Back to Journals » International Journal of Nanomedicine » Volume 21
Bidirectional Interactions Between Nanomaterials and Extracellular Vesicles: Molecular Insights and Translational Implications
Authors Sa J, Lv Y, He J, Wang K ![]() , Zhou J
, Zhou J
Received 1 December 2025
Accepted for publication 4 April 2026
Published 13 April 2026 Volume 2026:21 585732
DOI https://doi.org/10.2147/IJN.S585732
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Jie Huang
Jiazu Sa,1– 4,* YanCheng Lv,1– 4,* Junxian He,1– 4,* Ke Wang,5,6 Jie Zhou1– 4
1Department of Neurosurgery, The Affiliated Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China; 2Sichuan Clinical Medical Research Center for Neurosurgery, The Affiliated Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China; 3Neurological Diseases and Brain Function Laboratory, the Affiliated Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China; 4Academician (Expert) Workstation of Sichuan Province, the Affiliated Hospital of Southwest Medical University, Luzhou, 646000, People’s Republic of China; 5Key Laboratory of Advanced Technologies of Materials, Ministry of Education, School of Materials Science and Engineering, Southwest Jiaotong University, Chengdu, 610031, People’s Republic of China; 6Institute of Biomedical Engineering, College of Medicine, Southwest Jiaotong University, Chengdu, 610031, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ke Wang, Email [email protected] Jie Zhou, Email [email protected]
Abstract: Extracellular vesicles (EVs) are nanoscale carriers mediating intercellular communication via proteins, nucleic acids, and lipids. Concurrently, diverse nanomaterials (NMs) with tunable properties have emerged. Bidirectional interactions between NMs and EVs impact disease pathogenesis, diagnostics, and therapeutics. With translational relevance, NMs’ physicochemical features (size, charge, coatings) modulate EV release, cargo enrichment (proteins, miRNAs, lipids), and downstream processes including immune regulation, angiogenesis, and tumor microenvironment remodeling. Conversely, EVs internalize NMs, inducing oxidative stress, inflammation, or apoptosis. However, while NMs’ regulatory effects on EVs is well-characterized, the mechanisms of EVs-mediated NMs internalization, processing, and redistribution remain poorly understood—an asymmetry defining research priorities. Unresolved controversies persist—microenvironment-dependent immune effects, inconsistent cargo alterations, and the impact of EVs heterogeneity. This review synthesizes current knowledge on mutual NMs-EVs influences, emphasizing NMs-regulated EVs biogenesis/cargo/function and EVs uptake/response to NMs. We introduce the “NMs-EVs regulatory axis”—a bidirectional, dynamic network where NMs and EVs mutually shape function across molecular, cellular, and tissue levels, moving beyond linear carrier views. These interactions enable hybrid platforms for drug delivery, imaging, and diagnostics, positioning the NMs-EVs axis as a frontier in nanomedicine for precision therapeutics.
Keywords: intercellular communication, biosynthesis regulation, cargo remodeling, immune modulation, theranostic platforms, nanotoxicology
Introduction
Extracellular vesicles (EVs) are lipid bilayer-enclosed, non-self-replicating nanoscale particles that are widely present in various cell types and their secreted bodily fluids, including blood, urine, saliva, cerebrospinal fluid, and breast milk.1,2 According to the latest recommendations of the International Society for Extracellular Vesicles (ISEV), the subtyping of EVs still faces methodological challenges. Although EVs are traditionally classified based on size and biogenesis pathways into exosomes (30–150 nm), micro-vesicles (100–1000 nm), and apoptotic bodies (>1 um), current isolation techniques struggle to achieve absolute purification of each subtype, and there is overlap in size and shared markers among different subtypes. Based on their biogenesis pathways and size, EVs are typically classified into three categories. The first category consists of exosomes (30–150 nm) originating from the endosomal pathway, which are released after fusion of multivesicular bodies (MVBs) with the plasma membrane.2,3 Additionally, microvesicles (100–1000 nm) are formed by direct budding from the plasma membrane, and apoptotic bodies (>1 μm) originate from the fragmentation of the plasma membrane during apoptosis.4 EVs serve as information nanocarriers for intercellular communication, capable of transporting various functional molecules such as proteins, lipids, DNA, mRNA, and miRNA between different cell types and tissues, thereby participating in a wide range of physiological and pathological processes.4 In the immune system, EVs can exert dual effects of immune promotion or suppression by presenting antigens, regulating cytokine secretion, and altering the activation state of immune cells.5 In the tumor microenvironment, tumor-derived EVs can promote angiogenesis, inhibit immune cell killing functions, and create conditions for tumor metastasis by remodelling the matrix and inducing the pre-metastatic microenvironment in distant organs.5 Additionally, EVs play important roles in the development of neurodegenerative diseases, cardiovascular diseases, metabolic diseases, and others.5,6 Due to their stable presence in bodily fluids and their molecular characteristics reflecting cellular states, EVs are emerging as important candidate biomarkers for liquid biopsy and demonstrate potential in overcoming delivery barriers such as the blood-brain barrier (BBB). Despite their functional versatility, EVs exhibit remarkable heterogeneity in size, molecular cargo, membrane composition, and cellular origin. This heterogeneity, while biologically meaningful, poses significant challenges for studying NMs-EVs interactions: different EVs subpopulations may respond differently to NMs exposure, and bulk analysis of mixed EVs populations can obscure subtype-specific effects or even lead to conflicting conclusions. It is imperative to comprehend the manner in which NMs-EVs interactions are modulated by EVs heterogeneity to accurately interpret experimental data and promote the advancement of translational applications.
In recent years, the rapid development of nanomaterials (NMs) has provided multidimensional tools and strategies for life sciences and medical research.7 Nanoparticles (NPs) typically refer to structural units with characteristic dimensions ranging from 1 to 100 nm, but in practical applications, their size can be extended to 200 nm or even larger depending on functional requirements.8 Their composition can be inorganic, organic polymers, or bio-derived. Among these, the size, surface charge, and surface modification of NPs are the core parameters determining their in vivo behavior and biocompatibility.8,9 Additionally, surface charge influences particle-cell membrane interactions, for example, positively charged particles tend to bind to negatively charged cell membranes, enhancing cellular uptake efficiency but potentially increasing plasma protein adsorption and immune clearance risks.10 On the other hand, surface modification can significantly prolong circulation half-life, improve stability, and reduce non-specific adsorption.4 However, in certain cases, it may trigger anti-PEG immune responses, affecting repeated dosing efficacy.11
Multiple studies have demonstrated that NMs not only exhibit unique advantages in drug delivery and imaging diagnostics but also engage in complex bidirectional interactions with EVs.3,12 Additionally, exposure to exogenous or medical NMs can influence the biosynthesis, secretion levels, cargo types, and downstream biological effects of EVs by altering the dynamics of intracellular membrane systems, the activation status of signaling pathways, and cellular stress responses.13 Moreover, engineering strategies can combine NMs with EVs and, through surface modification with ligands, antibodies, or signal peptides, endow EVs with enhanced targeting and tissue penetration capabilities.3,14
Based on this, this paper will systematically explore the latest research progress on the interaction between NMs and EVs, focusing on two aspects. First, the regulatory mechanisms of NMs on the biosynthesis, secretion patterns, and functions of EVs, including cell skeleton restructuring, membrane dynamics changes, and regulatory pathways mediated by oxidative stress and inflammatory signals. Second, the biological responses of EVs to NMs, including uptake and distribution, metabolic degradation, immune regulation, and safety assessment. Additionally, we will delve into the application prospects of their combination in diagnostic and therapeutic fields, and discuss the challenges and strategies encountered in large-scale production, standardized quality control, and clinical translation, with the aim of providing theoretical references and technical guidelines for future integrated research in nanomedicine and EVs.
This review focuses specifically on biomedically relevant nanomaterials (eg, gold, iron oxide, silica, lipid-based nanoparticles) and their interactions with EVs derived from mammalian cells in the context of therapeutic and diagnostic applications. We exclude environmental nanomaterials such as ambient particulate matter and non-mammalian EVs systems, as their interaction dynamics and translational implications differ substantially. By integrating current knowledge on NMs-regulated EVs biogenesis, EVs-mediated NMs redistribution, and their combined functional outcomes, we aim to provide a cohesive conceptual framework-the NMs-EVs regulatory axis-that bridges fundamental nanobiology with next-generation precision medicine. Compared with previous reviews, this review systematically summarizes the current knowledge on the mutual influence between nanomaterials and extracellular vesicles, focusing on how nanomaterials regulate vesicle biogenesis, secretion, cargo composition, and functions, as well as how vesicles uptake and respond to nanomaterials. Additionally, it proposes future directions for the optimization of nanomaterials. Clinically and industrially, delineating the NMs-EVs interaction landscape is increasingly relevant. EVs are emerging as high-value biomarkers in liquid biopsy, and nano-enabled platforms, including EVs enrichment, nano-plasmonic sensing, and nanoparticle-assisted EVs capture, substantially improve diagnostic sensitivity.15 Conversely, EVs function as biocompatible carriers that enhance the biodistribution, immune evasion, and targeting precision of therapeutic nanomaterials.7,16 Understanding how NMs reshape EVs release and cargo selection informs the development of EVs-based biomarkers, improves nanotoxicity prediction, and accelerates NMs-EVs hybrid strategies for targeted therapy and theranostics.16 Collectively, this emerging regulatory axis bridges fundamental nanobiology with next-generation diagnostic and therapeutic innovation.
Influence of NMs on EVs Biogenesis and Composition
To better understand how nanomaterials shape EVs biology, it is necessary to first examine their fundamental influence on the early stages of vesicle formation. Among the various determinants, the physicochemical characteristics of NMs, including size, surface charge, and surface modifications, play a pivotal role in modulating the interaction with cellular membranes and intracellular trafficking systems. These intrinsic features not only dictate the uptake and intracellular routing of NMs, but also establish the baseline through which they can regulate EVs biogenesis.
