduce the release of proinflammatory factors (TNF-α, IL-6), but long-term excessive activation may lead to immunosuppression. Individualized treatment should be selected based on the stage of sepsis to achieve precise intervention.74
Oxytocin secreted by magnocellular neurons in the SON and PVN mitigates sepsis-induced organ damage through anti-inflammatory effects.47,48,75 The trigeminal nucleus (TN) releases calcitonin gene-related peptide (CGRP), which promotes a massive infiltration of immune cells into tissues, thereby inducing neurogenic inflammation.76 CGRP receptor antagonists (such as erenumab) can suppress neurogenic inflammation.77 ARC-mediated secretion of NPY suppresses IL-12 and TNF-α production in macrophages by activating Y1R, thereby converting macrophages to an M2 anti-inflammatory phenotype and reducing proinflammatory factor release.49 These preclinical findings suggest a potential basis and intervention targets for personalized precision treatment of sepsis.
Cholinergic Anti-Inflammatory Drugs
The cholinergic anti-inflammatory pathway forms a critical neuroimmune circuit in sepsis. In this reflex arc, peripheral inflammation is detected by vagal afferent fibers, which transmit signals to the NTS in the brainstem.87 The NTS serves as the integration center; upon activation, it stimulates vagal efferent fibers to release acetylcholine. Acetylcholine then acts on α7 nicotinic acetylcholine receptors (α7nAChR) in peripheral tissues, inhibiting the synthesis and release of pro-inflammatory cytokines such as TNF-α and thereby mitigating the systemic inflammatory response.88–91 Conversely, excessive sympathetic nervous system activation can inhibit this cholinergic pathway, exacerbating inflammatory dysregulation and organ dysfunction.92 Targeting this axis, α7nAChR agonists such as GTS-2178–80 and galantamine81,82 have demonstrated protective effects in sepsis models, highlighting the therapeutic potential of modulating this “neuroimmune hub”.
Neuroprotective Agents
Microglia and astrocytes are widely distributed across various brain regions of the central nervous system, playing crucial roles in maintaining brain homeostasis and neuroimmune function.93,94 In sepsis, microglia and astrocytes undergo a phenotypic shift from an anti-inflammatory M2 state to a pro-inflammatory M1 phenotype, leading to neuronal damage.95 In rodent models, magnesium ferrocyanide has been shown to alleviate sepsis-associated encephalopathy by inhibiting microglial activation and neuronal copper toxicity.83 Pioglitazone (PG) activates peroxisome proliferator-activated receptor gamma (PPARγ), attenuates microglial activation, enhances the survival of dopaminergic neurons in the nigrostriatal system, and exerts neuroprotective effects against sepsis-induced brain injury.84 These findings suggest potential directions for clinical treatment, and targeting the regulation of glial cell polarization represents a promising avenue for future investigation.
Vagus Nerve Stimulation
The core pathology of sepsis is the coexistence of an uncontrolled systemic inflammatory response and immunosuppression.96 Traditional anti-inflammatory drug strategies are often accompanied by immunosuppression or organ toxicity due to their broad mechanism of action. In recent years, advances in the “neuro-immune” network research have enabled the regulation of the body’s innate immune system through precise neural stimulation.
The afferent fibers of the vagus nerve originate from visceral organs and project to the NTS in the medulla oblongata, where they contribute to innate defense against inflammation through the cholinergic anti-inflammatory pathway (CAIP).97 Both pharmacological and electrical stimulation of the vagus nerve to activate the CAIP have demonstrated significant efficacy in mitigating cytokine storms.98,99 Vagus nerve stimulation (VNS) can provide more precise and targeted stimulation.100–102 A novel integrated strategy combining oral administration with ultrasound has been proposed and validated in the latest research: After ingesting capsaicin-coated piezoelectric nanoparticles, a portable low-intensity pulsed ultrasound device generates microcurrents in situ within the gastric wall, noninvasively activating the vagus nerve-cholinergic anti-inflammatory pathway. This approach significantly reduces systemic inflammation, organ damage, and mortality in septic mice, offering a safe and accessible new solution for bedside or home-based neuroimmunomodulation,3 as illustrated in Figure 6. Mughrabi et al implanted miniature “electrochemical probes” into the spleens of live mice: Real-time monitoring of NE release via fast-scan cyclic voltammetry (FSCV) provides instantaneous feedback on vagus nerve, greater splanchnic nerve, or splenic nerve stimulation intensity within seconds. This response negatively correlates with subsequent TNF-α suppression effects. By integrating splenic voltammetric electrodes with implantable stimulators, this approach delivers an immediate, quantitative “physiological benchmark” for neuroimmunoregulation.103
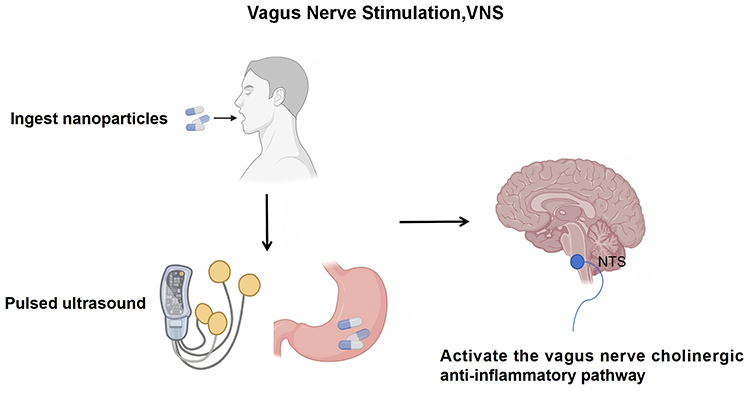 |
Figure 6 Diagram of a non‑invasive vagus nerve stimulation strategy for sepsis. Created with BioRender.com. Abbreviations: NTS, Nucleus Tractus Solitarius; VNS, Vagus Nerve Stimulation. |
Clinical studies have demonstrated that VNS can significantly improve the sepsis-induced coagulopathy (SIC) score at 24 hours in patients with sepsis-induced coagulopathy (SIC).104 The world’s first neuroimmunomodulatory device approved for immune disorders-SetPoint Medical’s vagus nerve stimulation device-has received official approval from the US Food and Drug Administration (FDA) for treating rheumatoid arthritis (FDA Premarket Approval P240039),105 marking the transition of neuroimmunomodulation from experimental research to clinical application. The translation of invasive VNS to septic patients requires navigating complex ethical and practical landscapes, including conducting rigorous risk-benefit analyses for critically ill patients, navigating the complexities of informed consent in this vulnerable population, and ensuring long-term safety and efficacy.
