Back to Archived Journals » Virus Adaptation and Treatment » Volume 9
Bovine leukemia virus: current perspectives
Authors Juliarena MA, Barrios CN, Lützelschwab CM, Esteban EN, Gutiérrez SE
Received 17 April 2017
Accepted for publication 15 July 2017
Published 10 August 2017 Volume 2017:9 Pages 13—26
DOI https://doi.org/10.2147/VAAT.S113947
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Jonathan Dinman
Marcela Alicia Juliarena,1 Clarisa Natalia Barrios,1 Claudia María Lützelschwab,1 Eduardo Néstor Esteban,2 Silvina Elena Gutiérrez1
1Department of Animal Health and Preventive Medicine, Veterinary Research Center of Tandil (CIVETAN), CIC-CONICET, Faculty of Veterinary Science, National University of the Center of Buenos Aires Province, Tandil, Argentina; 2BIOALPINA Program (GENIAL/COTANA), Colonia Alpina, Argentina
Abstract: Enzootic bovine leukosis, caused by bovine leukemia virus (BLV), is the most common neoplasm of dairy cattle. Although beef and dairy cattle are susceptible to BLV infection and BLV-associated lymphosarcoma, the disease is more commonly detected in dairy herds, mostly because of the management practices in dairy farms. The pathogenicity of BLV in its natural host, the bovine, depends mainly on the resistance/susceptibility genetics of the animal. The majority of infected cattle are asymptomatic, promoting the extremely high dissemination rate of BLV in many bovine populations. The important productive losses caused by the BLV, added to the health risk of maintaining populations with a high prevalence of infection with a retrovirus, generates the need to implement control measures. Different strategies to control the virus have been attempted. The most effective approach is to identify and cull the totality of infected cattle in the herd. However, this approach is not suitable for herds with high prevalence of infection. At present, no treatment or vaccine has proven effective for the control of BLV. Thus far, the genetic selection of resistant animals emerges as a natural strategy for the containment of the BLV dissemination. In natural conditions, most of the infected, resistant cattle can control the infection, and therefore do not pass the virus to other animals, gradually decreasing the prevalence of the herd.
Keywords: bovine leukemia virus, control, genetic resistance, BoLA-DRB3
Introduction
Bovine leukemia virus (BLV), the causative agent of enzootic bovine leukosis, is an exogenous Deltaretrovirus in the Retroviridae family. BLV is closely related to the human T-cell leukemia viruses (HTLV I to III) and simian T-cell leukemia viruses (STLVs), some of them are also associated with proliferative or neurologic diseases of human and nonhuman primates.1,2
Infection by BLV naturally occurs mainly in cattle, but also in zebus, buffalos, and capybaras. Other species such as sheep, goats, and rabbits can be experimentally infected.3
BLV infection is widely distributed in cattle-raising countries. An assessment of BLV infection in USA dairy operations in 2007 showed that 83.9% of them were seropositive for BLV.4 A more recent survey from cattle presented at slaughter in USA in the period 2014–2015 resulted in 38.6% BLV seropositive samples, with significantly higher infection rates for dairy compared to beef cattle.5 A national study from Canada in 1980 showed that 40% of its dairy herds and 11% of its beef herds were infected.6 In Argentina, 84% of the dairy herds were infected in 2001.7 On the other hand, most countries of the European Union, including France, Germany, Sweden, Spain, Denmark, Switzerland, United Kingdom, Belgium, and many others, are officially free of BLV. Other countries like Italy, Poland, and Portugal have extensive regions of their territory officially free of BLV, while the infection is restricted to small areas.8 The virus has been eliminated from dairy herds in Australia, but beef cattle remain infected at very low prevalences.9
BLV is a single-stranded diploid RNA virus which contains genetic information for structural proteins and enzymes (gag, env, reverse transcriptase, protease, and integrase). Three proteins: nucleocapsid, capsid, and matrix are produced from the Gag transcript, while the env gene encodes for surface and transmembrane proteins. The surface and transmembrane proteins work in a coordinated manner to accomplish binding and fusion to cellular membrane receptors during viral entry.10 The BLV provirus also encodes additional accessory genes from the pX region of the genome, which are transcribed by alternative splicing: Tax, Rex, and the less abundant R3 and G4 proteins. These nonstructural proteins have important influence in virus–host cell interactions. Tax and Rex are modulators of viral and cellular gene expression at transcriptional and posttranscriptional level. The other nonstructural proteins R3 and G4 influence in vivo replication and pathogenesis.11 Despite the virtual absence of expression of genomic and subgenomic transcripts from the 5′LTR, BLV abundantly expresses a cluster of RNA polymerase III-transcribed microRNAs (miRNAs).12 BLV miRNAs are not only expressed in tumors but also in the asymptomatic phase of the infection.13 Although dispensable for infectivity, miRNAs have been shown to modify at least six target genes related to apoptosis, immunity, cell signaling, and oncogenesis.14 It has been recently shown that BLV constitutively expresses antisense transcripts from the 3′LTR in all leukemic and asymptomatic carriers examined, reinforcing the concept that BLV provirus is far from silent. Although the precise role these RNA species play in the biology of BLV has not been fully explained, it seems that antisense transcripts may have a long noncoding RNA-like role, and hence may be silencing the 5′LTR. Transcriptional interference between antisense RNA and miRNA suggest a shared role in the regulation of BLV.15 The recent identification of BLV antisense transcripts taken in conjunction with the identification of the BLV microRNAs represent a major shift in our understanding of BLV pathogenesis. BLV and HTLV-1 are preferentially integrated near cancer drivers, altering the expression of host genes by different mechanisms, including the transcription of antisense chimeric viral–host RNAs. It seems that the cis-perturbation of cancer drivers located in the vicinity of the provirus is a major determinant of early clonal expansion in BLV-induced leukemogenesis.16
Biological properties of BLV and initial events following BLV infection
The main cellular target of BLV is the B lymphocyte, although other cell types such as monocytes, CD8 T-lymphocytes, and granulocytes could also be infected.3 Once BLV infects a cell, the RNA genome is copied into DNA by the virus-encoded reverse transcriptase. After experimental infection, an early and intense viral replication occurs by expression of virions, infection of target lymphocytes, reverse transcription, and integration of the provirus into the host genome (also known as the infectious cycle). Multiple clones generated at this early phase of the infection contribute to the peak in proviral load, which reaches maximal level at 4–8 weeks postinfection. Once the cellular and humoral specific immune responses are elicited, a massive depletion of these initial clones occurs, and the proviral load significantly decreases. The infection then spreads through clonal expansion of infected host cells, without evidence for reverse transcription.17–19 Specific antibodies are mainly directed toward the structural envelope gp51 and capsid p24 proteins. These antiviral activities persist throughout the animal’s life, indicating that the immune system is permanently stimulated by BLV antigens.17
A hallmark of BLV infection is the lack of viral protein expression at all stages of the disease. In fact, B lymphocytes harboring an integrated provirus do not produce in vivo detectable levels of protein or viral RNA.20 This fact, also observed in the HTLV-1, is related to suboptimal enhancers located in the LTR promoter. When these suboptimal enhancers are converted into consensus sequences by mutation, the basal transcriptional activity from the promoter is strongly increased, but viral replication in vivo is significantly reduced. It seems that these imperfect enhancers have been evolutionary selected by Deltaretroviruses to repress viral expression, which allows them to be maintained in their hosts facing also a strong adaptive immune response.21 However, once these cells are isolated and cultured in vitro, the provirus is de-repressed and a rapid increase in viral expression occurs, indicating that the provirus is maintained at a repressed stage in vivo.22 Acetylation of histones has been shown to induce the expression of BLV in vitro,23 and latency has also been associated to methylation of the BLV promoter,24 indicating that epigenetic factors may play an important role in the regulation of viral expression.
Pathology induced by BLV in its natural host
Throughout evolution, the relationship between BLV and its natural host, the bovine, has evolved toward minimal pathogenicity, resulting in the majority of BLV-infected cattle without clinical symptoms of disease. BLV infection fits well the “iceberg principle,” typical of many viral diseases. The majority (approximately 70%) of infected cattle are asymptomatic, while one-third of infected cattle develop a permanent and relatively stable increase in the number of B lymphocytes in peripheral blood, termed persistent lymphocytosis (PL).22 PL is the result of a disruption of the B cell homeostasis, which is a complex balance between proliferation and apoptosis rates. As a consequence of impaired cell proliferation and decreased cell death, the B cell turnover is reduced in PL cattle, which finally leads to the accumulation of infected cells in blood.25 The emergence of the tip of the iceberg, ie, the tumoral disease, occurs in about 1%–5% of the infected animals.26 The pathologic condition results from the accumulation of transformed lymphocytes in one or more organs, after a long latency period of 1–8 years. Lesions appear as white, firm tumor masses or as a diffuse tissue infiltrate in any organ; however, the abomasum, heart, visceral and peripheral lymph nodes, spleen, uterus, and kidneys are most frequently affected. The development of these tumors within essential organs leads to a series of functional defects that are ultimately incompatible with survival.27
Hematologically normal BLV-infected cattle (ie, those which do not develop PL or lymphosarcoma), which represent about 70% of infected cattle, comprise at least two groups which can be differentiated in terms of their proviral load in peripheral blood and antibody titers against the BLV major antigens. Therefore, upon infection with BLV, some cattle develop high proviral load (HPL) in peripheral blood (>100,000 BLV proviral copies/µg DNA) and high antibody titers against the most antigenic protein of BLV, ie, gp51. These animals are classified as HPL animals and are not statistically different from PL cattle in terms of proviral load in peripheral blood or humoral immune response to BLV. The other group of non-PL cattle, encompassing about 60% of non-PL cattle, comprises those animals which develop very low proviral load (LPL) in peripheral blood after BLV infection and low humoral immune response against BLV major antigens. LPL cattle usually harbor <100 BLV proviral copies in peripheral blood, which is often undetectable by the molecular methods routinely used (polymerase chain reaction [PCR] and real-time PCR). The low anti-BLV p24 antibody response is characteristic of LPL status, as these specific antibodies are undetectable in most LPL cattle, or are developed at very low titers28 (Table 1).