Physicochemical Properties of NMs Affecting EVs Biogenesis
The physicochemical properties of NMs largely determine their interaction with cell membrane systems, thereby influencing the generation and composition of Evs.7 In addition, particle size is a key factor influencing the selection of endocytosis pathways and membrane fusion kinetics.17 NPs smaller than 50 nm typically enter the endosomal system rapidly via clathrin-mediated endocytosis and are closely associated with EVs biogenesis during the transport process between early endosomes and MVBs.17,18 In contrast, particles with sizes between 50 and 150 nm tend to enter cells via macropinocytosis or membrane invagination-guided endocytosis pathways, which are often accompanied by significant membrane cytoskeletal reorganization and phospholipid redistribution, thereby indirectly regulating the release of microvesicles formed by exocytosis from the plasma membrane.7,18 When particle sizes exceed 200 nm, their efficiency of entering cells significantly decreases, with more particles influencing local signal transduction through prolonged contact with membrane surface receptors, thereby regulating the release frequency and cargo loading patterns of Evs.17 Moreover, surface charge is also a key parameter. Positively charged NPs can rapidly bind to negatively charged cell membrane phospholipids and proteins. This strong electrostatic interaction not only enhances particle endocytosis efficiency but may also activate calcium ion signals and actin reorganization, possibly further promoting EVs release.18 However, positively charged particles are more likely to induce changes in cell membrane permeability and stress responses, leading to alterations in vesicle membrane composition.19 In contrast, negatively charged or neutral particles have weaker binding affinity with membranes, and their effects on EVs biogenesis are more dependent on particle chemical composition and surface modifications.19
A critical yet often overlooked determinant in NMs-EVs interactions is the distinction between the synthetic identity and the biological identity of nanomaterials. The synthetic identity refers to the engineered physicochemical properties of NMs, such as size, shape, surface charge, and functional coatings, which govern their initial interactions with biological systems. However, upon entry into physiological fluids, NMs rapidly adsorb biomolecules-predominantly proteins-to form a protein corona, which confers a new biological identity that often overrides the synthetic one. This corona not modulates cellular uptake pathways and intracellular trafficking but also redefines how NMs interface with endomembrane systems involved in EVs biogenesis. For instance, corona-coated NMs have been shown to preferentially interact with ESCRT components, thereby influencing intraluminal vesicle formation and MVB maturation. Thus, explicitly distinguishing between synthetic and biological identity provides a mechanistic framework for understanding how NMs, as perceived by cells, differentially regulate EVs-mediated communication. Coatings and surface functionalization can also directly alter the affinity of particles for cell membranes and endosomal membranes. For example, polyethylene glycol (PEG) modification can prolong particle stability in the extracellular environment and reduce non-specific protein adsorption, but may also decrease direct interaction with membranes, thereby reducing regulatory capacity on EVs biogenesis.20,21 Conversely, NPs coated with extracellular matrix proteins or ligand molecules can bind to specific receptors, triggering local endocytosis and signal transduction cascades, promoting the generation of specific types of EVs and enriching related signaling proteins or nucleic acids.22 The effects of different types of NPs on EVs biogenesis exhibit high specificity. Therefore, when investigating the mechanisms underlying the interaction between NMs and EVs, it is essential to comprehensively consider these factors to accurately assess the impact of material interventions on EVs function and their potential biomedical applications.
Beyond these physicochemical determinants, the molecular machinery governing EVs biogenesis itself constitutes a critical layer of regulation. EVs formation occurs through two major pathways: ESCRT-dependent and ESCRT-independent mechanisms, which differ fundamentally in their molecular requirements and cargo selectivity. The ESCRT-dependent pathway relies on the sequential action of ESCRT 0, -I, -II, and -III complexes to drive inward budding of the endosomal membrane, culminating in intraluminal vesicle (ILV) formation and subsequent exosome release. In contrast, ESCRT-independent pathways utilize ceramide-mediated membrane curvature, tetraspanin-enriched microdomains, or lipid raft dynamics to facilitate the packaging of distinct cargo populations. Importantly, NMs may differentially engage these pathways based on their physicochemical properties. The effects of different types of NPs on EVs biogenesis exhibit high specificity. For instance, cationic nanoparticles have been demonstrated to enhance nSMase2 activity, thereby potentially promoting ESCRT-independent EVs production. Conversely, metal oxide nanoparticles have been shown to disrupt ESCRT-III assembly through the process of oxidative stress-mediated protein damage. It is imperative to ascertain which biogenesis pathway is perturbed by a specific NM to effectively predict the composition of the resulting EVs cargo and the ensuing functional outcomes, given that ESCRT-dependent and -independent EVs exhibit distinct protein and RNA repertoires.
Beyond size and surface charge, another critical yet often overlooked determinant is the formation of the “protein corona”, which occurs immediately when NMs enter biological fluids. The corona modifies nanoparticle identity, alters receptor binding patterns, and redefines the intracellular trafficking routes that ultimately shape EVs biogenesis.23 Recent studies have demonstrated that corona-coated NMs preferentially interact with ESCRT components, thereby modulating intraluminal vesicle formation and influencing MVB maturation.4,13,24 The engineered corona structures can be exploited to program EVs loading specificity, providing a tunable interface between NMs and the vesicular sorting machinery.5
The protein corona is not a static entity but undergoes dynamic evolution over time (“hard” vs. “soft” corona) and varies dramatically across biological fluids (eg, plasma vs. interstitial fluid vs. cerebrospinal fluid). This compositional variability can fundamentally alter NMs recognition by cellular receptors and subsequent intracellular trafficking, yet most studies examine corona formation under static conditions in a single fluid type. Understanding how corona dynamics influence EVs biogenesis across different tissue microenvironments remains a critical knowledge gap.
Alteration in EVs Cargo Composition
Nano-materials not only influence the generation of EVs but also profoundly alter the molecular composition of their internal cargo, including key components such as nucleic acids, proteins, and lipids.25 This regulatory effect is often achieved through cellular signal transduction networks, stress responses, and changes in membrane system dynamics, thereby endowing EVs with new biological functions or altering their original functional patterns.25 At the nucleic acid level, different types of NPs can selectively promote the enrichment of specific miRNAs in EVs. For example, after exposure to substrates with directionally aligned nanofibres or microgroove structures, endothelial cells undergo morphological elongation and cytoskeletal redistribution.26 This biophysical stimulus drives endothelial cells to secrete EVs enriched in miR-143 and miR-145.27,28 These miRNAs can upregulate markers associated with contractile phenotypes and inhibit abnormal proliferation in vascular smooth muscle cells, thereby playing a role in maintaining vascular homeostasis and vascular remodelling. However, whether this mechanism operates in other cell types or in vivo remains to be determined.
At the protein level, stimulation by NMs can induce a series of stress proteins and signal-regulating molecules to enter Evs.29,30 Additionally, HSP70 can enhance the fusion efficiency of EVs with receptor cell membranes, increasing the effectiveness of functional molecule delivery.31 Treatment with certain metal nanoparticles can also increase the levels of integrins, adhesion molecules, and immunomodulatory proteins in EVs, which may alter the organ distribution and immune recognition patterns of EVs in vivo.
In terms of lipid composition, exposure to NMs may alter the lipid profile of EVs membranes by regulating membrane phospholipid metabolism and lipid raft structure.31,32 For example, silver nanoparticles (Ag NPs) can induce changes in the levels of membrane phosphatidylserine and sphingolipids, which not only affect the membrane stability and fusion properties of EVs but may also alter the localization of membrane-associated signaling molecules.33,34 Additionally, EVs enriched with specific lipids exhibit higher affinity in interactions with endothelial cells and immune cells, which is of significant importance for the regulation of inflammatory responses and angiogenesis (Figure 1).34
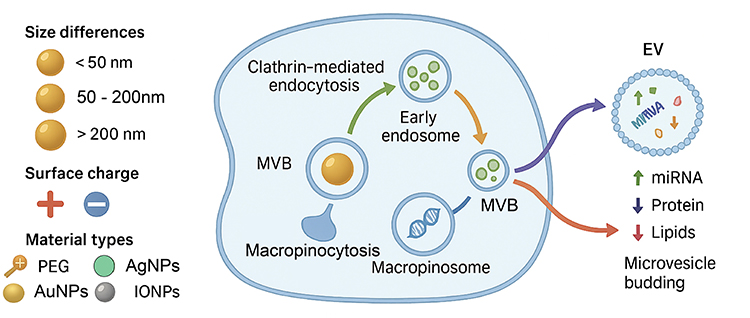 |
Figure 1 Nanomaterials with distinct physicochemical properties influence EVs biogenesis and cargo loading via multiple endocytic pathways. NMs differing in size, surface charge, and surface modifications (eg, PEG, Ag NPs, Au NPs, IO NPs) enter mammalian cells through clathrin-mediated endocytosis, caveolin-mediated endocytosis, or macropinocytosis, leading to early endosome formation and multivesicular body maturation. These endocytic routes modulate MVB dynamics and EVs release frequency, while also altering EVs cargo composition, including microRNAs (miRNAs), proteins, and lipids. Released EVs can originate from MVB-plasma membrane fusion or microvesicle budding directly from the plasma membrane. |
In addition, NMs can reshape EVs cargo profiles through mitochondrial respiration stress. In addition, NMs can reshape EVs cargo profiles through mitochondrial respiration stress. Sub-cytotoxic concentrations of metal-oxide NPs trigger mitochondrial ROS and activate NRF2/HO-1 signaling,35,36 subsequently enriching stress-responsive miRNAs such as miR-21-5p and miR-34a in EVs.37–39 However, these effects exhibit steep dose-response curves: low doses may induce adaptive stress responses (eustress) with distinct EV cargo profiles, while higher doses overwhelm cellular antioxidant capacity, leading to apoptotic EV release.40 Temporally, acute exposure favors rapid EV release with stress marker enrichment, whereas chronic exposure may select for adapted cell populations with fundamentally altered EV secretion patterns.4 These vesicles impose pro-survival or pro-senescence phenotypes on recipient cells, establishing a long-range mitochondrial stress communication axis.41,42 Collectively, these multimodal alterations in EVs cargo composition—spanning nucleic acids, proteins, and lipids—converge on a common theme: nanomaterials reprogram EVs into specialized signaling vectors that reflect the nature and intensity of the cellular stress response. The ability of NMs to impart distinct functional outcomes to EVs is attributable to a variety of mechanisms, including biophysically driven enrichment of microRNAs, stress-induced protein loading, lipid membrane remodeling, and mitochondrial stress-mediated shifts in microRNAs. These phenomena encompass vascular homeostasis, immune modulation, inflammatory amplification, and long-range stress communication. Significantly, the functional convergence of these cargo classes indicates that EVs-mediated intercellular signaling functions as an integrated readout of nanomaterial exposure, thereby positioning EVs as biosensors and effectors in the cellular response to nano-bio interactions.