Clinical Implications and Translational Significance of Neuroimmunity in Sepsis
The clinical relevance of neuroimmune crosstalk is profound within the intensive care setting. Sepsis-associated encephalopathy (SAE), characterised by delirium and cognitive disorders, is a prominent clinical manifestation of neuroinflammation with direct prognostic significance.106 Similarly, although Sepsis-induced ARDS is fundamentally derived from peripheral inflammation and endothelial damage,107 its pathophysiology may be significantly modulated by sympathetic-vagus nerve imbalances involving key brain stem nuclei (such as RVLM and NTS). Furthermore, this central autonomic dysregulation extends to the cardiovascular system, manifesting as sepsis-induced cardiac dysfunction (SICD). Here, aberrant sympathetic outflow and the consequent catecholamine surge can induce cardiomyocyte toxicity and myocardial depression, highlighting a pathological brain-heart axis driven by neuroimmune disturbances.108
Importantly, these central neuroimmune pathways exert profound influence over the peripheral vasculature. Endothelial injury,109 immunothrombosis,110 and microcirculatory dysfunction111 are well-established determinants of organ failure in sepsis, with sepsis-induced acute kidney injury serving as a prime example, driven by inflammatory and oxidative stress pathways as well as dysregulated innate immune signaling.112,113 Each of these processes is susceptible to modulation by neurogenic signals, as shown by improvements in oxygenation and microcirculatory parameters with iloprost treatment in COVID-19-related ARDS, which provides an important translational bridge highlighting the interplay between central neuroimmune regulation and peripheral endothelial function.114
Therefore, sepsis should be conceptualized as a syndrome encompassing central regulatory dysfunction, endothelial injury, and peripheral organ damage. This comprehensive view emphasises the therapeutic potential of neuromodulatory strategies aimed at rebalancing autonomic output. For example, VNS can not only reduce systemic inflammation, but also reduce secondary microcirculation disorders, serving as an adjunct to conventional organ support therapy in the treatment of sepsis. However, it is critical to acknowledge the models from which these mechanisms are derived. Such models often utilize homogeneous populations under controlled conditions, which may not fully capture the immense heterogeneity in etiology, comorbidities, and treatment responses seen in human sepsis. Notably, current mechanistic understanding relies heavily on preclinical models. Direct clinical evidence linking brain regional activity to immune outcomes in sepsis remains sparse. Therefore, future research must integrate multi-modal methods, such as advanced neuroimaging, continuous autonomic monitoring and deep immune phenotypes, to verify these pathways, reveal potential substitution or competition mechanisms, and finally determine clinically feasible phenotypes. Integrating perspectives and addressing challenges from neurological critical care is crucial to transforming these promising mechanisms into effective treatment. This gap underscores the significant translational journey required to move from promising experimental neuromodulation to safe, effective, and widely applicable clinical therapies.
Conclusion
The central nervous system (CNS) serves as the core hub of the immune regulatory network in sepsis, dynamically coordinating systemic inflammatory and immune responses through multi-regional and multi-level pathways. In recent years, therapeutic strategies based on neuroimmune mechanisms have achieved significant breakthroughs. From drug design targeting specific receptors (such as α7nAChR, CGRP, and NPY) to non-invasive neuromodulation technologies, these approaches have demonstrated promising translational potential.
To realize this potential, the field must now address key translational challenges. At a fundamental level, major scientific questions remain unanswered, particularly regarding how to achieve precise spatiotemporal control over complex neuroimmune circuits to ensure pathway specificity without systemic side effects. Moreover, successful translation requires overcoming patient heterogeneity (driven by genetics, comorbidities, and sepsis etiology) to enable personalized targeting, as well as defining the optimal timing for intervention across the dynamic stages of sepsis. Finally, rigorously evaluating the safety profiles of neuromodulatory devices and validating predictive biomarkers remain indispensable for guiding clinical therapy.
