Back to Journals » Neuropsychiatric Disease and Treatment » Volume 12
Brain oscillations in bipolar disorder and lithium-induced changes
Authors Atagün M
Received 16 November 2015
Accepted for publication 21 January 2016
Published 7 March 2016 Volume 2016:12 Pages 589—601
DOI https://doi.org/10.2147/NDT.S100597
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Murat İlhan Atagün
Department of Psychiatry, Yıldırım Beyazıt University Medical School, Cankaya, Ankara, Turkey
Abstract: Electroencephalography (EEG) studies in patients with bipolar disorder have revealed lower amplitudes in brain oscillations. The aim of this review is to describe lithium-induced EEG changes in bipolar disorder and to discuss potential underlying factors. A literature survey about lithium-induced EEG changes in bipolar disorder was performed. Lithium consistently enhances magnitudes of brain oscillations in slow frequencies (delta and theta) in both resting-state EEG studies as well as event-related oscillations studies. Enhancement of magnitudes of beta oscillations is specific to event-related oscillations. Correlation between serum lithium levels and brain oscillations has been reported. Lithium-induced changes in brain oscillations might correspond to lithium-induced alterations in neurotransmitters, signaling cascades, plasticity, brain structure, or biophysical properties of lithium. Therefore, lithium-induced changes in brain oscillations could be promising biomarkers to assess the molecular mechanisms leading to variability in efficacy. Since the variability of lithium response in bipolar disorder is due to the genetic differences in the mechanisms involving lithium, it would be highly promising to assess the lithium-induced EEG changes as biomarkers in genetic studies.
Keywords: lithium, bipolar disorder, brain oscillations, electroencephalography, biomarker
Introduction
Despite pharmacological advances and increasing number of medications, bipolar disorder remains debilitating. Almost one-half of regularly treated patients experience an episode every 2 years, and 73% of these patients experience an episode every 5 years.1 There are significant losses in patient care and pharmacoeconomics in the prolonged process of establishing optimal treatments for each individual.2 It is therefore necessary to further understand the mechanisms of action of the current treatments to identify biomarkers for optimizing the treatment of bipolar disorder.
Personalized treatments are essential to improve the outcomes in the treatment of bipolar disorder. One of the most promising paths is the detection of early signs of treatment responses with in vivo assessments of the brain. With high temporal resolution, the electroencephalography (EEG) measures brain electrophysiology. The activity of large cortical neuron populations is captured from scalp electrodes, and therefore, EEG is an indispensable method for the evaluation of integrative brain function at rest or during task-based experiments. Because the EEG signal comprises excitatory postsynaptic potentials of thalamocortical and corticocortical projections, this analysis evaluates changes in brain physiology (eg, neurotransmitters and network dynamics).3,4 Therefore, EEG is a promising technique with unique capabilities for identifying biomarkers for both the pathophysiology and the neuropsychopharmacology of mental disorders. Earlier researches have detected some predictors of lithium response in clinical EEG;5 however, several quantitative EEG studies have been published since then.
Since the EEG signal incorporates activity of diverse neuron populations, data analysis methods may enable options for the quantification of diverse brain functions. Brain oscillations might be observable reflections of specific brain functions through the formation of cortical codes in networks under different circumstances.4 The most commonly known example is event-related changes in both magnitude and transient phase resetting at specific frequencies, which are reflections of information processing in the brain.6 Furthermore, the maintenance of specific frequency bands across trials with time- and phase-locked features with respect to an event might also indicate consistent brain activity.6
The automatic responses of the sensory cortices in cases of changes in the characteristics of a stimulus are bottom-up tasks in EEG studies. Bottom-up (stimulus-driven) tasks (eg, the mismatch negativity [MMN], the dual-click paradigm [sensory gating, P50], and the loudness dependence of auditory-evoked potentials [LDAEP]) are independent of task-directed performance and are therefore appropriate for all clinical populations and pharmacological EEG studies. The selection of a specific type of stimulus and mental operation rely on cognitive activities, and thus cognitive tasks are top-down tasks. Top-down tasks are modulated through sensory, frontal, and parietal cortices associated with attention, decision making, and working memory.7 Therefore, a major advantage of top-down tasks is that these events are involved in a broader range of cognitive networks and mental operations. Because the most promising feature of the EEG is high temporal resolution, top-down tasks might facilitate the assessment of cognitive networks and external interventions.8
On the other hand, the brain comprises different networks, and resting-state EEG might provide information about the synchronous activity of the intrinsic corticocortical networks.9,10 This promising avenue might develop into translational studies, as shared functional and spatial characteristics of the intrinsic network architecture of the brain across species have been shown through neuroimaging.11 In addition, the majority of pharmacological EEG studies have been performed with resting-state EEGs.12 While functional magnetic resonance imaging studies characterize the spatial distribution of resting state networks, EEG measures offer unique information on the strength and synchronization of neuronal activity at high temporal frequencies. Alpha activity is a classic example, as it becomes active during rest and is augmented when the eyes are closed.
Brief review of EEG studies in bipolar disorder
EEG studies in bipolar disorder have revealed several abnormalities. There are overlapping and distinguishing points with major depression or schizophrenia. Psychosis and mood disorder signatures in the brain are potential biomarkers for further research. The aim of this review is to discuss the neurophysiological abnormalities in bipolar disorder and the effects of lithium. This section aims at briefly reviewing the recent findings of EEG studies in patients with bipolar disorder to extract implications from the neurobiological mechanisms of different EEG modalities.
A spontaneous EEG study reported that schizophrenia patients and their relatives displayed increased delta, theta, and beta activity compared with patients with bipolar disorder and healthy controls.10 Another spontaneous EEG study showed that patients with bipolar disorder and schizophrenia had augmented frontocentral slow beta oscillations, similar to their relatives.13 The authors also reported that delta activity increased in the schizophrenia group, whereas the fast alpha activity increased in the bipolar disorder group. In contrast, a study with a medication-free sample of euthymic bipolar disorder patients reported that the spontaneous alpha activity was highly reduced.14 Kam et al9 showed that bipolar disorder patients had significantly higher beta and gamma activity compared with schizophrenia patients and healthy control groups. Furthermore, the authors also indicated that alpha coherence was increased in both schizophrenia and bipolar disorder patients, whereas schizophrenia patients showed increased beta-1 (12–20 Hz) and delta coherence values. Differences between these results might reflect the differences in medication status observed in these studies.
Auditory stimulation at 40 Hz produces an auditory steady-state response (ASSR) whose generators are present in the primary auditory cortex on the medial aspect of Heschl’s gyrus.15 The ASSR represents the driving of the primary auditory cortex at a gamma frequency range associated with γ-amino butyric acid (GABA) control over fast oscillations. However, a recent study showed that blockade of N-methyl-d-aspartate (NMDA) receptors have increased the intertrial coherence at 20 and 40 Hz oscillations, whereas inhibition of GABAA receptors did not show any significant effect on 20 or 40 Hz oscillations.16 Studies with an ASSR task consistently showed a decrease in the evoke power,17,18 phase-locking factor,19 and altered asymmetry20 in bipolar disorder. Similarly, a magnetoencephalography study with an auditory speech-processing paradigm reported a lateralization abnormality in the speech assessment paradigm that discriminated bipolar disorder and schizophrenia.21
Sensory gating reduces perceived stimuli and eliminates irrelevant stimuli from the environment. This neurological filtering process protects attention resources of the brain and can be assessed through the dual-click paradigm in EEG studies.22 The suppression of the P50 response of the auditory cortex is generated through the inhibitory control of the frontal lobe.23 Patients with bipolar disorder show disturbed sensory gating.24 These findings suggest that bipolar I disorder patients have auditory gating deficits at both preattentive and early attentive levels.