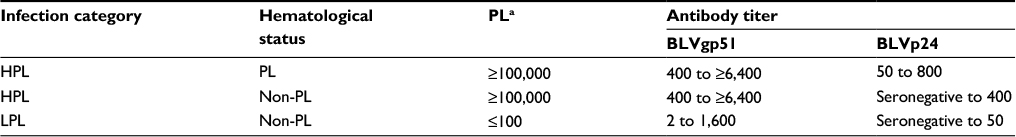 | Table 1 Classification of BLV-infected cattle in terms of hematological status and proviral load Notes: aProviral copies/μg DNA. Copyright ©2007. American Veterinary Medical Association. Adapted from Juliarena MA, Gutierrez SE, Ceriani C. Determination of proviral load in bovine leukemia virus-infected cattle with and without lymphocytosis. Am J Vet Res. 2007;68(11):1220–1225.28 Abbreviations: BLV, bovine leukemia virus; HPL, high proviral load; LPL, low proviral load; PL, persistent lymphocytosis. |
Is BLV a human pathogen? The zoonotic potential of BLV
The fact that BLV or BLV-infected cells are present in the milk of most naturally infected cows indicate that humans are often orally exposed to BLV.29 The first evidence of a possible implication of BLV in public health came from a study carried out in the 1970s, just a few years after BLV was discovered, in which 2 out of 6 chimpanzees fed from birth with unpasteurized milk from cows naturally infected with BLV, developed a fatal erythroleukemia.30
As pasteurization of milk completely inactivates BLV and BLV-infected cells,31 farm dwellers, who may take the milk from the supply before pasteurization is carried out, should be critically exposed to BLV. Therefore, the measurement of occurrence of disease in persons living on farms is critical in epidemiological studies aimed at evidencing association between BLV and human disease.
Initial extensive epidemiological studies carried out in USA, Denmark, and Sweden failed to demonstrate an association between human leukemia and bovine leukosis. Seroepidemiological studies also failed to detect anti-BLV antibodies in humans with different exposure to BLV (reviewed by Johnson32). The absence of BLV-specific sequences in 157 cases of childhood acute lymphoblastic leukemia or non-Hodgkin’s lymphoma and 136 controls in USA33 and in 517 cases of human leukemia and 162 lung cancer patients in Korea34 was further evidence against the implication of BLV in human disease.
Concern about the possibility of BLV linked to a human disease has reemerged because of findings of recent studies. In a first report, Buehring et al35 found reactivity to BLVp24 protein in immunoblots of 74% human sera and argued that serological methods used in initial studies were not sensitive enough to detect BLV-specific antibodies in humans. Although less than 10% of the people with specific antibodies stated direct contact with bovines or their biological products, the authors conclude that antibodies in humans may have been raised in response to oral exposure by consumption of heat-denatured BLV in bovine foodstuffs, or may be the consequence of human infection by BLV.35 Other studies have shown that sera from humans infected with HTLV-1 and 2 cross-react with BLVp24, due to an epitope shared by the related viruses.36–38
Further evidence for the possible transmission of BLV from cattle to humans was the detection of BLV sequences by PCR in 44% of breast tissues, and the most striking finding was BLVp24 expression detected by immunohistochemistry on the secretory epithelium of the mammary gland.39 A subsequent case–control study showed BLV sequences in samples from breast cancer patients being at a level twice that in normal breast tissue sections, concluding that the presence of BLV in breast tissues was associated with breast cancer.40
Although the precedent cited studies suggested an association between BLV and breast cancer, the evidence is still controversial. Neither anti-BLV antibodies nor BLV sequences were found by sensitive enzyme-linked immunosorbent assay or PCR in either healthy or breast cancer Chinese women.41 Recent studies with the more sensitive whole genome sequencing, however, did not support the previous evidence of association between BLV and human breast cancer. None of 32 billion sequencing reads retrieved from 51 breast cancers mapped on different strains of BLV.42 Additionally, extensive studies from RNA-seq data of more than 3,700 malignant human tumors, including 810 human breast adenocarcinomas, do not support the previous evidence of BLV expression in breast tissue.43,44
Independent of the lack of conclusive evidence for BLV as a human pathogen, the current concern about BLV should be a good opportunity to address the control and eradication of this prevalent oncogenic retrovirus from cattle herds.
Transmission
Natural or iatrogenic horizontal transmission of BLV is the major route of spread in most settings, primarily involving the transfer of allogeneic infected cells, as cell-free virus is rarely detected in vivo.45 Upon contact with susceptible cattle, infected lymphocytes from blood, secretions, or excretions of infected animals containing viable lymphocytes may potentially transmit the infection. Vertical transmission may occur by transplacental infection after immune competence is established (third month of gestation), although at low rates (between 3% and 8%).45,46 Postnatal vertical infection may occur from feeding calves with milk or colostrum from infected dams; however, specific antibodies obtained by natural passive transfer confer protection to calves.47 Iatrogenic procedures involving the contact of blood, when practiced without proper disinfection of instruments, are associated with BLV transmission. As infectivity is dependent on the number of infected lymphocytes contained in the injected blood, cows with PL and HPL are more efficient transmitters. On the other hand, LPL cattle, carrying an exiguous number of infected lymphocytes in peripheral blood, have been proven not to transmit the infection to adjacent uninfected cattle in natural conditions of management.48 Evidence has been reported on the role of blood sucking insects in the transmission of BLV, and the beneficial effect of insect vector control.49
Impact of BLV infection on production traits
The economic losses caused by the BLV were underestimated for many years, especially in countries of the American continent, without implementing BLV control measures. Therefore, most of these countries still face the burden of this disease due to the high prevalence of infection among cattle, and the economic cost that this entails.
The situation is different in Western European, where eradication programs were employed leading to negligible rates of BLV infection. BLV-free countries have begun to apply trade barriers to the import of infected livestock, which has a huge impact on the dairy industry.
The most obvious economic losses caused by the BLV are the loss on the cow’s production potential and the shortening of its life span due to death by lymphosarcoma/leukemia. Considering that nearly 5% of infected animals die during the period of 3–8 years of age and the current prevalence of infection in this age category of cows, the losses to the dairy sector are of the order of millions of dollars at the regional level. However, the economic cost is usually underestimated due to the fact that clinical signs are vague and the death by lymphosarcoma/leukemia is easily subdiagnosed.50 In addition, the indirect economic losses – such as the cost of replacing an animal in production, the diagnostic and veterinary care, and the loss of a calf and milk production over about 10 months – are generally not taken into account.51
The effect of BLV infection on production traits in dairy cows has been studied essentially by using two approaches: cow- and herd-level designs. In cow-level experimental design, study animals are classified according to their BLV infection status, and the production of both groups (BLV positive vs BLV negative) is compared. In herd-level experimental design, linear association is made between the prevalence of BLV infection and the production traits of the different herds in the study. This variation in experimental designs makes it difficult to compare the results of the different studies.
In early observations, it was found that BLV-infected cows had higher milk production than noninfected cows;52–54 however the genetic potential for milk production was significantly greater in seropositive cows.52 Therefore, the higher production of an animal was considered a risk factor for acquiring infection, due to the nature of the spread of BLV and management practices of most dairy herds.53 Later investigations were unable to find association between BLV infection and milk production traits,55–58 probably because of the difficulty to establish the date of BLV infection.59
Recent reports showed a negative effect of BLV infection on milk production. An inverse correlation between the prevalence of BLV infection and the milk production was found at the herd level in two studies.
In both cases, a linear association between higher BLV prevalence and decreased herd-level milk production was found, resulting in 115–218 kg milk loss/cow/yr (between 2.7% and 1% loss production/cow/yr) for every 10% increase in BLV infection prevalence.59,60 Similar results were obtained at cow level, with a decrease in milk production of 1.5%/infected cow/yr.61
Moreover, to reduce the possible influence of confounding factors, the animals were classified according to the number of calving and the lactation stage (early, middle, and late lactation). BLV-infected cows with ≥4 parities in their early and middle lactation stage had 13.2% and 14.9% reduction rate, respectively, compared with noninfected cows.62 Another study analyzed the lactation history and BLV infection status of 4,052 cows from 348 herds and showed that infected cows in their 2nd and 3rd lactations had significantly lower life milk production compared with their negative counterparts with the same number of lactations. No significant differences were found between infected and noninfected cows with >3 lactations.63
In spite of the more accurate observation obtained by considering the stage and the number of lactations, other factors like the intrinsic genetic merit of the animals should be considered as the reason for the differences encountered among the different studies.
The different phenotypes developed by BLV-infected animals can also influence on the effect on production traits. Cows with PL have an estimated milk loss production of 3%.64 Analysis of the effect of BLV infection on production in animals classified according to LPL and HPL phenotypes is in progress.