Collectively, these insights pave the way toward a precision medicine approach in sepsis, centered on aligning brain-region-specific interventions with individualized neuroimmune signatures. Translating this paradigm into clinical practice will require defining clinically actionable phenotypes based on autonomic function, central inflammatory markers, or other biomarker profiles. Future research would benefit from focusing on identifying specific patient subgroups, such as those with autonomic dysregulation, high risk of SAE, or distinct biomarker profiles, which could indicate greater responsiveness to neuroimmune-targeted therapies. Clinical evaluation of neuromodulatory approaches like vagus nerve stimulation should advance through structured study phases, progressing from mechanistic validation to controlled trials in sepsis cohorts. Furthermore, the development of integrated biomarker strategies, including neuroendocrine, autonomic, and inflammatory signatures, will be important for guiding patient selection and monitoring treatment responses in a more individualized manner. With the advancement of research methodologies and technological platforms, neuroimmunomodulatory strategies hold significant translational promise and could contribute to more targeted approaches in the clinical treatment of sepsis.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this study.
Author Contributions
Xuechun Zhou: Writing-review & editing, Writing – original draft, Investigation. Ying Tang: Writing-review & editing, Investigation. Hui Chen: Writing – original draft, Investigation. Wei Huang: Supervision, Writing – Review & Editing, Funding Acquisition. Haibo Qiu: Supervision, Project Administration, Writing – Review & Editing, Funding Acquisition. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by National Science and Technology Major Project (2022YFC2304600), the Noncommunicable Chronic Diseases-National Science and Technology Major Project (2023ZD0506500), Jiangsu Province Science and Technology Plan Project “Provincial Frontier Technology R&D Program” (BF2024054), National Natural Science Foundation of China (No. 82202390), Jiangsu Provincial Double-Innovation Doctor Program (JSSCBS20220106) and the Fundamental Research Funds for the Central Universities and Zhishan Scholars Programs of Southeast University (2242023R40034).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rudd KE, Johnson SC, Agesa KM, et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: analysis for the global burden of disease study. Lancet. 2020;395(10219):200–16. doi:10.1016/s0140-6736(19)32989-7
2. Cao M, Wang G, Xie J. Immune dysregulation in sepsis: experiences, lessons and perspectives. Cell Death Discov. 2023;9(1):465. doi:10.1038/s41420-023-01766-7
3. Mac CH, Nguyen GLT, Nguyen DTM, et al. Noninvasive vagus nerve electrical stimulation for immune modulation in sepsis therapy. J Am Chem Soc. 2025;147(10):8406–8421. doi:10.1021/jacs.4c16367
4. Jagot F, Gaston-Breton R, Choi AJ, et al. The parabrachial nucleus elicits a vigorous corticosterone feedback response to the pro-inflammatory cytokine IL-1β. Neuron. 2023;111(15):2367–2382.e6. doi:10.1016/j.neuron.2023.05.009
5. Singh U, Jiang J, Saito K, et al. Neuroanatomical organization and functional roles of PVN MC4R pathways in physiological and behavioral regulations. Mol Metab. 2022;55:101401. doi:10.1016/j.molmet.2021.101401
6. Moffitt JR, Bambah-Mukku D, Eichhorn SW, et al. Molecular, spatial, and functional single-cell profiling of the hypothalamic preoptic region. Science. 2018;362(6416). doi:10.1126/science.aau5324
7. Horvath K, Juhasz B, Kuti D, Ferenczi S, Kovacs KJ. Recruitment of corticotropin-releasing hormone (CRH) neurons in categorically distinct stress reactions in the mouse brain. Int J Mol Sci. 2023;24(14):11736. doi:10.3390/ijms241411736
8. Gusmao DO, de Sousa ME, de Sousa LMM, et al. GH-releasing hormone neurons regulate the hypothalamic-pituitary-somatotropic axis via short-loop negative feedback. Endocrinology. 2025;166(5). doi:10.1210/endocr/bqaf062
9. Li H, Jiang T, An S, et al. Single-neuron projectomes of mouse paraventricular hypothalamic nucleus oxytocin neurons reveal mutually exclusive projection patterns. Neuron. 2024;112(7):1081–1099.e7. doi:10.1016/j.neuron.2023.12.022
