Back to Archived Journals » International Journal of Wine Research » Volume 8
Botrytized wines – current perspectives
Received 27 July 2016
Accepted for publication 16 August 2016
Published 19 October 2016 Volume 2016:8 Pages 29—39
DOI https://doi.org/10.2147/IJWR.S100653
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Ildikó Magyar, János Soós
Department of Oenology, Institute of Viticulture and Oenology, Szent István University, Budapest, Hungary
Abstract: Botrytized wines are wine specialties made of overripe grapes infected by Botrytis cinerea with the form “noble rot”. Due to the particular characteristics of the noble rotted grape, these wines (eg, Tokaji Aszú, Sauternes, Trockenbeerenauslese types, etc) have many characteristic features, including higher or lower residual sugar content and unique aroma composition. The technology, biochemistry, and special characteristics of botrytized wines have been researched for a long time. This review outlines the main directions of the current studies, giving a brief overview on the recent findings. Beside the traditional wine types, noble rot is increasingly utilized in making newer sweet wine styles and straw (passito) wines, which generates a series of new interesting experimental results. The fungus–grape interactions during the noble rot, the induced botrytization, the microbial communities of botrytized wines, and the volatile compounds having key roles in the distinct aroma of these wine styles are being focused on in the current studies in this field.
Keywords: Botrytis, noble rot, sweet wines, passito wine, aroma
Introduction
Botrytized wines are a distinct category of the natural sweet wines produced from withered, shriveled grapes. While the other sweet styles such as late harvest wines, ice wines, and traditional straw (“passito”) wines are mostly made of physically dehydrated, concentrated grape juices, botrytized wines are produced from noble rotted grape, which is not only dehydrated but also infected and transformed by Botrytis cinerea under special conditions. In contrast to the usual detrimental activity of this fungus on grapes (gray rot), noble rot is a rare, desirable process, which greatly enhances grape quality. During noble rot, some of the compounds of the grape are decomposed, while new compounds are produced by the fungus. This modified chemical composition of the grape juice is highly concentrated due to the physical effect of dehydration, which is supported by the digested grape skin and by particular weather conditions (morning dust/fog and dry, sunny days). The wines produced from this extremely high sugar and extract content grape have many characteristic features, including higher or lower residual sugar content and unique aroma composition. Some distinctive descriptive terms associated with botrytized wines are peach, apricot, quince, tropical fruit, honey, and caramel. The aging process may add more nuances, eg, chocolate, tobacco, dried fruits, raisins, etc.
Listed in historical order, the oldest type of these wines is Tokaji Aszú in Hungary,1,2 followed by Trockenbeerenauslese (TBA) and Beerenauslese wines in Germany and Austria.3,4 Other famous names are Sauternes4 and “Sélection de Grains Nobles” wines in France. There is an increasing interest in producing botrytized sweet wines in Australia and South Africa, while they are also produced occasionally in California.1 Recently, the benefits of botrytization have become well recognized and utilized in making various passito wines in Italy (Picolit and Recioto), even in dry styles (Amarone).
Since the 1950s, extended research has been carried out on botrytized wine specialties. The technology, biochemistry, and special characteristics of botrytized sweet wines have recently been reviewed by several authors.1,3,4 While briefly summarizing the most important and previously confirmed knowledge in this field, the aim of this review is to give an overview of the most recent research on botrytized wines. In accordance with the topics of current publications, the focus of this review is on the interaction of the grape and B. cinerea during noble rot, the microbiological diversity of botrytized wines, and the sensory and chemical aspects of their aroma composition.
The botrytization process
B. cinerea (Figure 1) generally is very detrimental to wine quality when it causes bunch rot, also known as gray rot/mold, causing significant economic loss. Under particular conditions, B. cinerea generates a special form of infection called noble rot, which highly improves the quality of the grape.
 | Figure 1 Scanning electron micrographs of Botrytis cinerea conidiophore developing on the surface of noble rotted berry. |
The main factors that lead to noble rot are 1) appropriate climatic and microclimatic conditions, 2) appropriate grape variety (ripening time, anatomy of berry and bunch, phytoalexin production, etc), and 3) full or over-ripeness of the grape at the time of Botrytis invasion (when the vascular connection between the vine and berry ceases). There are only a few geographical–viticultural environments where all these conditions coincide relatively frequently.
The noble rot can be summarized as an interaction and balance between the enzymatic activity of B. cinerea and the concentrating effect of physical dehydration on the grape berry. Several authors have described the infection process, which has recently been reviewed,1,5 and hence only a brief summary is given in this review. Since Botrytis has limited ability to digest the intact cuticle of the berry skin, penetration occurs through stomata, microfissures, or wounds. The growth of the fungus is restricted to the outer layers of the epidermis, digesting the skin, extracting chemical compounds from the berry, and introducing metabolic products into the juice. The permeable skin supports the evaporation of water during sunny and windy days, which results in shriveling of the berry and several-fold concentration of the juice. The sugar content of the botrytized grape commonly reaches 350 g/L in Sauternes6 or even ~700 g/L,1,2 if the shriveling process is intensive, as in Tokaj (Figure 2).
 | Figure 2 Selective picking of botrytized and shriveled (“aszú”) berries in Tokaj. Note: Photo courtesy of Dr Zsuzsanna Bene. |
Botrytis produces numerous hydrolase and oxidase enzymes, which transform many components of the grape tissue and the juice. Oxidation of glucose leads to production of glycerol, >7 g/L,6 but it may exceed 30 g/L after further berry dehydration, eg, in Tokaji Aszú berries and TBA grapes.1,3 Other sugar alcohols (arabitol, mannitol, sorbitol, and inositol) and various carbonyl compounds are also produced. Part of the glucose is oxidized to gluconic acid, considered as another Botrytis indicator; however, its elevated level (up to 2–3 g/L) in botrytized berry can be partially explained by the activity of acetic acid bacteria. Taking part in this reaction is a glucose oxidase (GOX), not present in healthy grapes, but typical of botrytized grapes.7 Via oxidation of glucose to gluconic acid, it releases H2O2, which can generate further oxidation reactions, such as oxidation of tartaric acid to glyoxylic acid, ethanol to acetaldehyde, and glycerol to glycerol aldehyde. These compounds will further react with catechins and proanthocyanidins, influencing the color and thus the visual quality of the wine. GOX remains stable in wine conditions as it is relatively resistant to sulfites, and ascorbic acid and bentonite have little effect on its activity.7
One of the most studied enzyme families of Botrytis is an aggressive polyphenol oxidase (PPO) enzyme group called laccase.6–8 While GOX has an indirect effect on polyphenols, the direct role of PPO in the oxidation of a wide scale of phenols has been known for a long time.8 Although PPO is more stable in wine than the polyphenol oxidase enzymes of the grape, it is more sensitive to SO2 than GOX.7
Botrytis significantly influences the composition of nitrogenous substances in grapes. While the yeast-assimilable nitrogen content decreases and the amino acid pattern changes,3 production of exocellular fungal enzymes increases the protein content of the juice.6,9 A further specialty is a particular polysaccharide composition.
