Back to Journals » Infection and Drug Resistance » Volume 12
Association of the genes encoding Metallo-β-Lactamase with the presence of integrons among multidrug-resistant clinical isolates of Acinetobacter baumannii
Authors Amin M, Navidifar T ![]() , Saleh Shooshtari F
, Saleh Shooshtari F ![]() , Goodarzi H
, Goodarzi H
Received 8 December 2018
Accepted for publication 11 April 2019
Published 13 May 2019 Volume 2019:12 Pages 1171—1180
DOI https://doi.org/10.2147/IDR.S196575
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Joachim Wink
Mansour Amin,1,2 Tahereh Navidifar,2 Farkhondeh Saleh Shooshtari,2 Hamed Goodarzi2
1Infectious and Tropical Diseases Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Background: Metallo-β-Lactamases (MBL) are usually encoded on the gene cassettes harboring integrons and disseminated easily among Acinetobacter baumannii isolates. This study was aimed to investigate the association of the genes encoding MBL with the presence of class 1 and 2 integrons among multidrug-resistant (MDR) A.baumannii isolates.
Methodology: A total of 85 non-duplicated A.baumannii isolates were collected and evaluated for the amplification of blaOXA-51. The presence of genes encoding MBLs, including blaIMP, blaVIM, blaSIM, blaSPM, blaGIM, blaDIM and blaNDM, as well as intI 1 and intI 2 was evaluated by PCR. Also, the production of MBLs was screened phenotypically by the combination of EDTA and meropenem.
Results: In this study, 77 out of 85 isolates were MDR. Also, 34 isolates had only intI 1, 10 had only intI 2 and 15 had both intI 1 and intI 2. The phenotypic detection of MBLs was found in 30 isolates, among which blaVIM was as the most common the gene encoding MBL followed by blaIMP, blaSPM and blaSIM. The gene cassettes analysis revealed that class 1 integron is often responsible for transferring the genes harboring MBLs.
Conclusion: The production of MBLs among A. baumannii strains is one of the main mechanisms of resistance to carbapenems. Therefore, the development of inexpensive screening methods for the phenotypic detection of MBLs in clinical laboratories settings is essential. Also, our data revealed that the class 1 integron is often responsible for the dissemination of the MBL genes among A. baumannii isolates.
Keywords: acinetobacter baumannii, blaVIM, blaIMP, integron, Metallo-Beta-Lactamase
Introduction
Multidrug-resistant (MDR) bacterial strains have emerged as one of the leading causes of nosocomial infections worldwide. Infections caused by A. baumannii are frequent and increasing in hospitalized patients, especially in the intensive care units (ICU).1 Nowadays, the development of antibiotic resistance among A. baumannii strains is considered as one of the major public health concerns in hospital setting.2 Moreover, A. baumannii strains have a high capacity to acquire the multiple antibiotic resistance determinants through the mobile elements, such as integrons harboring single or multiple gene cassettes.
Integrons are conserved, transposon-like DNA elements that mostly encode antibiotic resistance determinants and have a high ability for chromosomal integration in bacteria.3 To date, several classes of integrons have been described; among them, class 1 and 2 integrons are frequently reported from MDR A. baumannii strains.4,5
Carbapenems have a potent activity against multidrug-resistant gram-negative bacilli and are usually the choice antibiotics against A.baumannii strains. However, the resistance rate to carbapenems in this bacterium is increasing throughout the world. The resistance to carbapenems can be led through various mechanisms, such as the production of Metallo-β-Lactamase and oxacillinase enzymes.6
More specially, the infections caused by Metallo-Beta-Lactamase (MBL)-producing organisms are associated with the high rates of morbidity and mortality.7 MBLs belong to class B beta-lactamases that can hydrolyze all beta-lactam classes except monobactams.8 MBLs are usually encoded on the gene cassettes harboring class 1 integron and disseminated easily in bacterial populations.9 To date, several MBLs were recognized such as the blaVIM, blaIMP, blaGIM, blaSPM, blaDIM, blaSIM and blaNDM which of those, the blaVIM and blaIMP allelic variants have emerged as the dominant MBLs worldwide.8,10 The high levels of resistance to carbapenems among MDR A. baumannii strains have made some demands for the reintroduction older antibiotics such as colistin and polymyxin B that had not been used for many years because of their toxicity.11 Moreover, recent studies have shown that gram-negative bacilli resistant to aminoglycosides, beta-lactams, and fluoroquinolones are often sensitive to polymyxin B.12 This study was aimed to investigate the association of the genes encoding MBLs with the presence of integrons among multidrug-resistant clinical isolates of Acinetobacter Baumannii.
Materials and methods
Bacterial isolates and identification
The present study was conducted from July 2017 to March 2018. A total of 85 A.baumannii clinical isolates were collected from different clinical samples of hospitalized patients in hospitals of Imam Khomeini and Taleghani in Ahvaz, Iran. The collected samples were as part of the routine hospital laboratory procedure and were transferred to Department of Microbiology, school of medicine, Ahvaz Jundishapur University of Medical Sciences. Then, they were cultured on Blood agar and MacConkey agar (Merck–Germany) and incubated for 24 hrs at 37°C. The gram-negative bacilli were monitored for more biochemical tests, including the sugar fermentation, motility, citrate utilization, urease, oxidative/fermentative glucose (O/F) test, catalase, oxidase and growth ability at 37°C and 42°C.13In addition, the identification of A. baumannii isolates was confirmed by the amplification of blaOXA-51-like gene using the previously described primers by Turton et al.14 The A. baumannii ATCC19606 was used as the reference strain.