10. Tossell K, Yu X, Giannos P, et al. Somatostatin neurons in prefrontal cortex initiate sleep-preparatory behavior and sleep via the preoptic and lateral hypothalamus. Nat Neurosci. 2023;26(10):1805–1819. doi:10.1038/s41593-023-01430-4
11. Reiche EM, Nunes SO, Morimoto HK. Stress, depression, the immune system, and cancer. Lancet Oncol. 2004;5(10):617–625. doi:10.1016/s1470-2045(04)01597-9
12. Li F, Jiang H, Shen X, et al. Sneezing reflex is mediated by a peptidergic pathway from nose to brainstem. Cell. 2021;184(14):3762–3773.e10. doi:10.1016/j.cell.2021.05.017
13. Koopman FA, Chavan SS, Miljko S, et al. Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. Proc Natl Acad Sci U S A. 2016;113(29):8284–8289. doi:10.1073/pnas.1605635113
14. Zou N, Zhou Q, Zhang Y, et al. Transcutaneous auricular vagus nerve stimulation as a novel therapy connecting the central and peripheral systems: a review. Int J Surg. 2024;110(8):4993–5006. doi:10.1097/js9.0000000000001592
15. Bonaz B, Sinniger V, Pellissier S. Anti-inflammatory properties of the vagus nerve: potential therapeutic implications of vagus nerve stimulation. J Physiol. 2016;594(20):5781–5790. doi:10.1113/jp271539
16. Tanaka S, Abe C, Abbott SBG, et al. Vagus nerve stimulation activates two distinct neuroimmune circuits converging in the spleen to protect mice from kidney injury. Proc Natl Acad Sci U S A. 2021;118(12). doi:10.1073/pnas.2021758118
17. Steyert MR, Li T, Piao X, Nowakowski TJ. Molecular signatures in cortical development. Annu Rev Neurosci. 2025;48(1):445–464. doi:10.1146/annurev-neuro-091823-014001
18. Fukutomi H, Glasser MF, Zhang H, et al. Neurite imaging reveals microstructural variations in human cerebral cortical gray matter. Neuroimage. 2018;182:488–499. doi:10.1016/j.neuroimage.2018.02.017
19. Rolls A. Immunoception: the insular cortex perspective. Cell Mol Immunol. 2023;20(11):1270–1276. doi:10.1038/s41423-023-01051-8
20. Kayyal H, Cruciani F, Chandran SK, et al. Retrieval of conditioned immune response in male mice is mediated by an anterior-posterior insula circuit. Nat Neurosci. 2025;28(3):589–601. doi:10.1038/s41593-024-01864-4
21. Ye M, Zhu H, Lu X, et al. Central innate immunization induces tolerance against post-traumatic stress disorder-like behavior and neuroinflammatory responses in male mice. Brain Behav Immun. 2024;122:368–387. doi:10.1016/j.bbi.2024.08.049
22. Barichello T, Generoso JS, Singer M, Dal-Pizzol F. Biomarkers for sepsis: more than just fever and leukocytosis-a narrative review. Crit Care. 2022;26(1):14. doi:10.1186/s13054-021-03862-5
23. Gao Q, Hernandes MS. Sepsis-associated encephalopathy and blood-brain barrier dysfunction. Inflammation. 2021;44(6):2143–2150. doi:10.1007/s10753-021-01501-3
24. Wang J, Zhong Z, Luo H, et al. Modulation of brain immune microenvironment and cellular dynamics in systemic inflammation. Theranostics. 2025;15(11):5153–5171. doi:10.7150/thno.107061
25. Kozak W, Kluger MJ, Soszynski D, et al. IL-6 and IL-1 beta in fever. Studies using cytokine-deficient (knockout) mice. Ann N Y Acad Sci. 1998;856:33–47. doi:10.1111/j.1749-6632.1998.tb08310.x
26. Borges-Rodriguez M, Shields CA, Travis OK, et al. Platelet inhibition prevents NLRP3 inflammasome activation and sepsis-induced kidney injury. Int J Mol Sci. 2021;22(19):10330. doi:10.3390/ijms221910330
27. Xiong S, Hong Z, Huang LS, et al. IL-1beta suppression of VE-cadherin transcription underlies sepsis-induced inflammatory lung injury. J Clin Invest. 2020;130(7):3684–3698. doi:10.1172/jci136908
28. Busch K, Kny M, Huang N, et al. Inhibition of the NLRP3/IL-1beta axis protects against sepsis-induced cardiomyopathy. J Cachexia Sarcopenia Muscle. 2021;12(6):1653–1668. doi:10.1002/jcsm.12763
29. Aslankoc R, Savran M, Ozmen O, Asci S. Hippocampus and cerebellum damage in sepsis induced by lipopolysaccharide in aged rats - Pregabalin can prevent damage. Biomed Pharmacother. 2018;108:1384–1392. doi:10.1016/j.biopha.2018.09.162
30. Savran M, Aslankoc R, Ozmen O, et al. Agomelatine could prevent brain and cerebellum injury against LPS-induced neuroinflammation in rats. Cytokine. 2020;127:154957. doi:10.1016/j.cyto.2019.154957
31. Papazoglou I, Lee JH, Cui Z, et al. A distinct hypothalamus-to-beta cell circuit modulates insulin secretion. Cell Metab. 2022;34(2):285–298.e7. doi:10.1016/j.cmet.2021.12.020