Furthermore, it has been reported that BLV infection is strongly associated with an increase in the somatic cell count (SCC) in milk, particularly in cows with >4 lactations.62 This increase of SCC may indicate that BLV-infected cattle could present higher incidence of mastitis than BLV-free animals. A high SCC in milk is undesirable for the dairy industry because it increases the acidity of milk and decreases the quality of cheese.
In conclusion, BLV infection can affect the productivity of the herd. The differences found in the studies of the influence of BLV infection on herd productivity could be due to the difficulty in determining the onset of BLV infection, the differences in the experimental designs used, the indirect effects of other diseases, and the size and the genetic background of the studied population.
Potential therapeutic treatment for BLV-infected cows
One feature of the BLV infection is that the virus is maintained in a silent state within the infected cells and, in this manner, is not detected by the immune system. Much effort has been put into identification of compounds capable of reversing the BLV latency to render infected cells susceptible to immune clearance.
The complexity of regulating gene expression in eukaryotic cells is achieved primarily by the joint operation of several different transcriptional regulatory proteins. Furthermore, the packaging of DNA into chromatin and its modification by methylation are key steps in the regulation of gene expression. One of the mechanisms involved in the remodeling of chromatin is the posttranslational modification of histones by acetylation. Acetylation occurs through the action of enzymes with histone acetyltransferase activity (HAT), and, conversely, removal of acetyl groups is mediated by a group of enzymes, the histone deacetylases (HDAC). Oversimplifying the mechanism involves HAT-mediated acetylation, which neutralizes the lysine located within the N-termini tail of histones, thus decreasing their affinity for DNA, thereby provoking the unfolding of the nucleosomes. This conformational change favors the access of transcription factors (for the complete description of the mechanism of action of HAT and HDAC, Grunstein65). Deacetylation, on the other hand, is frequently followed by histone methylation, forming highly repressive structures such as heterochromatin. Therefore, preventing the deacetylation of histones by inhibition of HDAC contributes to the activation of gene transcription.
A number of chemical substances have been found to possess HDAC-inhibitory effect with varying efficiencies and selectivity over the four different classes of HDACs.
HDAC inhibitors (HDACi) have gained much attention as therapeutic drugs for cancer and noncancer diseases (reviewed by Rotili et al,66 Beumer and Tawbi,67 De Souza and Chatterji68). Already in 1996, Moog and coworkers reported that valproic acid (VPA), a weak HDACi, markedly increased the HIV replication in various infected cell lines and primary cell cultures.69 Later, Chen et al70 were able to prove a dramatic reactivation of gene expression after treatment with trapoxin, a strong HDACi, in a transfected cell line carrying a silent, integrated copy of a virally transduced gene. By forcing the viral expression, a new potential therapeutic approach emerged to decrease the pool of latent retrovirus reservoirs. Furthermore, HDACis were also tested alone or in combination with antiretroviral drugs as therapeutic agents for HIV, STLV, and HTLV infections in preclinical and clinical trials.71–73
Since BLV-infected sheep is considered the animal model of HTLV-1 infection in humans, the studies on the epigenetic control of BLV viral expression in this animal model were aimed at helping delineate new therapeutic strategies for the disease in humans. The potential role of HDAC on the transcriptional control of the BLV expression in vitro and ex vivo was explored. An increase of more than 10 fold in the BLV promoter activity was observed when a cell line transfected with BLV was cultured in the presence of trichostatin A, a potent and specific HDACi. Moreover, the activity of trichostatin A and trapoxin, another HDACi, also modulated the expression of BLV ex vivo in infected peripheral blood mononuclear cells from sheep and cattle.23
Owing to the potential toxicity of the therapeutic dose of TSA in vivo, other HDACi were investigated. Valproate (the sodium salt of VPA), a drug used as anticonvulsant for certain types of seizures, is a weak HDACi. Although teratogenic, valproate exhibits low toxicity in adults and good pharmacokinetic properties in vivo.74 The effectiveness of the valproate therapy as an activator of transcription of the BLV LTR and of p24 synthesis was demonstrated in transient transfection experiments and in short-term cultures of primary B lymphocytes.75 Indeed, treatment with valproate caused a transient increase of proviral load, a decreased in lymphocyte number, and the induction of tumor regression in leukemic sheep.75 After valproate treatment, the authors observed a slow and continuous rate of tumor cell destruction, possibly attributed to the induction of an immune response subsequent to the proviral reactivation. In spite of these encouraging results in the leukemic sheep, the animals remained persistently infected. In a 2-year follow-up study, four out of five leukemic sheep showed variable responses to valproate treatment and, with different kinetics.76 Although the B cell number was restored, and the proviral load initially decreased in these sheep, the treatment discontinuation led to the reappearance of an uncontrollable leukemia and unresponsiveness to further administration of valproate. This refractoriness could be overcome by using other, more potent HDACi or by complementing with another chemotherapeutic drug.76
The use of latency-reversing agents on HTLV-1-infected individuals was attempted in patients suffering from HTLV-1-associated myelopathy/tropical spastic paraparesis, where a long-term reduction in the proviral load would be protective against collateral damage in the central nervous system.77 However, in spite of the initial reduction of proviral load, the clinical trials of VPA treatment in HTLV-1-associated myelopathy/tropical spastic paraparesis patients were proven to be ineffective in permanently reducing the proviral loads after 1–2 year investigation.72,77 Similar results were obtained for HIV-1-infected patients.78–80 It would seem that the combination of drugs provides a more effective reactivation and elimination of the persistently infected cells.81,82 Nevertheless, a more achievable approach would be to better identify the main mechanism(s) causing viral latency, and improve our knowledge on the relative contribution of each of the silencing mechanisms involved. In this way, it will be feasible to target and eliminate the latent virus reservoir with the appropriated drug(s).
Strategies for the control of BLV infection
Experimental vaccines to prevent BLV infection
Because BLV is a retrovirus, prospects for the development of an effective vaccine are challenging. Both cows and sheep develop a strong antiviral immune response to BLV primary infection, but, in the majority of the infected animals, neither humoral nor cell-mediated immune responses against BLV are efficient at eliminating infected cells that carry a transcriptionally silent virus.
Considering that maternal antibodies transferred to the offspring via colostrum protect the newborn from infection during several months, early attempts of immunization were aimed to induce neutralizing antibodies.54,83 Initially, a cell-derived vaccine, harboring a defective provirus expressing mainly env gene products was tested. The cell-based vaccine generated neutralizing antibodies and protected vaccinated sheep, but presented the risk of disease transmission since the provirus could be found after coculture with embryo cells derived from bovine testes.84–86
Inactivated virus vaccine candidates were also assayed, but only a fraction of the vaccinated animals was protected from a low-dose challenge. Curiously, the protected animals showed lower anti-BLV-gp51 and anti-BLV-p24 antibody titers compared to the unprotected ones.87,88 On the other hand, subunit vaccines promoted a humoral immune response but failed to prevent BLV infection in cattle.89,90 The first evidence that cell-mediated immune response could be involved in the suppression of BLV replication came about after vaccination with a BLV-env coding recombinant vaccinia virus (env-rVV), in naïve or BLV-infected animals. Although the env-rVV vaccine did not protect the naïve animals from the BLV infection after challenge, it significantly suppressed the dissemination of BLV in peripheral blood leukocytes. This effect did not correlate with the presence of the neutralizing or anti-BLV antibody titers observed in these animals, suggesting the stimulation of Th cells, possibly of the type 1, without the induction of a specific humoral immune response.91–93 Vaccination of mice and sheep using the rVV coding for BLV gp51 (BLVgp51-rVV) resulted in protective immunity, with the characteristics of a type 1 response.94–98 These observations are in agreement with the notion that animals that control the virus express higher levels of interleukin-12 and interferon-gamma and lower levels of interleukin-10.99–101 However, inoculation of cattle with the rVV-BLVgp51 resulted in a nonprotective type 2 immune response.90
Immunization of sheep with a minimal cytotoxic T-lymphocyte peptide epitope vaccine delayed the appearance of lymphosarcomas.102
Another approach employed was the injection of vectors containing the env and tax genes under the control of the cytomegalovirus103 or the Sr-alpha promoters, using the prime-boost strategy.104 Both DNA vaccines induced a strong immune response and protected the vaccinated animals from the challenge, but they did not prevent later infection, probably owing to short-lived stimulation.
More recently, efforts were directed toward the development of a recombinant live-attenuated BLV vaccine.
In the last decades, a series of mutant proviral clones were engineered. Mutations of the different viral genes contributed to the understanding of the pathogenesis of the disease,105–109 providing tools to analyze their function in vivo.110 Furthermore, these mutant proviruses brought to light their potential suitability as attenuated virus vaccines. The candidates studied included hybrid, simpler retrovirus derivatives from a wild type, lacking the regulatory genes and cis acting response sequences;107,111–113 noninfectious provirus deficient for cell fusion, but with wild-type-like propagation levels;114 and infectious provirus with impaired propagation ability.21,107,108,110,115–117
The perfect vaccine that induces neutralizing antibodies and protective cell-mediated immune responses remains elusive. Since animals are exposed to reinfection during their entire life,118 the pursuit of such aim, achieved mainly by live-attenuated vaccines, will demand not only the constant antigenic stimulus of an innocuous, replicating virus, but the capacity to induce a long-lived protective immune response. From the epidemiological point of view, it should also allow the differentiation between vaccinated and infected animals, and, most importantly, it must be safe.
