Back to Journals » Neuropsychiatric Disease and Treatment » Volume 10
Berberine exerts an anticonvulsant effect and ameliorates memory impairment and oxidative stress in a pilocarpine-induced epilepsy model in the rat
Authors Gao F, Gao Y, Liu Y, Wang L, Li Y
Received 25 August 2014
Accepted for publication 23 September 2014
Published 13 November 2014 Volume 2014:10 Pages 2139—2145
DOI https://doi.org/10.2147/NDT.S73210
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Wai Kwong Tang
Fei Gao,1,* Ying Gao,2,* Yang-feng Liu,3 Li Wang,4 Ya-jun Li1
1Department of Neurology, First Affiliated Hospital of Xi’an Medical University, Xi’an, People’s Republic of China; 2Department of Radiotherapy Oncology, First Affiliated Hospital of Medical College of Xi’an Jiaotong University, Xi’an, People’s Republic of China; 3Department of Neurology, People’s Liberation Army No. 451 Hospital, Xi’an, People’s Republic of China; 4Department of Scientific Research, First Affiliated Hospital of Xi’an Medical University, Xi’an, People’s Republic of China
*These authors contributed equally to this work
Abstract: Though new antiepileptic drugs are emerging, approximately a third of epileptic patients still suffer from recurrent convulsions and cognitive dysfunction. Therefore, we tested whether berberine (Ber), a vegetable drug, has an anticonvulsant property and attenuates memory impairment in a pilocarpine (Pilo)-induced epilepsy model in rats. The rats were injected with 400 mg/kg Pilo to induce convulsions, and Ber 25, 50, and 100 mg/kg were administrated by the intragastric route once daily 7 days before Pilo injection until the experiment was over. Convulsions were observed after Pilo injection. For the rats that developed status epilepticus (SE), malondialdehyde, glutathione levels, superoxide dismutase, and catalase activity in the hippocampus were measured 24 hours after SE. The rats received the Morris water-maze test 2 weeks after SE, and then were killed for fluoro-jade B staining to detect the degenerating neurons. We found Ber delayed latency to the first seizure and the time to develop SE in a dose-dependent manner. Malondialdehyde levels were decreased, while glutathione and catalase activity were strengthened in Ber-injected SE rats. In the Morris water-maze test, Ber decreased escape latency compared to saline-treated SE rats. Additionally, Ber reduced the number of fluoro-jade B-positive cells in the hippocampal CA1 region. Our data suggest that Ber exerts anticonvulsant and neuroprotective effects on Pilo-induced epilepsy in rats. Simultaneously, Ber attenuates memory impairment. The beneficial effect may be partly due to mitigation of the oxidative stress burden.
Keywords: status epilepticus, pilocarpine, memory impairment, oxidative stress, neuroprotection
Introduction
Epilepsy, a neurological disorder characterized by recurrent episodes of seizures due to an imbalance between cerebral excitability and inhibition, affects approximately 50 million people worldwide.1 Seizures in 20%–30% epileptic patients cannot be controlled effectively, though new antiepilepsy drugs (AEDs) are growing in number. A large number of patients suffer from cognitive dysfunction.2–4 The present AEDs can only ameliorate the occurrence of seizures, while they are not helpful for cognitive dysfunction. Moreover, cognitive impairment is one of the side effects of AEDs.5 Therefore, the need for newer and more efficacious AEDs is urgent.
The epilepsy model induced by pilocarpine (Pilo), a potent muscarinic cholinergic agonist, is a useful animal model to reproduce behavioral and electroencephalographic alterations similar to those in human epilepsy.6 The brain processes large amounts of O2 in a relatively small mass, and has a high content of substrates available for oxidation associated with low antioxidant activities, making it extremely susceptible to oxidative damage.7 Under normal conditions, there is a steady-state balance between the production of reactive oxygen species and their destruction by antioxidant systems. However, in the Pilo epilepsy model, this balance has been broken by increased reactive oxygen species production or/and by a decrease in cellular antioxidant systems, such as catalase (CAT) and superoxide dismutase (SOD). This imbalance contributes to irreversible neuronal damage of cell-membrane phospholipid, and it has been suggested as a possible mechanism of epileptic activity.8 Previous studies have suggested that antioxidative therapy may have beneficial effects on epilepsy.9,10
Berberine (Ber) is the major active constituent of Rhizoma coptidis and other plants, and has multiple pharmacological effects, including anticholinergic, antihypotensive, antiarrhythmic, antiosteoporotic, cardioprotective, antitumor and antimalarial effects.11 In central nervous system diseases, such as cerebral ischemia, Ber has neuroprotective function through its anti-inflammatory, antioxidative and antineuronal apoptosis pharmacological properties.12–14 Recent research has indicated that Ber has an anticonvulsant effect on a pentylenetetrazol–maximal electroshock–kainic acid-induced epileptic model in mice.15 However, the underlying mechanism remains unknown.