The loudness dependence of the auditory-evoked potentials task is based on serotonergic function in the auditory cortices, and the change of the tone loudness may determine the slope of auditory-evoked potentials.25 The paradigm is involved in clinical states and pharmacological mechanisms in bipolar disorder.26
MMN is a deflection after infrequent change in a repetitive sequence of sounds. The MMN detects changes when a memory trace representing the constant standard stimulus and the neural code of the stimulus with deviant parameters are discrepant.27 The MMN amplitude is based on NMDA receptor function.28 Although MMN studies have consistently shown disturbed responses in schizophrenia, the study results are variable in bipolar disorder, and a recent meta-analysis revealed that MMN was significantly decreased in bipolar disorder.29
The oddball paradigm is a discrimination task based on selection and counting a particular type of stimulus. The results of oddball paradigm experiments suggested significant differences between schizophrenia and bipolar disorder. Most of the studies enrolled patients taking various medications. Because pharmacological agents alter brain physiology, it is more accurate to examine medication-free patients in psychiatric populations. In a study with medication-free euthymic patients, reduced gamma coherence values in both euthymia30 and mania31 were reported. Medication-free patients with bipolar disorder showed significantly decreased amplitudes in both slow (4–6 Hz) and fast (6–8 Hz) theta frequency bands.32 Additionally, in the fast theta band responses of the patients, the decrease was only at target responses, whereas in the slow theta frequency band, theta responses were disturbed in both simple and target responses. In another study with medication-free patients, event-related delta oscillations of medication-free patients with bipolar disorder showed significantly decreased magnitudes.33 Thus, patients with bipolar disorder showed decreased amplitudes and reduced coherence values, and these findings indicate neural synchronization deficits in bipolar disorder.
Lithium-EEG studies
It has been consistently shown that lithium enhances resting-state delta and theta oscillations in spontaneous EEG studies of patients with bipolar disorder34–36 or in healthy volunteers.37,38 In another study using quantitative EEG to evaluate the effects of lithium alone and in combination with other therapies on resting EEG, lithium administration increased the amplitudes of delta, theta, alpha, and beta waves, showing a correlation between serum lithium levels and interhemispheric coherence, delta, and beta-2 (18–30 Hz) values, whereas risperidone and carbamazepine increased the amplitudes of alpha and beta-1 (13–18 Hz) frequencies.39 The authors added that the higher amplitudes in the left temporal brain areas of patients remained unresponsive to lithium, and the amplitude of theta EEG activity was higher in unresponsive patients upon admission to the study.39,40 Ulrich et al41,42 reported that lithium increases resting-state alpha activity in healthy males. In a study with a small sample size (n=27, five responsive to lithium and 22 nonresponsive), it has been reported that there was no statistically significant difference between groups in clinical EEG assessments.5 Another study reported that patients with bipolar disorder exhibiting nonepileptiform sharp wave activity showed a better response to valproate than to lithium.43
The number of task-based studies is relatively low for bipolar disorder. Early studies evaluating responses to lithium through auditory-evoked potentials reported that the amplitudes of the responses are positively correlated with the lithium response.44–46 In an LDAEP paradigm associated with serotonergic function,47 30 patients with bipolar or unipolar disorder who received lithium therapy for at least 3 years were examined, and LDAEP measures were found to be higher in lithium-responsive patients compared with those unresponsive to lithium therapy. Other studies using the LDAEP task reported that N1 and P2 responses predicted a response to lithium.48,49 Another study did not detect any relationship between the P300 response and auditory or visual event-related P300 responses.50
The aforementioned lithium-EEG studies were performed with heterogeneous samples and have enrolled patients with different mood statuses. In studies to predict lithium responses, the definition of the lithium response was not clear. However, the amplification of slow frequencies (delta and theta oscillations) is a consistent finding in both healthy volunteers and patients with bipolar disorder.
In a study using an auditory oddball paradigm, 16 patients with bipolar disorder on lithium monotherapy, 22 medication-free patients, and a control group similar to the other groups in terms of age, sex, and education were enrolled (Figure 1).51 In the lithium group, we observed a significant peak at 20 Hz in target responses in the Fourier transform analysis. The lithium group had significantly higher beta frequency oscillations in response to target stimuli under the auditory oddball paradigm. Interestingly, there was a negative correlation between serum lithium concentrations and amplitudes of event-related beta oscillations. Similarly, beta oscillations were significantly increased under the visual oddball paradigm.52 The lithium group also displayed significantly higher evoked and event-related alpha oscillations.53 This effect is specific to the cognitive component of the tasks for beta frequencies, reflecting its emergence rather than its location in cognitive networks.
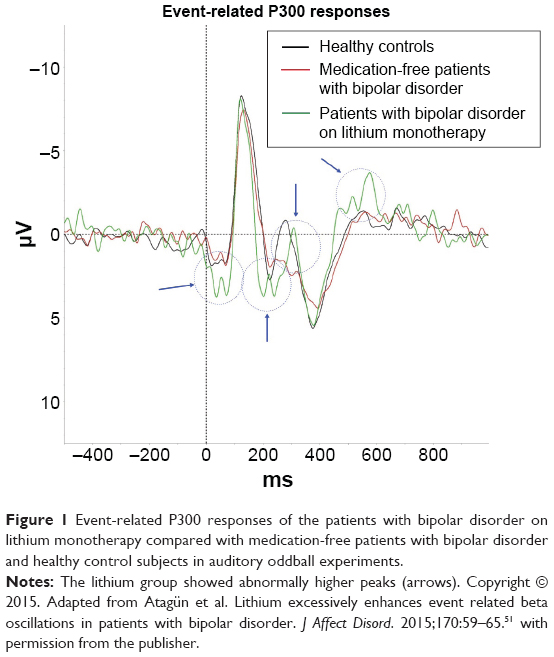 | Figure 1 Event-related P300 responses of the patients with bipolar disorder on lithium monotherapy compared with medication-free patients with bipolar disorder and healthy control subjects in auditory oddball experiments. |
Taken together, the amplitude-elevating effects of lithium seem to uniformly elevate all EEGs and EROs.39 Although specificity to target responses in beta oscillations might have specific attributions, general increases in all amplitudes indicate synaptic plasticity and altered balance between excitatory and inhibitory neurotransmission.3
A few studies have shown that valproate54,55 and carbamazepine56 do not exhibit the same pattern of change as lithium in the EEG. Özerdem et al54 reported that valproate had mixed effects on brain oscillations; patients undergoing valproate monotherapy showed increased alpha and decreased beta oscillations. This pattern is not similar to that of lithium-EEG, as lithium consistently amplifies all frequency bands. Compared with other mood stabilizers, the effects of lithium on the EEG signal are specific to lithium, and when compared with other psychotropic medication groups, including antipsychotics,57,58 antidepressants,59 stimulants,60 benzodiazepines,61 analgesics,62 or glutamatergic agents, such as ketamine63,64 and riluzole,65 lithium-induced EEG changes are hallmarks specific to bipolar disorder. However, the consistency of the findings across studies refers to the sensitivity of the changes. As these changes are sensitive, specific, and reproducible, lithium-induced EEG changes are consistent with the biomarker definition.66
Potential mechanisms of lithium-induced changes in brain oscillations
Influences on neurotransmitters, signaling cascades, neurotrophic factors, and neuroplasticity cascades are mechanisms of action for lithium.67 Mechanisms that modify integrative brain function with lithium might be involved in most of the mentioned mechanisms, and the relevance of these mechanisms with EEG changes will be discussed in the following sections.