A critical distinction must be drawn between stress-induced EVs—released under pathological or toxicological conditions and characterized by enrichment of damage-associated molecular patterns (DAMPs), stress proteins, and pro-inflammatory miRNAs—and physiologically functional EVs that mediate homeostatic intercellular communication. NMs exposure can blur this distinction, as even sub-cytotoxic concentrations may induce stress responses that qualitatively alter EVs cargo, potentially transforming homeostatic signaling vectors into mediators of pathology. Whether such stress-induced EVs represent adaptive responses or early indicators of toxicity remains an active area of investigation.
Functional Impact of Nanomaterial Modulated EVs
Having established how NMs shape the biogenesis and molecular composition of EVs, it is equally important to consider the downstream functional consequences of these modifications. Alterations in EVs quantity, cargo profile, and membrane properties inevitably translate into changes in their biological activities, influencing a wide spectrum of cellular processes and tissue-level responses. The following sections summarize the major domains in which nanomaterial-modulated EVs exert their impact, beginning with their central role in mediating cell-cell communication.
Impact on Cell-Cell Communication
EVs serve as important mediators of intercellular communication, regulating the signal networks and functional states of receptor cells by transporting bioactive molecules such as proteins, nucleic acids, and lipids during various physiological and pathological processes.43 Previous studies have shown that NMs can alter the cargo profile and membrane properties of EVs, thereby reshaping their role in intercellular communication and influencing the homeostasis and response patterns of the tissue microenvironment.43,44 This regulation has duality, as it may confer new functional advantages to EVs while also leading to abnormal activation or inhibition of existing signaling pathways.
The study of metal and oxide nanoparticles provides a typical example. Ferrite iron oxide nanoparticles (Fe3O4, IONPs) have been shown to regulate the levels of immune-regulatory molecules in tumor cell-derived EVs, leading to a significant upregulation of immune-suppressive pathways upon uptake by immune cells.8 This includes promoting the activation of regulatory T cells (Tregs) and the expression of immune checkpoint molecules, thereby weakening the host’s anti-tumor immune response.45,46 This EVs-mediated immune signal remodelling may accelerate immune escape during tumor progression. Similarly, exposure to gold nanoparticles (AuNPs) can induce cells to release EVs rich in HSP70 and pro-inflammatory factors.30 These EVs, when interacting with macrophages or dendritic cells, can activate inflammatory signaling pathways such as NF-κB, promoting the secretion of pro-inflammatory cytokines.47 In acute injury repair and infection defense, this inflammatory amplification effect may aid in lesion clearance and tissue regeneration, but in chronic inflammation or the tumor microenvironment, it may drive pathological inflammation and tumor progression.47
Additionally, NMs can indirectly regulate intercellular communication networks by influencing the miRNA load of Es.4 These EVs, upon being taken up by vascular smooth muscle cells, promote the expression of contractile phenotype markers and inhibit abnormal proliferation, thereby achieving the remodelling of vascular structure and function.48 This suggests that nanostructures can not only locally regulate the secretory behavior of a single cell type but also influence the functional state of adjacent or even distant tissues through EVs-mediated paracrine signals.49 Beyond oncology, the immunomodulatory consequences of NMs-EVs interactions are equally relevant in non-cancerous pathologies. For instance, in fibrotic diseases, exposure to silica nanoparticles or carbon nanotubes has been shown to induce epithelial cell-derived EVs enriched in pro-fibrotic mediators such as TGF-β1 and Wnt ligands, which activate fibroblasts and accelerate extracellular matrix deposition. Similarly, in the context of neuroinflammation, metal oxide nanoparticles can trigger microglial release of EVs carrying inflammasome components such as NLRP3 as well as IL-1β, and aggregation-prone proteins, thereby propagating neuroinflammatory signals and potentially exacerbating conditions such as Alzheimer’s or Parkinson’s disease. These examples underscore that the NMs-EVs regulatory axis operates across diverse pathological landscapes, influencing disease progression in fibrosis, neurodegeneration, and beyond.
In summary, NMs can alter the signal output patterns of EVs in intercellular communication by regulating their composition and physicochemical properties. This process involves the binding characteristics of EVs with receptor cell membranes, the selection of endocytic pathways, and the reconstruction of downstream transcriptional regulatory networks, ultimately determining the biological response of target cells.43 A deeper understanding of this process not only facilitates the targeted optimization of EVs’ therapeutic potential using NMs but also provides crucial evidence for assessing their risks in immune regulation, inflammatory responses, and tumor microenvironment remodelling.
Impact on Immune Modulation
Beyond regulating intercellular communication networks, EVs induced by NMs also exhibit significant functional shaping effects in the immune system.48 This process primarily occurs by altering the immune regulatory molecule profile carried by EVs, thereby influencing the activation state and response direction of immune cells.50 As mentioned earlier, nano-regulation can cause EVs to bias toward either immune suppression or inflammatory signals, and immune regulation is the specific manifestation of this bias at the tissue level. Multiple studies have shown that exposure to specific types of NMs can induce cells to secrete EVs rich in immune-suppressive molecules, including programmed death ligand 1 (PD-L1), transforming growth factor β (TGF-β), and interleukin 10 (IL-10). These molecules can exist in a stable form on the surface or within the lumen of EVs and, upon contact with immune cells, inhibit the activation of effector T cells through receptor-ligand interactions or endocytic delivery, thereby promoting the differentiation of regulatory T cells and overall downregulating the intensity of the immune response.48 In the tumor microenvironment, these immune-suppressive EVs aid cancer cells in evading immune surveillance.50 In transplant rejection or autoimmune disease models, the rational application of such EVs holds potential for mitigating excessive immune attacks.50 It is important to distinguish between the direct immunotoxicity of NMs and the EVs-mediated secondary immunomodulation discussed here. Direct immunotoxicity refers to the immediate effects of NMs on immune cells upon physical encounter, such as nanoparticle-induced NLRP3 inflammasome activation in macrophages or surface charge-dependent lymphocyte apoptosis. In contrast, EVs-mediated immunomodulation represents an indirect, signal-relayed mechanism: NMs first program donor cells to alter the molecular cargo of their secreted EVs, these EVs then transmit immunomodulatory signals to recipient immune cells, often at distant sites. This distinction is critical because EVs-mediated effects can outlast the initial exposure, propagate across tissues, and amplify the immunological impact of even low-dose NM exposure. By framing the discussion within this indirect pathway, this section focuses specifically on how NMs reprogram the immune system through EVs, rather than through direct nano-immune cell contact.
It is worth noting that the immune function of nano-regulated EVs is not unidirectionally inhibitory and may exhibit opposite effects under different immune contexts.48 For example, under conditions of infection or acute inflammation, some nano-induced EVs may enrich pro-inflammatory molecules and enhance immune responses, thereby accelerating pathogen clearance.51 In contrast, under conditions of chronic inflammation, transplant immune rejection, or autoimmune diseases, EVs rich in immune inhibitory molecules may become important tools for controlling immune overreactions.50 This context-dependent nature implies that when applying nanotechnology-regulated EVs for immune intervention, it is essential to precisely define the disease stage and immune status to optimize therapeutic efficacy and mitigate the occurrence of adverse effects (Figure 2).
 |
Figure 2 Bidirectional interactions between NMs and EVs. Upon entering the extracellular microenvironment, NMs rapidly acquire a protein Corona that modulates their biological identity and cellular interactions. After corona formation, NMs are internalized into cells primarily through clathrin-mediated endocytosis and other endocytic pathways (2–3). Inside the cytoplasm, NMs influence EVs biogenesis by modulating endosomal trafficking, ESCRT machinery activity, and MVB dynamics (3). These intracellular changes result in altered EVs cargo profiles, including upregulation of HSP70 (4). EVs subsequently mediate the redistribution of NMs to neighboring or distant cells, facilitating secondary delivery (5). The downstream biological consequences of NMs-EVs crosstalk include immune modulation, angiogenesis, and apoptosis. |
It is important to note that many studies infer immunomodulatory function from EVs molecular composition (eg, presence of PD-L1, TGF-β) rather than direct functional immunological assays. While correlational evidence is suggestive, definitive demonstration of immune modulation requires co-culture systems, primary immune cell functional assays (proliferation, cytokine secretion, cytotoxicity), and ideally in vivo validation. Future studies should prioritize functional immunological readouts to establish causal relationships between NMs-induced EVs cargo alterations and immune outcomes.
Impact on Angiogenesis
It is important to note that the majority of mechanistic insights into NMs-EVs interactions have emerged from in vitro studies, where controlled conditions allow for precise dissection of molecular events. In instances where they are available, the findings derived from in vivo models are highlighted. These models are advantageous in that they better recapitulate the complexity of physiological and pathological microenvironments, thereby underscoring their translational relevance. Angiogenesis plays a central role in tissue repair, regeneration, and tumor growth and metastasis.4 EVs can directly influence endothelial cell proliferation, migration, and lumen formation by transporting proangiogenic molecules and regulating signaling pathways.4 Under the regulation of NMs, the molecular composition and membrane properties of EVs often undergo changes, which can significantly affect the mode of action of EVs in the angiogenesis process, thereby exerting a bidirectional regulatory role in physiological and pathological environments (Figure 3).52,53
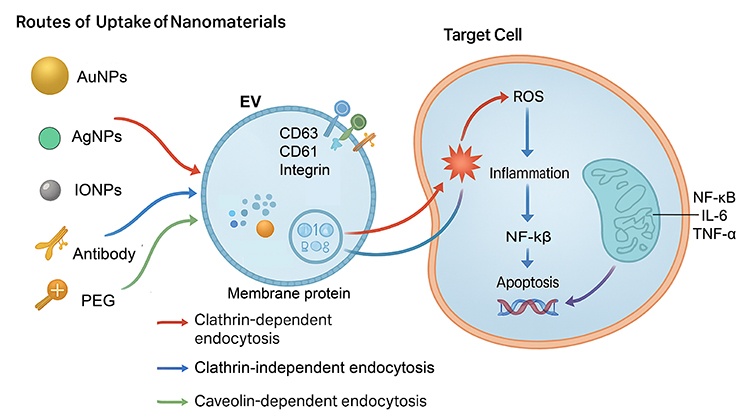 |
Figure 3 Routes of uptake of nanomaterials by EVs and their downstream effects on target cells. NMs with varying physicochemical properties, including Au NPs, Ag NPs, IO NPs, antibodies, and PEG-modified particles, can be incorporated into EVs via distinct endocytic pathways, such as clathrin-dependent endocytosis, clathrin-independent endocytosis, and caveolin-dependent endocytosis. Once loaded into EVs, these NMs can be delivered to target cells, where they induce oxidative stress through the generation of ROS, activate inflammatory signaling cascades (eg, NF-κB, IL-6, TNF-α), and trigger apoptosis, ultimately contributing to altered cellular functions or cell death. |
On the one hand, specific NMs can enhance the proangiogenic capacity of EVs. For example, tumor cells treated with AuNPs secrete EVs rich in vascular endothelial growth factor (VEGF), fibroblast growth factor 2 (FGF2), and proangiogenic-related miRNAs.54 These vesicles activate the PI3K/AKT and MAPK signaling pathways in endothelial cells, promoting cell proliferation and migration and accelerating the formation of new blood vessels.6 In the tumor microenvironment, this nanomaterial-induced EVs-mediated proangiogenic effect may accelerate the establishment of tumor blood supply networks, enhancing tumor invasiveness and metastatic potential.55 On the other hand, NMs can also inhibit angiogenesis by regulating the EVs cargo profile.55,56 Substrates with oriented nanofibre structures can alter the mechanical stress distribution on endothelial cells, leading to the secretion of EVs enriched in miR-143 and miR-145.53 These miRNAs can upregulate the expression of contractile phenotype genes in vascular smooth muscle cells, stabilize vascular wall structure, and to some extent restrict the formation of abnormal neovascularisation.4,52 These EVs can inhibit the activity of the VEGF signaling pathway in endothelial cells, reducing the formation of capillary-like structures.