The present study was therefore designed to evaluate anticonvulsant activity of Ber in Pilo-induced status epilepticus (SE) rats. We also evaluated the effects of Ber on oxidative stress, memory impairment, and neuronal degeneration in this process.
Materials and methods
Animals and experimental procedures
Male Sprague Dawley rats, weighing between 220 and 250 g were used in this study. The rats were housed in a controlled environment (21°C±1°C and 60% humidity, under a 12-hour light/dark cycle, lights on at 8 am). Food and water were available ad libitum. All animal procedures were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
The rats were randomly divided into six groups in this experiment: 1) control group (n=12), administered saline alone; 2) Ber 100 group (n=12), administered Ber (100 mg/kg) alone; 3) Pilo group (n=20), administered Pilo; 4) Pilo + Ber 25 group (n=20), administered both Ber (25 mg/kg) and Pilo; 5) Pilo + Ber 50 group (n=20), administered both Ber (50 mg/kg) and Pilo; 6) Pilo + Ber 100 group (n=20), administered with both Ber (100 mg/kg) and Pilo.
Ber was dissolved in 0.9% saline and administered by intragastric route once daily for 7 days before Pilo injection until the experiment was over. In the control group, equivalent saline was administered instead of Ber. All rats received methylscopolamine (1 mg/kg, intraperitoneally) 45 minutes before Pilo injection to minimize the peripheral effects of Pilo. Rats were continuously monitored to detect any behaviors indicative of seizure activity for 120 minutes following Pilo administration. Epileptic activity was graded according to the Racine scale.16 An animal was considered to be in SE when continuous generalized seizure activity was observed without normal behavior in seizure episodes.17 When rats had experienced SE for 1 hour, seizures were terminated by intraperitoneal injection of 10 mg/kg diazepam. All SE animals were given water-soaked food until they were able to eat normal dry-food pellets.
At 24 hours after SE, rats were killed for the valuation of lipid peroxidation, GSH, SOD, and CAT activity. The surviving SE rats received the Morris water maze (MWM) test 14 days after SE, and were killed for fluoro-jade B (FJB) stains after the MWM test.
Drugs and chemicals
All drugs were purchased from Sigma (St Louis, MO, USA). MDA, GSH, SOD, and CAT activity assay kits were purchased from Jiancheng Bioengineering Institute (Nanjing, People’s Republic of China).
Tissue preparation
The animals were deeply anesthetized (10% chloral hydrate, 350 mg/kg, intraperitoneally) and killed by decapitation. Their brains were rapidly dissected on ice to remove the hippocampus for determination of MDA, GSH, SOD, and CAT activities. The brain samples were weighed and kept at −70°C until analyzed. Each brain sample was then homogenized in 5% w/v 20 mM phosphate buffer, pH 7.4.
For the FJB stain, rats were deeply anesthetized and perfused transcardially with 0.9% saline (100 mL), followed by 4% paraformaldehyde (200 mL). Brains were removed and postfixed overnight in 4% paraformaldehyde, and then in a 30% sucrose solution at 4°C. When brains sank to the bottom of the solution, serial coronal sections (30 μm thickness) were made through the entire hippocampus using a sliding microtome.
Lipid peroxidation level
According to a previous experiment,18 lipid peroxidation level was determined by measuring the level of thiobarbituric reactive species. This is expressed as nanomoles of MDA/g wet tissue.
Catalase activity
This was assessed according to the methods of Aebi,19 in which the CAT reaction is stopped after 1 minute with a CAT inhibitor. This is expressed as mmol/min/mg protein.
Superoxide dismutase activity
As described by Misra and Fridovich,20 SOD activity is expressed as U/mg protein.
Reduced glutathione level
GSH level was assayed spectrophotometrically as described by Beutler et al21 and is expressed as nmol/mg protein.