More recent studies focus not only on the enzymatic activities of Botrytis in the grape but also on the metabolic response of the plant to Botrytis attack.9,10 Changes in the polyphenol composition of the berry result from a combination of the fungal PPO and GOX activity and the metabolic response of the grape to fungal infection. The total phenol content either decreases11,12 or possibly increases in highly dehydrated berries due to concentration. Flavonoids are the most important group of phenolics in wine. A significant accumulation of several flavonoid glycosides and flavanones was detected in noble rotted berries of Sémillon grape but the production of flavones and flavonols was suppressed during noble rot.10 In contrast, the presence of the fungus on Chenin blanc grape seems to induce, from one side, an increase in flavonols, such as catechin, epicatechin, and epicatechin gallate, and from the other side, a decrease in caftaric acid, coutaric acid, quercetin-3 glucoside, and kaempferol-3 glucoside.12 From the results, it seems that changes in the flavonoid composition show varietal influences and need further research.
The plant responds to the fungal infection by producing more stilbenes such as resveratrol and its derivates, which act as phytoalexins against the fungus. The evolution of these compounds depends on the grape variety and on the development of noble rot.10 In the last stage of noble rot, viniferin and astilbin could be used by the plant against Botrytis. However, the final level of stilbene derivates (trans-astringin, trans-resveratrol, trans-piceid, etc) is generally low in the botrytized juice and wine, probably because Botrytis enzymes of the laccase group, such as stilbene oxidase, are able to transform these molecules to oxidized compounds, which was more extensively reviewed by Teissedre and Donèche.4
In a significant, comprehensive study, Blanco-Ulate et al10 have found desirable interaction between a fruit and a fungus that stimulates pathways otherwise inactive in white-skinned berries. Using transcript and metabolite analysis of the grape tissue, they identified several genes that were upregulated due to Botrytis infection. They showed that noble rot stimulated the synthesis of anthocyanins in white-skinned grape (Sémillon), contributing to the pink and bluish color of the affected berries. Biosynthesis of stilbenes, terpineols, and fatty acid aroma precursors was also stimulated by Botrytis.
Modification of grape metabolism by Botrytis concerns the aroma potential of the grape as well, which was extensively studied by Thibon et al.13 Certain grape varieties (Sauvignon blanc, Sémillon) contain cysteine-S conjugate molecules, which are precursors of several important thiols released by yeasts during fermentation.13 These cysteinylated precursor compounds are S-3-(hexan-1-ol)-l-cysteine (P-3SH), S-3-(pentan-1-ol)-l-cysteine (P-3SP), S-3-(heptan-1-ol)-l-cysteine (P-3SHp), and S-3-(2-methylbutan-1-ol)-l-cysteine (P-2M3SB) and are converted by yeasts to strongly odiferous thiols in the wine (refer the “Aroma composition of botrytized wines” section).13,14 These precursors are present in the healthy grape as well, but their quantities dramatically increase due to noble rot. The botrytization process increased the P-3SH level in grapes ~100-fold in 1 week, compared to healthy grapes.15 The production pathway of this molecule was elucidated by Thibon et al.16 In model experiments using cell cultures, it was demonstrated that grapevine cells were able to produce P-3SH and that the presence of B. cinerea considerably increased the precursor level (up to 1,000-fold).
Since the noble rotted grape is superior to the withered grape in terms of wine quality, discriminating them objectively would be valuable from a practical standpoint. The main aim of several studies is to identify possible markers of Botrytis infection in dehydrated grapes.
PPO activity is generally accepted as an index of noble rot infection,6 although some controversial results have been presented.17 Using monoclonal antibodies, highly sensitive and specific immunoassays have also been elaborated, which are rapid and user friendly, supporting practical application in the wineries.18 More recently, protein determination by mass spectrometry analysis allowed researchers to identify specific proteins of B. cinerea that represent potential markers of the presence of the fungus in the withered grapes.19
Additionally, physical properties might be appropriate for differentiating botrytized and withered grapes. Instrumental measurements of chromatic parameters (CIEL*a*b*) on the berries of white grapes proved that only blue dehydrated grapes showed a chemical and volatile composition typical of the noble rot infection. It is of great relevance for wineries where passito wine production is carried out on a small scale, and manual selection of green, orange, and blue berries from the stalk enables winemakers to differentiate the wine characteristics.20 Measuring various physical parameters (eg, mechanical resistance of the skin, color, etc) the combination of puncture and compression tests with color (CIEL*a*b* parameters) showed a clear differentiation of botrytized berries in Chenin blanc grape (Loire Valley) and appeared to be the most adequate to establish markers of noble rot evolution.12 Phenolic composition was another discriminating marker in this study. Recently, the applicability of electronic tongue is under study for differentiation of botrytized and healthy juice or wine.21
Induced botrytization
The rare occurrence and uncertainty of noble rot have for a long time motivated researchers to obtain induced and/or controlled botrytization. Moreover, global climatic change will severely impact the delicate equilibrium needed for natural noble rot, a point which has to be considered in future research.5
Several promising experiments were reported on artificial postharvest infection of grape or juice. These methods had not been in practice, probably due to the high costs at that time.5 However, vineyard infection has been applied with success in the Napa valley, California, where a natural Botrytis isolate from Sémillon grape was used as a source of inoculum to induce noble rot in the production of Dolce Wine (Oakville, CA, USA). This strain was successfully used for artificial infection in the vineyard in several subsequent years, and the noble rotted grape has been used for commercial wine production as well as for several interesting research studies on botrytization.10,22–24 The draft genome sequence of this strain (BcDW1) has recently been published.10
Induction of Botrytis infection during postharvest withering of the grape has been increasing as a strategy in making straw (passito) wines (eg, Amarone and Recioto) in Italy. Although these wines are traditionally made of fully ripened but nonbotrytized grapes that are dehydrated in drying chambers after harvest, Botrytis infection occasionally may occur either in the vineyard or during withering. The incidence of spontaneous Botrytis infection is extremely uncertain, depending on seasonal conditions and withering techniques.10,11,25,26 Both research and practical investigations on different passito wine types have proved that Botrytis infection of the withering grape significantly improves aroma composition. Artificial postharvest infection by selected local B. cinerea strains during grape withering has been demonstrated to be suitable by several authors, and implementing this procedure in drying fruit rooms to standardize the level of grape botrytization is encouraged.11,27,28 For the controlled postharvest botrytization, several methods have been developed. In an internationally patented automatic system, the grape treatment is carried out in a drying room in various drying cycles. The first cycle involves dehydration at 76% relative humidity (RH) and 11°C for 30 days, followed by exposure to 100% RH for 7 days and a final cycle of 76% at 8–10°C.29 This method successfully induces activity of B. cinerea latently present on the bunches, and the resulting wine has characteristics typical of botrytized wines.5 Another method involves artificial infection of the grape with a commercial Botrytis inoculum in an induced mist and decreasing RH to 50% after 3–4 days. The whole botrytization and withering process requires 15–25 days, depending on the grape variety. Besides improving sensory characteristics of the passito wines, postharvest botrytization technologies offer a way to produce botrytized wines in the regions where it is not naturally possible or in traditional noble rot areas during bad seasons.5
Diversity of yeast and bacterium biota of botrytized grapes and wines
The microbiology of botrytized winemaking, with special regard to the alcoholic fermentation, is more complex than that of normal vinification. Its special features involve the modified composition of yeasts and bacteria on the botrytized grape and the high sugar content as well as other difficult environmental conditions of botrytized must and presence of possible inhibitors. As a result, atypical microbial communities of botrytized wines can be expected and are actually found.