Lithium-induced neurotransmitter changes
Lithium downregulates NMDA receptors and attenuates calcium metabolism and signaling in the brain.68 However, lithium activates α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, and it has been suggested that AMPA receptor activity mediates the effects of lithium.69 This mechanism potentially reflects the enhancement of brain oscillations. Additionally, Kainate receptors modulate synaptic activity.70 Type 2 Kainate receptors (GluK2) increase intracellular calcium concentrations and consequently enhance synaptic glutamatergic activity.71 Lithium selectively inhibits the mRNA and protein expression of GluK2 receptors in astrocytes and inhibits Kainate receptors.72
In the presynaptic terminal, 2 and 3 receptor subtypes of metabotropic glutamate receptors decrease the magnitude of single-neuron action-potential-generated transmitter release73 and reduce spontaneous transmitter release. Overall, the effect of this phenomenon increases the signal-to-noise ratio (SNR) at the synaptic level and increases the stability and consistency of the synaptic response. Consistently, Gandal et al74 proposed that in neuroimaging studies with psychotropic medications, the SNR is increased because of the effects of medications on GABA and glutamatergic neurotransmission. The activity of dopaminergic receptors in the prefrontal cortex might regulate the activity of pyramidal neurons75 and excitatory synapses.76 These receptors might also increase the input–output responses in pyramidal neurons,77 modulate persistent synaptic activity, and enhance the SNR.78 These mechanisms are all targets of lithium, and thus, lithium might increase the SNR in neuroimaging studies.
Despite the diversity of excitatory neurotransmitters in the brain, inhibitory neurotransmitters are scarce. In addition to other tasks, the tuning of excitation through inhibitory neurotransmission is particularly important for resonant and coherent brain activity. GABAergic interneurons are major sources of inhibitory neurotransmission in the brain, and this function is of particular importance for the production of gamma oscillations.79 GABAergic receptors are highly susceptible to neuroplasticity. The phosphorylation of GABAA receptors alters the density and function of these proteins.80 Various enzymes and cascades involved in phosphorylation include neurosteroids, protein kinase C, and Akt,81 which are targets for lithium.82 Additionally, glycogen synthase kinase 3β (GSK-3β) modulates the phosphorylation of GABAA receptors via the phosphorylation of synaptic proteins (ie, gephyrin), and the inhibition of GSK-3β through lithium precludes phosphorylation and thereby the degradation of receptors.83 Other studies have reported that cyclin-dependent kinase-5 (CDK-5),84 integrin signaling,85 and brain-derived neurotrophic factor86 modulate gephyrin phosphorylation. These cascades are also targets of lithium. Overall, lithium changes the GABAergic receptor density and function through various mechanisms, which should be translated into human neuroscience.
Other lithium-induced changes in the brain
Scalp EEG sensors measure the bioelectrical activity of neuron populations when they can summate properly. This summation is only possible when the directions of the vectors of the electrical fields are similar. Similar vectorial angles reflect similar angles of the apical dendrites of the pyramidal neurons typically turned through external space with similar alignments. If the order of the cells and their alignment change, then the summation of the vectors is attenuated, reflecting weakened summation. Therefore, the failure of vector summation might result from altered cellular connectivity and alignment. Reduced cortical thickness in the left superior temporal cortices has been associated with reduced P300 amplitudes in schizophrenia,87 and this finding was replicated in several studies.88–90 Similarly, occipital resting-state alpha oscillations were correlated with occipital gray matter density in a group of elderly subjects, including patients with Alzheimer’s disease and mild cognitive impairment, and in healthy older adults.91 The same group later reported that frontal event-related delta oscillations were correlated with the frontal cortex volume in patients with mild cognitive impairments and in healthy age-matched controls.92 These studies, showing the correlation between imaging modalities, strongly suggested the same underlying physiopathology of morphological and functional disturbances. Altered anatomic localization and the position of the pyramidal neuron layers are underlying reasons for the decreased amplitudes in psychotic spectrum disorders. As schizophrenia and bipolar disorder share similar pathogenesis,93,94 structural and functional changes in brain tissue might also be reasons for neurophysiological abnormalities in bipolar disorder.
Research on schizophrenia and bipolar disorder has demonstrated abnormalities in glial cells,95 the number of dendritic spines,96 synapse density,97 cortical thickness,98,99 cortical folding/sulcus patterns,100 and laminar microstructure.101 These data indicate a neurodevelopmental basis and progressive histopathological changes in the neurobiology of bipolar disorder and schizophrenia.102 Two recent meta-analyses reported that cortical thickness decreases with progression to psychosis, and meta-regressions reported that antipsychotics have an inverse relationship with the loss of cortical thickness.98,99 Despite confounding factors, lithium increases gray matter volume in bipolar disorder.103 A recent report indicated that lithium might increase the gray matter in the brain according to the GSK-3β gene variant.104 These authors suggested that the neurotrophic effects of lithium are associated with GSK-3β inhibition.
In addition, lithium might modify cortical surface folding.100 An increase in the gray matter through antipsychotics could result from increasing the neuronal soma size,105 synaptic density,106 and stimulating neurogenesis.107 Lithium increases synaptic strength,99 the number of dendrites,108 and neurogenesis.109 Lithium enhances long-term potentiation independent of neurogenesis,110 likely reflecting the stimulation of synapse formation cascades. Synapsin II is a gene encoding a phosphoprotein associated with synaptogenesis, synaptic plasticity, and transmission.111 Synapsin II gene-encoded protein levels are downregulated.112 Interestingly, a comparison between responders and nonresponders showed that lithium modulates synapsin II expression more in the responder group.113 The response to lithium might be associated with synapsin II expression, with respect to synaptic strength and neuroplasticity. Thus, cortical thickness, laminar alignment disturbances, reduced number of synapses and dendrites, and reduced synaptic strength result from disabled neurotrophic and neuroplasticity cascades. These changes might cause morphological reformation, leading to a recovery of the brain potentials. Medications could enhance magnitudes and reverse neuroimaging findings through the stimulation of neuroplastic cascades.
Cellular migration and laminar structure formation are associated with cellular and subcellular mechanisms that include protein families encoded by certain genes. The most commonly known genes are Neuregulin, Disbindin, and DISC-1, and these genes are abnormal in schizophrenia and bipolar disorder.114 Wnt/β catenin,115 Reelin,116 and CDK117 pathways are members of these protein families that act as major regulators of the settlement and connectivity of neurons in both embryonic life and adulthood. These protein families and cascades are responsive to pharmacological interventions and psychotropic medications.116,118 Further studies with interventions for the CDK, Reelin, and Wnt pathways might identify the determination processes and organization of the laminar structures of the cortex. The same pathology might disturb P300 responses through precise changes in the alignment of neurons at the cortical surface. However, to date, no study has compared the structural data with EEG measurements in patients with bipolar disorder, and these findings should be translated to patients with bipolar disorder in future studies.
Disturbances of brain oscillations in both bipolar disorder and schizophrenia are highly genetically inherited.24 The genes and protein families are potential causes of disturbances in integrative brain functions because the disturbed brain oscillations are amenable to these products. The relationship between P3 responses upon the auditory oddball paradigm and a number of genes in schizophrenia and psychotic bipolar disorder has recently been investigated.118 These authors reported that the genes involved in immune response complement pathways, cell adhesion, axon guidance, and neurogenesis were associated with P3 amplitudes in the frontal cortex. However, because genes are pleiotropic and multiple mechanisms influence each other in the brain, these findings should be cautiously evaluated. Further studies with appropriate designs should examine the effects of each cascade separately.
However, lithium excessively enhances brain oscillations, and therefore, it is possible to speculate that lithium might stimulate additional mechanisms. In addition to increasing synapse formation, lithium might also stimulate excitatory transmission and/or synaptic potentiation.