32. Rasiah NP, Loewen SP, Bains JS. Windows into stress: a glimpse at emerging roles for CRH(PVN) neurons. Physiol Rev. 2023;103(2):1667–1691. doi:10.1152/physrev.00056.2021
33. Pondeljak N, Lugović-Mihić L. Stress-induced interaction of skin immune cells, hormones, and neurotransmitters. Clin Ther. 2020;42(5):757–770. doi:10.1016/j.clinthera.2020.03.008
34. Cuberos Paredes E, Goyes D, Mak S, et al. Transcutaneous auricular vagus nerve stimulation inhibits mental stress-induced cortisol release-potential implications for inflammatory conditions. Physiol Rep. 2025;13(3):e70251. doi:10.14814/phy2.70251
35. Jacobs A, Derese I, Vander Perre S, et al. Dynamics and prognostic value of the hypothalamus-pituitary-adrenal axis responses to pediatric critical illness and association with corticosteroid treatment: a prospective observational study. Intensive Care Med. 2020;46(1):70–81. doi:10.1007/s00134-019-05854-0
36. Lewis A, Thant AA, Aslam A, Aung PPM, Azmi S. Diagnosis and management of adrenal insufficiency. Clin Med. 2023;23(2):115–118. doi:10.7861/clinmed.2023-0067
37. Fredrick FC, Meda AKR, Singh B, Jain R. Critical illness-related corticosteroid insufficiency: latest pathophysiology and management guidelines. Acute Crit Care. 2024;39(3):331–340. doi:10.4266/acc.2024.00647
38. Ramsay SD, Nenke MA, Meyer EJ, Torpy DJ, Young RL. Unveiling the novel role of circadian rhythms in sepsis and septic shock: unexplored implications for chronotherapy. Front Endocrinol. 2025;16:1508848. doi:10.3389/fendo.2025.1508848
39. Beltran-Garcia J, Osca-Verdegal R, Javega B, et al. Characterization of early peripheral immune responses in patients with sepsis and septic shock. Biomedicines. 2022;10(3):525. doi:10.3390/biomedicines10030525
40. Poller WC, Downey J, Mooslechner AA, et al. Brain motor and fear circuits regulate leukocytes during acute stress. Nature. 2022;607(7919):578–584. doi:10.1038/s41586-022-04890-z
41. Granton E, Brown L, Defaye M, et al. Biofilm exopolysaccharides alter sensory-neuron-mediated sickness during lung infection. Cell. 2024;187(8):1874–1888.e14. doi:10.1016/j.cell.2024.03.001
42. Warner MR, Senanayake PD, Ferrario CM, Levy MN. Sympathetic stimulation-evoked overflow of norepinephrine and neuropeptide Y from the heart. Circ Res. 1991;69(2):455–465. doi:10.1161/01.res.69.2.455
43. Sankar V, Webster NR. Clinical application of sepsis biomarkers. J Anesth. 2013;27(2):269–283. doi:10.1007/s00540-012-1502-7
44. Oztas B, Sahin D, Kir H, et al. The effect of leptin, ghrelin, and neuropeptide-Y on serum Tnf-Alpha, Il-1beta, Il-6, Fgf-2, galanin levels and oxidative stress in an experimental generalized convulsive seizure model. Neuropeptides. 2017;61:31–37. doi:10.1016/j.npep.2016.08.002
45. Xing L, Chen B, Qin Y, et al. The role of neuropeptides in cutaneous wound healing: a focus on mechanisms and neuropeptide-derived treatments. Front Bioeng Biotechnol. 2024;12:1494865. doi:10.3389/fbioe.2024.1494865
46. Chen WC, Liu YB, Liu WF, Zhou YY, He HF, Lin S. Neuropeptide Y is an immunomodulatory factor: direct and indirect. Front Immunol. 2020;11:580378. doi:10.3389/fimmu.2020.580378
47. Schiller M, Ben-Shaanan TL, Rolls A. Neuronal regulation of immunity: why, how and where? Nat Rev Immunol. 2021;21(1):20–36. doi:10.1038/s41577-020-0387-1
48. Mehdi SF, Pusapati S, Khenhrani RR, et al. Oxytocin and related peptide hormones: candidate anti-inflammatory therapy in early stages of sepsis. Front Immunol. 2022;13:864007. doi:10.3389/fimmu.2022.864007
49. Jiang J, Zou Y, Xie C, et al. Oxytocin alleviates cognitive and memory impairments by decreasing hippocampal microglial activation and synaptic defects via OXTR/ERK/STAT3 pathway in a mouse model of sepsis-associated encephalopathy. Brain Behav Immun. 2023;114:195–213. doi:10.1016/j.bbi.2023.08.023
50. Keszthelyi D. Cytokine-responsive vagal afferents and the nucleus of the solitary tract: orchestrators of immune function. Gastroenterology. 2024;167(4):814. doi:10.1053/j.gastro.2024.05.025
51. Jin H, Li M, Jeong E, Castro-Martinez F, Zuker CS. A body-brain circuit that regulates body inflammatory responses. Nature. 2024;630(8017):695–703. doi:10.1038/s41586-024-07469-y
52. Amorim MR, de Deus JL, Cazuza RA, et al. Neuroinflammation in the NTS is associated with changes in cardiovascular reflexes during systemic inflammation. J Neuroinflammation. 2019;16(1):125. doi:10.1186/s12974-019-1512-6