Antibiotic susceptibility testing
Antimicrobial susceptibility of A. baumannii isolates was determined by disc diffusion method according to the clinical and laboratory standards institute (CLSI) guidelines.15 Briefly, the bacterial suspensions were prepared in sterile normal saline to a turbidity equivalent of 0.5 McFarland standard. The used antibiotic discs were imipenem (10 μg), meropenem (10 μg), ceftazidime (30 μg), cefotaxime (30 μg), ciprofloxacin (5 μg), gentamicin (10 μg), amikacin (30 μg), tetracycline (30 μg), piperacillin (100 μg), cefepime(30 μg), piperacillin/tazobactam (100/10 μg), trimethoprim/sulphamethoxazole (1.25/23.75 μg), colistin (10 μg), ampicillin/sulbactam (10/10 μg), ceftriaxone (30 μg) and polymexin B (300U). Then, after 24 h incubation the diameters of the inhibition zones were measured in millimeters. Also, the minimum inhibitory concentrations (MICs) of colistin, meropenem and imipenem were measured using broth microdilution method and their results were interpreted according to CLSI (2018).15 In brief, for meropenem and imipenem, a MIC ≥8 µg/ml is considered as the breakpoint of resistant, as well as a MIC ≥4 µg/ml for colistin.
MDR Acinetobacter isolates are defined as strains that were resistant to at least three classes of antimicrobial agents, including all penicillins and cephalosporins, fluoroquinolones and aminoglycosides.16
Phenotypic detection of MBL production
First, the bacterial suspensions adjusted to 0.5 McFarland were streaked on Mueller Hinton agar plates using the Dacron swab. Then, two discs of meropenem (10 μg), one with 5 μL of 0.35 M EDTA and the other without EDTA were placed on a Mueller Hinton agar plate and incubated at 37°C for 16–18 hrs. The discs containing EDTA alone served as the negative control. A strain was considered to be MBL positive, if there was an increase of ≥7 mm in the inhibition zone around the imipenem + EDTA disc as compared to imipenem disc alone.17
ERIC-PCR typing and analysis
The genetic relationship of A. baumannii isolates was determined using the enterobacterial repetitive intergenic consensus-PCR (ERIC-PCR)18 with the primers sequences of ERIC-F (5′-ATGTAAGCTCCTGGGGATTCAC-3′) and ERIC-R (5′AAGTAAGTGACTGGGGTGA GCG-3′). The PCR reaction was performed in the final volume of 25 µL as follows: 1U Taq DNA polymerase, 1.5 mM MgCl2, 200 μM dNTPs, 0.35 μM of each primer, 10x PCR buffer, 6.5 μL of template DNA and distilled water up to a final volume of 25 μL. The amplification process was performed in Mastercycler Nexus Thermal Cycler Gradient (Eppendorf, Hamburg, Germany) with one cycle of initial denaturation at 94°C for 5 mins, followed by 35 cycles of denaturation at 94°C for 60 s, annealing at 57°C for 60 s, extension at 72°C for 80 s and a cycle of final extension at 72°C for 10 mins. The amplified products were visualized on agarose gel 1.5%, stained with safe stain. The data analysis was performed using the Gel Compare II software version 6.6 (Applied Math, Sint-Martens-Latem, Belgium). The similarity pattern was calculated using the Unweighted-Pair Group Method (UPGMA)/the Dice similarity coefficient with a position tolerance of 1%. Isolates with more than 90% similarity were considered as a clonal type.
Molecular method
The whole genomes of all MDR A. baumannii isolates were extracted using High Pure PCR Template Preparation Kit (Roche Diagnosis, Mannheim, Germany) according to manufacturer’s procedure. The Uniplex PCR reactions were performed for the presence of genes encoding intI1, intI2, blaIMP, blaVIM, blaDIM, blaGIM, blaSIM, blaNDM and blaSPM in a final volume of 25 μL, as described previously.19–22 In each PCR run, the distilled water was used as the negative control. The reaction mixture consisted of 1 U of AmpliTaq DNA polymerase, 1X PCR buffer, 1.5 mM MgCl2, 200 µM dNTPs, 3 μL of DNA and distilled water up to a final volume of 25 μL. The primer concentrations were as follows: 0.2 pmol/µL each of primers IntI1-F, IntI1-R, IntI2-F and IntI2-R; 0.45 pmol/µL each of primers blaVIM -F, blaVIM-R, blaIMP-F and blaIMP -R; 0.25 pmol/µL each of primers blaGIM-F, blaGIM-R, blaDIM-F and blaDIM-R; and 0.45 pmol/µL each of primers blaSIM-F, blaSIM-R, blaNDM-F, blaNDM-R, blaSPM-F and blaSPM-R. The amplification process was performed in a Mastercycler Nexus Thermal Cycler Gradient (Eppendorf, Hamburg, Germany) with one cycle initial denaturation at 95°C for 5 mins; 35 cycles with a denaturation temperature of 95°C for 45 s; annealing temperature of 51°C for the IntI1 and IntI2 genes, 54°C for the blaIMP and blaVIM genes, 53°C for the blaOXA-51-like gene, 52°C for the blaGIM, blaSIM and blaSPM genes, as well as 58°C for the blaNDM and blaDIM genes for 30 s and extension temperature of 72°C for 30 s, followed by a cycle of final extension at 72°C for 10 mins. All of the PCR products were visualized on 1% agarose gel stained with safe stain. DNA sequencing of PCR products was performed by (Bioneer, South Korea) for the determination of the MBL allelic variants.
Sequencing of integron gene cassettes
Amplification of the variable region of class 1 and 2 integrons was performed, as previously by Moura et al23. Then, the purification of the PCR products was performed by the QIAquick Gel Extraction Kit (Qiagen, Germany) and subjected to sequencing with an ABI Prism 377 automated sequencer (Applied Biosystems, USA). The obtained sequences were assembled using MEGA 724 and compared with those in the NCBI database using a BLAST program (
Statistical analysis
The descriptive statistics and Chi-Square test were performed in SPSS version 16.00 with a significance level of p<0.05.