Studies on the yeast biota of botrytized grapes in various countries have revealed a strong presence of Starmerella bacillaris (syn Candida zemplinina). This is known from very early research, but in various publications the name of the wine strains of the same species has been changed several times from the original Torulopsis bacillaris30 to C. stellata31 and then to C. zemplinina32–34 due to the changes in yeast taxonomy. Recently, a new classification has been suggested reinstating the species S. bacillaris, with the previous name C. zemplinina as an obligate synonym.35 In this review, however, we continue to use the synonymous name, since most of the enological studies refer to C. zemplinina.
C. zemplinina population significantly increases on botrytized berries in Tokaj during postharvest storage33 and likewise grows during withering of grapes for passito wine production;36 hence it has an important role in fermentation. C. zemplinina has several interesting enological properties: sugar tolerance,32,34 fructophilic character,22,37–39 good glycerol production,36,37,39,40 moderate or low acetic acid production,39 and a particular aroma profile.40,41 These properties make C. zemplinina a good candidate for starter culture in combination with Saccharomyces strains – not only in botrytized wines but also in normal wines as well.40–44
In botrytized wine fermentation, C. zemplinina frequently initiates the spontaneous fermentation and survives long after Saccharomyces strains have begun to dominate, particularly at lower temperatures.22,45 Its main contribution to the quality of botrytized wine is to lend a beneficial modification of the glucose–fructose ratio and enhance glycerol production.38 Its importance in the aroma composition of Tokaji Aszú was not confirmed,46 but seems significant in other wine styles.40
Various molecular genetic methods provide more and more possibilities for following the population dynamics of yeasts during fermentation. Using denaturing gradient gel electrophoresis (DGGE) during Dolce wine fermentation in California, C. zemplinina was detected as the dominant yeast throughout the early fermentation in certain vintage years.22 This species persisted throughout, with S. cerevisiae starting to appear only later in the fermentation. In other years, using terminal restriction fragment length polymorphism (RFLP)in the same wine type, the C. zemplinina population was not as pronounced at the beginning, although it was found to increase in the microbiota by the end of fermentation.23 Long prevalence of C. zemplinina was also found in fermenting Tokaji Aszú,38 Greek botrytized wine,45 and passito wine.36
Although C. zemplinina is known to be particularly associated with high sugar musts or wines, it is common on nonbotrytized grapes as well, although in lower populations. Recent genotyping of 163 wine-related strains from seven countries did not show any relationship between the genetic diversity and sweet/dry wine origin, suggesting that high sugar concentration in winemaking has no impact on C. zemplinina selection and adaptation. These surprising results indicate that C. zemplinina is not under selective pressure in winemaking environment.47 The reasons for its prevalence in botrytized wine fermentations need further explanation.
On the botrytized grape, other sugar-tolerant, non-Saccharomyces species, including Zygosaccharomyces bailii, Hanseniaspora species, Metschnikowia pulcherrima, and Candida lactis-condensi,23,33 Issatchenkia spp. or Kluyveromyces dobzhanskii,45 are present and contribute to the fermentation, although to a lesser extent and for a shorter time than C. zemplinina in general. Several M. pulcherrima are capable of making a positive contribution to volatile thiol release (3SH) in wines, essentially during the prefermentation stage in winemaking, when this microbiological subpopulation is dominant.48 Apart from C. zemplinina, a persistent dominance of Z. bailii throughout the fermentation was also reported in Botrytis-affected fermentation.45 Being sugar tolerant and resistant to various chemical preservatives, this species is considered an important spoilage yeast in winemaking.