Synchronization dynamics or cognitive functions? Bottom-up versus top-down influences
Lithium and neural synchronization: bottom-up mechanisms
Neuronal population behaviors often characterize the brain dynamics observed during the generation of synchronous oscillations. Both network effects119 and pacemaker cells120 stimulate the synchronous oscillation of neurons. The neuron population fires together because these cells respond to direct oscillatory input or cellular interactions might fire synchronously for neuron groups. Beyond the characteristics of the oscillation, adequate expansion of an oscillation might require sufficient networks and adequate cellular responses to the stimulus. Lithium has neurotrophic and neuroplastic effects that improve network integrity. In addition, lithium augments neuronal excitability through alterations in ion channel physiology. For example, altered sodium (Na) and potassium (K) channel physiology decreased after hyperpolarization, enhanced Na-K ATPase activity, increased the response rates of long-latency synapses,121 altered G-protein-gated K channels,122 and attenuated calcium channel responses to glutamate receptor signals.123 These in vitro studies consistently suggest that lithium salt might increase the excitability of the neurons. In short, although in the current literature there is a gap between the dynamics of large-scale networks and cellular activity, it is reasonable to speculate that lithium might enhance cortical excitability acutely by its biophysical properties, and chronically by improving network integrity. Enhancement of the synchronization capabilities may lead to increased phase precision and augmented magnitudes in neurophysiological assessments. To date, there are no studies reporting the effects of lithium on phase. Studies have consistently reported that delta and theta oscillations are prominently increased in patients using lithium, independent from tasks (Tables 1 and 2). Beta oscillations are selectively elevated upon event-related tasks.51,52 The molecular underpinnings of these effects maybe involved with the cognitive brain dynamics, and has to be observed in future studies. The nonselective effects on slow frequencies might be related with biophysical changes or increased volume conduction. However, changes in fast frequencies seem to be independent of chemical changes and are more likely to be involved with synaptic changes and network integrity. These proposals could be investigated by further brain oscillation studies assessing both magnitudes of brain oscillations and phase changes in bipolar disorder patients on lithium therapy.
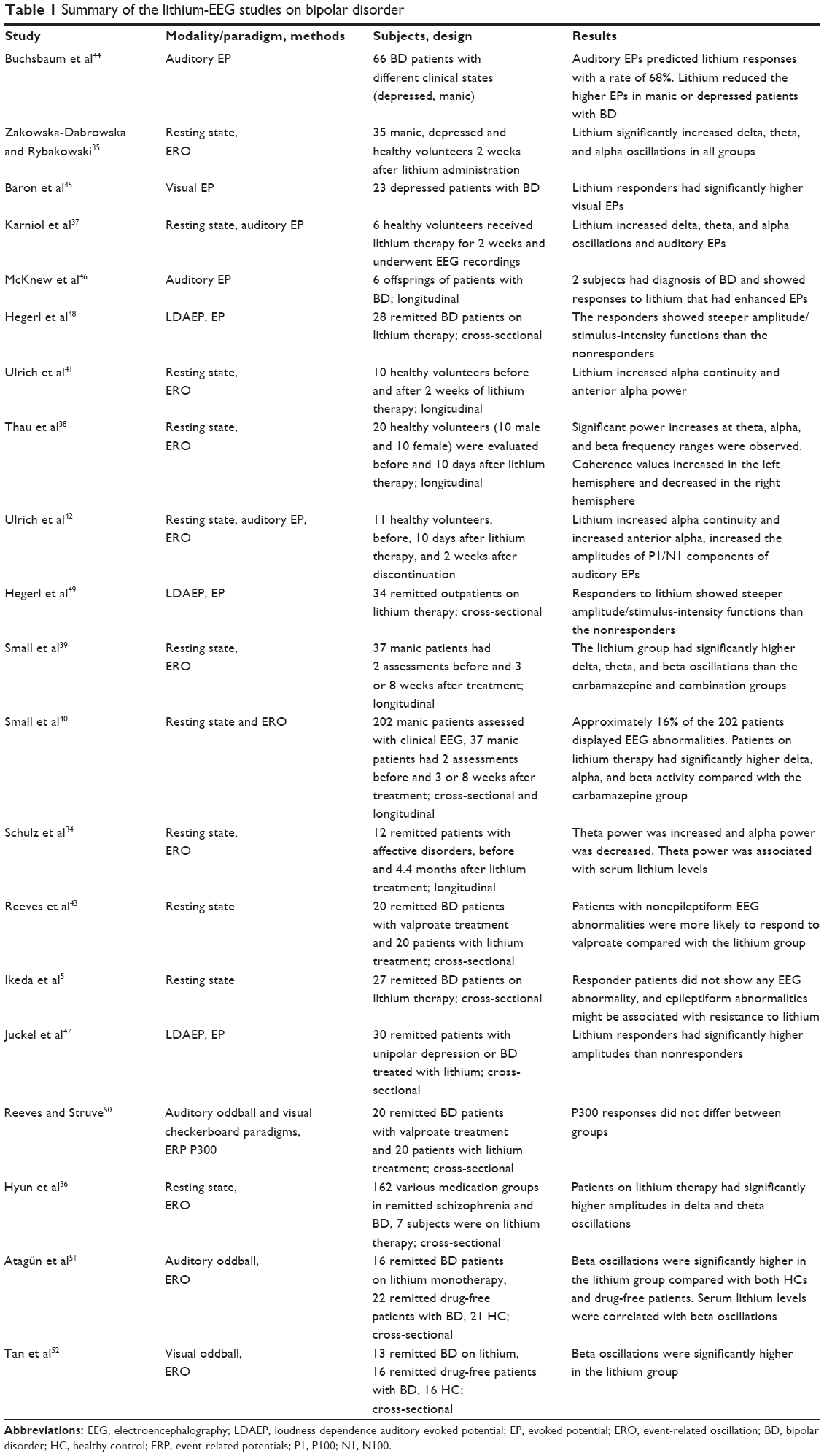 | Table 1 Summary of the lithium-EEG studies on bipolar disorder |
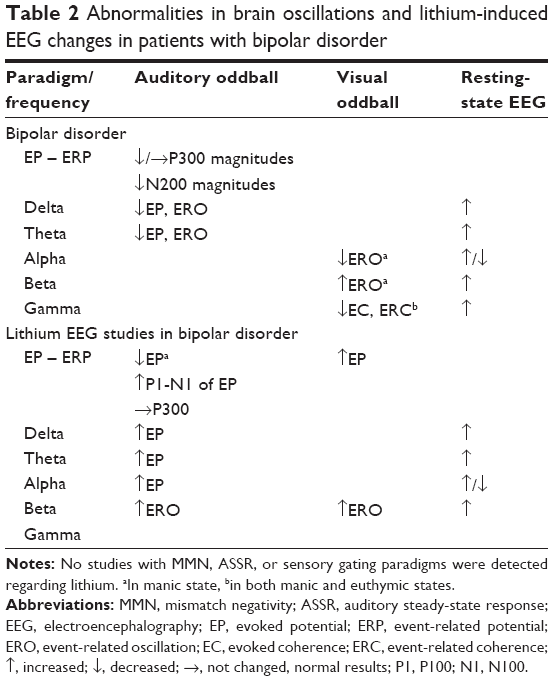 | Table 2 Abnormalities in brain oscillations and lithium-induced EEG changes in patients with bipolar disorder |
Cognitive effects of lithium and brain oscillations: top-down mechanisms
A top-down approach might imply an influence on changes in cognitive functions. Brain rhythms are associated with specific functions, and the derangement of a particular rhythm might be associated with a cognitive deficit.124 Bipolar disorder is characterized by dysfunction in several domains of cognition, likely associated with this network. Cognitive impairments in bipolar disorder include attention, memory, executive function, response inhibition, and information processing.125,126 The neurocognitive effects of lithium have been examined, and studies have reported that except for some motor disturbances, lithium does not generally cause a specific change in the cognitive function of patients with bipolar disorder.127,128 Briefly, the current literature does not indicate a clear relationship between the neurocognitive effects of lithium and EEG changes.