Results
Bacterial isolates and determination of antibiotic susceptibility
In this cross-sectional study, 85 non-duplicated A. baumannii isolates were collected from the different clinical samples, including burn wounds 22 (25.88%), tracheal secretion 31 (36.47%), blood 16 (18.82%), bronchial lavage 12 (14.11%) and urine 4 (4.7%) isolates and the mean age of the patients was 62.1±4.75 years. According to antibiotic susceptibility testing, 77 out of 85 (90.58%) A. baumannii isolates were identified as MDR.
In our study, among 77 MDR A. baumannii isolates, resistance to amikacin, ceftazidime, ceftriaxone, cefepime, ciprofloxacin, cefotaxime, gentamicin, imipenem, meropenem, piperacillin/tazobactam, piperacillin, ampicillin/sulbactam, trimethoprim/sulfamethoxazole and tetracycline was seen in 71 (92.2%), 75 (97.4%), 76 (98.7%), 75 (97.4%), 75 (97.4%), 76 (98.7%), 76 (98.7%), 69 (89.6%), 73 (94.8%), 75 (97.4%), 75 (97.4%), 43 (55.8%), 74 (96.1%) and 47 (61.03%) isolates, respectively. Also, all isolates were sensitive to polymyxin B and only two isolates were resistant to colistin. The MICs of carbapenems and colistin among 85 A.baumanni isolates are shown in Table 1.
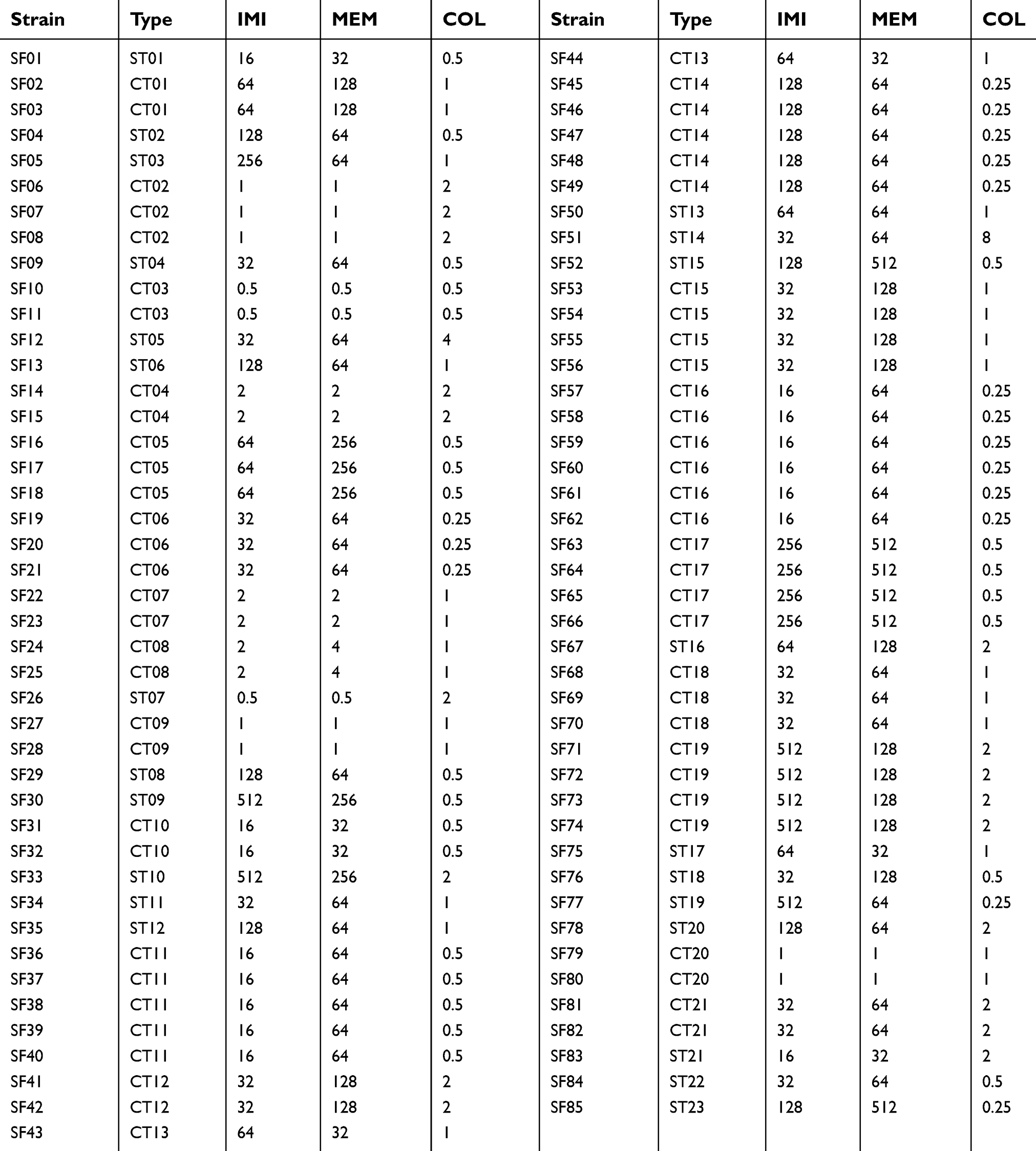 | Table 1 Distribution of resistance to meropenem, imipenem and colistin with regard to ERIC PCR types among 85 A. baumannii isolates |
ERIC-PCR analysis
In our study, 85 A.baumanni isolates were classified into 21 clone types and 23 single type of ERIC-PCR. Figure 1 is shown the dendrogram of ERIC-PCR of these isolates. Also, Table 1 shows the distribution of MICs of imipenem, meropenem and colistin among these isolates with respect to ERIC-PCR types. According to these results, there was a significant association (p<0.05) between the clone types and antibiotic susceptibility to carbapenem agents and colistin.