A claim of an achievement of a successful, attenuated vaccine involving a multiple genetically modified provirus has arisen in the last years.119 It is likely that this recombinant provirus is both mutated on the immunoreceptor tyrosine activation motifs cytoplasmic tail of the BLV-gp30 glycoprotein gene and has the G4 and R3 genes deleted.115 However, no proof of concept has been shown, nor has preliminary data been published concerning the identity of the provirus mutated/deleted genes, the immune protection conferred to the vaccinated animals in the long term, the safety assessment of the vaccine, and the proposed strategy to be used to differentiate vaccinated from infected animals. The recent evidence on the role of viral miRNA in BLV pathogenesis14 together with the finding that BLV proviruses are integrated near cancer drivers, whose expression is perturbed by the integrated proviruses,16 further emphasizes the need of caution on retroviral live-attenuated vaccines. It should also be borne in mind that the use of an attenuated retroviral vaccine implies a risk for disruption of genes upon insertion into the cellular genome, preventing their expression, and there is also the risk of reversion to virulent, oncogenic forms in the long run.120,121 In the case of BLV, the transformation event that leads to neoplasia is believed to take place at the time of infection, so even if an attenuated virus did not result in persistent infection, lymphosarcoma could still develop.17,116 Moreover, although rare, the possibility exists that the restoration of virulent variants can occur by transcomplementation with a wild-type strain during natural exposure or superinfection with wild-type virus, leading to a prolonged incubation time and a delayed manifestation of disease.110,122–124.
In summary, to date, no BLV vaccine is available and a suitable BLV preventive vaccine will take many years to materialize. As it is the case for other retroviral diseases, the virus capacity to evade adaptive immune responses, the early establishment of latent viral reservoirs, and the lack of clear immune correlates of protection represent unprecedented challenges for vaccine development. The vaccination of animals with live-attenuated retrovirus should be analyzed with caution.
As in the case of other retroviruses, the historical failure of other vaccines to induce a sterilizing immunity and provoke a latent infection must be considered. The oncogenic potential of integrating viruses and the reversion to pathogenic forms are the most obvious risks. On the other hand, persistent infection of animals vaccinated with an attenuated strain could lead to restrictions on trade in live cattle and dairy byproducts due to the intrinsic risk associated with a spread of a genetically engineered pathogen. The possibility of introducing a genetically modified live retrovirus that would potentially disseminate and evolve, even to a limited extent, within the herds entails a safety concern and an ethical problem.
Until a suitable vaccine arrives, other strategies should be considered to control the virus dissemination among herds.
Traditional BLV control approach
In this section, different strategies are considered for the control of BLV. The option chosen depends mainly on the herd infection prevalence, the animals’ value, and whether a governmental indemnity is offered to producers for culling seropositive cows.
The first step is to calculate the herd prevalence. A good approach is to establish the “herd profile,” by grouping animals in their 1st, 2nd, 3rd, and 4th or more lactations, and sample 10 cattle of each group. This method enables the estimation of lactation-specific BLV prevalence, which can help to identify areas of potential risk for BLV transmission.59
Once the prevalence is estimated, the information is used to select one of the following control methods. The most effective approach is to identify and cull the totality of infected cattle in the herd. The presence of BLV-specific antibodies in serum or milk is a reliable indicator of infection in cattle older than 6 months, and thus diagnosis is usually based on serological tests. Antibodies to BLVp24 are absent or develop at low titers in infected cattle with LPL. Therefore, the employment of serological tests using the gp51 as the antigen is preferred for diagnosis. Due to the high cost of culling the infected cattle, this approach is suitable for regions or herds with low incidence of the disease, and to be successful, it requires governmental economic compensation policies.125,126
Culling of seropositive cattle of outstanding genetic potential may be resisted by farmers. Embryo transfer from infected dams to seronegative recipients can be an effective means to control BLV transmission in such a situation.127–130
A slow but more economically viable alternative is the implementation of a two herd scheme, by which BLV-infected animals are maintained on the farm, but managed separately, with separate equipment or adequate disinfection of utensils. It requires the routine identification of new infections in the BLV-negative herd, and their transfer to the BLV-positive herd. Thus, the sensitivity of the test routinely used to diagnose the infection is critical. Generally, the gp51 enzyme-linked immunosorbent assay tests detect at least 10% more reactors than the agar-gel immunodiffusion test27 but, the diagnosis should be accompanied by appropriate management measures to minimize BLV transmission127 (Box 1). This approach, although quite demanding, has proved successful to control the transmission and even to eradicate the infection from dairy farms.131,132
 | Box 1 Main management measures aimed at minimizing BLV transmission Abbreviation: BLV, bovine leukemia virus. |
Up to 10% of calves born to infected dams are infected in utero; hence, care should be taken in preserving the negative status of the calves born uninfected to provide BLV-negative herd replacements. Feeding colostrum to newborn calves from its seropositive dam can provide protection for the first months of life;133–135 however, it also may potentially favor transmission. Reduction in the risk of BLV transmission via colostrum can be achieved by heating the colostrums at 63° for 30 min,136 or by feeding calves with colostrum and milk from seronegative cows or milk replacer. Other management practices aimed at reducing the iatrogenic risk of transmission should be considered. As colostral-derived antibodies persist for 6–9 months of age, calves should be tested serologically for BLV infection at this time.
Finally, a more conventional, although controversial, control plan is the implementation of biosafety and management measures aimed at minimizing the within-herd transmission of BLV. The efficacy of this approach can be undermined by human and environmental factors.
Control of BLV infection by genetic selection of resistant cattle: fundamentals and application
The major histocompatibility complex (MHC) plays a central role in the development of immune response against pathogens in all mammalian species. Genes encoding the MHC are highly polymorphic, and numerous associations between allelic variants and immune responsiveness and disease resistance are well documented. The MHC in bovine is termed the bovine leukocyte antigen system (BoLA). The BoLA locus is highly complex and contains about 154 predicted functional genes spanning about 4 centimorgan on chromosome 23 (BTA23). Cattle express one DR gene pair (DRA and DRB3). The coding sequence of DRA is monomorphic, while DRB3 gene is highly polymorphic.137
The polymorphism of MHC genes was associated with resistance or susceptibility to the dissemination of the BLV in the host. Among the MHC genes studied for association with BLV phenotypes, polymorphism in exon 2 of BoLA-DRB3 was demonstrated to be stronger compared to other studied genes.137–140
The nomenclature of BoLA alleles is gen*allele, and two nomenclatures are used to name the different alleles of these genes. One is based on the cut pattern identified by three restriction enzymes (PCR-restriction fragment length polymorphism [RFLP] technique), which can differentiate 57 different alleles. The other nomenclature, which has been adopted by the International Society for Animal Genetics, is based on the amino acid sequence. Currently, the amino acid sequences of 130 DRB3 alleles have been published, some of which are indistinguishable by PCR-RFLP.141
Initial studies showed that the resistance to PL caused by BLV was associated with the presence of BoLA-DRB3*11 allele in Holstein cattle.139 Also, it was demonstrated that BLV-infected cattle selected for the presence of the DRB3*11 allele carried less infected lymphocytes compared to other infected animals.142 The allele *11 (as determined by PCR-RFLP) comprises two variants, BoLA-DRB3*0901 and BoLA-DRB3*0902, that can be identified by sequencing. Our studies of genetic association between the polymorphism of BoLA-DRB3 with the different phenotypes in 230 BLV-infected cattle belonging to seven Holstein herds showed a stronger association of BoLA-DRB3*0902 with the LPL phenotype (odds ratio (OR) =8.24) than allele *11 itself (OR =5.82). Allele BoLA-DRB3*1701 also showed significant association with LPL profile (OR =3.46). The HPL phenotype was significantly associated only with one allele: BoLA-DRB3*1501 (allele *16 as determined by PCR-RFLP) (OR =0.36).143 We then assigned the DRB3 alleles to three categories: resistance (R) if the allele was associated with LPL phenotype, susceptibility (S) if the allele was associated with HPL phenotype, and neutral (N) if the allele was not associated with any phenotype.143 Interestingly, the alleles conferring resistance are the same for Japanese Black and Holstein cattle, but alleles associated with susceptibility differ in both breeds.144
There is a strong association between BoLA-DRB3*0902 and low antibody titers against the two main BLV structural proteins, env gp51 and gag p24. The other BoLA-DRB3 allele associated with resistance, DRB3*1701, is also significantly associated with low antibody titers against these BLV structural proteins.145,146 These animals have shown immune competence against other prevalent pathogens of dairy cattle; hence, the low anti-BLV titers are probably due to a poor antigenic stimulation of the extremely low viral load they carry.
Not all the LPL cattle harbor R alleles; however, the penetrance of R alleles for the LPL phenotype is notably high, particularly for the DRB3*902 (more than 80% cattle with DRB3*902 develop LPL phenotype).143,147 The findings above reviewed suggest that the BoLA DRB3 might influence the host response to BLV infection, or it can merely be the consequence of linkage disequilibrium and the fact that other genes and/or epigenetic or environmental factors might be involved in the regulation of BLV proviral load.