The role of nano-regulated EVs in the vascularization process exhibits significant environmental dependence. In ischemic tissue repair models, pro-angiogenic EVs can accelerate local perfusion recovery and tissue regeneration.55,56 However, in tumor or pathological vascular proliferation environments, anti-angiogenic EVs may help disrupt blood supply to abnormal tissues, thereby limiting lesion progression.6
While multiple studies demonstrate that NMs-exposed cells secrete EVs with altered pro-angiogenic potential, much of this evidence remains correlational—linking miRNA enrichment to endothelial tube formation without definitive mechanistic validation. Causal demonstration requires loss-of-function studies (eg, miRNA inhibitors, blocking antibodies) and in vivo angiogenesis assays (Matrigel plugs, aortic ring assays) to confirm that observed effects are directly mediated by specific EVs components rather than indirect or confounding factors.
Cellular Uptake and Response of EVs to NMs
Having identified the primary pathways for NMs entering EVs, the subsequent question is how the presence of these nanoparticles within EVs will influence their subsequent biological effects. In fact, the uptake process not only determines the efficiency and manner of NMs entering EVs but also establishes the foundation for their interactions with the internal components of EVs. Whether through endocytosis or membrane fusion-mediated incorporation, NPs invariably alter the physicochemical properties and cargo state of EVs to some extent. This alteration can profoundly impact their signaling functions, in vivo distribution, and interaction patterns with target cells. Therefore, a thorough investigation of the biological effects arising from NMs-EVs interactions is crucial for understanding their dual roles in drug delivery, immunomodulation, and potential toxicity, while also providing a theoretical foundation for future applications.
As illustrated in Figure 2, nanomaterials engage in a bidirectional regulatory loop with EVs. After entering body fluids, NMs rapidly adsorb biomolecules to form a protein corona, which defines their cellular uptake properties and immunological visibility.23 Following internalization, predominantly through clathrin-mediated pathways, NMs localize to endosomal compartments and modulate key steps of EVs biogenesis, including ESCRT activity and MVB maturation.4,57 These processes collectively reshape the molecular cargo of secreted EVs, leading to the release of EVs enriched in stress proteins such as HSP70.58,59 In turn, EVs serve as vehicles that redistribute NMs to distal tissues, amplifying biological responses. Ultimately, this NMs-EVs interaction axis contributes to phenotype changes including immune modulation, angiogenic activation, and programmed cell death.6,60
Mechanisms of EVs Uptake of NMs
In the bidirectional relationship between NMs and EVs, in addition to the regulation of EVs biogenesis and function by NMs, EVs themselves can also take up and respond to exogenous NPs.4 This process not only affects the distribution and metabolism of NMs in the body, but may also alter the composition and function of EVs, thereby triggering a chain reaction of downstream biological effects.61 Before discussing the mechanisms, it is important to clarify the two distinct levels at which EVs can internalize nanomaterials. The first involves cellular uptake followed by EVs packaging: NMs are internalized by parental cells, processed through endosomal pathways, and subsequently incorporated into intraluminal vesicles during multivesicular body (MVB) formation, leading to their release within EVs. The second involves post-release uptake: once secreted, mature EVs can directly bind and internalize exogenous NMs from the extracellular milieu through endocytosis or membrane fusion. These two modes are not mutually exclusive and may coexist in biological systems, yet they differ fundamentally in their kinetics, molecular determinants, and functional consequences. The following section focuses primarily on the post-release uptake of NMs by isolated EVs, unless otherwise specified.
The uptake of NMs by EVs primarily relies on various endocytic and membrane interaction mechanisms.61 Among these, clathrin-dependent endocytosis is one of the most common pathways, suitable for NPs with smaller diameters and specific ligands on their surfaces.62 This endocytic pathway depends on the specific binding of receptors on the EVs membrane with molecules on the nanoparticle surface, triggering the formation and internalization of clathrin-coated pits. Caveolin-dependent endocytosis is typically associated with larger or hydrophobic-modified nanoparticles.63 This pathway utilizes cholesterol- and sphingomyelin-rich membrane domains for particle internalization and is commonly observed in EVs subpopulations involved in signal molecule transport.63 Additionally, macropinocytosis plays a crucial role in the uptake of larger-sized NPs by EVs.64 This process involves the formation of large vesicles through membrane invagination, which encapsulate exogenous particles and enter the cell.
In addition to the classic endocytosis mechanism, membrane fusion is another important pathway for EVs interaction with NMs.11,13 Some NPs coated with lipids or biomimetic membranes can directly fuse with EVs membranes, thereby integrating their cargo or surface molecules into the EVs structure.10 This process exhibits distinct temperature-dependent and membrane lipid fluidity characteristics, suggesting a potential association with the phospholipid composition and lipid raft structure of EVs membranes.7 EVs membrane proteins play a crucial role in the molecular recognition of nanomaterial uptake. For example, tetraspanins (CD9, CD63, CD81) and members of the integrin family can serve as molecular interfaces for the recognition and binding of NMs.65 Studies have shown that NPs designed with surface ligands targeting these membrane proteins can significantly enhance their binding efficiency and internalization rate with EVs, thereby enabling selective delivery or signal regulation.2 This property provides a theoretical foundation for using NMs to achieve targeted modification of EVs and creates conditions for the development of hybrid delivery systems.
Biological Effects of NMs-EVs Interactions
After being taken up by EVs, the presence of nanoparticles may not only alter the physicochemical properties of EVs but also exert profound effects on their biological functions and in vivo distribution patterns.17 Given that EVs inherently possess the ability to transport bioactive molecules across cell types, NPs incorporated into EVs can be transported to distant target cells or tissues, thereby altering the original delivery specificity and efficiency.48 Particularly in EVs subpopulations with tissue-tropic characteristics, the integration of NPs may enhance or reorient their organ distribution patterns, enabling the delivery of their cargo beyond their natural biological range. The classification of EVs into exosomes, microvesicles, and apoptotic bodies follows traditional nomenclature based on biogenesis pathways. However, according to MISEV2023 guidelines, definitive assignment to these subtypes requires rigorous experimental evidence, such as demonstrating endosomal origin for exosomes, which is not available in all cited studies.
NPs may also interfere with the normal signal transduction functions of EVs. Certain metal or oxide nanoparticles can directly bind to nucleic acids or proteins within the EVs lumen, thereby altering their conformation and activity and ultimately affecting signal transduction in downstream receptor cells.51 For example, ferrite nanoparticles can interact with iron-containing proteins, heat shock proteins, or enzymes in EVs, altering their stability and activity states.66 Additionally, the surfaces of NPs may adsorb signal molecules within EVs, causing some key factors to lose their activity during delivery or be taken up by non-specific target cells, thereby reducing the precision of the signal.67
When exposure to NPs is too high or accumulates over time, interactions between NMs and EVs may also trigger toxic effects and disrupt signal networks.10,66 Moreover, EVs containing high loads of NPs may trigger excessive oxidative stress and inflammatory responses when taken up by immune cells or endothelial cells, leading to cellular damage or even tissue dysfunction.67 On the other hand, NPs carried by EVs may continuously interfere with intercellular physiological signal transmission, disrupting microenvironmental homeostasis.7 It is worth noting that the impact of NPs on EVs function is highly dependent on various factors, including nanoparticle size, morphology, surface modification, chemical composition, as well as exposure dose and duration.7,14 Therefore, when designing EVs-based nano-delivery systems or strategies to modulate EVs function using NMs, it is essential to systematically evaluate these variables to ensure enhanced targeting and delivery efficiency while minimizing potential risks (Figure 3). Beyond the immediate immunomodulatory effects discussed above, the immunogenicity of NMs-EVs complexes raises critical safety considerations. First, the protein corona formed on NMs upon exposure to biological fluids can alter their immune recognition, corona composition shaped by NMs properties and the biological milieu-determines whether NMs are recognized as “self” or “non-self” by immune cells EVs, as carriers of NMs, may either shield or expose corona epitopes, thereby modulating subsequent immune responses. Second, repeated exposure to NM-EVs complexes carries the risk of inducing anti-drug antibodies (ADA) or hypersensitivity reactions, particularly if the complexes trigger B-cell activation or T-helper cell polarization. Third, the long-term persistence of EVs in circulation-coupled with their ability to deliver NMs to lymphoid tissues-raises concerns about chronic immune stimulation, potentially leading to autoimmunity or immune exhaustion These immunogenicity challenges underscore the need for comprehensive preclinical evaluation, including assessments of ADA formation, complement activation, and cytokine release syndrome, alongside standard toxicity studies.
