Back to Journals » Neuropsychiatric Disease and Treatment » Volume 14
Association of HLA alleles with autism
Authors Sayad A, Akbari MT, Noroozi R, Omrani MD ![]() , Inoko H, Taheri M
, Inoko H, Taheri M ![]() , Ghafouri-Fard S
, Ghafouri-Fard S ![]()
Received 6 September 2018
Accepted for publication 8 November 2018
Published 27 November 2018 Volume 2018:14 Pages 3259—3265
DOI https://doi.org/10.2147/NDT.S186673
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Arezou Sayad,1 Mohammad Taghi Akbari,2 Rezvan Noroozi,3 Mir Davood Omrani,1,4 Hidetoshi Inoko,5 Mohammad Taheri,6 Soudeh Ghafouri-Fard1
1Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran; 2Department of Medical Genetics, Faculty of Medical Sciences, Tarbiat Modares University, Tehran, Iran; 3Photochemistry Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran; 4Urogenital Stem Cell Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran; 5Department of Molecular Life Science, Tokai University, Hiratsuka, Japan; 6Student Research Committee, Shahid Beheshti University of Medical Sciences, Tehran, Iran
Background: Autism spectrum disorders (ASD) are a group of heterogeneous neurodevelopmental disorders known by impaired social interaction and activities and abnormal repetitive behavior. As a multifactorial disorder, several genetic and immunological factors have been shown to be implicated in its pathogenesis.
Methods: Among them are certain human leukocyte antigen (HLA) alleles. In the current study, we genotyped HLA-A, -B & DRB alleles in 103 Iranian ASD patients and 180 age, gender, and ethnic-matched healthy controls.
Results: After Boferroni correction no allele or haplotype was associated with genetic susceptibility to ASD in Iranian population.
Conclusion: Future studies are needed to assess contribution of immunological factors such as HLA alleles in ASD pathogenesis.
Keywords: autism, HLA typing, immune response
Introduction
Autism spectrum disorders (ASD) (MIM 209850) are a group of various neurodevelopmental disorders known by impaired social interaction and activities in addition to abnormal repetitive behavior.1 Although the main cause of ASD is unknown, as a multifactorial disorder, both genetic and environmental factors are involved in the development of ASD.2 In addition, based on the results of the numerous studies regarding the connections between the immune system and the nervous system, immunological factors are thought to exert a role in the pathogenesis of ASD.3 Abnormal cytokine profiles, reduced lymphocyte numbers, reduced T-cell mitogen response, and the dysregulation of serum immunoglobulin levels are among immunological imbalances reported in ASD patients.3 Therefore, most of the genetic studies in ASD have focused on the immune system-related genes. The human leukocyte antigen (HLA) region on chromosome 6p21 codes for proteins, which participate in various biological functions including histocompatibility, inflammation, immune response regulation, and the complement cascade.4 Consequently, this region is of particular interest in the assessment of immune dysregulation detected in ASD patients. In addition, HLA proteins have been shown to be involved in neural cell communications, synaptic function, cerebral hemispheric specialization, as well as central nervous system (CNS) development.4 Certain HLA alleles have been shown to be associated with ASD. For instance, an association has been found between ASD and HLA-DRB1 alleles5 as well as complement C4 alleles.6 Furthermore, the null allele of the C4B gene, the extended haplotype B*44–C30–DRB*04, and the third hypervariable region (HVR-3) of certain DRBl alleles have been shown to be associated with ASD.7 Based on the importance of HLA alleles in the development of ASD, we designed a case–control study to assess the association between HLA class I and II alleles/haplotypes and ASD.