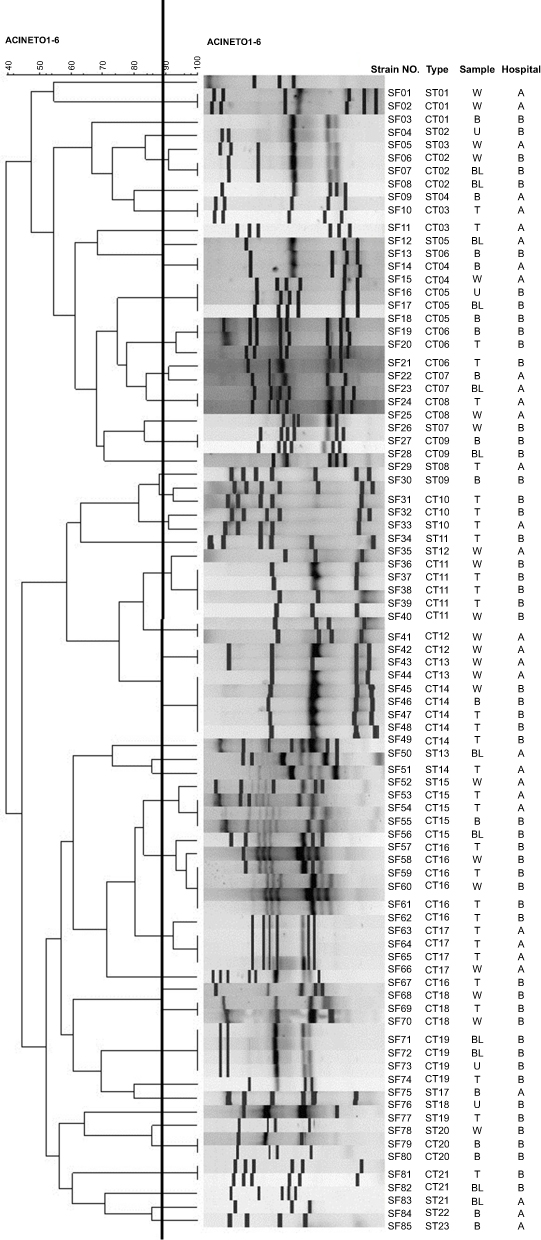 | Figure 1 Dendrogram of 85 A. baumannii clinical isolates based on ERIC-PCR types. Abbreviations: CT, clone type; ST, single type; W, burn wound; T, tracheal secretion; B, blood; BL, bronchial lavage; U, urine; Hospital A, Imam Khomeini; B, Taleghani Hospital. |
Detection of genes encoding MBLs and intI1 and intI2
In our study, the frequency rates of the genes encoding blaIMP, blaVIM, blaSIM and blaSPM, among 77 MDR A. baumannii isolates were 10 (12.98%), 17 (22.07%), 2 (2.59%) and 4 (5.19%), respectively. In addition, none of the genes encoding blaGIM, blaDIM and blaNDM was detected in these isolates. Also, none of the genes encoding MBLs was detected in non-MDR isolates.
Moreover, 7 isolates carried only the blaIMP gene derivatives, 14 carried only the blaVIM gene derivatives, 3 carried both the blaVIM and blaIMP genes derivatives, 4 carried only the blaSPM-1 gene and 2 carried only the blaSIM-1 gene. The distribution of allelic variants of blaIMP and blaVIM is shown in Table 2. According to these results, blaVIM-2 was the most prevalent variant of blaVIM gene. In this study, the amplification of the intI 1 and intI2 genes was performed using PCR. Of the 77 MDR A. baumannii isolates, 34 had only intI 1, 10 had only int 2 and 15 had both the intI 1 and intI 2 genes.
 | Table 2 Pattern of allelic variants of blaIMP and blaVIM |
Association of phenotypic detection of MBL production with genes encoding MBLs
Among 73 carbapenem-resistant A. baumannii isolates, 30 were phenotypically as MBL-producing isolates. Moreover, of these 30 isolates, 7 carried only the blaIMP gene derivatives, 14carried only the blaVIM gene derivatives, 2 carried both the blaVIM and blaIMP gene derivatives, 4 carried the blaSPM-1 gene and 2 carried the blaSIM-1 gene. However, one strain did not carry any gene encoding MBL. Overall, 29 isolates presenting MBL phenotype carried at least one of the MBL genes, confirming the efficacy of the phenotypic detection of MBL producing strains with the PCR results.
On the other hand, the phenotypic detection of MBL was negative for one blaVIM positive A. baumannii isolate and one blaIMP positive isolates.
Association of the presence of integrons with genes encoding MBLs among MDR A. baumannii
Table 3 indicates the distribution of gene cassettes carrying MBLs among integron-positive A.baumannii isolates. Eight gene cassette arrays were detected within class 1 integron and three gene cassette arrays within class 2 integron. The most prevalent gene cassette arrays among positive class 1 integron isolates, blaIMP-19,aacA31, blaOXA-21,aadA-1 and blaVIM-1,qacED-1, were detected among 10 isolates. According to these results, blaVIM allelic variants were as the part of gene cassettes incorporated into class 1 integron among 10 isolates and as the part of gene cassettes in class 2 integron among 2 isolates. On the other hand, blaIMP derivatives were as the part of gene cassettes incorporated into class 1 integron among 4 isolates and into class 2 integron among 1 isolate. Also, 2 isolates carried both blaVIM and blaIMP allelic variants in gene cassettes incorporated into class 1 integron and one isolate carried only blaIMP in gene cassette incorporated into class 1 integron. In addition, 2 isolates carring blaVIM and 2 isolates carring blaIMP were lack either intI1 or intI2.