FJB staining
FJB staining of brain slices was carried out to identify neurons undergoing degeneration.22 Floating sections were mounted on slides and dried at room temperature. Sections were then rehydrated by sequential soaking in 100% ethanol (5 minutes), 70% ethanol (2 minutes), and distilled water (2 minutes). After 15 minutes, sections were incubated in 0.06% potassium permanganate and rinsed for 1 minute in distilled water, then immersed in a solution containing 0.0004% FJB and 0.1% acetic acid for 30 minutes. After washing, the sections were coverslipped. The number of FJB positive neuronal cells was calculated in per millimeter length of the hippocampal CA1 pyramidal cell layer.
Morris water-maze test
The MWM test is widely used for assessment of spatial memory. A test pool (diameter 140 cm, 60 cm deep) was filled with water (water temperature 22°C±1°C) made opaque by the addition of a white nontoxic paint. A platform (10×10 cm) was placed 1.5 cm below the surface of the water. The rats were placed into the pool at four different starting points. Then, rats were tested for their ability to locate the hidden platform. Rats were given two trials per day, and a trial ended when the rat escaped onto the platform. The escape latency for each trial was recorded. Each trial was started and ended manually by the experimenter. The movement of the rats was monitored by a digital camera (HVR-S270C; Sony, Tokyo, Japan).
Statistical analysis
All measurement data are expressed as means ± standard error of mean. One-way analysis of variance (ANOVA) followed by the least significant difference post hoc test was used for the analysis of FJB stain and oxidative stress study data. Two-way repeated-measure ANOVA was used to analyze data in the MWM test. For the analysis of categorical data, Fisher’s exact test was used. P<0.05 was considered statistically significant.
Results
Berberine relieved pilocarpine-induced convulsions in rats
After Pilo injection, all rats inhibited convulsive behaviors, such as head bobbing, scratching, and chewing. In the Pilo group, the first episode of seizures occurred at 16.7±2.37 minutes. However, 25 mg/kg Ber treatment significantly delayed latency to first seizure (P<0.05). In the 50 mg/kg and 100 mg/kg Ber groups, latency to first seizure was 33.8±2.75 min and 45.2±2.70 min, respectively. Compared to the Pilo group rats, latency was significantly longer (P<0.01). Time to SE in the Pilo + Ber 25 group rats was delayed significantly compared with that in the Pilo group rats (P<0.05). After Ber 50 mg/kg and Ber 100 mg/kg injection, time to SE increased to 40.8±3.13 and 50.5±3.34 minutes, respectively. They both increased significantly compared with that in the Pilo group. Rats in the control and Ber 100 groups showed no seizures in the 2 hours after saline or Ber injection. Nineteen of 20 rats developed SE and nine rats died in the Pilo group, while 14 of 20 rats developed SE and three rats died in the Pilo + Ber 50 group. The percentage SE between the two groups was significantly different (P<0.05). The incidence of SE also significantly decreased in the Pilo + Ber 100 group compared to that in the Pilo group (P<0.01). Percentage survival in Ber 100 mg/kg-treated SE rats was 90%. The experimental data are shown in Table 1.
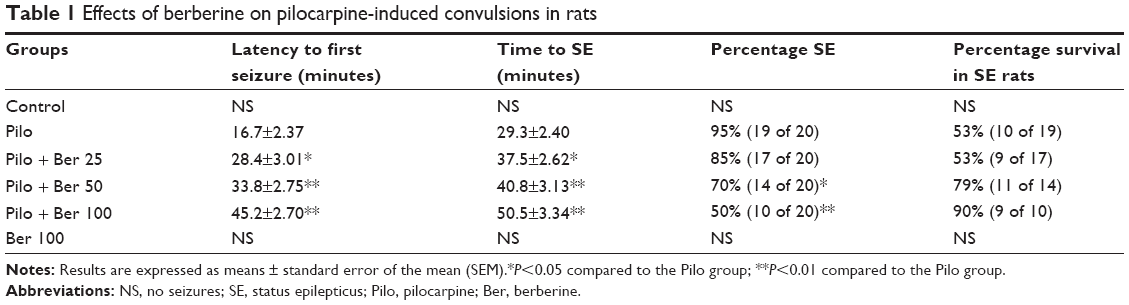 | Table 1 Effects of berberine on pilocarpine-induced convulsions in rats |
Berberine decreased the degree of oxidative stress in the hippocampus 24 hours after pilocarpine-induced SE in rats
At 24 hours after SE, the CAT level of hippocampus significantly increased in Pilo-group rats compared to saline-treated control rats (P<0.01, four to six rats per group, Figure 1A). In the Ber 50 and Ber 100 groups, hippocampal CAT levels were significantly higher than in the saline-treated SE rats (P<0.01, four to six rats per group, Figure 1A). There was a significant fall in GSH levels in hippocampal tissue of SE rats compared to saline-treated control rats (P<0.01, four to six rats per group, Figure 1B). In the Ber 25 group, GSH levels in the hippocampus significantly increased compared with saline-treated SE rats (P<0.05, four to six rats per group, Figure 1B). With increased Ber dose, there were significantly growing GSH levels of hippocampal tissue in the 50 and 100 mg/kg Ber treatment groups (both P<0.01, four to six rats per group, Figure 1B). The effects of administration of Ber on lipid peroxidation are depicted in Figure 1C. Compared with saline-treated control rats, there was a significant rise in MDA levels in the hippocampus in SE rats. Ber (25, 50 and 100 mg/kg) significantly reduced MDA levels in the hippocampus compared to SE rats in the Pilo group (P<0.05, P<0.01, and P<0.01, respectively, four to six rats per group, Figure 1C). As in our previous study,23 the SOD level in hippocampus showed no significant change between SE rats and saline-treated control rats. Three doses of Ber injection had no effect on SOD level (all P>0.05, four to six rats per group, Figure 1D). Ber 100 mg/kg per se did not influence MDA, CAT, SOD, or GSH levels.