Interestingly, EVs not only internalize NMs but also serve as natural shuttles redistributing nanoparticles across tissues.34,59 Recent research demonstrated that EVs can encapsulate surface-modified nanoparticles and transfer them to distal endothelial or parenchymal cells, bypassing classical clearance pathways.4 This EVs-mediated transport markedly alters nanoparticle bio-distribution, indicating that EVs form an endogenous secondary circulation system for nanomaterials.68 Beyond these functional alterations, the recognition that EVs can internalize and redistribute NMs introduces a conceptual paradigm shift: EVs function as an endogenous secondary circulation system for nanomaterials. This concept redefines the in vivo fate of NMs in several fundamental ways. First, it challenges the classical view that NM biodistribution is solely governed by their physicochemical properties and mononuclear phagocyte system clearance. Instead, EVs-mediated transport introduces a biological vector layer that can override synthetic identity, enabling NMs to access distal tissues and cellular niches beyond their natural reach. Second, this secondary circulation operates on different timescales-EVs can sequester, protect, and gradually release NMs, prolonging their biological half-life and creating a depot effect that extends therapeutic windows. Third, by exploiting the intrinsic tissue-tropism of EVs subpopulations, NMs can acquire hijacked targeting specificity, opening avenues for precision delivery without complex surface engineering. Collectively, framing EVs as a secondary circulatory system not only advances our understanding of nano-bio interactions but also provides a unified conceptual framework for designing next-generation nanotherapeutics-drugs that will actively harness rather than passively adapt to this endogenous redistribution pathway.
While the concept of EVs as an endogenous secondary circulation system for nanomaterials is intriguing, its quantitative contribution relative to classical biodistribution routes (mononuclear phagocyte system clearance, renal filtration, hepatic metabolism) remains undefined. Current evidence demonstrates that EVs-mediated transport can alter NMs distribution to specific tissues (eg, brain, tumors), but the fraction of total NM dose routed through this pathway, its kinetics, and its relative importance across different NMs types and administration routes have not been systematically quantified. Addressing this gap will require tracer studies, pharmacokinetic modeling, and comparative analyses in EVs-depleted vs. EVs-replete conditions.
In summary, the incorporation of NMs into EVs presents a dual-faced paradigm with both therapeutic opportunities and safety concerns. To enhance clarity, the key beneficial outcomes and potential risks are summarized below. Therapeutic Opportunities includes Enhanced biodistribution, EVs can redirect NMs to distal tissues, bypassing classical clearance pathways and enabling access to previously inaccessible sites. Prolonged circulation, EVs-mediated secondary circulation extends the biological half-life of NMs, creating a depot effect that widens therapeutic windows. Acquired targeting, Tissue-tropic EVs subpopulations can impart targeting specificity to NMs without complex surface engineering. Synergistic delivery, EVs and NMs can co-deliver therapeutic payloads, combining the biocompatibility of EVs with the functionality of NMs. Safety concerns involves signal disruption, NMs may bind to EVs cargo molecules, altering their conformation and interfering with normal intercellular signaling. Off-target effects, NMs-laden EVs may be taken up by non-target cells, reducing signaling precision and causing unintended biological effects. Toxic amplification, EVs loaded with high doses of NMs can trigger oxidative stress inflammatory responses, and cellular damage upon uptake by immune or endothelial cells. Microenvironment disturbance: Persistent EVs-mediated NMs signaling may disrupt tissue homeostasis and contribute to pathological remodeling. The balance between these beneficial and adverse outcomes is governed by multiple factors, including NMs physicochemical properties exposure dose, duration, and the biological context of EVs-cell interactions. Therefore, harnessing the NMs-EVs axis for therapeutic applications requires systematic optimization to maximize benefits while minimizing risks.
Therapeutic and Diagnostic Applications of NMs- EVs Interactions
In the preceding section, we focused on exploring the uptake mechanisms of EVs by nanomaterials and the resulting biological effects, including the remodeling of signaling pathways, alterations in distribution patterns, and potential toxicity risks. These insights not only reveal the complex bidirectional interactions between NMs and EVs but also provide a theoretical foundation for their potential value in medical applications. Indeed, it is precisely because EVs possess both intrinsic biocompatibility and programmability that their combination with NMs increasingly demonstrates unique advantages in disease treatment and diagnosis. Consequently, a growing number of studies are exploring how to harness this interaction to develop EVs into highly efficient drug delivery vehicles, sensitive biomarkers, and integrated diagnostic and therapeutic nanoplatforms.
EVs as Nanocarriers in Drug Delivery
EVs are considered ideal biopharmaceutical carriers due to their natural lipid bilayer structure derived from cell membranes, low immunogenicity, and excellent biocompatibility.69 Compared to traditional synthetic nanocarriers, EVs exhibit greater stability and lower clearance rates during circulation in vivo, and their membrane surface proteins and glycosylation modifications confer specific tissue and cell targeting capabilities.69 Additionally, EVs can cross various biological barriers, including the BBB, without triggering significant immune rejection, making them uniquely advantageous for the treatment of central nervous system (CNS) diseases.4 Compared to synthetic nanocarriers such as liposomes or polymeric nanoparticles, EVs offer distinct advantages derived from their biological origin: they exhibit lower immunogenicity due to self-derived membrane proteins, possess intrinsic tissue-targeting capabilities conferred by surface adhesion molecules, and can naturally traverse biological barriers like the BBB without the need for extensive surface engineering. These features position EVs as uniquely equipped vectors for applications where synthetic carriers often face limitations, such as CNS drug delivery and immune-evasive systemic circulation.
The introduction of NMs provides programmable enhancement strategies for the drug delivery function of EVs. Through surface functionalization or hybridization with NPs, the targeting ability, drug loading capacity, and tissue penetration capability of EVs can be significantly improved.6,70 For example, incorporating Au NPs, superparamagnetic IONPs, or quantum dots onto the surface of EVs not only enables visual tracking through their optical or magnetic response properties but also enhances affinity for specific receptor cells through ligand modification.6 Furthermore, NMs can co-load small-molecule drugs, nucleic acids, or proteins with EVs through electrostatic adsorption, hydrophobic interactions, or chemical coupling, thereby increasing drug loading capacity and improving release kinetics.29 Some strategies also utilise the photothermal or magnetothermal effects triggered by NMs to achieve exogenous, controllable drug release from EVs, thereby enhancing local therapeutic efficacy.2
In the treatment of CNS diseases, functionalized EVs can be loaded with neuroprotective agents, anti-inflammatory drugs, or nucleic acid therapeutic molecules.7 Through surface engineering of NMs, their efficiency in crossing the BBB can be enhanced, while reducing non-specific distribution in peripheral tissues. In tumor-targeted therapy, nano-functionalized EVs can deliver chemotherapeutic drugs, siRNA, or immunomodulators, significantly increasing drug accumulation at tumor sites while reducing systemic toxicity.7,17 Additionally, in anti-inflammatory drug delivery, nano-functionalized EVs can be designed to exhibit higher affinity in inflammatory microenvironments, enabling targeted delivery of glucocorticoids, non-steroidal anti-inflammatory drugs, or anti-inflammatory nucleic acid molecules to precisely regulate local immune responses.17,51
In summary, the advantages of EVs as natural drug carriers combined with the functionalization strategies of NMs not only significantly enhance the targeting, drug loading capacity and penetration of delivery systems but also provide feasible solutions for the precise treatment of various diseases.6,70 However, this strategy still faces numerous challenges in clinical translation, including the stability of large-scale production, batch-to-batch consistency, and long-term safety assessment. Therefore, future research should explore more efficient nanomaterial and EVs composite platforms while ensuring controllability and safety to advance their clinical application in CNS diseases, tumor therapy, and inflammatory diseases.
EVs as Diagnostic Biomarkers Upon Nanomaterial Exposure
The exposure of NMs within biological organisms may not only affect cellular function but also significantly alter the molecular composition of EVs, including key components such as proteins, miRNAs, and lipids.5,50 These changes can reflect the stress state of cells under nanostimulation, the activation patterns of signaling pathways, and potential pathological processes, thereby demonstrating potential as exposure biomarkers and tools for early disease diagnosis.2 Existing studies have shown that exposure to specific metal or oxide nanoparticles can induce characteristic changes in the protein profile of EVs. For example, tumor cells exposed to superparamagnetic IONPs released EVs with significantly elevated levels of the immune regulatory protein PD-L1,71 the inflammation-associated molecule HMGB1,72 and the heat shock protein HSP70.73 Therefore, these EVs-derived molecular features not only serve as biological fingerprints of nanomaterial exposure but may also indicate associated disease risks. It is important to contextualize the translational readiness of NMs-induced EVs biomarkers, which span a spectrum from experimental discovery to early clinical validation. While the concept is supported by compelling preclinical evidence, most of the examples discussed below remain at the experimental or preclinically validated stage, with clinical adoption requiring further validation in human cohorts and standardization of detection platforms.
In the field of early tumor diagnosis, changes in the molecular profile of EVs induced by exposure to NMs hold promise for the development of liquid biopsy platforms.2 For example, EVs rich in proangiogenic factors or specific miRNAs may indicate the early onset of tumor microenvironment remodelling.27 In neurological diseases, EVs released by NMs after crossing the BBB and interacting with neurons or glial cells may carry molecular markers associated with neuroinflammation, protein misfolding, or synaptic damage, such as neurofilament protein (NfL) or β-amyloid, providing feasible clues for the early diagnosis of Alzheimer’s disease, Parkinson’s disease, and other conditions.74,75 In cardiovascular injury detection, cardiomyocytes exposed to specific NMs can secrete EVs rich in cardiac injury markers, which can be detected through blood tests to achieve non-invasive, early identification of cardiac injury.76
Compared with traditional biomarkers, EVs-derived biomarkers exhibit high stability in various body fluids such as blood, urine, and saliva, and can reflect the molecular characteristics of the lesion site.6 Furthermore, by combining signal amplification and detection platforms with NMs, high-sensitivity detection of EVs biomarkers can be achieved in minute samples, thereby significantly improving the accuracy and accessibility of early diagnosis.