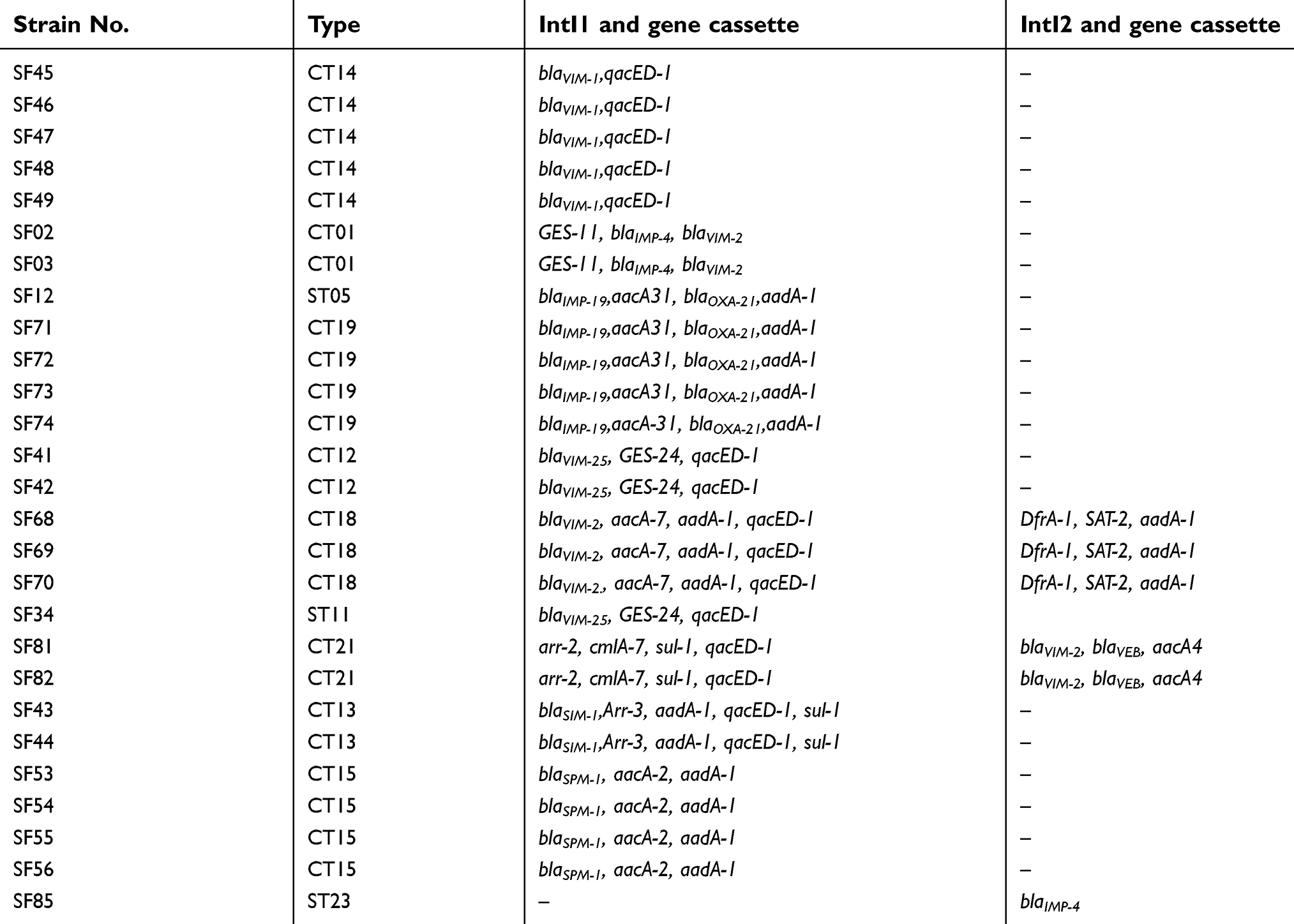 | Table 3 Distribution of gene cassettes carrying MBLs among integron-positive A.baumannii isolates |
According to the results shown in Table 3, the isolates belonging to a same clone type had the similar gene cassette array in class 1 and 2 integron.
Discussion
A.baumannii is an important nosocomial pathogen with the high associated mortality. In the last few years, the resistance to the almost commonly prescribed antibiotics among A.baumannii strains is increasing which will cause a treatment challenge in the future.25
The results of our study showed that 90.58% of A. baumannii isolates were MDR. In agreement with our results, the high prevalence of MDR A. baumannii isolates was reported from other studies, ranged from 49.6% to 100%.26–31The multidrug antibiotic resistance has often limited the efficacy of the common therapeutic options especially for the strains that are resistant to carbapenems. In the current study, the resistance rates to carbapenem agents (imipenem or meropenem) were similar to a previous study by Shoja et al32 in the same region during 2011 to 2012 years, indicating that the prevalence of MDR A.baumannii isolates is still high in our region.
Our results showed that the antibiotic resistance rates to amikacin, ceftazidime, ceftriaxone, cefepime, ciprofloxacin, cefotaxime, gentamicin, meropenem, piperacillin/tazobactam and piperacillin among MDR A. baumannii strains were more than 90%. Similar to our work, Mirnejad et al33, Huang et al5 and Taherikalani et al34 also reported the high percentages of the antibiotic resistance among A. baumannii isolates.
As mentioned earlier, polymyxins are recommended as the antibiotic choices for MDR A. baumannii infections. In our study, all isolates were susceptible to polymyxin B which was in concordance with the studies conducted by Najar Peerayeh et al35 and Shoja et al32 in Iran. However, in contrast to our results, the higher resistance rates to polymyxin B were reported in other regions of Iran, including 14% in Tehran,36 16% in Tabriz37 and 11% in Kermanshah.38 It seems that this growing resistance could be due to the excessive usage of this antibiotic in the treatment of severe infections. Surprisingly, the resistance level to polymyxin B in Brazil39 was much high (81.5%). This high resistance might be due to the prolonged use of this antibiotic agent in treatment of carbapenem-resistant A. baumannii infections in this country.39 Our results showed that the majority of A. baumannii isolates were susceptible to colistin which is in agreement with a previous study32 in our region, suggesting polymyxin B and colistin are still the most effective antibiotic agents against MDR A. baumannii strains.