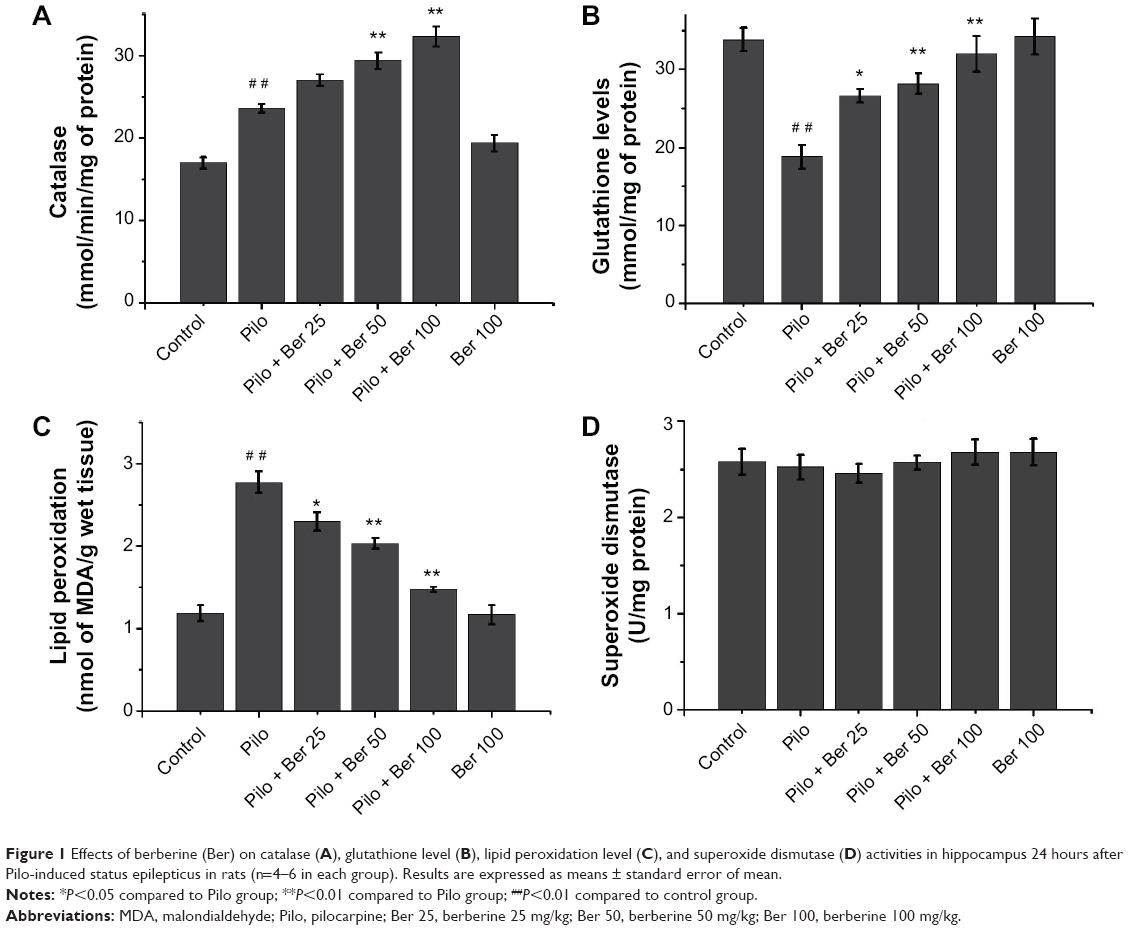 | Figure 1 Effects of berberine (Ber) on catalase (A), glutathione level (B), lipid peroxidation level (C), and superoxide dismutase (D) activities in hippocampus 24 hours after Pilo-induced status epilepticus in rats (n=4–6 in each group). Results are expressed as means ± standard error of mean. |
Berberine prevented memory impairment 2 weeks after pilocarpine-induced SE in rats
The mean escape latency for the trained rats decreased over the course of the learning trials in all the groups. Two-way repeat-measure ANOVA revealed that Ber (50 and 100 mg/kg) significantly decreased escape latency in SE rats compared to saline-treated SE rats (P<0.05 and P<0.01, respectively, five rats per group). However, Ber 25 mg/kg had no effect on escape latency. There was no significant difference in escape latency between saline-treated control rats and Ber 100 mg/kg alone-treated rats (P>0.05, five rats per group, Figure 2).