Mechanisms of pharmacological interventions on brain oscillations
Fast oscillations are generated for synchrony between specific brain regions. Beta oscillations have been associated with memory formation and recall, stimulus saliency, attention, and motor movements.51 Synchronous gamma oscillations might generate beta oscillations via synaptic potentiation.129 It has been shown that after the activation of a synaptic plasticity cascade, consistent plastically altered networks produce beta oscillations.79 Furthermore, the inhibition of AMPA receptors immediately abolishes beta oscillations.129 However, after the blockade of GABAergic interneuron activity in synchronously oscillating hippocampal areas, beta oscillations continued.130 Another recent study has reported that the degradation of perineuronal nets wrapping up parvalbumin-containing interneurons significantly enhanced beta oscillations, whereas with the decreased maturity of the neurons, gamma oscillations might decrease and beta oscillations might significantly increase.131 These studies suggest that gamma oscillations depend on GABAergic cells and that beta oscillations are related with glutamatergic AMPA receptors.79,129,130,131
Oscillations generate synchronous neuronal discharge patterns across large-scale networks, irrespective of axonal conduction delays. However, the disruption of myelin might prolong the conduction time in networks, thereby disrupting synchronicity and coherence in mental disorders.132 The myelination of excitatory neurons projecting onto GABAergic interneurons determines the level of activation, and thereby, GABAergic neurotransmission.133 Because inhibitory (GABAergic) neurotransmission determines gamma oscillations, Traub et al129 proposed that excitatory inputs to interneurons might determine the frequency of oscillation in neuron populations. The duration of inhibitory synaptic control influences the fast frequencies, and the amplitude of the synaptic GABA response determines the power of the oscillation.130 Considering that atypical antipsychotics134 and lithium135 stimulate the maturation of oligodendrocytes and the synthesis and maturation of myelin, it is possible that these drugs provide better GABAergic activity and thus potentiate fast frequency oscillations.
Conclusion
To conclude, EEG studies have consistently reported abnormalities in bipolar disorder, and lithium induces typical EEG changes. It is possible to draw the following conclusions from the current EEG literature:
- Bipolar disorder is characterized through various abnormalities in brain oscillations (Table 2). In addition to the abnormalities listed in Table 2, patients with bipolar disorders display decreased MMN amplitudes,29 decreased evoked gamma power in ASSR experiments,18 and disturbed sensory gating.24 These abnormalities reflect disturbances in integrative brain functions deriving from various etiological mechanisms, including changes in neuronal excitability, neuroplasticity, neurotransmitters, and cortical thinning.
- Lithium enhances slow frequencies in either resting-state or task-based studies. However, the enhancement of the fast frequencies might be specific to task demands and stimulus characteristics, as lithium selectively enhances event-related beta oscillations.
- The potentiation of the brain oscillations might imply various brain-wide effects of lithium, including neuroplastic and neurotransmitter changes as well as biophysical changes. Furthermore, lithium increases cortical thickness and brain volume, and structural changes induced by lithium might be related with the lithium-induced EEG changes. Enhancement of beta oscillations might also correspond with AMPA and GABA receptor activities and neuroplastic changes. It seems that a variety of mechanisms are involved in the changes in the brain oscillations. Enhancement of the slow frequencies might correspond with all the mechanisms mentioned in this paper, whereas enhancement of fast frequencies is related with changes in signaling cascades and synaptic plasticity.
Future studies with bipolar disorder patients undergoing lithium therapy might involve phase and gamma oscillations. In addition, studying the length of lithium use would be beneficial, as biophysical properties of lithium salt suggest acute enhancement in conductivity of brain tissues and electrophysiological activity in neurons. MMN, ASSR, and sensory gating paradigms have shown abnormalities in bipolar disorder; however, to date, no studies have examined the neurophysiological effects of lithium with these modalities. Lithium-induced EEG changes could be used as biomarkers to assess the genetic differences in signaling and neuroplasticity, leading to differential efficacies of lithium that may underlie the variability of lithium response in bipolar disorder. To optimize treatment strategies, brain oscillations are promising biomarker candidates, as these effects assess integrative brain functions and responses to pharmacological agents. Future studies with larger samples and standard protocols might be promising for examining the pharmacological mechanisms of lithium.
Disclosure
The author reports no conflicts of interest in this work.
References
Perlis RH, Ostacher MJ, Patel JK, et al. Predictors of recurrence in bipolar disorder: primary outcomes from the Systematic Treatment Enhancement Program for Bipolar Disorder (STEP-BD). Am J Psychiatry. 2006;163:217–224. | ||
Kleinman L, Lowin A, Flood E, et al. Costs of bipolar disorder. Pharmacoeconomics. 2003;21:601–622. | ||
Buzsaki G. Rhythms of the Brain. New York, NY: Oxford University Press; 2006. | ||
Başar E. Brain Oscillations Principals and Approaches. Berlin, Germany: Springer; 1998. | ||
Ikeda A, Kato N, Kato T. Possible relationship between electroencephalogram finding and lithium response in bipolar disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2002;26:903–907. | ||
Makeig S, Debener S, Onton J, Delorme A. Mining event-related brain dynamics. Trends Cogn Sci. 2004;8:204–210. | ||
Wagner AD, Schacter DL, Rotte M, et al. Building memories: remembering and forgetting of verbal experiences as predicted by brain activity. Science. 1998;281:1188–1191. | ||
Kenemans JL, Kahkonen S. How human electrophysiology informs psychopharmacology: from bottom-up driven processing to top-down control. Neuropsychopharmacology. 2011;36:26–51. | ||
Kam JW, Bolbecker AR, O’Donnell BF, et al. Resting state EEG power and coherence abnormalities in bipolar disorder and schizophrenia. J Psychiatr Res. 2013;47:1893–1901. | ||
Venables NC, Bernat EM, Sponheim SR. Genetic and disorder-specific aspects of resting state EEG abnormalities in schizophrenia. Schizophr Bull. 2009;35:826–839. | ||
Smucny J, Wylie KP, Tregellas JR. Functional magnetic resonance imaging of intrinsic brain networks for translational drug discovery. Trends Pharmacol Sci. 2014;35:397–403. | ||
Jobert M, Wilson FJ, Ruigt GS, et al. Guidelines for the recording and evaluation of pharmaco-EEG data in man: the International Pharmaco-EEG Society (IPEG). Neuropsychobiology. 2012;66:201–220. | ||
Narayanan B, O’Neil K, Berwise C, et al. Resting state electroencephalogram oscillatory abnormalities in schizophrenia and psychotic bipolar patients and their relatives from the bipolar and schizophrenia network on intermediate phenotypes study. Biol Psychiatry. 2014;76:456–465. | ||