Thus, BoLA-DRB3*0902 allele is an excellent marker for the selection of resistant animals in the current populations of Holstein breed.137,145,147 Its allele frequencies in different Holstein populations are relatively high (5%–15%).143,147–152 BoLA-DRB3*0902 animals with LPL phenotype did not transmit the infection at herd level in natural conditions,48 and under experimental conditions, it was necessary to inoculate a large volume of blood in order to infect sheep.145,147 Furthermore, to date, the BoLA-DRB3*0902 allele has not been associated with susceptibility to other infectious agent, nor has it negatively affected production or reproduction traits.137,153 Moreover, R alleles are associated with resistance to intramammary infection and higher production traits.149,150 Thus, expanding the population of cattle harboring these alleles in order to control BLV infection would additionally increase resistance to mastitis. Identification of cattle carrying the BoLA-DRB3*902 allele can be easily achieved by PCR and real-time PCR.154
Based on the abovementioned results on the BoLA-DRB3*0902 heterozygous animals, we have proposed a BLV control and eradication program based on genetic selection. This program is specially designed for herds with high prevalence of BLV, where the traditional approaches of control are economically unfeasible. The BLV control and eradication program has been designed in two steps: as the first step, HPL cattle should be replaced by cattle harboring the BoLA-DRB3*0902 marker. Once culling of HPL animals has been completed, as the second step, LPL cattle should be replaced by BLV-negative cattle.143 The selected animals for the replacement in the first step should carry the BoLA-DRB3*0902 allele in heterozygosis with other alleles not associated with any BLV phenotype.147 Once all infected cattle in the herd are LPL, any BLV-negative cattle could be used as replacement, no matter which BoLA allele it carries. Finally, it should be mentioned that a potential risk exists in expanding or segregating BoLA genotype-selected populations, due to the relevant role of MHC genes in the immune response, which might increase susceptibility to other infectious agents. Thus, although the *0902 allele has not been negatively associated with any disease, this plan only proposes the genetic selection of cattle until the HPL animals have been replaced in the herd.
In brief, the BLV control and eradication program proposed is based on two main premises: 1) BLV-infected cattle with LPL phenotype do not transmit the virus and 2) the development of LPL phenotype can be predicted by a marker (BoLA-DRB3*0902). To test the first premise and simulate the second step of the proposed control plan, an experiment was conducted in which BLV-infected-LPL cattle carrying the BoLA-DRB3*0902 marker cohabited with BLV-free cattle for 20 months. While no BLV-free cattle became infected in this experimental herd, new infections occurred in other herds of the region.48 This situation simulates the second step of the proposed control plan.
Preliminary data from a commercial dairy farm with 72.7% infection prevalence (63.3% of HPL infected animals) have provided support for the first step of the control plan. The increase of the allelic frequency of BoLA-DRB3*0902 in this herd resulted in an increase in the percentage of LPL infected animals.
One of the main limitations for the widespread application of the proposed control plan is the availability of breeders with the desirable productive and reproductive traits, which also carry the selection marker. Although the number of bulls with the genetic marker available has increased in recent years, to increase the allelic frequency of BoLA-DRB3*902 in the population while maintaining the genetic variability of the herds, it would be necessary for more bulls to carry this allele. The allelic frequency of BoLA-DRB*902 is relatively high, and its pattern of inheritance is codominant, facilitating the expansion of its frequency by directed crossbreeding or artificial insemination. The growing demand of semen from bulls carrying the BoLA-DRB3*0902 marker is likely to generate much interest in the international market of semen.
Concluding remarks
Due to the considerable increase of BLV prevalence in cattle populations in certain areas of the world, there is growing concern about its negative effects on cattle health and production and its potential hazard on human health. Currently, no vaccine or treatment has proven to be effective. For many countries, the opportunity to implement “the test and cull” scheme of control has passed. The selection of cattle carrying the BoLA-DRB3*0902 marker emerges as a natural strategy for the containment of the BLV dissemination.
Disclosure
The authors report no conflicts of interest in this work.
References
Maclachlan NJ, Dubovi EJ. Retroviridae. In: Press A, editor. Fenner’s Veterinary Virology. 4th ed: Elsevier Science; 2011:243–274. | ||
Barbeau B, Hiscott J, Bazarbachi A, et al. Conference highlights of the 16th International Conference on Human Retrovirology: HTLV and related retroviruses, 26–30 June 2013, Montreal, Canada. Retrovirology. 2014;11:19. | ||
Schwartz I, Levy D. Pathobiology of bovine leukemia virus. Vet Res. 1994;25(6):521–536. | ||
USDA. Bovine Leukosis Virus (BLV) on U.S. Dairy Operations, 2007; 2008. | ||
Bauermann FV, Ridpath JF, Dargatz DA. Bovine leukemia virus seroprevalence among cattle presented for slaughter in the United States. J Vet Diagn Invest. 2017. | ||
Samagh BS, Kellar JA. Seroepidemiological survey of bovine leukaemia virus infection in Canadian cattle. Paper presented at: Fourth International Symposium on Bovine Leukosis; 1982; Luxembourg. | ||
Trono KG, Perez-Filgueira DM, Duffy S, Borca MV, Carrillo C. Seroprevalence of bovine leukemia virus in dairy cattle in Argentina: comparison of sensitivity and specificity of different detection methods. Vet Microbiol. 2001;83(3):235–248. | ||
European Commission. Bovine and swine diseases. Annual Report. 2014. | ||
Voges H. Reports from Industry Surveillance and Disease Control Programmes: New Zealand Dairy Enzootic Bovine Leukosis (EBL) Control Scheme SURVEILLANCE 38 37. 2011. Available from http://www.sciquest.org.nz/elibrary/download/72863/New_Zealand_Dairy_Enzootic_Bovine_Leukosis_%28EBL%29_C.pdf? Accessed July 11, 2017. | ||
Lairmore MD. Animal models of bovine leukemia virus and human T-lymphotrophic virus type-1: insights in transmission and pathogenesis. Annu Rev Anim Biosci. 2014;2:189–208. | ||
Willems L, Kerkhofs P, Dequiedt F, et al. Attenuation of bovine leukemia virus by deletion of R3 and G4 open reading frames. Proc Natl Acad Sci U S A. 1994;91(24):11532–11536. | ||
Kincaid RP, Burke JM, Sullivan CS. RNA virus microRNA that mimics a B-cell oncomiR. Proc Natl Acad Sci U SA. 2012;109(8):3077–3082. | ||
Rosewick N, Momont M, Durkin K, et al. Deep sequencing reveals abundant noncanonical retroviral microRNAs in B-cell leukemia/lymphoma. Proc Natl Acad Sci U S A. 2013;110(6):2306–2311. | ||
Gillet NA, Hamaidia M, de Brogniez A, et al. Bovine leukemia virus small noncoding RNAs are functional elements that regulate replication and contribute to oncogenesis in vivo. PLoS Pathog. 2016;12(4):e1005588. | ||
Durkin K, Rosewick N, Artesi M, et al. Characterization of novel bovine leukemia virus (BLV) antisense transcripts by deep sequencing reveals constitutive expression in tumors and transcriptional interaction with viral microRNAs. Retrovirology. 2016;13(1):33. | ||
Rosewick N, Durkin K, Artesi M, et al. Cis-perturbation of cancer drivers by the HTLV-1/BLV proviruses is an early determinant of leukemogenesis. Nat Commun. 2017;8:15264. | ||
Florins A, Gillet N, Asquith B, et al. Cell dynamics and immune response to BLV infection: a unifying model. Front Biosci. 2007;12:1520–1531. | ||
Pomier C, Alcaraz MT, Debacq C, et al. Early and transient reverse transcription during primary deltaretroviral infection of sheep. Retrovirology. 2008;5:16. | ||
Gillet NA, Gutierrez G, Rodriguez SM, et al. Massive depletion of bovine leukemia virus proviral clones located in genomic transcriptionally active sites during primary infection. PLoS Pathog. 2013;9(10):e1003687. | ||
Kettmann R, Marbaix G, Cleuter Y, Portetelle D, Mammerickx M, Burny A. Genomic integration of bovine leukemia provirus and lack of viral RNA expression in the target cells of cattle with different responses to BLV infection. Leuk Res. 1980;4(6):509–519. | ||
Merezak C, Pierreux C, Adam E, et al. Suboptimal enhancer sequences are required for efficient bovine leukemia virus propagation in vivo: implications for viral latency. J Virol. 2001;75(15):6977–6988. | ||