Participants and methods
Participants
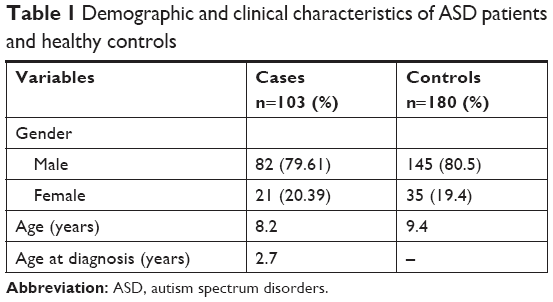
The current study has been designed as a case–control study to evaluate the association of HLA alleles and haplotypes with ASD in Iranian population. A total of 103 Iranian ASD patients and 180 age, gender, and healthy controls from the same ethnic group were enrolled in the study. Two clinical psychologists confirmed ASD diagnosis based on the criteria defined in the Diagnostic and Statistical Manual of Mental Disorders (DSM-V)8 and the Autism Diagnostic Inventory-Revised (ADI-R).9 Detailed neurological examination, medical history assessment, and genetic counseling were performed to exclude patients with any simultaneous disorder such as Fragile X syndrome, attention-deficit hyperactive disorder (ADHD), and epilepsy from the study. Controls were selected from volunteers in whom absence of neurological disorders was confirmed through obtaining medical history and physical examinations. Written informed consent forms were obtained from all participants of the study. In addition, for children aged below 16 years, an informed consent was obtained from their parents. The study was approved by the Shahid Beheshti University of Medical Sciences’ ethical committee. The research was conducted in accordance with the Declaration of Helsinki as revised in 1989.
Sample collection and DNA extraction
Saliva samples have been gathered from all participants as stated in a previous report.10 Genomic DNA was extracted from buccal epithelial cells in the mouthwash samples by using the GeneAll Exgene cell SVmini DNA kit (cat no 106-152). Afterward, we measured the purity and concentration of extracted DNA using the WPA Biowave II UV/Visible Spectrophotometer (serial no 80-3003-75) as stated in the manual.
HLA-A, -B, and -DRB genotyping
The low-resolution HLA-A, B, DR SSP kit (HLA-A-B-DR SSP Combi Tray; Olerup Diagnostic Gmbh, Mannheim, Germany) was used for HLA-DNA typing. This method applies amplification with sequence-specific primers, electrophoresis of the PCR products on 2.0% agarose gel stained with safety dye, and observation under UV transilluminator. The resolution of the method is comparable with serologic analysis of HLA-A, HLA-B, and HLA-DRB1 loci. The results are analyzed using the SCORE software arranged by the company. Besides, HLA-A/B/DRB1 haplotypes were determined according to the known linkage disequilibrium of HLA-A, -B, and -DRB1 alleles using an expectation–maximization (EM) algorithm in the R statistical computing environment.
Statistical analyses
Chi-squared test with Yates correction and Fisher’s exact test with Bonferroni correction were applied for the comparison of distribution of HLA-A, HLA-B, and HLA-DRB1 alleles and haplotypes between ASD patients and healthy controls. Besides, OR with 95% CIs was measured to identify the risks conferred by each haplotype. Statistical analyses were carried out using the SPSS (Released 2011, IBM SPSS Statistics for Windows, Version 20.0; IBM Corporation, Armonk, NY, USA). We manually counted all HLA genotypes to confirm the absence of any missing genotype. The level of significance was set at P-values ≤0.05.