53. El-Naggar AE, Helmy MM, El-Gowilly SM, El-Mas MM. Adenosine A1 receptors of the medullary solitary tract arbitrate the nicotine counteraction of neuroinflammation and cardiovascular dysfunction in septic rats. Sci Rep. 2023;13(1):17818. doi:10.1038/s41598-023-44601-w
54. El-Naggar AE, Helmy MM, El-Gowilly SM, El-Mas MM. Suppression by central adenosine A3 receptors of the cholinergic defense against cardiovascular aberrations of sepsis: role of PI3K/MAPKs/NFkappaB signaling. Front Pharmacol. 2024;15:1418981. doi:10.3389/fphar.2024.1418981
55. Chan JY, Ou CC, Wang LL, Chan SH. Heat shock protein 70 confers cardiovascular protection during endotoxemia via inhibition of nuclear factor-kappaB activation and inducible nitric oxide synthase expression in the rostral ventrolateral medulla. Circulation. 2004;110(23):3560–3566. doi:10.1161/01.Cir.0000143082.63063.33
56. Chan JY, Wang LL, Ou CC, Chan SH. Downregulation of angiotensin subtype 1 receptor in rostral ventrolateral medulla during endotoxemia. Hypertension. 2003;42(1):103–109. doi:10.1161/01.Hyp.0000077984.85616.23
57. Mi Y, Yu H, Wang P, et al. Tyrosine hydroxylase-positive neurons in the rostral ventrolateral medulla mediate sympathetic activation in sepsis. Life Sci. 2024;358:123118. doi:10.1016/j.lfs.2024.123118
58. Abdelnaby M, Sallam MY, Helmy MM, El-Gowelli HM, El-Mas MM. Role of central inflammatory and oxidative pathways in the morphine exacerbation of cardiovascular effects of sepsis in rats. Pharmaceuticals. 2025;18(6):882. doi:10.3390/ph18060882
59. Li W, Zhu H, Zou X, et al. A brain-to-lung signal from GABAergic neurons to ADRB2(+) interstitial macrophages promotes pulmonary inflammatory responses. Immunity. 2025;58(8):2069–2085.e9. doi:10.1016/j.immuni.2025.05.005
60. Koren T, Yifa R, Amer M, et al. Insular cortex neurons encode and retrieve specific immune responses. Cell. 2021;184(24):5902–5915.e17. doi:10.1016/j.cell.2021.10.013
61. Ahmed A, Basile EJ, Ahmad M, et al. Pure insular cortex infarct in sepsis-induced hypoxic ischemic encephalopathy. Cureus. 2021;13(12):e20634. doi:10.7759/cureus.20634
62. Joaquim LS, Steiner B, Farias B, et al. Sepsis compromises post-ischemic stroke neurological recovery and is associated with sex differences. Life Sci. 2024;349:122721. doi:10.1016/j.lfs.2024.122721
63. Florentino D, Della Giustina A, de Souza Goldim MP, et al. Early life neuroimmune challenge protects the brain after sepsis in adult rats. Neurochem Int. 2020;135:104712. doi:10.1016/j.neuint.2020.104712
64. Mittli D. Inflammatory processes in the prefrontal cortex induced by systemic immune challenge: focusing on neurons. Brain Behav Immun Health. 2023;34:100703. doi:10.1016/j.bbih.2023.100703
65. Fernández-Calle R, Vicente-Rodríguez M, Gramage E, et al. Pleiotrophin regulates microglia-mediated neuroinflammation. J Neuroinflammation. 2017;14(1):46. doi:10.1186/s12974-017-0823-8
66. Qin Z, Shi DD, Li W, et al. Berberine ameliorates depression-like behaviors in mice via inhibiting NLRP3 inflammasome-mediated neuroinflammation and preventing neuroplasticity disruption. J Neuroinflammation. 2023;20(1):54. doi:10.1186/s12974-023-02744-7
67. Sharshar T, Gray F, Lorin de la Grandmaison G, et al. Apoptosis of neurons in cardiovascular autonomic centres triggered by inducible nitric oxide synthase after death from septic shock. Lancet. 2003;362(9398):1799–1805. doi:10.1016/s0140-6736(03)14899-4
68. Akasaka S, Nomura M, Nishii H, et al. The hypothalamo-pituitary axis responses to lipopolysaccharide-induced endotoxemia in mice lacking inducible nitric oxide synthase. Brain Res. 2006;1089(1):1–9. doi:10.1016/j.brainres.2006.02.112
69. Li HR, Liu Q, Zhu CL, et al. beta-Nicotinamide mononucleotide activates NAD+/SIRT1 pathway and attenuates inflammatory and oxidative responses in the hippocampus regions of septic mice. Redox Biol. 2023;63:102745. doi:10.1016/j.redox.2023.102745
70. Gofton TE, Young GB. Sepsis-associated encephalopathy. Nat Rev Neurol. 2012;8(10):557–566. doi:10.1038/nrneurol.2012.183
71. Tauber SC, Djukic M, Gossner J, Eiffert H, Bruck W, Nau R. Sepsis-associated encephalopathy and septic encephalitis: an update. Expert Rev Anti Infect Ther. 2021;19(2):215–231. doi:10.1080/14787210.2020.1812384
72. Xu F, Han L, Wang Y, et al. Prolonged anesthesia induces neuroinflammation and complement-mediated microglial synaptic elimination involved in neurocognitive dysfunction and anxiety-like behaviors. BMC Med. 2023;21(1):7. doi:10.1186/s12916-022-02705-6