Ferrer JF. Bovine lymphosarcoma. Adv Vet Sci Comp Med. 1980;24:1–68. | ||
Merezak C, Reichert M, Van Lint C, et al. Inhibition of histone deacetylases induces bovine leukemia virus expression in vitro and in vivo. J Virol. 2002;76(10):5034–5042. | ||
Pierard V, Guiguen A, Colin L, et al. DNA cytosine methylation in the bovine leukemia virus promoter is associated with latency in a lymphoma-derived B-cell line: potential involvement of direct inhibition of cAMP-responsive element (CRE)-binding protein/CRE modulator/activation transcription factor binding. J Biol Chem. 2010;285(25):19434–19449. | ||
Debacq C, Asquith B, Reichert M, Burny A, Kettmann R, Willems L. Reduced cell turnover in bovine leukemia virus-infected, persistently lymphocytotic cattle. J Virol. 2003;77(24):13073–13083. | ||
Ferrer JF, Marshak RR, Abt DA, Kenyon SJ. Relationship between lymphosarcoma and persistent lymphocytosis in cattle: a review. J Am Vet Med Assoc. 1979;175(7):705–708. | ||
Radostitis O, Gay C, Hinchcliff K, Constable P. Disease associated with viruses and chlamydia I. In: Elsevier, editor. Veterinary Medicine. 10th ed. Elsevier; 2007:1209–1221. | ||
Juliarena MA, Gutierrez SE, Ceriani C. Determination of proviral load in bovine leukemia virus-infected cattle with and without lymphocytosis. Am J Vet Res. 2007;68(11):1220–1225. | ||
Ferrer JF, Kenyon SJ, Gupta P. Milk of dairy cows frequently contains a leukemogenic virus. Science. 1981;213(4511):1014–1016. | ||
McClure HM, Keeling ME, Custer RP, Marshak RR, Abt DA, Ferrer JF. Erythroleukemia in two infant chimpanzees fed milk from cows naturally infected with the bovine C-type virus. Cancer Res. 1974;34(10):2745–2757. | ||
Chung YS, Prior HC, Duffy PF, Rogers RJ, Mackenzie AR. The effect of pasteurisation on bovine leucosis virus-infected milk. Aust Vet J. 1986;63(11):379–380. | ||
Johnson RK. Bovine leukemia virus and enzootic bovine leukosis. Veterinary Bulletin. 1992;62(4):287–312. | ||
Bender AP, Robison LL, Kashmiri SV, et al. No involvement of bovine leukemia virus in childhood acute lymphoblastic leukemia and non-Hodgkin’s lymphoma. Cancer Res. 1988;48(10):2919–2922. | ||
Lee J, Kim Y, Kang CS, et al. Investigation of the bovine leukemia virus proviral DNA in human leukemias and lung cancers in Korea. J Korean Med Sci. 2005;20(4):603–606. | ||
Buehring GC, Philpott SM, Choi KY. Humans have antibodies reactive with Bovine leukemia virus. AIDS Res Hum Retroviruses. 2003;19(12):1105–1113. | ||
Onuma M, Tsukiyama K, Ohya KI, Morishima Y, Ohno R. Detection of cross-reactive antibody to BLV p24 in sera of human patients infected with HTLV. Microbiol Immunol. 1987;31(2):131–137. | ||
Maruyama K, Fukushima T, Mochizuki S. Cross-reactive antibodies to BLV and HTLV in bovine and human hosts with retrovirus infection. Vet Immunol Immunopathol. 1989;22(3):265–273. | ||
Zandomeni RO, Carrera-Zandomeni M, Esteban E, Ferrer JF. The trans-activating C-type retroviruses share a distinct epitope(s) that induces antibodies in certain infected hosts. J Gen Virol. 1991;72(Pt 9):2113–2119. | ||
Buehring GC, Shen HM, Jensen HM, Choi KY, Sun D, Nuovo G. Bovine leukemia virus DNA in human breast tissue. Emerg Infect Dis. 2014;20(5):772–782. | ||
Buehring GC, Shen HM, Jensen HM, Jin DL, Hudes M, Block G. Exposure to bovine leukemia virus is associated with breast cancer: a case-control study. PLoS One. 2015;10(9):e0134304. | ||
Zhang R, Jiang J, Sun W, et al. Lack of association between bovine leukemia virus and breast cancer in Chinese patients. Breast Cancer Res. 2016;18(1):101. | ||
Gillet NA, Willems L. Whole genome sequencing of 51 breast cancers reveals that tumors are devoid of bovine leukemia virus DNA. Retrovirology. 2016;13:75. | ||
Tang KW, Alaei-Mahabadi B, Samuelsson T, Lindh M, Larsson E. The landscape of viral expression and host gene fusion and adaptation in human cancer. Nat Commun. 2013;4:2513. | ||
Khoury JD, Tannir NM, Williams MD, et al. Landscape of DNA virus associations across human malignant cancers: analysis of 3,775 cases using RNA-Seq. J Virol. 2013;87(16):8916–8926. | ||
Hopkins SG, DiGiacomo RF. Natural transmission of bovine leukemia virus in dairy and beef cattle. Vet Clin North Am Food Anim Pract. 1997;13(1):107–128. | ||
Lassauzet ML, Thurmond MC, Johnson WO, Holmberg CA. Factors associated with in utero or periparturient transmission of bovine leukemia virus in calves on a California dairy. Can J Vet Res. 1991;55(3):264–268. | ||
Romero CH, Cruz GB, Rowe CA. Transmission of bovine leukaemia virus in milk. Trop Anim Health Prod. 1983;15(4):215–218. | ||
Juliarena MA, Barrios CN, Ceriani MC, Esteban EN. Hot topic: bovine leukemia virus (BLV)-infected cows with low proviral load are not a source of infection for BLV-free cattle. J Dairy Sci. 2016;99(6):4586–4589. | ||
Ooshiro M, Konnai S, Katagiri Y, et al. Horizontal transmission of bovine leukemia virus from lymphocytotic cattle, and beneficial effects of insect vector control. Vet Rec. 2013;173(21):527. | ||
Anderson RK, Sorensen DK, Perman V, Dirks VA, Snyder MM, Bearman JE. Selected epizootiologic aspects of bovine leukemia in Minnesota (1961–1965). Am J Vet Res. 1971;32:563–577. | ||
Rhodes JK, Pelzer KD, Johnson YJ. Economic implications of bovine leukemia virus infection in mid-Atlantic dairy herds. J Am Vet Med Assoc. 2003;223(3):346–352. | ||
Wu MC, Shanks RD, Lewin HA. Milk and fat production in dairy cattle influenced by advanced subclinical bovine leukemia virus infection. Proc Natl Acad Sci U S A. 1989;86(3):993–996. | ||
Pollari FL, Wangsuphachart VL, DiGiacomo RF, Evermann JF. Effects of bovine leukemia virus infection on production and reproduction in dairy cattle. Can J Vet Res. 1992;56(4):289–295. | ||
Callebaut I, Voneche V, Mager A, et al. Mapping of B-neutralizing and T-helper cell epitopes on the bovine leukemia virus external glycoprotein gp51. J Virol. 1993;67(9):5321–5327. | ||
Jacobs RM, Heeney JL, Godkin MA, et al. Production and related variables in bovine leukaemia virus-infected cows. Vet Res Commun. 1991;15(6):463–474. | ||
Kale M, Bulut O, Yapk O, et al. Effects of subclinical bovine leukemia virus infection on some production parameters in a dairy farm in southern Turkey. J S Afr Vet Assoc. 2007;78(3):130–132. | ||
Tiwari A, VanLeeuwen JA, Dohoo IR, et al. Production effects of pathogens causing bovine leukosis, bovine viral diarrhea, paratuberculosis, and neosporosis. J Dairy Sci. 2007;90(2):659–669. | ||
Sorge US, Lissemore K, Cantin R, Kelton DF. Short communication: Milk ELISA status for bovine leukosis virus infection is not associated with milk production in dairy cows. J Dairy Sci. 2011;94(10):5062–5064. | ||
Erskine RJ, Bartlett PC, Byrem TM, Render CL, Febvay C, Houseman JT. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J Dairy Sci. 2012;95(2):727–734. | ||
Ott SL, Johnson R, Wells SJ. Association between bovine-leukosis virus seroprevalence and herd-level productivity on US dairy farms. Prev Vet Med. 2003;61(4):249–262. | ||
Norby B, Bartlett PC, Byrem TM, Erskine RJ. Effect of infection with bovine leukemia virus on milk production in Michigan dairy cows. J Dairy Sci. 2016;99(3):2043–2052. | ||
Yang Y, Fan W, Mao Y, et al. Bovine leukemia virus infection in cattle of China: association with reduced milk production and increased somatic cell score. J Dairy Sci. 2016;99(5):3688–3697. | ||
Nekouei O, VanLeeuwen J, Stryhn H, Kelton D, Keefe G. Lifetime effects of infection with bovine leukemia virus on longevity and milk production of dairy cows. Prev Vet Med. 2016;133:1–9. | ||
Da Y, Shanks RD, Stewart JA, Lewin HA. Milk and fat yields decline in bovine leukemia virus-infected Holstein cattle with persistent lymphocytosis. Proc Natl Acad Sci U S A. 1993;90(14):6538–6541. | ||
Grunstein M. Histone acetylation in chromatin structure and transcription. Nature. 1997;389(6649):349–352. | ||
Rotili D, Simonetti G, Savarino A, Palamara AT, Migliaccio AR, Mai A. Non-cancer uses of histone deacetylase inhibitors: effects on infectious diseases and beta-hemoglobinopathies. Curr Top Med Chem. 2009;9(3):272–291. | ||
Beumer JH, Tawbi H. Role of histone deacetylases and their inhibitors in cancer biology and treatment. Curr Clin Pharmacol. 2010;5(3):196–208. | ||
De Souza C, Chatterji BP. HDAC inhibitors as novel anti-cancer therapeutics. Recent Pat Anticancer Drug Discov. 2015;10(2):145–162. | ||
Moog C, Kuntz-Simon G, Caussin-Schwemling C, Obert G. Sodium valproate, an anticonvulsant drug, stimulates human immunodeficiency virus type 1 replication independently of glutathione levels. J Gen Virol. 1996;77 (Pt 9):1993–1999. | ||