Theranostic Platforms Combining NMs and EVs
A theranostic platform integrating NMs with EVs has been developed to leverage the biocompatibility and targeting properties of EVs, combined with the imaging, drug delivery, and stimulus-responsive characteristics of NMs, to achieve simultaneous diagnosis and treatment of diseases.77 This strategy integrates diagnostic signal probes and therapeutic payloads within the same carrier system, enabling real-time monitoring of drug delivery and treatment responses. Additionally, treatment dosage and timing can be adjusted based on imaging feedback, thereby enhancing therapeutic efficacy and reducing unnecessary systemic exposure.6,7 Table 1 summarizes the characteristics, mechanisms, and applications of representative NMs on EVs biology.2,7 Currently, superparamagnetic IO NPs, Au NPs, and quantum dots are widely used for EVs functionalization to confer visualization capabilities such as MRI, PAI, and SERS.78,79 On the other hand, diagnostic and therapeutic integrated platforms can co-load chemotherapy drugs, siRNA, miRNA, proteins, or immunomodulators with nanoimaging probes into EVs.78,79 By leveraging the stimulus-responsive properties of NMs, controlled drug release can be achieved, enabling high drug concentrations at disease sites while reducing systemic toxicity.12 For example, by using near-infrared light to stimulate Au NPs-EVs complexes, not only can the tumor cell membrane be destroyed through photothermal effects, but the drugs inside the EVs can also be triggered to release, thereby forming a dual physical and chemical killing effect.80
 |
Table 1 Representative NMs Influencing EVs Biology |
In CNS diseases, the potential for integrated diagnosis and treatment using EVs and nanomaterial platforms is particularly prominent.12,79 Functionalized EVs can cross the BBB to deliver neuroprotective agents or anti-inflammatory drugs to the site of neural injury, while fluorescent or MRI probes enable real-time monitoring of drug distribution and action within the brain. In cardiovascular diseases, functionalized EVs can target ischemic myocardial regions to deliver vasculogenic factors and monitor improvements in local perfusion, providing dynamic imaging evidence for regenerative therapies.78
The advantage of this integrated diagnosis and treatment model lies in its ability to integrate multiple functions into a single platform, thereby simplifying treatment processes, improving treatment efficiency, and enabling personalized interventions. However, its clinical translation still faces several challenges, including the stability and consistency of NMs and EVs complexes in large-scale preparation, the optimization of the balance between imaging and therapeutic functions, and the assessment of long-term safety and immune response risks.
Challenges and Future Directions
Although significant progress has been made in the study of interactions between NMs and EVs in terms of mechanism elucidation and application exploration, several key challenges still hinder their clinical translation.9 First, EVs themselves exhibit high heterogeneity, which is not only reflected in the diversity of particle size distribution and biogenic pathways but also manifested in differences in cargo composition, membrane protein profiles, and tissue sources.4 This heterogeneity complicates the results of nanomaterial and EVs interactions and introduces unpredictability, while also posing technical challenges for separation and purification. Existing separation methods struggle to balance efficiency, purity, and reproducibility, particularly when large-scale, high-consistency production is required for clinical applications.7 Obtaining batch-stable EVs subpopulations with well-defined functional properties remains an urgent challenge that needs to be addressed. Moreover, many mechanistic insights remain derived from specific cell lines under controlled conditions, and their generalizability across cell types and physiological contexts requires further validation.
Methodological challenges further complicate data interpretation. EVs isolation methods (ultracentrifugation, size exclusion chromatography, precipitation) introduce distinct biases in EVs subpopulation recovery, potentially confounding comparisons across studies. Similarly, nanoparticle aggregation in biological media can alter effective size and surface properties, leading to variable cellular interactions that are often unaccounted for in experimental designs. Future studies should report detailed characterization of both EVs isolation efficiency and NMs colloidal stability to enable cross-study comparability.
Secondly, the toxicological assessment standards for NMs are still unclear, and this is particularly evident in composite platforms that incorporate EVs.84 Nanotoxicological implications of NMs-EVs interactions have not been integrated into existing risk-assessment frameworks. Conventional nanotoxicology appraises the direct effects of NMs on exposed cells/tissues. However, EVs-mediated indirect effects, such as the propagation of stress signals to distant sites, the amplification of inflammatory responses, and the long-term reprogramming of tissue microenvironments, represent secondary toxicity mechanisms that are not encompassed by current regulatory protocols. We propose that EVs parameters (secretion rates, cargo profiles, functional activity in recipient cells) should be incorporated into next-generation nanotoxicity screening platforms as sensitive indicators of biological perturbation.3
Third, the dynamic processes of interactions between NMs and EVs involve multi-scale, multi-level biophysical and biochemical events, including the formation of protein coronas around NPs in body fluids, initial contact with cell membranes, endocytosis and membrane fusion mechanisms, intracellular transport and processing, and redistribution via EVs.77 Single experimental systems are insufficient to fully elucidate these processes. Therefore, future research should integrate multi-scale collaborative research methods, including high-resolution imaging, multi-omics analysis, and computational modelling, to establish spatiotemporal dynamic models of nanomaterial and EVs interactions. This will facilitate the prediction of biological outcomes under different nanomaterial parameters and guide the iterative optimization of platform design. Beyond the scientific and technical hurdles discussed above, the clinical translation of NMs-EVs hybrid platforms faces regulatory and safety assessment challenges that are fundamental to their approval and commercialization. First, batch-to-batch variability remains a critical concern. EVs isolated from cell cultures exhibit intrinsic heterogeneity in size, cargo composition, and membrane protein profiles, which can be further amplified by NM loading or surface modification. This variability complicates the establishment of consistent quality control metrics and poses challenges for reproducibility across production batches Second, EVs source control is a regulatory prerequisite. Depending on the intended application, EVs may be derived from various cell types, each with distinct safety profiles and regulatory pathways. Autologous versus allogeneic sources, donor variability, and the risk of tumorigenic potential must be rigorously evaluated Third, Good Manufacturing Practice (GMP) considerations introduce additional layers of complexity. Unlike synthetic nanocarriers, which benefit from well-established manufacturing workflows, EVs-based products require GMP-compliant cell culture systems, scalable isolation methods, and validated characterization assays to ensure product identity, purity, potency, and sterility. The absence of standardized regulatory frameworks specifically tailored to NMs-EVs hybrids further compounds these challenges, as existing guidelines for nanomedicines and cell-derived products may not fully capture the unique features of these combinatorial platforms.
Finally, multifunctional platforms combining NMs and EVs hold great potential in precision medicine, particularly in achieving integrated diagnosis and treatment, overcoming biological barriers, and enabling personalized disease intervention.14,85 By integrating imaging probes, therapeutic payloads, and targeting ligands into EVs-based nanocomposite systems, real-time disease monitoring and precise intervention can be achieved. However, to achieve this goal, breakthroughs are needed in material controllability, standardized preparation, scalable production, and long-term safety, along with the accumulation of sufficient data support from preclinical and clinical studies. To address these regulatory and safety challenges, future efforts should prioritize the development of standardized protocols and reference materials for NMs-EVs hybrid production and characterization. Establishing industry-wide consensus on critical quality attributes (CQAs) such as EVs particle concentration, NMs loading efficiency, surface marker profiles, and functional potency-will be essential for regulatory acceptance. Furthermore, adaptive regulatory pathways that allow for iterative refinement as understanding of NMs-EVs interactions evolves could accelerate clinical translation while maintaining safety oversight. Collaboration between academia, industry, and regulatory agencies (eg, FDA, EMA) is urgently needed to establish fit-for-purpose guidelines that accommodate the unique biological characteristics and manufacturing complexities of NMs-EVs platforms. Ultimately, the successful clinical translation of NMs-EVs platforms will depend not only on scientific innovation but also on regulatory readiness and manufacturing robustness. Addressing batch variability, source control, and GMP compliance from the earliest stages of development will be critical to navigating the path from bench to bedside.
Conclusion
The interaction between NMs and EVs represents an important intersection between nanobiology and intercellular communication research. This interplay provides new opportunities to understand the biological fate of exogenous nanostructures in vivo and to develop innovative diagnostic and therapeutic platforms. Increasing evidence indicates that NMs can modulate EVs biogenesis, cargo composition, and membrane properties, thereby reshaping EVs-mediated signaling pathways involved in immune regulation, angiogenesis, and tissue remodelling. Conversely, EVs can function as natural carriers capable of internalization and transporting nanoparticles, enabling targeted delivery and functional enhancement and providing a structural foundation for multifunctional nanomedicine platforms.
In therapeutic applications, nano-functionalized EVs exhibit enhanced targeting capability, improved drug-loading efficiency, and an increased capacity to overcome biological barriers, demonstrating potential in the treatment of CNS disorders, precision oncology, and anti-inflammatory therapies. From a diagnostic perspective, NMs-induced alterations in EVs molecular profiles may serve as sensitive biomarkers for exposure assessment and early disease detection. When integrated with advanced nanotechnology detection technologies, these signatures may support the development of highly sensitive, multi-parameter liquid biopsy systems. In addition, emerging theranostic platforms integrating diagnostic monitoring and therapeutic intervention offer promising strategies for precision medicine (Figure 4).
 |
Figure 4 Schematic overview of the NMs-EVs regulatory axis: mechanisms, applications, and challenges. |
Despite these advances, several challenges remain. Beyond the scientific and technical hurdles discussed above, the clinical translation of NMs-EVs hybrid platforms faces regulatory and safety assessment challenges that are fundamental to their approval and commercialization. Batch-to-batch variability, EVs source control, and Good Manufacturing Practice (GMP) compliance introduce additional layers of complexity. The absence of standardized regulatory frameworks specifically tailored to NMs-EVs hybrids further compounds these challenges, as existing guidelines for nanomedicines and cell-derived products may not fully capture the unique features of these combinatorial platforms. The intrinsic heterogeneity of EVs populations, difficulties in their isolation and purification, and the lack of standardized toxicological evaluation frameworks for NMs continue to limit clinical translation. Future research should integrate multi-scale imaging, multi-omics approaches, and computational modelling to better define the dynamic interactions between NMs and EVs. On this basis, platform design, safety assessment, and manufacturing processes can be further optimized. Establishing standardized preparation protocols, robust quality control criteria, and clinical validation strategies will be essential for translating nano-EVs systems from laboratory studies to clinical practice.
Looking forward, advances in NMs-EVs biology may reshape the landscape of precision medicine. By elucidating how nanomaterials regulate EVs secretion, cargo programming, and biodistribution, this research field could enable next-generation liquid biopsy technologies with improved sensitivity and molecular specificity. Likewise, hybrid NMs-EVs delivery systems may offer unprecedented control over targeting accuracy, immune compatibility, and therapeutic payload design, positioning the NMs-EVs axis as a foundational framework for future diagnostic nanotechnology and personalized nanotherapeutics.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
This study was supported by Sichuan Science and Technology Program (2025ZNSFSC0544), National Natural Science Foundation project (82372825), and Luzhou Science and Technology Program (2023JYJ028, 2024JYJ053).
Disclosure
The authors declare no competing interests.