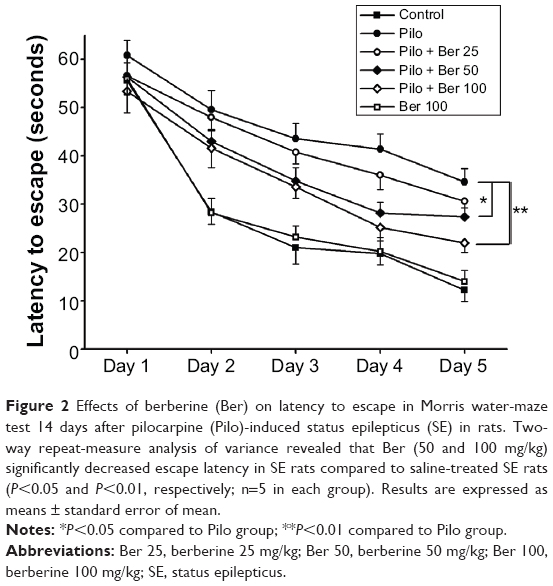 | Figure 2 Effects of berberine (Ber) on latency to escape in Morris water-maze test 14 days after pilocarpine (Pilo)-induced status epilepticus (SE) in rats. Two-way repeat-measure analysis of variance revealed that Ber (50 and 100 mg/kg) significantly decreased escape latency in SE rats compared to saline-treated SE rats (P<0.05 and P<0.01, respectively; n=5 in each group). Results are expressed as means ± standard error of mean. |
Berberine alleviated the number of FJB-positive cells in hippocampal CA1 20 days after pilocarpine-induced SE in rats
At 20 days after SE, no FJB-positive cells (degenerating neurons) were detected in hippocampus CA1 in control rats or Ber 100 mg/kg alone-treated rats. Abundant FJB-positive cells were observed in CA1 of saline-treated SE rats (Figure 3A). Compared with the saline-treated SE rats, the Ber 100 mg/kg-treated SE rats had significantly lower amounts of FJB-positive cells in CA1 (P<0.01, five rats per group, Figure 3D). The other two Ber dosages also decreased the FJB-positive cells in CA1 compared to the saline-treated SE rats (Ber 50 versus Pilo group, P<0.01; Ber 25 versus Pilo group, P<0.05; respectively; five rats per group, Figure 3B and C).