73. Chung HY, Wickel J, Hahn N, et al. Microglia mediate neurocognitive deficits by eliminating C1q-tagged synapses in sepsis-associated encephalopathy. Sci Adv. 2023;9(21):eabq7806. doi:10.1126/sciadv.abq7806
74. Chaudhuri D, Nei AM, Rochwerg B, et al. Executive summary: guidelines on use of corticosteroids in critically ill patients with sepsis, acute respiratory distress syndrome, and community-acquired pneumonia focused update 2024. Crit Care Med. 2024;52(5):833–836. doi:10.1097/ccm.0000000000006171
75. Mehdi SF, Pusapati S, Anwar MS, et al. Glucagon-like peptide-1: a multi-faceted anti-inflammatory agent. Front Immunol. 2023;14:1148209. doi:10.3389/fimmu.2023.1148209
76. Manion J, Musser MA, Kuziel GA, et al. C. difficile intoxicates neurons and pericytes to drive neurogenic inflammation. Nature. 2023:622(7983):611–618. doi:10.1038/s41586-023-06607-2
77. Edvinsson L, Haanes KA, Warfvinge K, Krause DN. CGRP as the target of new migraine therapies - successful translation from bench to clinic. Nat Rev Neurol. 2018;14(6):338–350. doi:10.1038/s41582-018-0003-1
78. Zhou J, Wu K, Ma Y, et al. GTS-21 alleviates sepsis-induced atrial fibrillation susceptibility by modulating macrophage polarization and Neuregulin-1 secretion. Int Immunopharmacol. 2025;154:114561. doi:10.1016/j.intimp.2025.114561
79. Kong W, Kang K, Gao Y, et al. GTS-21 protected against LPS-induced sepsis myocardial injury in mice through alpha7nAChR. Inflammation. 2018;41(3):1073–1083. doi:10.1007/s10753-018-0759-x
80. Yang A, Wu CH, Matsuo S, Umene R, Nakamura Y, Inoue T. Activation of the alpha7nAChR by GTS-21 mitigates septic tubular cell injury and modulates macrophage infiltration. Int Immunopharmacol. 2024;138:112555. doi:10.1016/j.intimp.2024.112555
81. Li G, Zhou CL, Zhou QS, Zou HD. Galantamine protects against lipopolysaccharide-induced acute lung injury in rats. Braz J Med Biol Res. 2016;49(2):e5008. doi:10.1590/1414-431x20155008
82. Chang EH, Chavan SS, Pavlov VA. Cholinergic control of inflammation, metabolic dysfunction, and cognitive impairment in obesity-associated disorders: mechanisms and novel therapeutic opportunities. Front Neurosci. 2019;13:263. doi:10.3389/fnins.2019.00263
83. Zhang Y, Xin J, Zhao D, et al. Magnesium hexacyanoferrate mitigates sepsis-associated encephalopathy through inhibiting microglial activation and neuronal cuproptosis. Biomaterials. 2025;321:123279. doi:10.1016/j.biomaterials.2025.123279
84. Zhang X, Li R, Chen MY, et al. Investigating the potential mechanism of pioglitazone in sepsis-related brain injury through transcriptomics. Gene. 2024;931:148892. doi:10.1016/j.gene.2024.148892
85. Kanczkowski W, Chatzigeorgiou A, Grossklaus S, Sprott D, Bornstein SR, Chavakis T. Role of the endothelial-derived endogenous anti-inflammatory factor Del-1 in inflammation-mediated adrenal gland dysfunction. Endocrinology. 2013;154(3):1181–1189. doi:10.1210/en.2012-1617
86. Woiciechowsky C, Schoning B, Lanksch WR, Volk HD, Docke WD. Mechanisms of brain-mediated systemic anti-inflammatory syndrome causing immunodepression. J Mol Med. 1999;77(11):769–780. doi:10.1007/s001099900051
87. Zhang FC, Weng RX, Li D, et al. A vagus nerve dominant tetra-synaptic ascending pathway for gastric pain processing. Nat Commun. 2024;15(1):9824. doi:10.1038/s41467-024-54056-w
88. Pena G, Cai B, Liu J, et al. Unphosphorylated STAT3 modulates alpha 7 nicotinic receptor signaling and cytokine production in sepsis. Eur J Immunol. 2010;40(9):2580–2589. doi:10.1002/eji.201040540
89. Wang H, Yu M, Ochani M, et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature. 2003;421(6921):3848. doi:10.1038/nature01339
90. Xu H, Boychuk JA, Boychuk CR, Uteshev VV, Smith BN. Nicotine enhances inhibition of mouse vagal motor neurons by modulating excitability of premotor GABAergic neurons in the nucleus tractus solitarii. J Neurophysiol. 2015;113(4):1165–1174. doi:10.1152/jn.00614.2014
91. Smith DV, Uteshev VV. Heterogeneity of nicotinic acetylcholine receptor expression in the caudal nucleus of the solitary tract. Neuropharmacology. 2008;54(2):445–453. doi:10.1016/j.neuropharm.2007.10.018
92. Rana M, Fei-Bloom Y, Son M, et al. Constitutive vagus nerve activation modulates immune suppression in sepsis survivors. Front Immunol. 2018;9:2032. doi:10.3389/fimmu.2018.02032