References
1. Man K, Brunet MY, Jones MC, Cox SC. Engineered extracellular vesicles: tailored-made nanomaterials for medical applications. Nanomaterials. 2020;10(9):1838. doi:10.3390/nano10091838
2. Zhang Y, Zhao L, Li Y, et al. Advanced extracellular vesicle bioinformatic nanomaterials: from enrichment, decoding to clinical diagnostics. J Nanobiotechnol. 2023;21(1):366. doi:10.1186/s12951-023-02127-3
3. Phillips W, Willms E, Hill AF. Understanding extracellular vesicle and nanoparticle heterogeneity: novel methods and considerations. Proteomics. 2021;21(13–14):e2000118. doi:10.1002/pmic.202000118
4. Lima TSM, Souza W, Geaquinto LRO, et al. Nanomaterial exposure, extracellular vesicle biogenesis and adverse cellular outcomes: a scoping review. Nanomaterials. 2022;12(7):1231. doi:10.3390/nano12071231
5. Jiang H, Kumarasamy RV, Pei J, et al. Integrating engineered nanomaterials with extracellular vesicles: advancing targeted drug delivery and biomedical applications. Front Nanotechnol. 2025;6. doi:10.3389/fnano.2024.1513683
6. Zocchi MR, Tosetti F, Benelli R, Poggi A. Cancer nanomedicine special issue review anticancer drug delivery with nanoparticles: extracellular vesicles or synthetic nanobeads as therapeutic tools for conventional treatment or immunotherapy. Cancers. 2020;12(7):1886. doi:10.3390/cancers12071886
7. Gurunathan S, Kang MH, Qasim M, Khan K, Kim JH. Biogenesis, membrane trafficking, functions, and next generation nanotherapeutics medicine of extracellular vesicles. Int J Nanomed. 2021;16:3357–21. doi:10.2147/IJN.S310357
8. Chen Y, Hou S. Recent progress in the effect of magnetic iron oxide nanoparticles on cells and extracellular vesicles. Cell Death Discov. 2023;9(1):195. doi:10.1038/s41420-023-01490-2
9. Taylor ML, Giacalone AG, Amrhein KD, Wilson RE Jr, Wang Y, Huang X. Nanomaterials for molecular detection and analysis of extracellular vesicles. Nanomaterials. 2023;13(3):524. doi:10.3390/nano13030524
10. Busatto S, Pham A, Suh A, Shapiro S, Wolfram J. Organotropic drug delivery: synthetic nanoparticles and extracellular vesicles. Biomed Microdevices. 2019;21(2):46. doi:10.1007/s10544-019-0396-7
11. Witwer KW, Wolfram J. Extracellular vesicles versus synthetic nanoparticles for drug delivery. Nat Rev Mater. 2021;6(2):103–106. doi:10.1038/s41578-020-00277-6
12. Choi HK, Chen M, Goldston LL, Lee KB. Extracellular vesicles as nanotheranostic platforms for targeted neurological disorder interventions. Nano Converg. 2024;11(1):19. doi:10.1186/s40580-024-00426-5
13. Jeppesen DK, Zhang Q, Franklin JL, Coffey RJ. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol. 2023;33(8):667–681. doi:10.1016/j.tcb.2023.01.002
14. Man K, Eisenstein NM, Hoey DA, Cox SC. Bioengineering extracellular vesicles: smart nanomaterials for bone regeneration. J Nanobiotechnol. 2023;21(1):137. doi:10.1186/s12951-023-01895-2
15. Hoshino A, Kim HS, Bojmar L, et al. Extracellular vesicle and particle biomarkers define multiple human cancers. Cell. 2020;182(4):1044–1061.e1018. doi:10.1016/j.cell.2020.07.009
16. Cecchin R, Troyer Z, Witwer K, Morris KV. Extracellular vesicles: the next generation in gene therapy delivery. Mol Ther. 2023;31(5):1225–1230. doi:10.1016/j.ymthe.2023.01.021
17. Yáñez-Mó M, Siljander PRM, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4(1):27066. doi:10.3402/jev.v4.27066
18. Manno M, Bongiovanni A, Margolis L, Bergese P, Arosio P. The physico-chemical landscape of extracellular vesicles. Nature Rev Bioeng. 2025;3(1):68–82. doi:10.1038/s44222-024-00255-5
19. Gurunathan S, Kang M-H, Kim J-H. A comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int J Nanomed. 2021;16:1281–1312. doi:10.2147/IJN.S291956
20. Essa S, Rabanel JM, Hildgen P. Effect of polyethylene glycol (PEG) chain organization on the physicochemical properties of poly (D, L-lactide)(PLA) based nanoparticles. Eur J Pharm Biopharm. 2010;75(2):96–106. doi:10.1016/j.ejpb.2010.03.002
21. Sun C, Du K, Fang C, et al. PEG-mediated synthesis of highly dispersive multifunctional superparamagnetic nanoparticles: their physicochemical properties and function in vivo. ACS nano. 2010;4(4):2402–2410. doi:10.1021/nn100190v
22. Zhu M, Nie G, Meng H, Xia T, Nel A, Zhao Y. Physicochemical properties determine nanomaterial cellular uptake, transport, and fate. Acc Chem Res. 2013;46(3):622–631. doi:10.1021/ar300031y
23. Park SJ. Protein-nanoparticle interaction: corona formation and conformational changes in proteins on nanoparticles. Int J Nanomed. 2020;15:5783–5802. doi:10.2147/IJN.S254808
24. Han S, da Costa Marques R, Simon J, et al. Endosomal sorting results in a selective separation of the protein Corona from nanoparticles. Nat Commun. 2023;14(1):295. doi:10.1038/s41467-023-35902-9
25. Sivanantham A, Jin Y. Impact of storage conditions on EV integrity/surface markers and cargos. Life. 2022;12(5):697. doi:10.3390/life12050697
26. Dixson AC, Dawson TR, Di Vizio D, Weaver AM. Context-specific regulation of extracellular vesicle biogenesis and cargo selection. Nat Rev Mol Cell Biol. 2023;24(7):454–476. doi:10.1038/s41580-023-00576-0
27. Ding Y, Hu Q, Gan J, et al. Effect of miR-143-3p from extracellular vesicles of porcine uterine luminal fluid on porcine trophoblast cells. Animals. 2022;12(23):3402. doi:10.3390/ani12233402
28. Xu Y, Qin S, An T, Tang Y, Huang Y, Zheng L. MiR‐145 detection in urinary extracellular vesicles increase diagnostic efficiency of prostate cancer based on hydrostatic filtration dialysis method. Prostate. 2017;77(10):1167–1175. doi:10.1002/pros.23376
29. Bhavsar V, Sahu A, Taware R. Stress-induced extracellular vesicles: insight into their altered proteomic composition and probable physiological role in cancer. Mol Cell Biochem. 2025;480(4):2025–2041. doi:10.1007/s11010-024-05121-x
30. Ullah M, Liu DD, Rai S, Concepcion W, Thakor AS. HSP70-mediated NLRP3 inflammasome suppression underlies reversal of acute kidney injury following extracellular vesicle and focused ultrasound combination therapy. Int J Mol Sci. 2020;21(11):4085. doi:10.3390/ijms21114085
31. Li Y, Cao H, Qiu D, et al. The proteomics analysis of extracellular vesicles revealed the possible function of heat shock protein 60 in Helicobacter pylori infection. Can Cell Inter. 2023;23(1):272. doi:10.1186/s12935-023-03131-1
32. Mir B, Goettsch C. Extracellular vesicles as delivery vehicles of specific cellular cargo. Cells. 2020;9(7):1601. doi:10.3390/cells9071601
33. Gaurav I, Thakur A, Kumar G, et al. Delivery of apoplastic extracellular vesicles encapsulating green-synthesized silver nanoparticles to treat citrus canker. Nanomaterials. 2023;13(8):1306. doi:10.3390/nano13081306
34. Wu X, Tang T, Wei Y, Cummins KA, Wood DK, Pang HB. Extracellular vesicles mediate the intercellular exchange of nanoparticles. Adv Sci. 2022;9(7):2102441. doi:10.1002/advs.202102441
35. Cui G, Li Z, Cao F, et al. Activation of Nrf2/HO-1 signaling pathway attenuates ROS-mediated autophagy induced by silica nanoparticles in H9c2 cells. Environ Toxicol. 2021;36(7):1389–1401. doi:10.1002/tox.23134
36. Ruan L, Li H, Zhang J, et al. Chemical transformation and cytotoxicity of iron oxide nanoparticles (IONPs) accumulated in mitochondria. Talanta. 2023;251:123770. doi:10.1016/j.talanta.2022.123770
37. Martinez-Arroyo O, Flores-Chova A, Mendez-Debaets M, et al. Enrichment of REDOXIFIBROMIR miR-21-5p in plasma exosomes of hypertensive patients with renal injury. Int J Mol Sci. 2025;26(2):590. doi:10.3390/ijms26020590
38. Li JW, Wei L, Han Z, Chen Z. Mesenchymal stromal cells-derived exosomes alleviate ischemia/reperfusion injury in mouse lung by transporting anti-apoptotic miR-21-5p. Eur J Pharmacol. 2019;852:68–76. doi:10.1016/j.ejphar.2019.01.022
39. Fulzele S, Mendhe B, Khayrullin A, et al. Muscle-derived miR-34a increases with age in circulating extracellular vesicles and induces senescence of bone marrow stem cells. Aging. 2019;11(6):1791–1803. doi:10.18632/aging.101874
40. Pink M, Verma N, Schmitz-Spanke S. Benchmark dose analyses of toxic endpoints in lung cells provide sensitivity and toxicity ranking across metal oxide nanoparticles and give insights into the mode of action. Toxicol Lett. 2020;331:218–226. doi:10.1016/j.toxlet.2020.06.012
41. Buttiens K, Maksoudian C, Perez Gilabert I, et al. Inorganic nanoparticles change cancer-cell-derived extracellular vesicle secretion levels and cargo composition, resulting in secondary biological effects. ACS Appl Mater Interfaces. 2024;16(1):66–83. doi:10.1021/acsami.3c12680