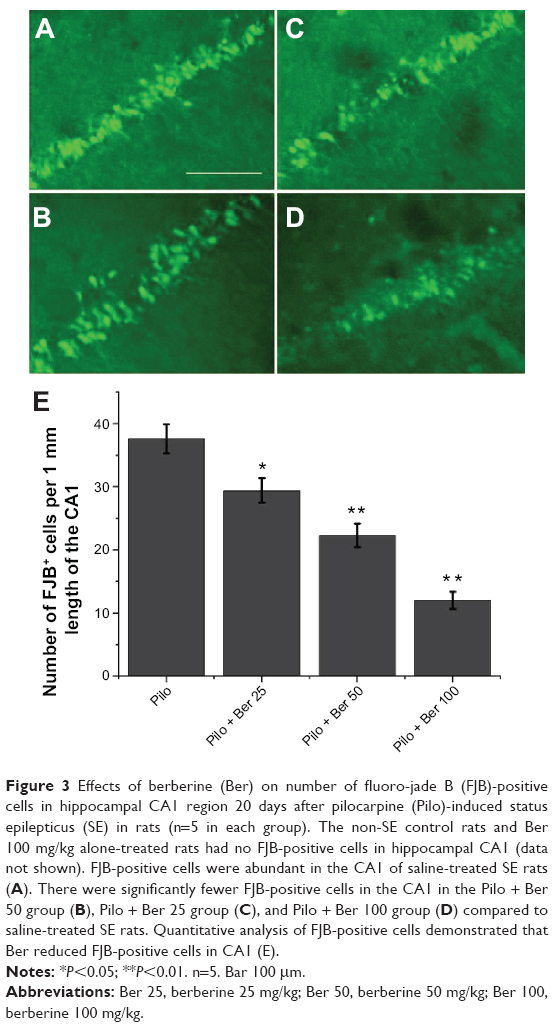 | Figure 3 Effects of berberine (Ber) on number of fluoro-jade B (FJB)-positive cells in hippocampal CA1 region 20 days after pilocarpine (Pilo)-induced status epilepticus (SE) in rats (n=5 in each group). The non-SE control rats and Ber 100 mg/kg alone-treated rats had no FJB-positive cells in hippocampal CA1 (data not shown). FJB-positive cells were abundant in the CA1 of saline-treated SE rats (A). There were significantly fewer FJB-positive cells in the CA1 in the Pilo + Ber 50 group (B), Pilo + Ber 25 group (C), and Pilo + Ber 100 group (D) compared to saline-treated SE rats. Quantitative analysis of FJB-positive cells demonstrated that Ber reduced FJB-positive cells in CA1 (E). |
Discussion
In this study, our data suggested that Ber exerts dose-dependent anticonvulsive activity in a Pilo-induced epilepsy model in rats. Simultaneously, Ber reduced the degree of oxidative stress in the hippocampus, and alleviated neuronal degeneration in hippocampal CA1 region in SE rats. Memory impairment in SE rats was also reduced by the administration of Ber.
Ber 25, 50, and 100 mg/kg all performed an anticonvulsant function. They delayed the onset of first seizure, and reduced time to SE. Ber 100 mg/kg decreased the numbers of rats developing SE and the mortality of SE rats. This result was similar to a previous study.15 The dosage of Ber used in the previous study was much lower than that in our experiment. In our preliminary dosage studies, low dosage, such as 10 mg/kg, had no effects on the seizure activity (data not shown). This may be attributed to differences in experimental animals, epileptic model, and pathway of administration.
Oxidative stress, an imbalance between prooxidants and antioxidants, has been indicated to play an important role in the pathogenesis of seizures.8 Abnormal values of oxidants and antioxidant enzymes are detected in the blood of children with refractory epilepsy.24 Reactive oxygen species have been implicated in the development of seizures and SE induced by Pilo.25,26 In our study, there was an increase in lipid peroxidation in the hippocampus of rats 24 hours after SE, which was in line with previous studies.23,25 The increased lipid peroxidation may do harm to the protein, lipids, and deoxyribonucleic acid in the brain.27 It may account for the underlying mechanisms for the deficits in cognitive function.28 Our data indicated that Ber can markedly prevent the rise in brain-lipid peroxidation demonstrated by decreased MDA level. It suggested that Ber had antioxidative potential in a Pilo-induced SE model in rats, and this pharmacological property may be dose-dependent.
CAT and SOD are important antioxidative enzymes in the brain. They can protect neuronal cells through the removal of free radicals. At 24 hours after SE, the level of CAT significantly increased in the hippocampus, in contrast to the control rats. This result is consistent with previous studies.23,25 The increase in CAT activity may be a compensatory mechanism accompanied by an immediate increase in free radicals.29 However, the activity of SOD remained unaltered at 24 hours after SE. Ber treatment had no effect on level of SOD, which suggested Ber cannot strengthen SOD activity in this condition.
GSH is another important antioxidant agent in the brain. In the present study, Pilo-induced SE decreased the GSH level in the rat hippocampus, which damaged the antioxidant defense system and may partially have been responsible for seizures and neuronal degeneration. Freitas et al also observed that hippocampal GSH contents were decreased 24 hours after SE.25 Ber significantly increased GSH levels in a dose-dependent manner compared with Pilo-induced SE rats. This demonstrated that Ber may strengthen hippocampal antioxidative ability through increasing the level of CAT and GSH, but not SOD, at this time point in Pilo-induced SE rats.