However, the major part of fermentation of botrytized wines is normally dominated by Saccharomyces species, just as in normal wines. Concerning the population dynamics in the course of spontaneous fermentation, a late appearance and getting dominance of Saccharomyces strains are typical23,38 The Saccharomyces biota of botrytized wines seems different from that of normal wines. The most striking characteristic is the high frequency of S. bayanus var. uvarum strains in botrytized musts and fermentations.49 While S. bayanus var. uvarum is a relatively rare wine yeast in most wine regions, its significant presence was confirmed in Tokaj,31,38,50 Sauternes,51 and Amarone52,53 wines as well. Whether these Saccharomyces strains are generally present on the botrytized grape is not completely clear. From the reports, it seems that these strains can be isolated from the grape only after selective enrichment, if at all.23,33 Various studies, using different molecular methods, report on a huge genetic diversity of Saccharomyces cerevisiae in botrytized wines either in Tokaj31,50 or in Sauternes51,54 or in north Italy.52 In contrast to this, S. bayanus var. uvarum exhibits low polymorphism in chromosome size49 and also its mtDNA was found to be less polymorphic than that of S. cerevisiae.49 Enological properties of S. bayanus are significantly different from those of S. cerevisiae,37,53,55 especially regarding its high glycerol and succinic acid production and particular aroma profile (with high 2-phenylethanol production), as well as a pronounced cryophilic character. Viniculture and winemaking at low temperatures seem to be the specific ecological niche for S. bayanus var. uvarum.49
To accelerate and control the difficult fermentation process, yeast starter cultures have long been used in fermentation of Sauternes and BA/TBA, wines, as well as in the newer style botrytized wines. This practice is spreading in Tokaji Aszú fermentation too. The predictability can be improved and the microbiological diversity of fermentation can be reduced in this way, and the impact of the local yeast biota still remains significant.23,38 Of the commercial starter cultures, only a few strains can cope with the difficult conditions of botrytized must, so finding the appropriate strain requires thorough consideration and practical experiments. Selection of local strains, which are well adapted to the particular conditions, is an important perspective.38,56 The use of S. bayanus strains as single starter cultures seems sufficient only at low temperature, because the local S. cerevisiae strains easily outgrow them. Combined applications of S. bayanus and S. cerevisiae and/or non-Saccharomyces species are under study.57
Selecting a local strain or choosing commercial yeast appropriate for botrytized wine fermentation has special criteria, in addition to high sugar tolerance. Since the high sugar content and Botrytis glucans dramatically enhance the acetic acid production of yeasts, a low acetic acid production is essential.1,38 The diversity of different Saccharomyces strains in enological traits involves aroma production (higher alcohols, fermentation esters, etc), including their capacity to release volatile thiols by cleavage of S–C bonds in the precursor molecules present in grapes.14 This capacity is particularly important in fermentation of botrytized Sauvignon blanc and Sémillon wines.
In contrast to yeasts, the bacterium biota of botrytized wines is not well studied. It has long been known that acetic acid bacterium population reaches high levels on the digested berry skin, leading to an elevated acetic acid, keto-gluconic, and gluconic acid concentration in the juice, but little is known about other taxa. A recent study utilizing a next-generation sequencing technique24 has determined the bacterial diversity of botrytized wines to be far higher than previously realized. Several previously unreported bacteria (eg, Chryseobacterium, Methylobacterium, Sphingomonas, etc) were identified on wines in 2 years of Dolce fermentation. As expected, Saccharomyces inoculation exerted selective pressure on bacterial diversity in these fermentations, most notably suppressing abundance of acetic acid bacteria (Gluconobacter). In both the vintages studied, Rhodospirillales (predominantly Acetobacter, Gluconobacter, and Gluconacetobacter) were the most dominant bacteria detected, with secondary populations of Lactobacillales. The LAB community of Dolce was also abnormal compared to normal wines, comprising primarily Leuconostoc and Lactococcus in addition to the more typical Lactobacillus and Pediococcus; notably, Oenococcus was entirely absent.
Aroma composition of botrytized wines
The chemical changes induced by noble rot and complemented and combined with the yeast activities during alcoholic fermentation result in a special, very complex chemical composition of botrytized wines. The main characteristics have been well studied and reviewed by several authors.1,3,6 The most important differences compared to normal wines involve residual sugar content from the grape, high glycerol and gluconic acid concentrations (considered Botrytis indicators in the juice and wine), modified organic acid pattern, high level of carbonyl compounds (with high sulfite binding capacity), and low level of yeast-assimilable nitrogen. A further specialty of botrytized wines is the higher polyphenol content and antioxidant capacity as well as a particular polysaccharide and protein composition arising from the noble rotted grape. These properties lead to technological difficulties in stabilization and clarification, as reviewed in earlier publications.
Concerning the studies on the chemical composition of botrytized wines, the most significant development can be found in the aroma research. The particular aroma ranges of various botrytized wines are determined by several interacting factors. The primary factors are the Botrytis activity and the respective metabolic responses of the grape. In addition, alcoholic fermentation (yeast, temperature, and nutrients) and maturation of wines (time, level of oxidation, sulfite addition, and type of cooperage) significantly influence the final style and sensory character of these wines.