In our study, the blaIMP and blaVIM allelic variants were recognized as the most common genes encoding MBLs in the majority of isolate with the positive results in the phenotypic detection of MBL. However, in the one isolate that was phenotypically positive for MBL production, any gene encoding MBL was not detected using PCR. It seems that MBL phenotype in this isolate was caused by other mechanisms rather than the presence of genes encoding MBLs that unfortunately were not considered in our study.
In consistent with our work, Lee et al40 in Seoul found the blaIMP and blaVIM genes allelic variants in most A.baumannii isolates, whereas the blaSIM-1 gene was recognized only in a few isolates. However, in contrast to our results, Shahcheraghi et al41 in Iran did not find either blaIMP or blaVIM genes, instead the blaSPM gene was recognized in the A. baumannii isolates.
In our study, the phenotypic detection of MBL was negative in one blaVIM -positive isolate and one blaIMP - positive isolate. Similar to our study, Ikonomidis et al42 also, reported two A. baumannii isolates harboring blaVIM-1 gene which were phenotypically negative for MBL production. Moreover, to find the reason of this phenomenon, the researchers evaluated the blaVIM-1 expression in these two isolates, indicating that one of these isolates had a weak P1 promoter, and both these isolates had the inactivated P2 promoters. Hence, the blaVIM-1 expression level was reduced significantly and these isolates showed a negative phenotype in MBL test.
The integrons as the mobile genetic elements play an important role in the dissemination of antibiotic resistance determinants among A.baumannii isolates. In recent years, the frequency rates of integrons are increasing, so that they have caused a serious threat for the spread of antibiotic resistance elements.43
In our study, the prevalence of the intI1 gene was more than the intI2 gene that is in agreement with the results obtained from studies of Huang et al5 in China, Japoni et al44 and Taherikalani et al34 in Iran. However, unlike our study, Mirnejad et al33 in Tehran and Ramírez et al43 in Buenos Aires found higher frequency of the intI2 gene than the intI1 gene. The difference in data is often dependent on the integron classes of clones which are widely disseminated in the community and nosocomial settings.
Our results showed that class 1 integron is often responsible for transferring the gene cassettes harboring MBLs, especially the blaVIM and blaIMP allelic variants. In consistent with our results, Tsakris et al45 and Mendes et al46 associated the presence of class 1 integron with gene cassettes encoding blaVIM and blaIMP allelic variants. Moreover, Mendes et al indicated the presence of the blaIMP-1 gene in the gene cassette of blaIMP1_ aac(6′)-31_ aadA1 which was plasmid located in five of the seven isolates. Also, Goudarzi et al47 showed the presence of gene cassettes encoding blaVIM and blaIMP allelic within both class 1 and 2 integrons, suggesting the class 1 integron has the important role in the horizontal transfer of gene cassettes encoding MBLs.46
In our study, the most prevalent gene cassette arrays among positive class 1 integron and MBLs isolates were blaIMP-19_aacA31_blaOXA-21_aadA-1 and blaVIM-1_qacED-1.
In consistent with our results, Goudarzi et al47 showed seven different gene cassettes in 89 class 1 integron-carrying isolates and three gene cassettes in 15 class 2 integron-harboring A. baumannii isolates that among them, five different gene cassettes harbored gene encoding MBLs (VIM-25-GES-24-qacF, IMP-4, VIM-2-VEB-aacA4 and GES-11-IMP-4-VIM-2).
In our study, the majority of gene cassettes encoding MBL genes harbored genes encoding resistance to aminoglycosides as shown in a previous study by Farshadzadeh et al.48 Moreover, they indicated that gene cassettes encoding resistance to aminoglycosides were present in the majority of MDR A. baumannii isolates, suggesting the high-level resistance rates to aminoglycoside agents among A. baumannii isolates.
Also, according to the results obtained from ERIC-PCR analysis, the isolates belonging to a same clone type had the similar gene cassette array in class 1 and 2 integrons, indicating the importance of molecular typing methods in epidemiological studies for finding the distribution of clonal types disseminated in a hospital or a geographical region.
Conclusion
We demonstrated a high prevalence of resistance to carbapenems, as well as the genes encoding MBLs among MDR A. baumannii isolates. Hence, the results of our study showed that MBLs have an important role in the resistance to carbapenem among MDR A. baumannii isolates. Therefore, the development of simple and inexpensive screening methods for detecting MBL production in microbiology laboratories is essential. In this study, we indicated polymyxins as the only option of effective antibiotic in vitro against MDR A. baumannii isolates. Also, our data revealed that the class I integron had a significant role in the dissemination of blaVIM gene among clinical isolates of A. baumannii in Ahvaz, Iran.
Acknowledgments
This work is a part of M. Sc. thesis of Farkhondeh Saleh Shooshtari which has been approved in the Department of Microbiology of Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran. The authors thank the Health Research Institute, Infectious and Tropical Diseases Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran for financial support. This study was supported by funds from Health Research Institute, Infectious and Tropical Diseases Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran.
Author contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ghajavand H, Esfahani BN, Havaei SA, Moghim S, Fazeli H. Molecular identification of Acinetobacter baumannii isolated from intensive care units and their antimicrobial resistance patterns. Adv Biomed Res. 2015;4:110.