Cognitive impairment is frequently observed in epileptic patients. Though the underlying mechanisms are not well known, growing evidence indicates that oxidative stress is one of the important causes leading to memory impairment after SE.30 Oxidative damage to the rat synapse in the hippocampus has been previously reported to contribute to the deficit of cognitive functions.31–33 In our study, Ber alleviated memory impairment in rats with epilepsy induced by Pilo. Ber had no effect on the memory function in non-SE control rats. This suggests that this dosage of Ber can suppress seizure activity while having no side effect of memory impairment. On the other hand, neuron loss is one of the mechanisms underlying cognitive impairment. Ber alleviated hippocampal neuronal degeneration, demonstrated by decreasing CA1 FJB-positive cells, in our study. It also performed a neuroprotective function in other central nervous system diseases. Besides, antioxidation, anti-inflammation, and antiapoptosis may be involved in this process.12,13,34 Additionally, Ber blocked the transient outward potassium current and delayed the rectifier potassium current in a concentration-dependent manner in acutely isolated CA1 pyramidal neurons.35 It is possible that a block of potassium currents is one of the mechanisms underlying its neuroprotective action.
In conclusion, the results of this study revealed that Ber exerts an anticonvulsant effect against Pilo-induced seizures in rats. Simultaneously, Ber attenuates memory impairment and neuronal degeneration in the hippocampus. At least in part, its beneficial effect is due to its potential to mitigate the oxidative stress burden. Therefore, this study further confirms the anticonvulsant activity of Ber, although more studies are necessary to elucidate the complete mechanism involved in these effects.
Acknowledgments
This work was supported by grants from the Xi’an Science and Technology Bureau (No.YF07168), the Shaanxi Health and Family Planning Commission (No.2010H27), the National Natural Science Foundation of China (No. 81301937) and the International Cooperation Foundation of Shaanxi Province of China (No. 2013KW-27-03).
Disclosure
The authors report no conflicts of interest in this work.
References
Dua T, de Boer HM, Prilipko LL, Saxena S. Epilepsy care in the world: results of an ILAE/IBE/WHO global campaign against epilepsy survey. Epilepsia. 2006;47(7):1225–1231. | ||
Kapur N. Transient epileptic amnesia – a clinical update and a reformulation. J Neurol Neurosurg Psychiatry. 1993;56(11):1184–1190. | ||
Thompson PJ, Trimble MR. Anticonvulsant serum levels: relationship to impairments of cognitive functioning. J Neurol Neurosurg Psychiatry. 1983;46(3):227–233. | ||
Jambaqué I, Dellatolas G, Dulac O, Ponsot G, Signoret JL. Verbal and visual memory impairment in children with epilepsy. Neuropsychologia. 1993;31(12):1321–1337. | ||
Ortinski P, Meador KJ. Cognitive side effects of antiepileptic drugs. Epilepsy Behav. 2004;5 Suppl 1:S60–S65. | ||
Curia G, Longo D, Biagini G, Jones RS, Avoli M. The pilocarpine model of temporal lobe epilepsy. J Neurosci Methods. 2008;172(2): 143–157. | ||
de Freitas RL, Santos IM, de Souza GF, Tome Ada R, Saldanha GB, de Freitas RM. Oxidative stress in rat hippocampus caused by pilocarpine-induced seizures is reversed by buspirone. Brain Res Bull. 2010;81(4–5):505–509. | ||
Sudha K, Rao AV, Rao A. Oxidative stress and antioxidants in epilepsy. Clin Chim Acta. 2001;303(1–2):19–24. | ||
Kamida T, Fujiki M, Ooba H, Anan M, Abe T, Kobayashi H. Neuroprotective effects of edaravone, a free radical scavenger, on the rat hippocampus after pilocarpine-induced status epilepticus. Seizure. 2009; 18(1):71–75. | ||
Shin EJ, Jeong JH, Chung YH, et al. Role of oxidative stress in epileptic seizures. Neurochem Int. 2011;59(2):122–137. | ||
Imanshahidi M, Hosseinzadeh H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytother Res. 2008;22(8):999–1012. | ||
Hong JS, Chu YK, Lee H, et al. Effects of berberine on hippocampal neuronal damage and matrix metalloproteinase-9 activity following transient global cerebral ischemia. J Neurosci Res. 2012;90(2):489–497. | ||