The volatile compounds identified in sweet botrytized wines belong to numerous chemical classes (Table 1). Earlier research showed that while the terpene content typically decreases in these wines, numerous hydroxy-, oxo-, and dicarboxylic acid esters, acetals, and lactones form, which are present in lower concentrations or absent in normal wines.58–61
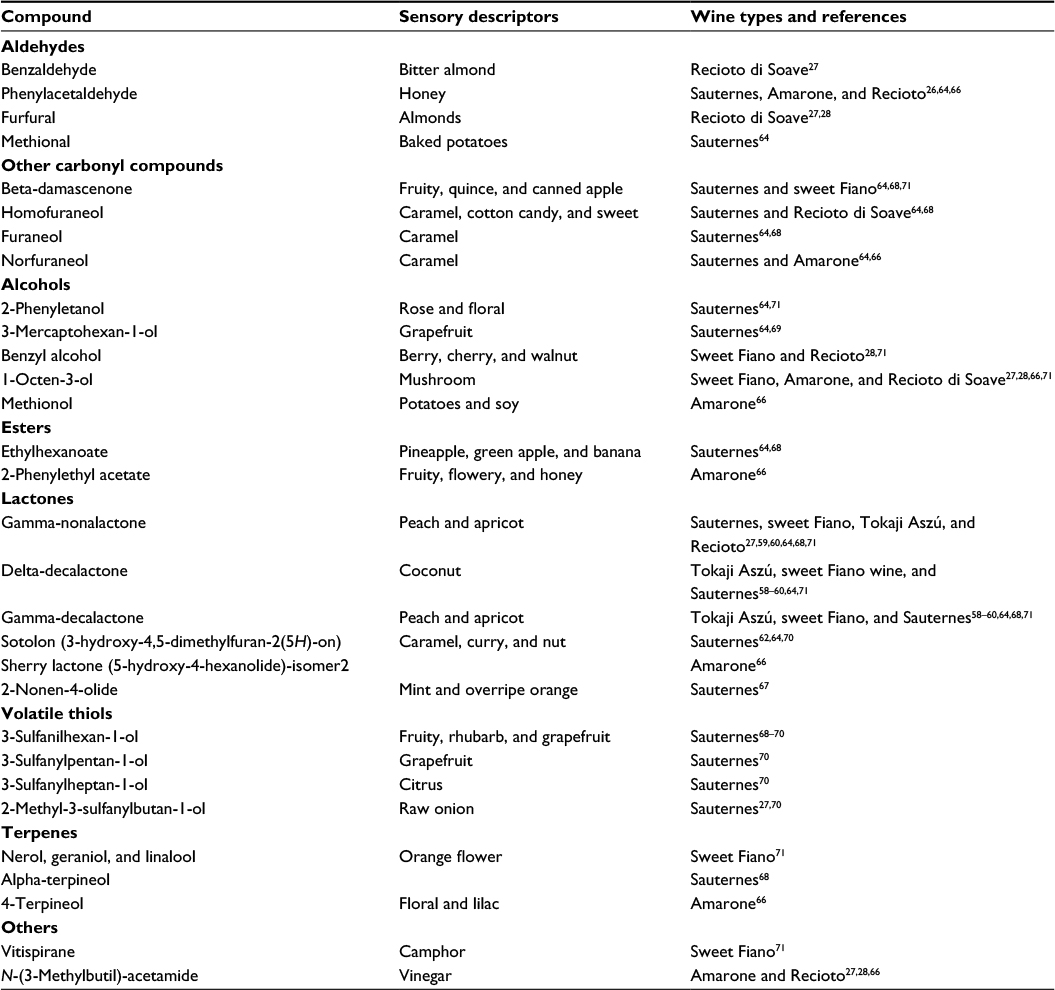 | Table 1 Volatile compounds significantly associated with botrytized wines as characteristic aroma substances |
The first research on the chemical nature of botrytized aroma character focused mainly on heterocyclic odoriferous compounds. One of the first compounds identified as a key odorant in these wines was sotolon (4,5-dimethyl-3-hydroxy-2(5H)-furanone), having a sweetish, caramel, or nut taste.62 Its presence has been repeatedly confirmed in botrytized wines.63,64 However, it is also a typical compound in some nonbotrytized wines such as flor sherry and fortified sweet wines such as Port.65
Among heterocyclic compounds, beta-lactones (coconut) and gamma-lactones (peach and apricot)20,59,61,64,66 were confirmed as key aroma compounds. The use of the gas chromatography/mass spectrometry/olfactometry technique has significantly contributed to the understanding of botrytized aroma character. More recently, a new lactone, 2-nonen-4-olide, was identified in Sauternes wines.67 While this molecule alone presents minty and fruity nuances, together with other volatile compounds it contributes to the overripe fruit (particularly orange) aroma and typicality in Sauternes sweet wines.
In addition to lactones, higher alcohols, aldehydes, and esters, eg, ethyl-hexanoate (pineapple and banana), phenylethanol (rose), phenylacetaldehyde (honey), beta-damascenone (fruity, quince, and canned apple) and different furanones (caramel), have important roles in the distinct aroma of botrytized wines.64,68 These compounds, however, do not fully explain the citrus-like nuances. In the last decade, studies on Sauternes wines using gas chromatography–olfactometry methods revealed the importance of polyfunctional thiols in this respect, especially sulfanylaldehydes, sulfanylalcohols, and sulfanylesters.
Volatile thiols are extremely strong odorants; many of them bad smelling (eg, onion), but a few have pleasant, citrus-like odor. Several volatile thiols are an important part of the varietal character of Sauvignon blanc and Semillon wines made from healthy grapes as well.69 Tominaga et al69 presented first that 3-sulfanylhexan-1-ol (3SH) is present at very high concentration and contributes to the citrus/grapefruit nuance of Sauternes sweet wines. During the last decade, several new volatile thiols have been identified as important odorants of Sauternes wines, such as 3-methyl-3-sulfanylbutanal (bacon-petroleum),63 3-sulfanylpentan-1-ol (3SP, citrus and sulfur), 3-sulfanylheptan-1-ol (3SHp, grapefruit), and 2-methyl-3-sulfanylbutan-1-ol (2M3SB, raw onion).70 Thiols are practically not present in grape, only in wines, and are released from precursor molecules during alcoholic fermentation, as presented in a previous section. Botrytis infection highly enhances the formation of the precursors in the grape, and the various yeast strains have a further impact on the final composition of thiols in the wine.
These important findings on the role of volatile thiols in Botrytis aroma originated from the studies of botrytized Sauvignon blanc and Semillon wines in the Sauternes region. Whether these molecules have similar importance in botrytized wines from other grape varieties (eg, Furmint in Tokaj or Riesling, Gewürztraminer in Germany, Chenin blanc in South Africa) has remained unclear and needs further research.
As discussed in the “Induced botrytization” section, there is an increasing interest in improving the aroma composition with notes of botrytization in some passito wines. This might occur naturally on the grapevine or as a postharvest process during the withering of the grape in climatized chambers, but artificial botrytization is also a strategy in some wineries. Table 2 shows the comparison of the main volatile compounds most affected by Botrytis in a dry red passito wine Amarone, when healthy grape or partially botrytized grape was vinified.66 These data clearly show the great impact of Botrytis and with respect to the various compounds, they generally confirm the earlier findings obtained from traditional sweet botrytized wines. In another study, the combined effect of Botrytis and the yeast strain was studied in Recioto di Soave microvinifications.27 Although the effect of the botrytization on wine aroma was greater (revealing Botrytis products such as 1-octen-3-ol, gamma-nonalactone, and benzaldehyde) than that of yeasts, the contribution of the latter also proved significant. Large variations between yeasts (local strains or commercial starters) were observed for several compounds (eg, ethyl esters, lactones, phenylacetaldehyde, isovaleric acid, and N-(3-methylbutyl)acetamide). The results indicate the importance of the choice of strain to modulate wine flavor in passito wines.
 | Table 2 Aroma compounds detected by SPE-GC-MS in dry, red Amarone wines obtained from healthy grape must (W-100) and from mixed healthy and botrytized grape musts at different percentages (W-80: 20% botrytized, and W60: 40% botrytized) Notes: Of the 53 compounds determined only the compounds whose concentrations varied statistically >30% in botrytized wines with respect to healthy grape wine are shown. Adapted with permission from Fedrizzi B, Tosi E, Simonato B, et al. Changes in wine aroma composition according to botrytized berry percentage: a preliminary study on Amarone wine. Food Technol Biotechnol. 2011;49(4):529–535.66 Mean values with different letters in superscript within a row are significantly different (P<0.05). Abbreviation: SPE-GC-MS, gas chromatography-mass spectrometry. |
Botrytized wines are typically aged for a long time either in barrels (like Tokaji Aszú) or in bottles (Sauternes, TBA wines). The length, the oxygen ingress, the level of SO2, and the material of the cooperage have a huge impact on the aroma composition of the new botrytized wines. Although the variations in these factors significantly determine the style of the finished wine, these effects are not as fully researched as the impact of botrytization.