2. Hanlon GW. The emergence of multidrug resistant Acinetobacter species: a major concern in the hospital setting. Lett Appl Microbiol. 2005;41(5):375–378. doi:10.1111/j.1472-765X.2005.01791.x
3. Gillings MR. Integrons: past, present, and future. Microbiol Mol Biol Rev. 2014;78(2):257–277. doi:10.1128/MMBR.00056-13
4. Martins N, Picão RC, Adams-Sapper S, Riley LW, Bm M. Association of class 1 and 2 integrons with multidrug-resistant Acinetobacter baumannii international clones and Acinetobacter nosocomialis isolates. Antimicrob Agents Chemother. 2015;59(1):698–701. doi:10.1128/AAC.02415-14
5. Huang C, Long Q, Qian K, et al. Resistance and integrin characterization of Acinetobacter baumannii in a teaching hospital in Chongqing, China. New Microbes New Infect. 2015;8:103–108. doi:10.1016/j.nmni.2015.09.015
6. Quale J, Bratu S, Landman D, Heddurshetti R. Molecular epidemiology and mechanisms of carbapenem resistance in Acinetobacter baumannii endemic in New York City. Clin Infect Dis. 2003;37(2):214–220. doi:10.1086/375875
7. Tsakris A, Poulou A, Kristo I, et al. Large dissemination of VIM-2-metallo-{beta}-lactamase-producing pseudomonas aeruginosa strains causing health care-associated community-onset infections. J Clin Microbiol. 2009;47(11):3524–3529. doi:10.1128/JCM.01099-09
8. Bebrone C. Metallo-beta-lactamases (classification, activity, genetic organization, structure, zinc coordination) and their superfamily. Biochem Pharmacol. 2007;74(12):1686–1701. doi:10.1016/j.bcp.2007.05.021
9. Khosravi Y, Tay ST, Vadivelu J. Analysis of integrons and associated gene cassettes of metallo-β-lactamase-positive Pseudomonas aeruginosa in Malaysia. J Med Microbiol. 2011;60(Pt7):988–994. doi:10.1099/jmm.0.029868-0
10. Anoar KA, Ali FA, Omer SA. Detection of metallo [Beta]-lactamase enzyme in somegram negative bacteria isolated from burn patients in Sulaimani city, Iraq. Eur Sci J. 2014;10(3):485–496.
11. Zavascki AP, Goldani LZ, Li J, Nation RL. Polymyxin B for the treatment of multidrug-resistant pathogens: a critical review. J Antimicrob Chemother. 2007;60(2):1206–1215. doi:10.1093/jac/dkm357
12. Evans ME, Feola DJ, Rapp RP. Polymyxin B sulfate and colistin: old antibiotics for emerging multiresistant gram-negative bacteria. Ann Pharmacother. 1999;33(9):960–967. doi:10.1345/aph.18426
13. Hall GS. Non-fermenting and miscellaneous gram-negative bacilli. In: Mahon CR, Lehman DC, Manuselis G, Heights M, editors. Textbook of Diagnostic Microbiology. 4th ed. Mo: Saunders/Elsevier; 2011:482–501.
14. Turton JF, Woodford N, Glover J, Yarde S, Kaufmann ME, Pitt TL. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J Clin Microbiol. 2006;44:2974–2976. doi:10.1128/JCM.01021-06
15.
16. Manchanda V, Sanchaita S, Singh N. Multidrug resistant acinetobacter. J Glob Infect Dis. 2010;2(3):291–304. doi:10.4103/0974-777X.68538
17. Yong D, Lee K, Yum JH, Shin HB, Rossolini GM, Chong Y. Imipenem-EDTA disk method for differentiation of metallo-beta-lactamase-producing clinical isolates of Pseudomonas spp. and Acinetobacter spp. J Clin Microbiol. 2002;40(10):3798–3801.
18. Tavakol M, Momtaz H, Mohajeri P, Shokoohizadeh L, Tajbakhsh E. Genotyping and distribution of putative virulence factors and antibiotic resistance genes of Acinetobacter baumannii strains isolated from raw meat. Antimicrob Resist Infect Control. 2018;7:120. doi:10.1186/s13756-018-0405-2
19. Koeleman JGM, Stoof J, Van Der Bijl MW, Cmje V-G, Savelkoul PHM. Identification of epidemic strains of Acinetobacter baumannii by integrase gene PCR. J Clin Microbiol. 2001;39(1):8–13. doi:10.1128/JCM.39.1.8-13.2001
20. Fallah F, Noori M, Hashemi A, et al. Prevalence of blaNDM, blaPER, blaVEB, blaIMP, and blaVIM Genes among Acinetobacter baumannii Isolated from Two Hospitals of Tehran, Iran. Scientifica (Cairo). 2014;245162.
21. Aksoy MD, Çavuşlu Ş, Tuğrul HM. Investigation of Metallo Beta lactamases and oxacilinases in carbapenem resistant Acinetobacter baumannii strains isolated from inpatients. Balkan Med J. 2015;32(1):79–83. doi:10.5152/balkanmedj.2015.15302
22. Mlynarcik P, Roderova M, Kolar M. Primer evaluation for PCR and its application for detection of carbapenemases in enterobacteriaceae. Jundishapur J Microbiol. 2016;9(1):e29314. doi:10.5812/jjm
23. Moura A, Henriques I, Ribeiro R, Correia A. Prevalence and characterization of integrons from bacteria isolated from a slaughterhouse wastewater treatment plant. J Antimicrob Chemother. 2007;60(6):1243–1250. doi:10.1093/jac/dkm340
24. Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870–1874. doi:10.1093/molbev/msw054
25. Gaynes R, Edwards JR. National nosocomial infections surveillance system. Overview of nosocomial infections caused by gram-negative bacilli. Clin Infect Dis. 2005;41(6):848–854. doi:10.1086/444499
26. Banerjee T, Mishra A, Das A, Sharma S, Barman H, Yadav G. High prevalence and endemicity of multidrug resistant Acinetobacter spp. in intensive care unit of a tertiary care Hospital, Varanasi, India. J Pathog. 2018;2018:9129083. doi:10.1155/2018/9129083
27. Begum S, Hasan F, Hussain S, Ali Shah A. Prevalence of multi drug resistant Acinetobacter baumannii in the clinical samples from Tertiary Care Hospital in Islamabad, Pakistan. Pak J Med Sci. 2013;29(5):1253–1258.