42. Crewe C, Funcke JB, Li S, et al. Extracellular vesicle-based interorgan transport of mitochondria from energetically stressed adipocytes. Cell Metab. 2021;33(9):1853–1868.e1811. doi:10.1016/j.cmet.2021.08.002
43. Van niel G, Carter DR, Clayton A, Lambert DW, Raposo G, Vader P. Challenges and directions in studying cell–cell communication by extracellular vesicles. Nat Rev Mol Cell Biol. 2022;23(5):369–382. doi:10.1038/s41580-022-00460-3
44. Zieske JD, Hutcheon AE, Guo X. Extracellular vesicles and cell–cell communication in the cornea. Anat Record. 2020;303(6):1727–1734. doi:10.1002/ar.24181
45. Wang S, Mao Y, Rong S, et al. Engineering magnetic extracellular vesicles mimetics for enhanced targeting chemodynamic therapy to overcome ovary cancer. ACS Appl Mater Interfaces. 2024;16(30):39021–39034. doi:10.1021/acsami.4c06862
46. Mehta KJ. Iron-oxide-nanoparticle-tagged mesenchymal stem cell exosomes in therapeutics. Curr Stem Cell Rep. 2024;11(1):1. doi:10.1007/s40778-024-00242-x
47. Buzas EI. The roles of extracellular vesicles in the immune system. Nat Rev Immunol. 2023;23(4):236–250. doi:10.1038/s41577-022-00763-8
48. Kalluri R. The biology and function of extracellular vesicles in immune response and immunity. Immunity. 2024;57(8):1752–1768. doi:10.1016/j.immuni.2024.07.009
49. Schatz D, Vardi A. Extracellular vesicles—new players in cell–cell communication in aquatic environments. Curr Opin Microbiol. 2018;43:148–154. doi:10.1016/j.mib.2018.01.014
50. Zhou X, Xie F, Wang L, et al. The function and clinical application of extracellular vesicles in innate immune regulation. Cell Mol Immunol. 2020;17(4):323–334. doi:10.1038/s41423-020-0391-1
51. Wen C, Seeger RC, Fabbri M, Wang L, Wayne AS, Jong AY. Biological roles and potential applications of immune cell-derived extracellular vesicles. J Extracell Vesicles. 2017;6(1):1400370. doi:10.1080/20013078.2017.1400370
52. Giusti I, Delle Monache S, Di Francesco M, et al. From glioblastoma to endothelial cells through extracellular vesicles: messages for angiogenesis. Tumor Biol. 2016;37(9):12743–12753. doi:10.1007/s13277-016-5165-0
53. Ye ZW, Yu ZL, Chen G, Jia J. Extracellular vesicles in tumor angiogenesis and resistance to anti‐angiogenic therapy. Cancer Sci. 2023;114(7):2739–2749. doi:10.1111/cas.15801
54. Feng Q, Zhang C, Lum D, et al. A class of extracellular vesicles from breast cancer cells activates VEGF receptors and tumour angiogenesis. Nat Commun. 2017;8(1):14450. doi:10.1038/ncomms14450
55. Todorova D, Simoncini S, Lacroix R, Sabatier F, Dignat-George F. Extracellular vesicles in angiogenesis. Circulat Res. 2017;120(10):1658–1673. doi:10.1161/CIRCRESAHA.117.309681
56. Kholia S, Ranghino A, Garnieri P, et al. Extracellular vesicles as new players in angiogenesis. Vasc Pharmacol. 2016;86:64–70. doi:10.1016/j.vph.2016.03.005
57. Lee YJ, Shin KJ, Chae YC. Regulation of cargo selection in exosome biogenesis and its biomedical applications in cancer. Exp Mol Med. 2024;56(4):877–889. doi:10.1038/s12276-024-01209-y
58. Linder M, Pogge von Strandmann E. The role of extracellular HSP70 in the function of tumor-associated immune cells. Cancers. 2021;13(18):4721. doi:10.3390/cancers13184721
59. Adamiak K, Sidoryk-Węgrzynowicz M, Dąbrowska-Bouta B, Sulkowski G, Strużyńska L. Plastic nanoparticles interfere with extracellular vesicle pathway in primary astrocytes. Ecotoxicol Environ Saf. 2024;286:117180.
60. Abebaw D, Akelew Y, Adugna A, et al. Extracellular vesicles: immunomodulation, diagnosis, and promising therapeutic roles for rheumatoid arthritis. Front Immunol. 2024;15:1499929. doi:10.3389/fimmu.2024.1499929
61. Wang X, Wang WX. Cellular journey of nanomaterials: theories, trafficking, and kinetics. Aggregate. 2023;4(6):e372. doi:10.1002/agt2.372
62. Ye C, Hu X-S, Wang Z-W, et al. Involvement of clathrin-dependent endocytosis in cellular dsRNA uptake in aphids. Insect Biochem Mol Biol. 2021;132:103557. doi:10.1016/j.ibmb.2021.103557
63. Wang H, Shan X, Ren M, Shang M, Zhou C. Nucleosomes enter cells by clathrin-and caveolin-dependent endocytosis. Nucleic Acids Res. 2021;49(21):12306–12319. doi:10.1093/nar/gkab1121
64. Salloum G, Bresnick AR, Backer JM. Macropinocytosis: mechanisms and regulation. Biochem J. 2023;480(5):335–362. doi:10.1042/BCJ20210584
65. Kowalczyk A, Gajda-Walczak A, Ruzycka-Ayoush M, et al. Parallel SPR and QCM-D quantitative analysis of CD9, CD63, and CD81 tetraspanins: a simple and sensitive way to determine the concentration of extracellular vesicles isolated from human lung cancer cells. Anal Chem. 2023;95(25):9520–9530. doi:10.1021/acs.analchem.3c00772
66. Fernández-Afonso Y, Asín L, Beola L, et al. Iron speciation in animal tissues using AC magnetic susceptibility measurements: quantification of magnetic nanoparticles, ferritin, and other iron-containing species. ACS Appl Bio Mater. 2022;5(5):1879–1889. doi:10.1021/acsabm.1c01200
67. Özçelik S. Copper ferrite nanoparticles: structural, magnetic, optical, photocatalytic activity and blood compatibility properties. BioNanoScience. 2023;13(3):958–972. doi:10.1007/s12668-023-01130-0
68. Gupta D, Wiklander OPB, Wood MJA, El-Andaloussi S. Biodistribution of therapeutic extracellular vesicles. Extracell Vesicles Circ Nucl Acids. 2023;4(2):170–190. doi:10.20517/evcna.2023.12
69. Osorio-Méndez JJ, Gómez-Grosso LA, Montoya-Ortiz G, Novoa-Herrán S, Domínguez-Romero Y. Small Extracellular Vesicles from Breast Cancer Cells Induce Cardiotoxicity. Int J Mol Sci. 2025;26(3):945. doi:10.3390/ijms26030945
70. Badila E, Japie C, Vrabie A-M, Badila A, Georgescu A. Cardiovascular disease as a consequence or a cause of cancer: potential role of extracellular vesicles. Biomolecules. 2023;13(2):321. doi:10.3390/biom13020321
71. Gao Y, Chen X, Wang B, et al. Engineering platelets with PDL1 antibodies and iron oxide nanoparticles for postsurgical cancer immunotherapy. ACS Appl Bio Mater. 2022;6(1):257–266. doi:10.1021/acsabm.2c00869
72. Dai Z, Tang J, Gu Z, et al. Eliciting immunogenic cell death via a unitized nanoinducer. Nano Lett. 2020;20(9):6246–6254. doi:10.1021/acs.nanolett.0c00713
73. Basaki M, Keykavusi K, Sahraiy N, Akbari G, Hejazi M. Small heat shock protein’s gene expression response to iron oxide nanoparticles in the brain. Biol Trace Elem Res. 2022;200(4):1791–1798. doi:10.1007/s12011-021-02761-8
74. Caruso Bavisotto C, Scalia F, Marino Gammazza A, et al. Extracellular vesicle-mediated cell–cell communication in the nervous system: focus on neurological diseases. Int J Mol Sci. 2019;20(2):434. doi:10.3390/ijms20020434
75. Jin Q, Wu P, Zhou X, Qian H, Xu W, Alzahrani F. Extracellular vesicles: novel roles in neurological disorders. Stem Cells Int. 2021;2021(1):6640836. doi:10.1155/2021/6640836
76. Fu S, Zhang Y, Li Y, Luo L, Zhao Y, Yao Y. Extracellular vesicles in cardiovascular diseases. Cell Death Discovery. 2020;6(1):68. doi:10.1038/s41420-020-00305-y
77. Wu Q, Fu S, Xiao H, et al. Advances in extracellular vesicle nanotechnology for precision theranostics. Adv Sci. 2023;10(3):2204814. doi:10.1002/advs.202204814
78. Bei Y, Das S, Rodosthenous RS, et al. Extracellular vesicles in cardiovascular theranostics. Theranostics. 2017;7(17):4168. doi:10.7150/thno.21274
79. Rakshit T, Pal S. Extracellular vesicles for drug delivery and theranostics in vivo. JACS Au. 2024;4(2):318–327. doi:10.1021/jacsau.3c00611
80. Ailuno G, Baldassari S, Lai F, Florio T, Caviglioli G. Exosomes and extracellular vesicles as emerging theranostic platforms in cancer research. Cells. 2020;9(12):2569. doi:10.3390/cells9122569
81. de Souza W, Gemini-Piperni S, Grenho L, et al. Titanium dioxide nanoparticles affect osteoblast-derived exosome cargos and impair osteogenic differentiation of human mesenchymal stem cells. Biomater Sci. 2023;11(7):2427–2444. doi:10.1039/D2BM01854C
82. Mu X, Hu K, Wei A, Bai J, Feng L, Jiang J. TiO2 nanoparticles promote tumor metastasis by eliciting pro-metastatic extracellular vesicles. J Nanobiotechnol. 2023;21(1):392. doi:10.1186/s12951-023-02142-4
83. Yeh YT, Zhou Y, Zou D, et al. Rapid Size-Based Isolation of Extracellular Vesicles by Three-Dimensional Carbon Nanotube Arrays. ACS Appl Mater Interfaces. 2020;12(11):13134–13139. doi:10.1021/acsami.9b20990
84. Aafreen S, Feng J, Wang W, Liu G. Theranostic extracellular vesicles: a concise review of current imaging technologies and labeling strategies. Extracell Vesicles Circ Nucl Acids. 2023;4(1):107. doi:10.20517/evcna.2023.01
85. Gong L, Zhang Y, Liu C, Zhang M, Han S. Application of radiosensitizers in cancer radiotherapy. Int J Nanomed. 2021;Volume 16:1083–1102. doi:10.2147/IJN.S290438
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.