In a survey on aged Sauternes wines,68 most polyfunctional thiols (3-sulfanylpropyl acetate, 2-sulfanylethyl acetate, 3-methyl-3-sulfanylbutanal, etc) completely disappeared after 2 years of bottle aging in a cellar. Only 3-sulfanylhexan-1-ol (3SH) was found in aged samples at concentrations above its threshold value. In contrast to the thiols, most other key odorants found in the young noble rot wine were still detected 5–6 years after harvest. In the same study, a new lactone, abhexon, was found, for the first time, to be synthesized during bottle aging.
Fedrizzi et al25 compared Amarone wines from botrytized and nonbotrytized grapes, under oxidative and anaerobic conditions. This research highlighted peculiar evolutions for several volatile compounds in botrytized wine. In particular, benzaldehyde showed a tangible increment during the oxidative aging, with a rate much higher than that reported for non-Botrytis red wines. On the other hand, several sulfides disappeared after just 15 days of oxidative aging.
Conclusion
Noble rot of the grape is a complex microbiological and biochemical process, which has long been under investigation. The older studies elucidated the direct role of Botrytis enzymes in transformation of the substrates present in grape. While confirming most of the previous findings, recent publications apply a transcriptomic and metabolomic approach to understand the response of the grape plant to the stress caused by Botrytis under the conditions of noble rot. These responses led to a modulation of the grape metabolism (eg, enhanced production of certain polyphenols such as stilbenes and anthocyanins, terpenes, or cysteinylated thiol precursors), which contribute to the distinct composition and sensory character of these wines.
In recent years, benefits of the botrytization process have been thoroughly studied and successfully implemented in the technology of passito wines, where the postharvest withering of grapes in fruit-drying rooms is the key process for the production. Another significant area of current research is the diversity of microbiota and its importance in alcoholic fermentation. Use of carefully selected Saccharomyces and non-Saccharomyces yeast species as oligo starter culture is a promising direction in the control of the difficult fermentation process.
Thanks to the developments of the analytical methods, aroma composition of botrytized wines is under extensive research, elucidating the various effects (grape, Botrytis, dehydration, yeasts, etc). These investigations widely confirmed the old results and presented many new findings.
Producing botrytized wines is a demanding task for winemakers. In spite of significant advances, this area needs further research. The multivariate interactions among grape variety, Botrytis, yeast species, wine matrix, and temperature effects make this topic very complex and in need of better understanding. Timing and implementation of harvest, processing the special grape, induction and stoppage of alcoholic fermentation, preventing sweet wines from refermentation during the time of aging, and chemical and microbiological stabilization present the major technological challenges and need further research of an applied nature.
Disclosure
The authors report no conflicts of interest in this work.
References
Magyar I. Botrytized wines. Adv Food Nutrit Res. 2011;63:147–206. | ||
Kerényi Z. Tokaj. In: Sweet, Reinforced and Fortified Wines: Grape Biochemistry, Technology and Vinification. Editors: F Mencarelli and P Tonutti. John Wiley & Sons; Oxford; 2013:269–276. | ||
Dittrich HH, Grossmann H. Mikrobiologie des Weines. 4th ed. Stuttgart: Ulmer; 2011. | ||
Teissedre LP, Donèche B. Botrytized wines: sauternes, German wines. In: Sweet, Reinforced and Fortified Wines: Grape Biochemistry, Technology and Vinification. Editors: F Mencarelli and P Tonutti. John Wiley & Sons; Oxford; 2013:285–299. | ||
Vannini A, Chilosi G. Botrytis infection: grey mould and noble rot. In: Sweet, Reinforced and Fortified Wines: Grape Biochemistry, Technology and Vinification. John Wiley and Sons; Oxford; 2013:159–169. | ||
Ribéreau-Gayon P, Dubourdieu D, Donèche B, Lonvaud A. Botrytized sweet wines (Sauternes and Tokay). Handbook Enol. 2006;1:449–458. | ||
Vivas N, De Vivas Gaulejac N, Vitry C, et al. Occurrence and specificity of glucose oxidase (E.C: 1.1.3.4) in botrytized sweet white wine. Comparison with lacease (E.C: 1.10.3.2), considered as the main responsible factor for oxidation in this type of wine. Vitis J. Grapevine Res. 2010;49(3):113–120. | ||
Salgues M, Cheynier V, Gunata Z, Wylde R. Oxidation of grape juice 2-s-glutathionyl caffeoyl tartaric acid by Botrytis cinerea laccase and characterization of a new substance: 2,5-di-S-glutathionyi caffeoyl tartaric acid. J Food Sci. 1986;51(5):1191–1194. | ||
Hong YS, Martinez A, Liger-Belair G, Jeandet P, Nuzillard JM, Cilindre C. Metabolomics reveals simultaneous influences of plant defence system and fungal growth in Botrytis cinerea-infected Vitis vinifera cv. Chardonnay berries. J Exp Bot. 2012;63(16):5773–5785. | ||
Blanco-Ulate B, Amrine KCH, Collins TS, et al. Developmental and metabolic plasticity of white-skinned grape berries in response to botrytis cinerea during noble rot. Plant Physiol. 2015;169(4):2422–2443. | ||
Lorenzini M, Azzolini M, Tosi E, Zapparoli G. Postharvest grape infection of Botrytis cinerea and its interactions with other moulds under withering conditions to produce noble-rotten grapes. J Appl Microbiol. 2013;114(3):762–770. | ||
Carbajal-Ida D, Maury C, Salas E, Siret R, Mehinagic E. Physico-chemical properties of botrytised Chenin blanc grapes to assess the extent of noble rot. Eur Food Res Technol. 2016;242(1):117–126. | ||