28. De Francesco MA, Ravizzola G, Peroni L, Bonfanti C, Manca N. Prevalence of multidrug-resistant Acinetobacter baumannii and Pseudomonas aeruginosa in an Italian hospital. J Infect Public Health. 2013;6(3):179–185. doi:10.1016/j.jiph.2012.11.006
29. Tuan Anh N, Nga TV, Tuan HM, et al. Molecular epidemiology and antimicrobial resistance phenotypes of Acinetobacter baumannii isolated from patients in three hospitals in southern VIETNAM. J Med Microbiol. 2017;66(1):46–53. doi:10.1099/jmm.0.000418
30. Shrestha S, Tada T, Miyoshi-Akiyama T, et al. Molecular epidemiology of multidrug-resistant Acinetobacter baumannii isolates in a university hospital in Nepal reveals the emergence of a novel epidemic clonal lineage. Int J Antimicrob Agents. 2015;46(5):526–531. doi:10.1016/j.ijantimicag.2015.07.012
31. Castilho SRA, Godoy CSM, Guilarde AO, et al. Acinetobacter baumannii strains isolated from patients in intensive care units in Goiânia, Brazil: molecular and drug susceptibility profiles. PLoS One. 2017;12(5):e0176790. doi:10.1371/journal.pone.0176790
32. Shoja S, Moosavian M, Rostami S, et al. Dissemination of carbapenem-resistant Acinetobacter baumannii in patients with burn injuries. J Chin Med Assoc. 2017;80(4):245–252. doi:10.1016/j.jcma.2016.10.013
33. Mirnejad R, Mostofi S, Masjedian F. Antibiotic resistance and carriage class 1and 2 integrons in clinical isolates of Acinetobacter baumannii from Tehran,Iran. Asian Pac J Trop Biomed. 2013;3(2):140–145. doi:10.1016/S2221-1691(13)60038-6
34. Taherikalani M, Fatolahzadeh B, Emaneini M, Soroush S, Mm F. Distribution of different carbapenem resistant clones of Acinetobacter baumannii in Tehran hospitals. New Microbiol. 2009;32(3):265–271.
35. Najar Peerayeh S, Karmostaji A. Molecular identification of resistance determinants, integrons and genetic relatedness of extensively drug resistant Acinetobacter baumannii isolated from hospitals in Tehran, Iran. Jundishapur J Microbiol. 2015;8(7):e27021. doi:10.5812/jjm
36. Bahador A, Taheri M, Pourakbari B, et al. Emergence of rifampicin, tigecycline, and colistin-resistant Acinetobacter baumannii in Iran; spreading of MDR strains of novel International Clone variants. Microb Drug Resist. 2013;19(5):397–406. doi:10.1089/mdr.2012.0233
37. Aliakbarzade K, Farajnia S, Karimi Nik A, Zarei F, Tanomand A. Prevalence of aminoglycoside resistance genes in Acinetobacter baumannii isolates. Jundishapur J Microbiol. 2014;7(10):e11924. doi:10.5812/jjm.11924
38. Mohajeri P, Farahani A, Feizabadi MM, Ketabi H, Abiri R, Najafi F. Antimicrobial susceptibility profiling and genomic diversity of Acinetobacter baumannii isolates: A study in western Iran. Iran J Microbiol. 2013;5(3):195–202.
39. Genteluci GL, Gomes DB, Souza MJ, Carvalho KR, Villas-Bôas MH. Emergence of polymyxin B-resistant Acinetobacter baumannii in hospitals in Rio de Janeiro. J Bras Patol Med Lab. 2016;52(2):91–95.
40. Lee K, Yum JH, Yong D, et al. Novel acquired metallobeta- lactamase gene, bla(SIM-1), in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother. 2005;49(11):4485–4491. doi:10.1128/AAC.49.11.4485-4491.2005
41. Shahcheraghi F, Abbasalipour M, Feizabadi M, Ebrahimipour G, Akbari N. Isolation and genetic characterization of metallo-β-lactamase and carbapenamase producing strains of Acinetobacter baumannii from patients at Tehran hospitals. Iran J Microbiol. 2011;3(2):68–74.
42. Ikonomidis A, Ntokou E, Maniatis AN, Tsakris A, Pournaras S. Hidden VIM-1 metallobeta-lactamase phenotypes among Acinetobacter baumannii clinical isolates. J Clin Microbiol. 2008;46(1):346–349. doi:10.1128/JCM.01670-07
43. Ramírez MS, Stietz MS, Vilacoba E, et al. Increasing frequency of class 1 and 2 integrons in multidrug-resistant clones of Acinetobacter baumannii reveals the need for continuous molecular surveillance. Int J Antimicrob Agents. 2011;37(2):175–177. doi:10.1016/j.ijantimicag.2010.10.006
44. Japoni S, Japoni A, Farshad S, Ali AA, Jamalidoust M. Association between existence of integrons and multi-drug resistance in Acinetobacter isolated from patients in southern Iran. Pol J Microbiol. 2011;60(2):163–168.
45. Tsakris A, Ikonomidis A, Pournaras S, et al. VIM-1 metallo-beta-lactamase in Acinetobacter baumannii. Emerg Infect Dis. 2006;12(6):981–983.
46. Mendes RE, Castanheira M, Toleman MA, Sader HS, Jones RN, Walsh TR. Characterization of an integron carrying blaIMP-1 and a new aminoglycoside resistance gene, aac(6‘)-31, and its dissemination among genetically unrelated clinical isolates in a Brazilian hospital. Antimicrob Agents Chemother. 2007;51(7):2611–2614. doi:10.1128/AAC.00838-06
47. Goudarzi H, Azad M, Seyedjavadi SS, et al. Characterization of integrons and associated gene cassettes in Acinetobacter baumannii strains isolated from intensive care unit in Tehran, Iran. J Acute Dis. 2016;5(5):386–392. doi:10.1016/j.joad.2016.08.004
48. Farshadzadeh Z, Hashemi FB, Rahimi S, et al. Wide distribution of carbapenem resistant Acinetobacter baumannii in burns patients in Iran. Front Microbiol. 2015;6:1146. doi:10.3389/fmicb.2015.01146
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.