Başar E, Güntekin B, Atagün I, et al. Brain’s alpha activity is highly reduced in euthymic bipolar disorder patients. Cogn Neurodyn. 2012;6:11–20. | ||
Pantev C, Roberts LE, Elbert T, et al. Tonotopic organization of the sources of human auditory steady-state responses. Hear Res. 1996;101:62–74. | ||
Sullivan EM, Timi P, Hong LE, O’Donnell P. Effects of NMDA and GABA-A receptor antagonism on auditory steady-state synchronization in awake behaving rats. Int J Neuropsychopharmacol. 2015;18:pyu118. | ||
Oda Y, Onitsuka T, Tsuchimoto R, et al. Gamma band neural synchronization deficits for auditory steady state responses in bipolar disorder patients. PLoS One. 2012;7:e39955. | ||
O’Donnell BF, Hetrick WP, Vohs JL, et al. Neural synchronization deficits to auditory stimulation in bipolar disorder. Neuroreport. 2004;15:1369–1372. | ||
Rass O, Krishnan G, Brenner CA, et al. Auditory steady state response in bipolar disorder: relation to clinical state, cognitive performance, medication status, and substance disorders. Bipolar Disord. 2010;12:793–803. | ||
Reite M, Teale P, Rojas DC, et al. MEG auditory evoked fields suggest altered structural/functional asymmetry in primary but not secondary auditory cortex in bipolar disorder. Bipolar Disord. 2009;11:371–381. | ||
Oribe N, Onitsuka T, Hirano S, et al. Differentiation between bipolar disorder and schizophrenia revealed by neural oscillation to speech sounds: an MEG study. Bipolar Disord. 2010;12:804–812. | ||
Freedman R, Adler LE, Gerhardt GA, et al. Neurobiological studies of sensory gating in schizophrenia. Schizophr Bull. 1987;13:669–678. | ||
Weisser R, Weisbrod M, Roehrig M, et al. Is frontal lobe involved in the generation of auditory evoked P50? Neuroreport. 2001;12:3303–3307. | ||
Hall MH, Levy DL, Salisbury DF, et al. Neurophysiologic effect of GWAS derived schizophrenia and bipolar risk variants. Am J Med Genet B Neuropsychiatr Genet. 2014;165B(1):9–18. | ||
Hegerl U, Juckel G. Intensity dependence of auditory evoked potentials as an indicator of central serotonergic neurotransmission: a new hypothesis. Biol Psychiatry. 1993;33:173–187. | ||
Park YM, Lee SH. Clinical usefulness of loudness dependence of auditory evoked potentials (LDAEP) in patients with bipolar disorder. Psychiatry Investig. 2013;10:233–237. | ||
Naatanen R. Attention and Brain Function. Hillsdale, NJ: Lawrence Erlbaum Associates; 1992. | ||
Javitt DC, Steinschneider M, Schroeder CE, Arezzo JC. Role of cortical N-methyl-d-aspartate receptors in auditory sensory memory and mismatch negativity generation: implications for schizophrenia. Proc Natl Acad Sci U S A. 1996;93:11962–11967. | ||
Chitty KM, Lagopoulos J, Lee RS, et al. A systematic review and meta-analysis of proton magnetic resonance spectroscopy and mismatch negativity in bipolar disorder. Eur Neuropsychopharmacol. 2013;23:1348–1363. | ||
Ozerdem A, Guntekin B, Atagun I, et al. Reduced long distance gamma (28–48 Hz) coherence in euthymic patients with bipolar disorder. J Affect Disord. 2011;132:325–332. | ||
Ozerdem A, Güntekin B, Saatçi E, et al. Disturbance in long distance gamma coherence in bipolar disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2010;34:861–865. | ||
Atagün MI, Güntekin B, Ozerdem A, et al. Decrease of theta response in euthymic bipolar patients during an oddball paradigm. Cogn Neurodyn. 2013;7:213–223. | ||
Atagün MI, Güntekin B, Maşali B, et al. Decrease of event-related delta oscillations in euthymic patients with bipolar disorder. Psychiatry Res. 2014;223:43–48. | ||
Schulz C, Mavrogiorgou P, Schröter A, et al. Lithium-induced EEG changes in patients with affective disorders. Neuropsychobiology. 2000;42(Suppl 1):33–37. | ||
Zakowska-Dabrowska T, Rybakowski J. Lithium-induced EEG changes: relation to lithium levels in serum and red blood cells. Acta Psychiatr Scand. 1973;49:457–465. | ||
Hyun J, Baik MJ, Kang UG. Effects of psychotropic drugs on quantitative EEG among patients with schizophrenia-spectrum disorders. Clin Psychopharmacol Neurosci. 2011;9:78–85. | ||
Karniol IG, Dalton J, Lader MH. Acute and chronic effects of lithium chloride on physiological and psychological measures in normals. Psychopharmacology (Berl). 1978;57:289–294. | ||
Thau K, Rappelsberger P, Lovrek A, et al. Effect of lithium on the EEG of healthy males and females. A probability mapping study. Neuropsychobiology. 1989;20:158–163. | ||
Small JG, Milstein V, Malloy FW, et al. Topographic EEG studies of mania. Clin Electroencephalogr. 1998;29:59–66. | ||
Small JG, Milstein V, Malloy FW, et al. Clinical and quantitative EEG studies of mania. J Affect Disord. 1999;53:217–224. | ||
Ulrich G, Frick K, Stieglitz RD, Müller-Oerlinghausen B. Interindividual variability of lithium-induced EEG changes in healthy volunteers. Psychiatry Res. 1987;20:117–127. | ||
Ulrich G, Herrmann WM, Hegerl U, Müller-Oerlinghausen B. Effect of lithium on the dynamics of electroencephalographic vigilance in healthy subjects. J Affect Disord. 1990;20:19–25. | ||
Reeves RR, Struve FA, Patrick G. Does EEG predict response to valproate versus lithium in patients with mania? Ann Clin Psychiatry. 2001;13:69–73. | ||
Buchsbaum M, Goodwin F, Murphy D, Borge G. AER in affective disorders. Am J Psychiatry. 1971;128:19–25. | ||
Baron M, Gershon ES, Rudy V, et al. Lithium carbonate response in depression. Prediction by unipolar/bipolar illness, average-evoked response, catechol-O-methyl transferase, and family history. Arch Gen Psychiatry. 1975;32:1107–1111. | ||
McKnew DH, Cytryn L, Buchsbaum MS, et al. Lithium in children of lithium-responding parents. Psychiatry Res. 1981;4:171–180. | ||
Juckel G, Mavrogiorgou P, Bredemeier S, et al. Loudness dependence of primary auditory-cortex-evoked activity as predictor of therapeutic outcome to prophylactic lithium treatment in affective disorders – a retrospective study. Pharmacopsychiatry. 2004;37:46–51. | ||
Hegerl U, Ulrich G, Müller-Oerlinghausen B. Auditory evoked potentials and response to lithium prophylaxis. Pharmacopsychiatry. 1987;20:213–216. | ||
Hegerl U, Wulff H, Müller-Oerlinghausen B. Intensity dependence of auditory evoked potentials and clinical response to prophylactic lithium medication: a replication study. Psychiatry Res. 1992;44:181–190. | ||
Reeves RR, Struve FA. Auditory and visual P300 evoked potentials do not predict response to valproate versus lithium in patients with mania. Ann Clin Psychiatry. 2005;17:47. | ||
Atagün MI, Güntekin B, Tan D, et al. Lithium excessively enhances event related beta oscillations in patients with bipolar disorder. J Affect Disord. 2015;170:59–65. | ||
Tan D, Ozerdem A, Güntekin B, et al. Increased β frequency (15–30 Hz) oscillatory responses in euthymic bipolar patients under lithium monotherapy. Clin EEG Neurosci. Epub 2014 Dec 2. | ||
Atagün MI, Guntekin B, Tan D, et al. Lithium excessively enhances alpha and beta oscillatory responses in bipolar disorder. Biol Psychiatry. 2013;73:187S. | ||
Özerdem A, Güntekin B, Tunca Z, Başar E. Brain oscillatory responses in patients with bipolar disorder manic episode before and after valproate treatment. Brain Res. 2008;1235:98–108. | ||