93. Kwon HS, Koh SH. Neuroinflammation in neurodegenerative disorders: the roles of microglia and astrocytes. Transl Neurodegener. 2020;9(1):42. doi:10.1186/s40035-020-00221-2
94. Liddelow SA, Marsh SE, Stevens B. Microglia and astrocytes in disease: dynamic duo or partners in crime? Trends Immunol. 2020;41(9):820–835. doi:10.1016/j.it.2020.07.006
95. Hu J, Xie S, Zhang H, Wang X, Meng B, Zhang L. Microglial activation: key players in sepsis-associated encephalopathy. Brain Sci. 2023;13(10):1453. doi:10.3390/brainsci13101453
96. Venet F, Monneret G. Advances in the understanding and treatment of sepsis-induced immunosuppression. Nat Rev Nephrol. 2018;14(2):121–137. doi:10.1038/nrneph.2017.165
97. Matteoli G, Boeckxstaens GE. The vagal innervation of the gut and immune homeostasis. Gut. 2013;62(8):1214–1222. doi:10.1136/gutjnl-2012-302550
98. Kelly MJ, Breathnach C, Tracey KJ, Donnelly SC. Manipulation of the inflammatory reflex as a therapeutic strategy. Cell Rep Med. 2022;3(7):100696. doi:10.1016/j.xcrm.2022.100696
99. Fang YT, Lin YT, Tseng WL, et al. Neuroimmunomodulation of vagus nerve stimulation and the therapeutic implications. Front Aging Neurosci. 2023;15:1173987. doi:10.3389/fnagi.2023.1173987
100. Austelle CW, Cox SS, Wills KE, Badran BW. Vagus nerve stimulation (VNS): recent advances and future directions. Clin Auton Res. 2024;34(6):529–547. doi:10.1007/s10286-024-01065-w
101. Murray K, Cremin M, Tay E, et al. Inhibition of acute lung inflammation by a neuroimmune circuit induced by vagal nerve stimulation. Sci Adv. 2025;11(23):eadw7080. doi:10.1126/sciadv.adw7080
102. Johnson RL, Wilson CG. A review of vagus nerve stimulation as a therapeutic intervention. J Inflamm Res. 2018;11:203–213. doi:10.2147/jir.S163248
103. Mughrabi IT, Gerber M, Jayaprakash N, et al. Voltammetry in the spleen assesses real-time immunomodulatory norepinephrine release elicited by autonomic neurostimulation. J Neuroinflammation. 2023;20(1):236. doi:10.1186/s12974-023-02902-x
104. Yang C, Zhu J, Zhang Y, et al. Effects of transcutaneous auricular vagus nerve stimulation on sepsis-induced coagulopathy: protocol for a randomised controlled trial. BMJ Open. 2025;15(8):e102904. doi:10.1136/bmjopen-2025-102904
105. Premarket Approval (PMA) Database [homepage on the Internet]. Silver Spring (MD): U.S. Food and Drug Administration; 2024. Available from: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfpma/pma.cfm?id=P240039.
106. Manabe T, Heneka MT. Cerebral dysfunctions caused by sepsis during ageing. Nature Reviews Immunology. 2021;22(7):444–458. doi:10.1038/s41577-021-00643-7
107. Wang L, Tang Y, Tang J, et al. Endothelial cell-derived extracellular vesicles expressing surface VCAM1 promote sepsis-related acute lung injury by targeting and reprogramming monocytes. J Extracell Vesicles. 2024. doi:10.1002/jev2.12423
108. Hollenberg SM, Singer M. Pathophysiology of sepsis-induced cardiomyopathy. Nat Rev Cardiol. 2021;18(6):424–434. doi:10.1038/s41569-020-00492-2
109. Joffre J, Hellman J, Ince C, Ait-Oufella H. Endothelial Responses in Sepsis. Am J Respir Crit Care Med. 2020;202(3):361–370. doi:10.1164/rccm.201910-1911tr
110. Frantzeskaki F, Armaganidis A, Orfanos SE. Immunothrombosis in acute respiratory distress syndrome: cross talks between inflammation and coagulation. Respiration. 2016;93(3):212–225. doi:10.1159/000453002
111. Zhang H, Wang Y, Qu M, et al. Neutrophil, neutrophil extracellular traps and endothelial cell dysfunction in sepsis. Clin Transl Med. 2023. doi:10.1002/ctm2.1170
112. Raheem AK, Abu-Raghif AR, Abbas AH, Ridha-Salman H, Oubaid EN. Quercetin mitigates sepsis-induced renal injury via inhibiting inflammatory and oxidative pathways in mice. J Mol Histol. 2025;56(3):184. doi:10.1007/s10735-025-10442-2
113. Shareef SM, Kathem SH, Ridha-Salman H. L-carvone alleviates lipopolysaccharide-Induced acute kidney injury via modulating MyD88-dependent and independent signaling pathways in Mice. Mol Biol Rep. 2025;53(1):39. doi:10.1007/s11033-025-11213-8
114. Sarı Küçük R, Uluç K, Çolakoğlu ŞM, Berktaş CK, Mutlu S, Turgut N. The effect of using iloprost on prognosis in COVID-19 patients with ARDS: a retrospective clinical study. Eur Rev Med Pharmacol Sci. 2023;27(9):4269–4279. doi:10.26355/eurrev_202305_32337
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.