Chen WY, Townes TM. Molecular mechanism for silencing virally transduced genes involves histone deacetylation and chromatin condensation. Proc Natl Acad Sci U S A. 2000;97(1):377–382. | ||
Lehrman G, Hogue IB, Palmer S, et al. Depletion of latent HIV-1 infection in vivo: a proof-of-concept study. Lancet. 2005;366(9485):549–555. | ||
Lezin A, Gillet N, Olindo S, et al. Histone deacetylase mediated transcriptional activation reduces proviral loads in HTLV-1 associated myelopathy/tropical spastic paraparesis patients. Blood. 2007;110(10):3722–3728. | ||
Afonso PV, Mekaouche M, Mortreux F, et al. Highly active antiretroviral treatment against STLV-1 infection combining reverse transcriptase and HDAC inhibitors. Blood. 2010;116(19):3802–3808. | ||
Phiel CJ, Zhang F, Huang EY, Guenther MG, Lazar MA, Klein PS. Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J Biol Chem. 2001;276(39):36734–36741. | ||
Achachi A, Florins A, Gillet N, et al. Valproate activates bovine leukemia virus gene expression, triggers apoptosis, and induces leukemia/lymphoma regression in vivo. Proc Natl Acad Sci U S A. 2005;102(29):10309–10314. | ||
Gillet N, Vandermeers F, de Brogniez A, et al. Chemoresistance to valproate treatment of bovine leukemia virus-infected sheep; Identification of improved HDAC inhibitors. Pathogens. 2012;1(2):65–82. | ||
Olindo S, Belrose G, Gillet N, et al. Safety of long-term treatment of HAM/TSP patients with valproic acid. Blood. 2011;118(24):6306–6309. | ||
Sagot-Lerolle N, Lamine A, Chaix ML, et al. Prolonged valproic acid treatment does not reduce the size of latent HIV reservoir. AIDS. 2008;22(10):1125–1129. | ||
Archin NM, Cheema M, Parker D, et al. Antiretroviral intensification and valproic acid lack sustained effect on residual HIV-1 viremia or resting CD4+ cell infection. PLOS ONE. 2010;5(2):e9390. | ||
Routy JP, Tremblay CL, Angel JB, et al. Valproic acid in association with highly active antiretroviral therapy for reducing systemic HIV-1 reservoirs: results from a multicentre randomized clinical study. HIV Med. 2012;13(5):291–296. | ||
Boostani R, Vakili R, Hosseiny SS, et al. Triple therapy with prednisolone, pegylated interferon and sodium valproate improves clinical outcome and reduces human T-cell leukemia virus type 1 (HTLV-1) proviral load, tax and HBZ mRNA expression in patients with HTLV-1-associated myelopathy/tropical spastic paraparesis. Neurotherapeutics. 2015;12(4):887–895. | ||
Darcis G, Kula A, Bouchat S, et al. An in-depth comparison of latency-reversing agent combinations in various in vitro and ex vivo HIV-1 latency models identified bryostatin-1+JQ1 and ingenol-B+JQ1 to potently reactivate viral gene expression. PLoS Pathog. 2015;11(7): e1005063. | ||
Callebaut I, Mornon JP, Burny A, Portetelle D. The bovine leukemia virus (BLV) envelope glycoprotein gp51 as a general model for the design of a subunit vaccine against retroviral infection: mapping of functional sites through immunological and structural data. Leukemia. 1994;8(Suppl 1):S218–S221. | ||
Altaner C, Altanerova V, Ban J, et al. Cell-derived vaccine against bovine leukaemia virus infection. Zentralbl Veterinarmed B. 1988;35(10):736–746. | ||
Altaner C, Ban J, Altanerova V, Janik V. Protective vaccination against bovine leukaemia virus infection by means of cell-derived vaccine. Vaccine. 1991;9(12):889–895. | ||
Altaner C, Ban J, Altanerova V, Cerny L. Experimental transmission of the bovine leukaemia virus genome by cell contact. Folia Biol (Praha). 1987;33(6):400–409. | ||
Miller JM, Van Der Maaten MJ. Evaluation of an inactivated bovine leukemia virus preparation as an immunogen in cattle. Ann Rech Vet. 1978;9(4):871–877. | ||
Fukuyama S, Kodama K, Hirahara T, et al. Protection against bovine leukemia virus infection by use of inactivated vaccines in cattle. J Vet Med Sci. 1993;55(1):99–106. | ||
Burkhardt H, Rosenthal S, Wittmann W, et al. [Immunization of young cattle with gp51 of the bovine leukosis virus and the subsequent experimental infection]. Arch Exp Veterinarmed. 1989;43(6):933–942. | ||
Cherney TM, Schultz RD. Viral status and antibody response in cattle inoculated with recombinant bovine leukemia virus-vaccinia virus vaccines after challenge exposure with bovine leukemia virus-infected lymphocytes. Am J Vet Res. 1996;57(6):812–818. | ||
Ohishi K, Suzuki H, Yamamoto T, et al. Protective immunity against bovine leukaemia virus (BLV) induced in carrier sheep by inoculation with a vaccinia virus-BLV ENV recombinant: association with cell-mediated immunity. J Gen Virol. 1991;72(8):1887–1892. | ||
Ohishi K, Suzuki H, Yasutomi Y, et al. Augmentation of bovine leukemia virus (BLV)-specific lymphocyte proliferation responses in ruminants by inoculation with BLV env-recombinant vaccinia virus: their role in the suppression of BLV replication. Microbiol Immunol. 1992;36(12):13171323. | ||
Ohishi K, Ikawa Y. T cell-mediated destruction of bovine leukemia virus-infected peripheral lymphocytes by bovine leukemia virus env-vaccinia recombinant vaccine. AIDS Res Hum Retroviruses. 1996;12(5):393–398. | ||
Gatei MH, Naif HM, Kumar S, et al. Protection of sheep against bovine leukemia virus (BLV) infection by vaccination with recombinant vaccinia viruses expressing BLV envelope glycoproteins: correlation of protection with CD4 T-cell response to gp51 peptide 51–70. J Virol. 1993;67(4):1803–1810. | ||
Okada K, Ikeyama S, Ohishi K, et al. Involvement of CD8+ T cells in delayed-type hypersensitivity responses against bovine leukemia virus (BLV) induced in sheep vaccinated with recombinant vaccinia virus expressing BLV envelope glycoprotein. Vet Pathol. 1993;30(2):104–110. | ||
Ohishi K, Kabeya H, Amanuma H, Onuma M. Peptide-based bovine leukemia virus (BLV) vaccine that induces BLV-Env specific Th-1 type immunity. Leukemia. 1997;11(Suppl 3):223–226. | ||
Hislop AD, Good MF, Mateo L, et al. Vaccine-induced cytotoxic T lymphocytes protect against retroviral challenge. Nat Med. 1998;4(10):1193–1196. | ||
Portetelle D, Limbach K, Burny A, et al. Recombinant vaccinia virus expression of the bovine leukaemia virus envelope gene and protection of immunized sheep against infection. Vaccine. 1991;9(3):194–200. | ||
Pyeon D, Splitter GA. Interleukin-12 p40 mRNA expression in bovine leukemia virus-infected animals: increase in alymphocytosis but decrease in persistent lymphocytosis. J Virol. 1998;72(8):6917–6921. | ||
Yakobson B, Brenner J, Ungar-Waron H, Trainin Z. Cellular immune response cytokine expression during the initial stage of bovine leukemia virus (BLV) infection determines the disease progression to persistent lymphocytosis. Comp Immunol Microbiol Infect Dis. 2000;23(3):197–208. | ||
Usui T, Konnai S, Ohashi K, Onuma M. Interferon-gamma expression associated with suppression of bovine leukemia virus at the early phase of infection in sheep. Vet Immunol Immunopathol. 2007;115(1–2):17–23. | ||
Mateo L, Gardner J, Suhrbier A. Delayed emergence of bovine leukemia virus after vaccination with a protective cytotoxic T cell-based vaccine. AIDS Res Hum Retroviruses. 2001;17(15):1447–1453. | ||
Brillowska A, Dabrowski S, Rulka J, Kubis P, Buzala E, Kur J. Protection of cattle against bovine leukemia virus (BLV) infection could be attained by DNA vaccination. Acta Biochim Pol. 1999;46(4):971–976. | ||
Usui T, Konnai S, Tajima S, et al. Protective effects of vaccination with bovine leukemia virus (BLV) Tax DNA against BLV infection in sheep. J Vet Med Sci. 2003;65(11):1201–1205. | ||
Derse D, Martarano L. Construction of a recombinant bovine leukemia virus vector for analysis of virus infectivity. J Virol. 1990;64(1):401–405. | ||
Milan D, Nicolas JF. Activator-dependent and activator-independent defective recombinant retroviruses from bovine leukemia virus. J Virol. 1991;65(4):1938–1945. | ||
Boris-Lawrie K, Altanerova V, Altaner C, Kucerova L, Temin HM. In vivo study of genetically simplified bovine leukemia virus derivatives that lack tax and rex. J Virol. 1997;71(2):1514–1520. | ||
Kerkhofs P, Gatot JS, Knapen K, et al. Long-term protection against bovine leukaemia virus replication in cattle and sheep. J Gen Virol. 2000;81(Pt 4):957–963. | ||
Willems L, Chen G, Portetelle D, Mamoun R, Burny A, Kettmann R. Structural and functional characterization of mutants of the bovine leukemia virus transactivator protein p34. Virology. 1989;171(2):615–618. | ||
Willems L, Kettmann R, Dequiedt F, et al. In vivo infection of sheep by bovine leukemia virus mutants. J Virol. 1993;67(7):4078–4085. | ||