Ozerdem A, Güntekin B, Atagün MI, Başar E. Brain oscillations in bipolar disorder in search of new biomarkers. Suppl Clin Neurophysiol. 2013;62:207–221. | ||
Panagopoulos GR, Thomaides T, Tagaris G, Karageorgiou CL. Auditory event related potentials in patients with epilepsy on sodium valproate monotherapy. Acta Neurol Scand. 1997;96:62–64. | ||
Javitt DC, Spencer KM, Thaker GK, et al. Neurophysiological biomarkers for drug development in schizophrenia. Nat Rev Drug Discov. 2008;7:68–83. | ||
Galderisi S, Mucci A. Psychophysiology in psychiatry: new perspectives in the study of mental disorders. World Psychiatry. 2002;1:166–168. | ||
Jaworska N, Protzner A. Electrocortical features of depression and their clinical utility in assessing antidepressant treatment outcome. Can J Psychiatry. 2013;58:509–514. | ||
Klorman R, Brumaghim JT. Stimulant drugs and ERPs. Electroencephalogr Clin Neurophysiol Suppl. 1991;42:135–141. | ||
Fitzgerald AC, Wright BT, Heldt SA. The behavioral pharmacology of zolpidem: evidence for the functional significance of α1-containing GABA(A) receptors. Psychopharmacology (Berl). 2014;231:1865–1896. | ||
Malver LP, Brokjaer A, Staahl C, et al. Electroencephalography and analgesics. Br J Clin Pharmacol. 2014;77:72–95. | ||
Horacek J, Brunovsky M, Novak T, et al. Subanesthetic dose of ketamine decreases prefrontal theta cordance in healthy volunteers: implications for antidepressant effect. Psychol Med. 2010;40:1443–1451. | ||
Hong LE, Summerfelt A, Buchanan RW, et al. Gamma and delta neural oscillations and association with clinical symptoms under subanesthetic ketamine. Neuropsychopharmacology. 2010;35:632–640. | ||
Saletu B, Grunberger J, Anderer P, Linzmayer L. Effects of the novel neuroprotective agent, riluzole, on human brain function and behavior: I. Double-blind, placebo-controlled EEG mapping and psychometric studies under normoxia. Methods Find Exp Clin Pharmacol. 1996;18:55–66. | ||
Scarr E, Millan MJ, Bahn S, et al. Biomarkers for psychiatry: the journey from fantasy to fact, a report of the 2013 CINP think tank. Int J Neuropsychopharmacol. 2015;18(10):pyv042. | ||
Manji HK, Potter WZ, Lenox RH. Signal transduction pathways. Molecular targets for lithium’s actions. Arch Gen Psychiatry. 1995;52:531–543. | ||
Nonaka S, Hough CJ, Chuang DM. Chronic lithium treatment robustly protects neurons in the central nervous system against excitotoxicity by inhibiting N-methyl-d-aspartate receptor-mediated calcium influx. Proc Natl Acad Sci U S A. 1998;95:2642–2647. | ||
Gould TD, O’Donnell KC, Dow ER, et al. Involvement of AMPA receptors in the antidepressant-like effects of lithium in the mouse tail suspension test and forced swim test. Neuropharmacology. 2008;54:577–587. | ||
Jane DE, Lodge D, Collingridge GL. Kainate receptors: pharmacology, function and therapeutic potential. Neuropharmacology. 2009;56:90–113. | ||
Parpura V, Baker BJ, Jeras M, Zorec R. Regulated exocytosis in astrocytic signal integration. Neurochem Int. 2010;57:451–459. | ||
Li B, Zhang S, Li M, et al. Down-regulation of GluK2 kainate receptor expression by chronic treatment with mood-stabilizing anti-convulsants or lithium in cultured astrocytes and brain, but not in neurons. Neuropharmacology. 2009;57:375–385. | ||
Kamiya H, Ozawa S. Dual mechanism for presynaptic modulation by axonal metabotropic glutamate receptor at the mouse mossy fibre-CA3 synapse. J Physiol. 1999;518(Pt 2):497–506. | ||
Gandal MJ, Edgar JC, Klook K, Siegel SJ. Gamma synchrony: towards a translational biomarker for the treatment-resistant symptoms of schizophrenia. Neuropharmacology. 2012;62:1504–1518. | ||
Gurden H, Takita M, Jay TM. Essential role of D1 but not D2 receptors in the NMDA receptor-dependent long-term potentiation at hippocampal-prefrontal cortex synapses in vivo. J Neurosci. 2000;20:RC106. | ||
Onn SP, Wang XB, Lin M, Grace AA. Dopamine D1 and D4 receptor subtypes differentially modulate recurrent excitatory synapses in prefrontal cortical pyramidal neurons. Neuropsychopharmacology. 2006;31:318–338. | ||
Thurley K, Leibold C, Gundlfinger A, et al. Phase precession through synaptic facilitation. Neural Comput. 2008;20:1285–1324. | ||
Kroener S, Chandler LJ, Phillips PE, Seamans JK. Dopamine modulates persistent synaptic activity and enhances the signal-to-noise ratio in the prefrontal cortex. PLoS One. 2009;4:e6507. | ||
Whittington MA, Faulkner HJ, Doheny HC, Traub RD. Neuronal fast oscillations as a target site for psychoactive drugs. Pharmacol Ther. 2000;86:171–190. | ||
Stelzer A, Kay AR, Wong RK. GABAA-receptor function in hippocampal cells is maintained by phosphorylation factors. Science. 1988;241:339–341. | ||
Petrini EM, Barberis A. Diffusion dynamics of synaptic molecules during inhibitory postsynaptic plasticity. Front Cell Neurosci. 2014;8:300. | ||
Marx CE, Yuan P, Kilts JD, et al. Neuroactive steroids, mood stabilizers, and neuroplasticity: alterations following lithium and changes in Bcl-2 knockout mice. Int J Neuropsychopharmacol. 2008;11:547–552. | ||
Tyagarajan SK, Ghosh H, Yevenes GE, et al. Regulation of GABAergic synapse formation and plasticity by GSK3β-dependent phosphorylation of gephyrin. Proc Natl Acad Sci U S A. 2011;108:379–384. | ||
Kalbouneh H, Schlicksupp A, Kirsch J, Kuhse J. Cyclin-dependent kinase 5 is involved in the phosphorylation of gephyrin and clustering of GABAA receptors at inhibitory synapses of hippocampal neurons. PLoS One. 2014;9:e104256. | ||
Charrier C, Machado P, Tweedie-Cullen RY, et al. A crosstalk between β1 and β3 integrins controls glycine receptor and gephyrin trafficking at synapses. Nat Neurosci. 2010;13:1388–1395. | ||
Wuchter J, Beuter S, Treindl F, et al. A comprehensive small interfering RNA screen identifies signaling pathways required for gephyrin clustering. J Neurosci. 2012;32:14821–14834. | ||
McCarley RW, Salisbury DF, Hirayasu Y, et al. Association between smaller left posterior superior temporal gyrus volume on magnetic resonance imaging and smaller left temporal P300 amplitude in first-episode schizophrenia. Arch Gen Psychiatry. 2002;59:321–331. | ||
Fusar-Poli P, Crossley N, Woolley J, et al. Gray matter alterations related to P300 abnormalities in subjects at high risk for psychosis: longitudinal MRI-EEG study. Neuroimage. 2011;55:320–328. | ||
Fusar-Poli P, Crossley N, Woolley J, et al. White matter alterations related to P300 abnormalities in individuals at high risk for psychosis: an MRI-EEG study. J Psychiatry Neurosci. 2011;36:239–248. | ||
Guo Q, Tang Y, Li H, et al. Both volumetry and functional connectivity of Heschl’s gyrus are associated with auditory P300 in first episode schizophrenia. Schizophr Res. 2014;160:57–66. | ||
Babiloni C, Del Percio C, Boccardi M, et al. Occipital sources of resting-state alpha rhythms are related to local gray matter density in subjects with amnesic mild cognitive impairment and Alzheimer’s disease. Neurobiol Aging. 2015;36:556–570. | ||
Yener GG, Emek-Savas DD, Lizio R, et al. Frontal delta event-related oscillations relate to frontal volume in mild cognitive impairment and healthy controls. Int J Psychophysiol. Epub February 7, 2015. | ||
Moller HJ. Bipolar disorder and schizophrenia: distinct illnesses or a continuum? J Clin Psychiatry. 2003;64(Suppl 6):23–27. | ||