Thibon C, Shinkaruk S, Jourdes M, Bennetau B, Dubourdieu D, Tominaga T. Aromatic potential of botrytized white wine grapes: identification and quantification of new cysteine-S-conjugate flavor precursors. Anal Chim Acta. 2010;660(1–2):190–196. | ||
Thibon C, Marullo P, Claisse O, Cullin C, Dubourdieu D, Tominaga T. Nitrogen catabolic repression controls the release of volatile thiols by Saccharomyces cerevisiae during wine fermentation. FEMS Yeast Res. 2008;8(7):1076–1086. | ||
Thibon C, Dubourdieu D, Darriet P, Tominaga T. Impact of noble rot on the aroma precursor of 3-sulfanylhexanol content in Vitis vinifera L. cv Sauvignon blanc and Semillon grape juice. Food Chem. 2009;114(4):1359–1364. | ||
Thibon C, Cluzet S, Mérillon JM, Darriet P, Dubourdieu D. 3-slfanylhexanol precursor biogenesis in grapevine cells: the stimulating effect of Botrytis cinerea. J Agric Food Chem. 2011;59(4):1344–1351. | ||
Dewey FM, Hill M, DeScenzo R. Quantification of botrytis and laccase in winegrapes. Am J Enol Vitic. 2008;59(1):47–54. | ||
Dewey FM, Meyer U. Rapid, quantitative Tube immunoassays for on-site detection of Botrytis, Aspergillus and Penicillium antigens in grape juice. Anal Chim Acta. 2004;513(1):11–19. | ||
Lorenzini M, Millioni R, Franchin C, Zapparoli G, Arrigoni G, Simonato B. Identification of potential protein markers of noble rot infected grapes. Food Chem. 2015;179:170–174. | ||
Rolle L, Giordano M, Giacosa S, et al. CIEL *a *b * parameters of white dehydrated grapes as quality markers according to chemical composition, volatile profile and mechanical properties. Anal Chim Acta. 2012;732:105–113. | ||
Soós J, Várvölgyi E, Bázár G, Felföldi J, Magyar I, Kovács Z. Electronic tongue in botrytized wine authentication. Poster presented at: 16th International Symposium on Olfaction and Electronic Nose, June 28-July 1; 2015; Dijon. | ||
Mills DA, Johannsen EA, Cocolin L. Yeast diversity and persistence in botrytis-affected wine fermentations. Appl Environ Microbiol. 2002;68(10):4884–4893. | ||
Bokulich NA, Hwang CF, Liu S, Boundy-Mills KL, Mills DA. Profiling the yeast communities of wine fermentations using terminal restriction fragment length polymorphism analysis. Am J Enol Vitic. 2012;63(2):185–194. | ||
Bokulich NA, Joseph CML, Allen G, Benson AK, Mills DA. Next-generation sequencing reveals significant bacterial diversity of botrytized wine. PLoS One. 2012;7(5):e36357. | ||
Fedrizzi B, Zapparoli G, Finato F, et al. Model aging and oxidation effects on varietal, fermentative, and sulfur compounds in a dry botrytized red wine. J Agric Food Chem. 2011;59(5):1804–1813. | ||
Tosi E, Fedrizzi B, Azzolini M, Finato F, Simonato B, Zapparoli G. Effects of noble rot on must composition and aroma profile of Amarone wine produced by the traditional grape withering protocol. Food Chem. 2012;130(2):370–375. | ||
Azzolini M, Tosi E, Faccio S, Lorenzini M, Torriani S, Zapparoli G. Selection of Botrytis cinerea and Saccharomyces cerevisiae strains for the improvement and valorization of Italian passito style wines. FEMS Yeast Res. 2013;13(6):540–552. | ||
Tosi E, Azzolini M, Lorenzini M, et al. Induction of grape botrytization during withering affects volatile composition of Recioto di Soave, a “passito”-style wine. Eur Food Res Technol. 2013;236(5):853–862. | ||
Ferrarini R, Casarotti EM, Zanella G. Botrytis cinerea noble form induction on grapes during withering. Am J Enol Vitic. 2009:60.399A. | ||
Peynaud E, Domercq MC. [Study of the yeasts of the Gironde] Etude des levures de la Gironde. Ann Technol Agric. 1953;4:265–300. | ||
Antunovics Z, Csoma H, Sipiczki M. Molecular and genetic analysis of the yeast flora of botrytized Tokaj wines. Bull OIV. 2003;76:380–397. | ||
Sipiczki M. Candida zemplinina sp. nov., an osmotolerant and psychrotolerant yeast that ferments sweet botrytized wines. Int J Syst Evol Microbiol. 2003;53(6):2079–2083. | ||
Magyar I, Bene Z. Morphological and taxonomic study on mycobiota of noble rotted grapes in the Tokaj wine district. Acta Aliment. 2006;35(2):237–246. | ||
Tofalo R, Chaves-López C, Di Fabio F, et al. Molecular identification and osmotolerant profile of wine yeasts that ferment a high sugar grape must. Int J Food Microbiol. 2009;130(3):179–187. | ||
Duarte FL, Pimentel NH, Teixeira A, Fonseca A. Saccharomyces bacillaris is not a synonym of Candida stellata: reinstatement as Starmerella bacillaris comb. nov. Antonie Van Leeuwenhoek Int J Gen Mol Microbiol. 2012;102(4):653–658. | ||
Rantsiou K, Campolongo S, Alessandria V, Rolle L, Torchio F, Cocolin L. Yeast populations associated with grapes during withering and their fate during alcoholic fermentation of high-sugar must. Aust J Grape Wine Res. 2013;19(1):40–46. | ||
Magyar I, Tóth T. Comparative evaluation of some oenological properties in wine strains of Candida stellata, Candida zemplinina, Saccharomyces uvarum and Saccharomyces cerevisiae. Food Microbiol. 2011;28(1):94–100. | ||
Magyar I, Tóth T, Pomázi A. Oenological characterization of indigenous yeasts involved in fermentation of Tokaji Aszú. Bull OIV. 2008;81:35–43. | ||
Tofalo R, Schirone M, Torriani S, et al. Diversity of Candida zemplinina strains from grapes and Italian wines. Food Microbiol. 2012;29(1):18–26. | ||
Englezos V, Torchio F, Cravero F, et al. Aroma profile and composition of Barbera wines obtained by mixed fermentations of Starmerella bacillaris (synonym Candida zemplinina) and Saccharomyces cerevisiae. LWT – Food Sci Technol. 2016;73:567–575. | ||
Andorrà I, Berradre M, Mas A, Esteve-Zarzoso B, Guillamón JM. Effect of mixed culture fermentations on yeast populations and aroma profile. LWT – Food Sci Technol. 2012;49(1):8–13. | ||
Comitini F, Gobbi M, Domizio P, et al. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011;28(5):873–882. | ||
Suzzi G, Arfelli G, Schirone M, Corsetti A, Perpetuini G, Tofalo R. Effect of grape indigenous Saccharomyces cerevisiae strains on Montepulciano d’Abruzzo red wine quality. Food Res Int1. 2012;46(1):22–29. | ||