Zhang X, Wang C, Li Y, et al. Neuroprotection of early and short-time applying berberine in the acute phase of cerebral ischemia: Up-regulated pAkt, pGSK and pCREB, down-regulated NF-κB expression, ameliorated BBB permeability. Brain Res. 2012;1459:61–70. | ||
Gu L, Li N, Yu W, et al. Berberine reduces rat intestinal tight junction injury induced by ischemia-reperfusion associated with the suppression of inducible nitric oxide synthesis. Am J Chin Med. 2013;41(6):1297–1312. | ||
Bhutada P, Mundhada Y, Bansod K, Dixit P, Umathe S, Mundhada D. Anticonvulsant activity of berberine, an isoquinoline alkaloid in mice. Epilepsy Behav. 2010;18(3):207–210. | ||
Racine RJ. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr Clin Neurophysiol. 1972;32(3):281–294. | ||
Chu K, Jung KH, Lee ST, et al. Erythropoietin reduces epileptogenic processes following status epilepticus. Epilepsia. 2008;49(10):1723–1732. | ||
Ruiz-Larrea MB, Leal AM, Liza M, Lacort M, de Groot H. Antioxidant effects of estradiol and 2-hydroxyestradiol on iron-induced lipid peroxidation of rat liver microsomes. Steroids. 1994;59(6):383–388. | ||
Aebi H. Catalase in vitro. Methods Enzymol. 1984;105:121–126. | ||
Misra HP, Fridovich I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J Biol Chem. 1972;247(10):3170–3175. | ||
Beutler E, Duron O, Kelly BM. Improved method for the determination of blood glutathione. J Lab Clin Med. 1963;61:882–888. | ||
Schmued LC, Hopkins KJ. Fluoro-jade: novel fluorochromes for detecting toxicant-induced neuronal degeneration. Toxicol Pathol. 2000;28(1):91–99. | ||
Liu YF, Gao F, Li XW, et al. The anticonvulsant and neuroprotective effects of baicalin on pilocarpine-induced epileptic model in rats. Neurochem Res. 2012;37(8):1670–1680. | ||
Saad K, Hammad E, Hassan AF, Badry R. Trace element, oxidant, and antioxidant enzyme values in blood of children with refractory epilepsy. Int J Neurosci. 2014;124(3):181–186. | ||
Freitas RM, Vasconcelos SM, Souza FC, Viana GS, Fonteles MM. Oxidative stress in the hippocampus after pilocarpine-induced status epilepticus in Wistar rats. FEBS J. 2005;272(6):1307–1312. | ||
Freitas RM, Sousa FC, Vasconcelos SM, Viana GS, Fonteles MM. Pilocarpine-induced status epilepticus in rats: lipid peroxidation level, nitrite formation, GABAergic and glutamatergic receptor alterations in the hippocampus, striatum and frontal cortex. Pharmacol Biochem Behav. 2004;78(2):327–332. | ||
Liang LP, Patel M. Seizure-induced changes in mitochondrial redox status. Free Radic Biol Med. 2006;40(2):316–322. | ||
Keller JN, Schmitt FA, Scheff SW, et al. Evidence of increased oxidative damage in subjects with mild cognitive impairment. Neurology. 2005;64(7):1152–1156. | ||
Frantseva MV, Velazquez JL, Hwang PA, Carlen PL. Free radical production correlates with cell death in an in vitro model of epilepsy. Eur J Neurosci. 2000;12(4):1431–1439. | ||
Ancelin ML, Christen Y, Ritchie K. Is antioxidant therapy a viable alternative for mild cognitive impairment? Examination of the evidence. Dement Geriatr Cogn Disord. 2007;24(1):1–19. | ||
Biessels GJ, Kamal A, Ramakers GM, et al. Place learning and hippocampal synaptic plasticity in streptozotocin-induced diabetic rats. Diabetes. 1996;45(9):1259–1266. | ||
Fukui K, Onodera K, Shinkai T, Suzuki S, Urano S. Impairment of learning and memory in rats caused by oxidative stress and aging, and changes in antioxidative defense systems. Ann N Y Acad Sci. 2001;928:168–175. | ||
Gispen WH, Biessels GJ. Cognition and synaptic plasticity in diabetes mellitus. Trends Neurosci. 2000;23(11):542–549. | ||
Hu J, Chai Y, Wang Y, et al. PI3K p55γ promoter activity enhancement is involved in the anti-apoptotic effect of berberine against cerebral ischemia-reperfusion. Eur J Pharmacol. 2012;674(2–3):132–142. | ||
Wang F, Zhao G, Cheng L, Zhou HY, Fu LY, Yao WX. Effects of berberine on potassium currents in acutely isolated CA1 pyramidal neurons of rat hippocampus. Brain Res. 2004;999(1):91–97. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.