Boris-Lawrie K, Temin HM. Genetically simpler bovine leukemia virus derivatives can replicate independently of Tax and Rex. J Virol. 1995;69(3):1920–1924. | ||
Kucerova L, Altanerova V, Altaner C, Boris-Lawrie K. Bovine leukemia virus structural gene vectors are immunogenic and lack pathogenicity in a rabbit model. J Virol. 1999;73(10):8160–8166. | ||
Altanerova V, Holicova D, Kucerova L, Altaner C, Lairmore MD, Boris-Lawrie K. Long-term infection with retroviral structural gene vector provides protection against bovine leukemia virus disease in rabbits. Virology. 2004;329(2):434–439. | ||
Gatot JS, Callebaut I, Mornon JP, et al. Conservative mutations in the immunosuppressive region of the bovine leukemia virus transmembrane protein affect fusion but not infectivity in vivo. J Biol Chem. 1998;273(21):12870–12880. | ||
Rodriguez SM, Florins A, Gillet N, et al. Preventive and therapeutic strategies for bovine leukemia virus: lessons for HTLV. Viruses. 2011;3(7):1210–1248. | ||
Dequiedt F, Hanon E, Kerkhofs P, et al. Both wild-type and strongly attenuated bovine leukemia viruses protect peripheral blood mononuclear cells from apoptosis. J Virol. 1997;71(1):630–639. | ||
Reichert M, Cantor GH, Willems L, Kettmann R. Protective effects of a live attenuated bovine leukaemia virus vaccine with deletion in the R3 and G4 genes. J Gen Virol. 2000;81(Pt 4):965–969. | ||
Asfaw Y, Tsuduku S, Konishi M, et al. Distribution and superinfection of bovine leukemia virus genotypes in Japan. Arch Virol. 2005;150(3):493–505. | ||
Gutierrez G, Rodriguez SM, de Brogniez A, et al. Vaccination against delta-retroviruses: the bovine leukemia virus paradigm. Viruses. 2014;6(6):2416–2427. | ||
Dong B, Silverman RH, Kandel ES. A natural human retrovirus efficiently complements vectors based on murine leukemia virus. PLoS One. 2008;3(9):e3144. | ||
Lee SW, Markham PF, Coppo MJ, et al. Attenuated vaccines can recombine to form virulent field viruses. Science. 2012;337(6091):188. | ||
Watanabe T, Inoue E, Mori H, Osawa Y, Okazaki K. Delayed-onset enzootic bovine leukosis possibly caused by superinfection with bovine leukemia virus mutated in the pol gene. Arch Virol. 2015;160(8):2087–2091. | ||
Iwabu Y, Goto T, Tsuji S, et al. Superinfection of human immunodeficiency virus type 1 (HIV-1) to cell clone persistently infected with defective virus induces production of highly cytopathogenic HIV-1. Microbes Infect. 2006;8(7):1773–1782. | ||
Iwabu Y, Mizuta H, Kawase M, Kameoka M, Goto T, Ikuta K. Superinfection of defective human immunodeficiency virus type 1 with different subtypes of wild-type virus efficiently produces infectious variants with the initial viral phenotypes by complementation followed by recombination. Microbes Infect. 2008;10(5):504–513. | ||
Ferrer JF. Bovine leukosis: natural transmission and principles of control. J Am Vet Med Assoc. 1979;175(12):1281–1286. | ||
DiGiacomo RF. The epidemiology and control of bovine leukemia virus infection. Veterinary Medicine. 1992;87:248–257. | ||
Bartlett PC, Sordillo LM, Byrem TM, et al. Options for the control of bovine leukemia virus in dairy cattle. J Am Vet Med Assoc. 2014;244(8):914–922. | ||
Eaglesome MD, Mitchell D, Betteridge KJ, et al. Transfer of embryos from bovine leukaemia virus-infected cattle to uninfected recipients: preliminary results. Vet Rec. 1982;111(6):122–123. | ||
DiGiacomo RF, Studer E, Evermann JF, Evered J. Embryo transfer and transmission of bovine leukosis virus in a dairy herd. J Am Vet Med Assoc. 1986;188(8):827–828. | ||
DiGiacomo RF, McGinnis LK, Studer E, Evermann JF. Failure of embryo transfer to transmit BLV in a dairy herd. Vet Rec. 1990;127(18):456. | ||
Suh GH, Lee JC, Lee CY, et al. Establishment of a bovine leukemia virus-free dairy herd in Korea. J Vet Sci. 2005;6(3):227–230. | ||
Shettigara PT, Samagh BS, Lobinowich EM. Control of bovine leukemia virus infection in dairy herds by agar gel immunodiffusion test and segregation of reactors. Can J Vet Res. 1989;53(1):108–110. | ||
Lassauzet ML, Johnson WO, Thurmond MC, Stevens F. Protection of colostral antibodies against bovine leukemia virus infection in calves on a California dairy. Can J Vet Res. 1989;53(4):424–430. | ||
Nagy DW, Tyler JW, Kleiboeker SB. Decreased periparturient transmission of bovine leukosis virus in colostrum-fed calves. J Vet Intern Med. 2007;21(5):1104–1107. | ||
Van Der Maaten MJ, Miller JM, Schmerr MJ. Effect of colostral antibody on bovine leukemia virus infection of neonatal calves. Am J Vet Res. 1981;42(9):1498–1500. | ||
Sprecher DJ, Pelzer KD, Lessard P. Possible effect of altered management practices on seroprevalence of bovine leukemia virus in heifers of a dairy herd with history of high prevalence of infection. J Am Vet Med Assoc. 1991;199(5):584–588. | ||
Gutiérrez SE, Esteban EN, Lützelschwab CM, Juliarena MA. Major histocompatibility complex-associated resistance to infectious diseases: the case of bovine leukemia virus infection. In: Abubakar M, editor. Trends and Advances in Veterinary Genetics. Croatia: InTech; 2017. | ||
Lewin HA. Disease resistance and immune response genes in cattle: strategies for their detection and evidence of their existence. J Dairy Sci. 1989;72(5):1334–1348. | ||
Xu A, van Eijk MJ, Park C, Lewin HA. Polymorphism in BoLA-DRB3 exon 2 correlates with resistance to persistent lymphocytosis caused by bovine leukemia virus. J Immunol. 1993;151(12):6977–6985. | ||
Zanotti M, Poli G, Ponti W, et al. Association of BoLA class II haplotypes with subclinical progression of bovine leukaemia virus infection in Holstein-Friesian cattle. Anim Genet. 1996;27(5):337–341. | ||
IPD. Immuno Polymorphism Database - MHC Database. BoLA Nomenclature, Cattle, BoLA DRB3. Available from https://www.ebi.ac.uk/cgi-bin/ipd/mhc/view_nomenclature.cgi?bola.drb3. Accessed July 05, 2016. | ||
Mirsky ML, Olmstead C, Da Y, Lewin HA. Reduced bovine leukaemia virus proviral load in genetically resistant cattle. Anim Genet. 1998;29(4):245–252. | ||
Juliarena MA, Poli M, Sala L, et al. Association of BLV infection profiles with alleles of the BoLA-DRB3.2 gene. Anim Genet. 2008;39(4):432–438. | ||
Miyasaka T, Takeshima SN, Jimba M, et al. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese Black cattle. Tissue Antigens. 2012;81(2):72–82. | ||
Esteban EN, Poli M, Poiesz B, et al. Bovine leukemia virus (BLV), proposed control and eradication programs by marker assisted breeding of genetically resistant cattle. In: Rechi LJ, editor. Animal Genetics. Hauppauge, NY: Nova Science Publishers, Inc.; 2009:107–130. | ||
Juliarena MA, Poli M, Ceriani C, et al. Antibody response against three widespread bovine viruses is not impaired in Holstein cattle carrying bovine leukocyte antigen DRB3.2 alleles associated with bovine leukemia virus resistance. J Dairy Sci. 2009;92(1):375–381. | ||
Juliarena M. Aportes al estudio del rol del virus y del hospedador en el perfil de infección causado por el virus de la leucosis bovina. Tesis Doctoral. FCV-UNCPBA. 2008. | ||
Miyasaka T, Takeshima SN, Sentsui H, Aida Y. Identification and diversity of bovine major histocompatibility complex class II haplotypes in Japanese Black and Holstein cattle in Japan. J Dairy Sci. 2012;95(1):420–431. | ||
Dietz AB, Cohen ND, Timms L, Kehrli ME, Jr. Bovine lymphocyte antigen class II alleles as risk factors for high somatic cell counts in milk of lactating dairy cows. J Dairy Sci. 1997;80(2):406–412. | ||
Rupp R, Hernandez A, Mallard BA. Association of bovine leukocyte antigen (BoLA) DRB3.2 with immune response, mastitis, and production and type traits in Canadian Holsteins. J Dairy Sci. 2007;90(2):1029–1038. | ||
Sharif S, Mallard BA, Wilkie BN, et al. Associations of the bovine major histocompatibility complex DRB3 (BoLA-DRB3) alleles with occurrence of disease and milk somatic cell score in Canadian dairy cattle. Anim Genet. 1998;29(3):185-193. | ||
Nassiry MR, Shahroodi FE, Mosafer J, et al. Analysis and frequency of bovine lymphocyte antigen (BoLA-DRB3) alleles in Iranian Holstein cattle. Genetika. 2005;41(6):817–822. | ||
Lützelschwab CM, Forletti A, Cepeda R, Esteban EN, Confalonieri O, Gutiérrez SE. Co-infection with Mycobacterium bovis does not alter the response to bovine leukemia virus in BoLA DRB3*0902, genetically resistant cattle. Res Vet Sci. 2016;109:10–16. | ||
Forletti A, Juliarena MA, Ceriani C, Amadio AF, Esteban E, Gutierrez SE. Identification of cattle carrying alleles associated with resistance and susceptibility to the bovine leukemia virus progression by real-time PCR. Res Vet Sci. 2013;95(3):991–995. |
 © 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2017 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.