Di Maio S, Genna G, Gandolfo V, Amore G, Ciaccio M, Oliva D. Presence of Candida zemplinina in sicilian musts and selection of a strain for wine mixed fermentations. S Afr J Enol Vitic. 2012;33(1):80–87. | ||
Nisiotou AA, Spiropoulos AE, Nychas GJE. Yeast community structures and dynamics in healthy and Botrytis-affected grape must fermentations. Appl Environ Microbiol. 2007;73(21):6705–6713. | ||
Tóth-Markus M, Magyar I, Kardos K, Bánszky L, Maráz A. Study of Tokaji Aszú wine flavour by solid phase microextraction method. Acta Aliment. 2002;31(4):343–354. | ||
Masneuf-Pomarede I, Juquin E, Miot-Sertier C, et al. The yeast Starmerella bacillaris (synonym Candida zemplinina) shows high genetic diversity in winemaking environments. FEMS Yeast Res. 2015;15(5):fov045. | ||
Zott K, Thibon C, Bely M, Lonvaud-Funel A, Dubourdieu D, Masneuf-Pomarede I. The grape must non-Saccharomyces microbial community: impact on volatile thiol release. Int J Food Microbiol. 2011;151(2):210–215. | ||
Naumov GI, Naumova ES, Martynenko NN, Masneuf-Pomaréde I. Taxonomy, ecology, and genetics of the yeast Saccharomyces bayanus: a new object for science and practice. Microbiology. 2011;80(6):735–742. | ||
Naumov GI, Naumova ES, Antunovics Z, Sipiczki M. Saccharomyces bayanus var. uvarum in Tokaj wine-making of Slovakia and Hungary. Appl Microbiol Biotechnol. 2002;59(6):727–730. | ||
Naumov GI, Masneuf I, Naumova ES, Aigle M, Dubourdieu D. Association of Saccharomyces bayanus var. uvarum with some French wines: genetic analysis of yeast populations. Res Microbiol. 2000;151(8):683–691. | ||
Torriani S, Zapparoli G, Suzzi G. Genetic and phenotypic diversity of Saccharomyces sensu stricto strains isolated from Amarone wine. Antonie Van Leeuwenhoek Int J Gen Mol Microbiol. 1999;75(3):207–215. | ||
Tosi E, Azzolini M, Guzzo F, Zapparoli G. Evidence of different fermentation behaviours of two indigenous strains of Saccharomyces cerevisiae and Saccharomyces uvarum isolated from Amarone wine. J Appl Microbiol. 2009;107(1):210–218. | ||
Börlin M, Venet P, Claisse O, Salin F, Legras JL, Masneuf-Pomarede I. Cellar-associated Saccharomyces cerevisiae population structure revealed high-level diversity and perennial persistence at Sauternes wine estates. Appl Environ Microbiol. 2016;82(10):2909–2918. | ||
Masneuf-Pomarède I, Bely M, Marullo P, Lonvaud-Funel A, Dubourdieu D. Reassessment of phenotypic traits for Saccharomyces bayanus var. uvarum wine yeast strains. Int J Food Microbiol. 2010;139(1–2):79–86. | ||
Miki T, Ito Y, Kuroha K, Izawa S, Shinohara T. Potential of yeasts isolated in botrytized grape juice to be new wine yeasts. Food Sci Technol Res. 2008;14(4):345–350. | ||
Bely M, Stoeckle P, Masneuf-Pomarède I, Dubourdieu D. Impact of mixed Torulaspora delbrueckii-Saccharomyces cerevisiae culture on high-sugar fermentation. Int J Food Microbiol. 2008;122(3):312–320. | ||
Schreier P, Drawert F, Kerènyi Z, Junker A. GLC-mass-spectrometrical investigation of the volatile components of wines VI. Aroma compounds of Tokaj Aszu wines. a) Neutral compounds. Z Lebensm Unters Forch. 1976;161(3):249–258. | ||
Miklósy É, Kalmár Z, Kerényi Z. Identification of some characteristic aroma compounds in noble rotted grape berries and Aszú wines from Tokaj by GC-MS. Acta Aliment. 2004;33(3):215–226. | ||
Miklósy É, Kalmár Z, Pölös V, Kerényi Z. Study of volatile aroma components in young Tokaji Aszu wines by GC-MS. Chromatographia. 2000;51(suppl 2):S305–S308. | ||
Miklósy E, Kerényi Z. Comparison of the volatile aroma components in noble rotted grape berries from two different locations of the Tokaj wine district in Hungary. Anal Chim Acta. 2004;513(1):177–181. | ||
Masuda M, Okawa EIC, Nishimura KIC, Yunome H. Identification of 4,5-dimethyl-3-hydroxy-2(5//)-furanone (sotolon) and ethyl 9-hydroxynonanoate in botrytised wine and evaluation of the roles of compounds characteristic of it. Agric Biol Chem. 1984;48(11):2707–2710. | ||
Bailly S, Jerkovic V, Marchand-Brynaert J, Collin S. Aroma extraction dilution analysis of Sauternes wines. Key role of polyfunctional thiols. J Agric Food Chem. 2006;54(19):7227–7234. | ||
Sarrazin E, Dubourdieu D, Darriet P. Characterization of key-aroma compounds of botrytized wines, influence of grape botrytization. Food Chem. 2007;103(2):536–545. | ||
Ferreira A, Barbe J, Ertrand A. 3-hydroxy- 4,5-dimethyl- 2(5H)-furanone: a key odorant of the typical aroma of oxidative aged Port wine. J Agric Food Chem. 2003;51(15):4356–4363. | ||
Fedrizzi B, Tosi E, Simonato B, et al. Changes in wine aroma composition according to botrytized berry percentage: a preliminary study on Amarone wine. Food Technol Biotechnol. 2011;49(4):529–535. | ||
Stamatopoulos P, Frérot E, Darriet P. Evidence for perceptual interaction phenomena to interpret typical nuances of ‘overripe’ fruity aroma in bordeaux dessert wines. In: Guthrie B, Beauchamp J, Buettner A, Lavine BK, editors. ACS Symposium Series. Vol. 1191. American Chemical Society; Washington. 2015:87–101. | ||
Bailly S, Jerkovic V, Meurée A, Timmermans A, Collin S. Fate of key odorants in sauternes wines through aging. J Agric Food Chem. 2009;57(18):8557–8563. | ||
Tominaga T, Baltenweck-Guyot R, Peyrot des Gachons C, Dubourdieu D. Contribution of volatile thiols to the aromas of white wines made from several Vitis vinifera grape varieties. Am J Enol Vitic. 2000;51(2):178–181. | ||
Sarrazin E, Shinkaruk S, Tominaga T, Bennetau B, Frérot E, Dubourdieu D. Odorous impact of volatile thiols on the aroma of young botrytized sweet wines: identification and quantification of new sulfanyl alcohols. J Agric Food Chem. 2007;55(4):1437–1444. | ||
Genovese A, Gambuti A, Piombino P, Moio L. Sensory properties and aroma compounds of sweet Fiano wine. Food Chem. 2007;103(4):1228–1236. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.