Atagün MI, Şikoglu EM, Can SS, et al. Investigation of Heschl’s gyrus and planum temporale in patients with schizophrenia and bipolar disorder: a proton magnetic resonance spectroscopy study. Schizophr Res. 2015;161:202–209. | ||
Ongur D, Drevets WC, Price JL. Glial reduction in the subgenual prefrontal cortex in mood disorders. Proc Natl Acad Sci U S A. 1998;95:13290–13295. | ||
Konopaske GT, Lange N, Coyle JT, Benes FM. Prefrontal cortical dendritic spine pathology in schizophrenia and bipolar disorder. JAMA Psychiatry. 2014;71:1323–1331. | ||
Kim HJ, Thayer SA. Lithium increases synapse formation between hippocampal neurons by depleting phosphoinositides. Mol Pharmacol. 2009;75:1021–1030. | ||
Vita A, De Peri L, Deste G, Sacchetti E. Progressive loss of cortical gray matter in schizophrenia: a meta-analysis and meta-regression of longitudinal MRI studies. Transl Psychiatry. 2012;2:e190. | ||
Fusar-Poli P, Smieskova R, Kempton MJ, et al. Progressive brain changes in schizophrenia related to antipsychotic treatment? A meta-analysis of longitudinal MRI studies. Neurosci Biobehav Rev. 2013;37:1680–1691. | ||
Penttila J, Paillere-Martinot ML, Martinot JL, et al. Cortical folding in patients with bipolar disorder or unipolar depression. J Psychiatry Neurosci. 2009;34:127–135. | ||
Chance SA, Casanova MF, Switala AE, Crow TJ. Auditory cortex asymmetry, altered minicolumn spacing and absence of ageing effects in schizophrenia. Brain. 2008;131:3178–3192. | ||
Savitz JB, Price JL, Drevets WC. Neuropathological and neuromorphometric abnormalities in bipolar disorder: view from the medial prefrontal cortical network. Neurosci Biobehav Rev. 2014;42:132–147. | ||
Moore GJ, Bebchuk JM, Wilds IB, et al. Lithium-induced increase in human brain grey matter. Lancet. 2000;356:1241–1242. | ||
Benedetti F, Poletti S, Radaelli D, et al. Lithium and GSK-3β promoter gene variants influence cortical gray matter volumes in bipolar disorder. Psychopharmacology (Berl). 2015;232:1325–1336. | ||
Rajkowska G, Selemon LD, Goldman-Rakic PS. Neuronal and glial somal size in the prefrontal cortex: a postmortem morphometric study of schizophrenia and Huntington disease. Arch Gen Psychiatry. 1998;55:215–224. | ||
Sweet RA, Henteleff RA, Zhang W, et al. Reduced dendritic spine density in auditory cortex of subjects with schizophrenia. Neuropsychopharmacology. 2009;34:374–389. | ||
Newton SS, Duman RS. Neurogenic actions of atypical antipsychotic drugs and therapeutic implications. CNS Drugs. 2007;21:715–725. | ||
Park SW, Lee JG, Seo MK, et al. Effects of mood-stabilizing drugs on dendritic outgrowth and synaptic protein levels in primary hippocampal neurons. Bipolar Disord. 2015;17:278–290. | ||
Chen G, Rajkowska G, Du F, et al. Enhancement of hippocampal neurogenesis by lithium. J Neurochem. 2000;75:1729–1734. | ||
Son H, Yu IT, Hwang SJ, et al. Lithium enhances long-term potentiation independently of hippocampal neurogenesis in the rat dentate gyrus. J Neurochem. 2003;85:872–881. | ||
Cesca F, Baldelli P, Valtorta F, Benfenati F. The synapsins: key actors of synapse function and plasticity. Prog Neurobiol. 2010;91:313–348. | ||
Vawter MP, Thatcher L, Usen N, et al. Reduction of synapsin in the hippocampus of patients with bipolar disorder and schizophrenia. Mol Psychiatry. 2002;7:571–578. | ||
Cruceanu C, Alda M, Grof P, et al. Synapsin II is involved in the molecular pathway of lithium treatment in bipolar disorder. PLoS One. 2012;7:e32680. | ||
Coyle JT. Glutamate and schizophrenia: beyond the dopamine hypothesis. Cell Mol Neurobiol. 2006;26:365–384. | ||
Meffre D, Grenier J, Bernard S, et al. Wnt and lithium: a common destiny in the therapy of nervous system pathologies? Cell Mol Life Sci. 2014;71:1123–1148. | ||
Gleeson JG, Walsh CA. Neuronal migration disorders: from genetic diseases to developmental mechanisms. Trends Neurosci. 2000;23:352–359. | ||
Jorda EG, Verdaguer E, Canudas AM, et al. Implication of cyclin-dependent kinase 5 in the neuroprotective properties of lithium. Neuroscience. 2005;134:1001–1011. | ||
Narayanan B, Ethridge LE, O’Neil K, et al. Genetic sources of subcomponents of event-related potential in the dimension of psychosis analyzed from the B-SNIP study. Am J Psychiatry. 2015;172:466–478. | ||
Traub RD. Neuronal Networks of the Hippocampus. New York, NY: Cambridge University Press; 1991. | ||
Llinas RR. The intrinsic electrophysiological properties of mammalian neurons: insights into central nervous system function. Science. 1988;242:1654–1664. | ||
Butler-Munro C, Coddington EJ, Shirley CH, Heyward PM. Lithium modulates cortical excitability in vitro. Brain Res. 2010;1352:50–60. | ||
Farhy Tselnicker I, Tsemakhovich V, Rishal I, et al. Dual regulation of G proteins and the G-protein-activated K+ channels by lithium. Proc Natl Acad Sci U S A. 2014;111:5018–5023. | ||
Sourial-Bassillious N, Rydelius PA, Aperia A, Aizman O. Glutamate-mediated calcium signaling: a potential target for lithium action. Neuroscience. 2009;161:1126–1134. | ||
Basar E, Basar-Eroglu C, Karakas S, Schurmann M. Gamma, alpha, delta, and theta oscillations govern cognitive processes. Int J Psychophysiol. 2001;39:241–248. | ||
Atagün MI, Balaban OD, Yesilbas D, et al. Effect of lateralization on motor and mental speed in bipolar disorder. Klin Psikofarmakol B. 2012;22:297–306. | ||
Bora E, Yucel M, Pantelis C. Cognitive endophenotypes of bipolar disorder: a meta-analysis of neuropsychological deficits in euthymic patients and their first-degree relatives. J Affect Disord. 2009;113:1–20. | ||
Wingo AP, Wingo TS, Harvey PD, Baldessarini RJ. Effects of lithium on cognitive performance: a meta-analysis. J Clin Psychiatry. 2009;70:1588–1597. | ||
Atagün MI, Balaban OD, Lordoglu DY, Evren C. Lithium and valproate may affect motor and sensory speed in patients with bipolar disorder. Klin Psikofarmakol B. 2013;23:305–314. | ||
Traub RD, Whittington MA, Buhl EH, et al. On the mechanism of the γ → β frequency shift in neuronal oscillations induced in rat hippocampal slices by tetanic stimulation. J Neurosci. 1999;19:1088–1105. | ||
Whittington MA, Jefferys JG, Traub RD. Effects of intravenous anaesthetic agents on fast inhibitory oscillations in the rat hippocampus in vitro. Br J Pharmacol. 1996;118:1977–1986. | ||
Steullet P, Cabungcal JH, Cuenod M, Do KQ. Fast oscillatory activity in the anterior cingulate cortex: dopaminergic modulation and effect of perineuronal net loss. Front Cell Neurosci. 2014;8:244. | ||
Bartzokis G. Neuroglialpharmacology: white matter pathophysiologies and psychiatric treatments. Front Biosci (Landmark Ed). 2011;16:2695–2733. | ||
Takahashi N, Sakurai T, Davis KL, Buxbaum JD. Linking oligodendrocyte and myelin dysfunction to neurocircuitry abnormalities in schizophrenia. Prog Neurobiol. 2011;93:13–24. | ||
Ren Y, Wang H, Xiao L. Improving myelin/oligodendrocyte-related dysfunction: a new mechanism of antipsychotics in the treatment of schizophrenia? Int J Neuropsychopharmacol. 2013;16:691–700. | ||
Meffre D, Massaad C, Grenier J. Lithium chloride stimulates PLP and MBP expression in oligodendrocytes via Wnt/β-catenin and Akt/CREB pathways. Neuroscience. 2015;284:962–971. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.