Results
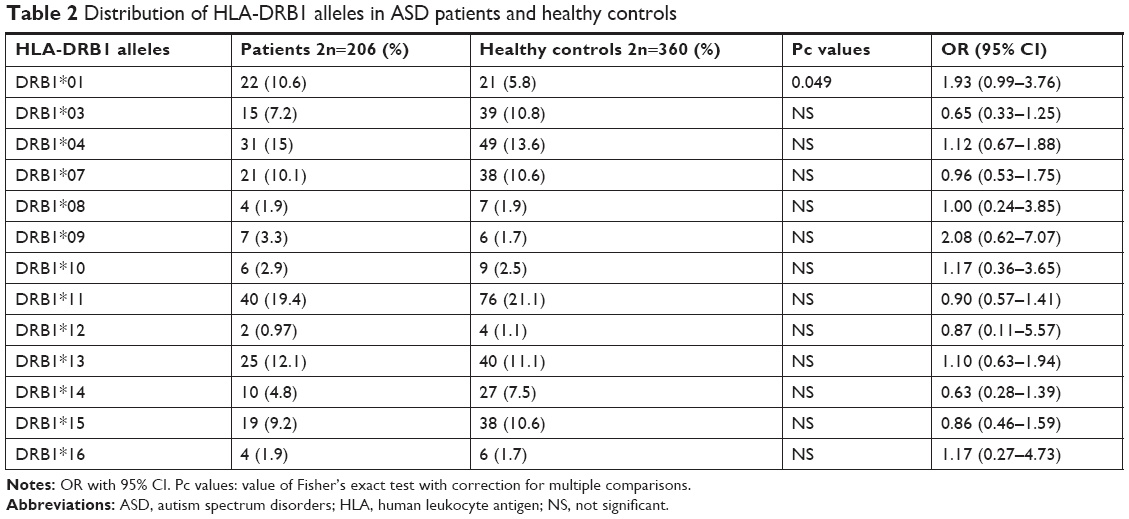
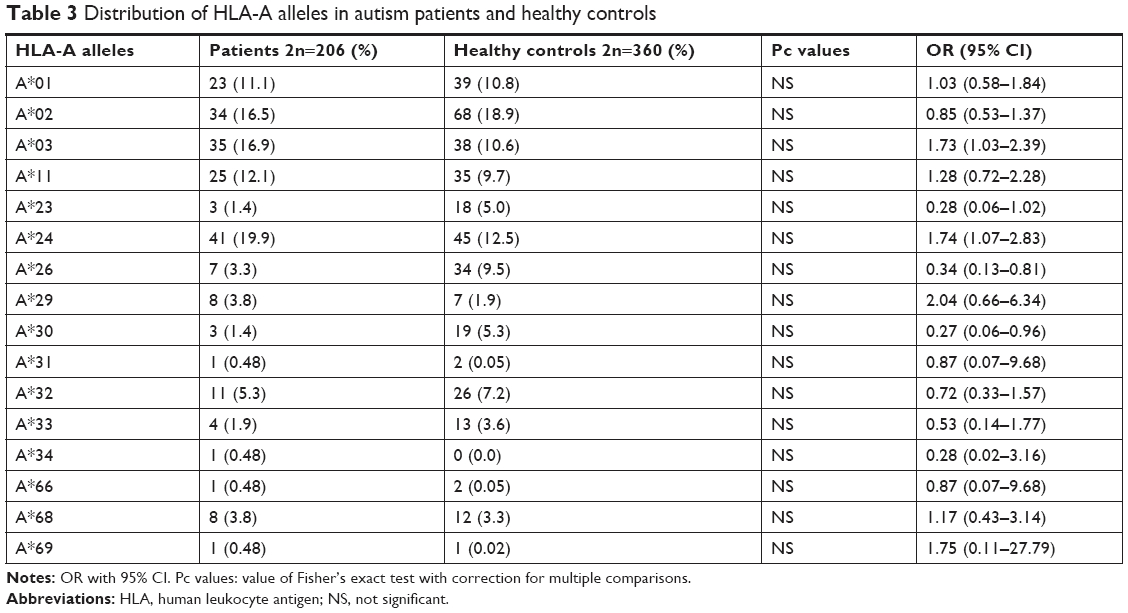
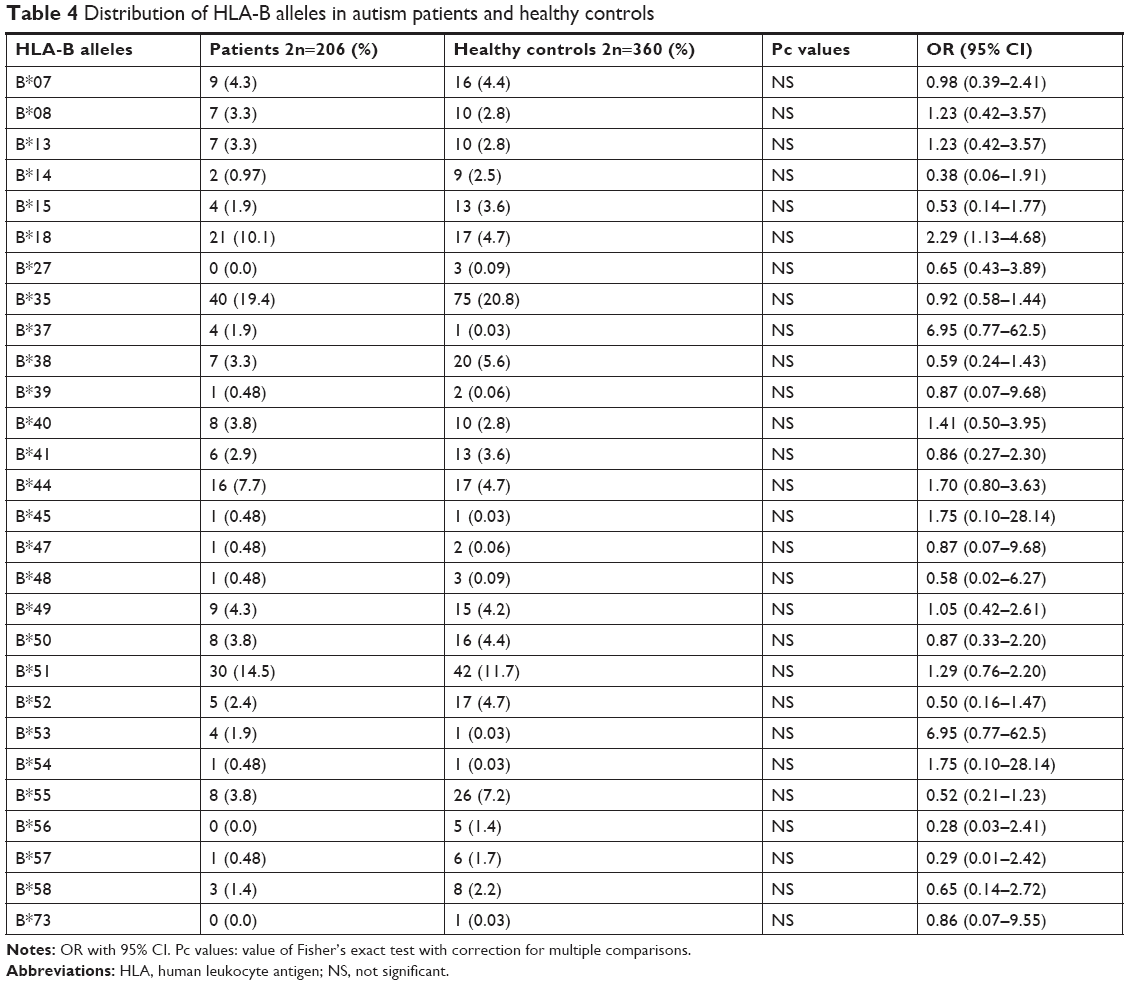
The values for age and sex of study participants as well as age at the diagnosis of ASD in patients are summarized in Table 1. Tables 2–4 show the frequencies of assessed HLA alleles at 2n level in ASD patients and healthy controls. Comparison of HLA-A, -B, and -DRB1 alleles’ frequencies between two groups of participants showed no significant difference in allele frequencies between patients and healthy controls. Table 5 shows distribution of the HLA-A/B/DRB1 haplotypes among both groups of the study. After Bonferroni correction, no haplotypes have been significantly more prevalent among patients compared with healthy subjects.
 | Table 1 Demographic and clinical characteristics of ASD patients and healthy controls |
 | Table 2 Distribution of HLA-DRB1 alleles in ASD patients and healthy controls |
 | Table 3 Distribution of HLA-A alleles in autism patients and healthy controls |
 | Table 4 Distribution of HLA-B alleles in autism patients and healthy controls |
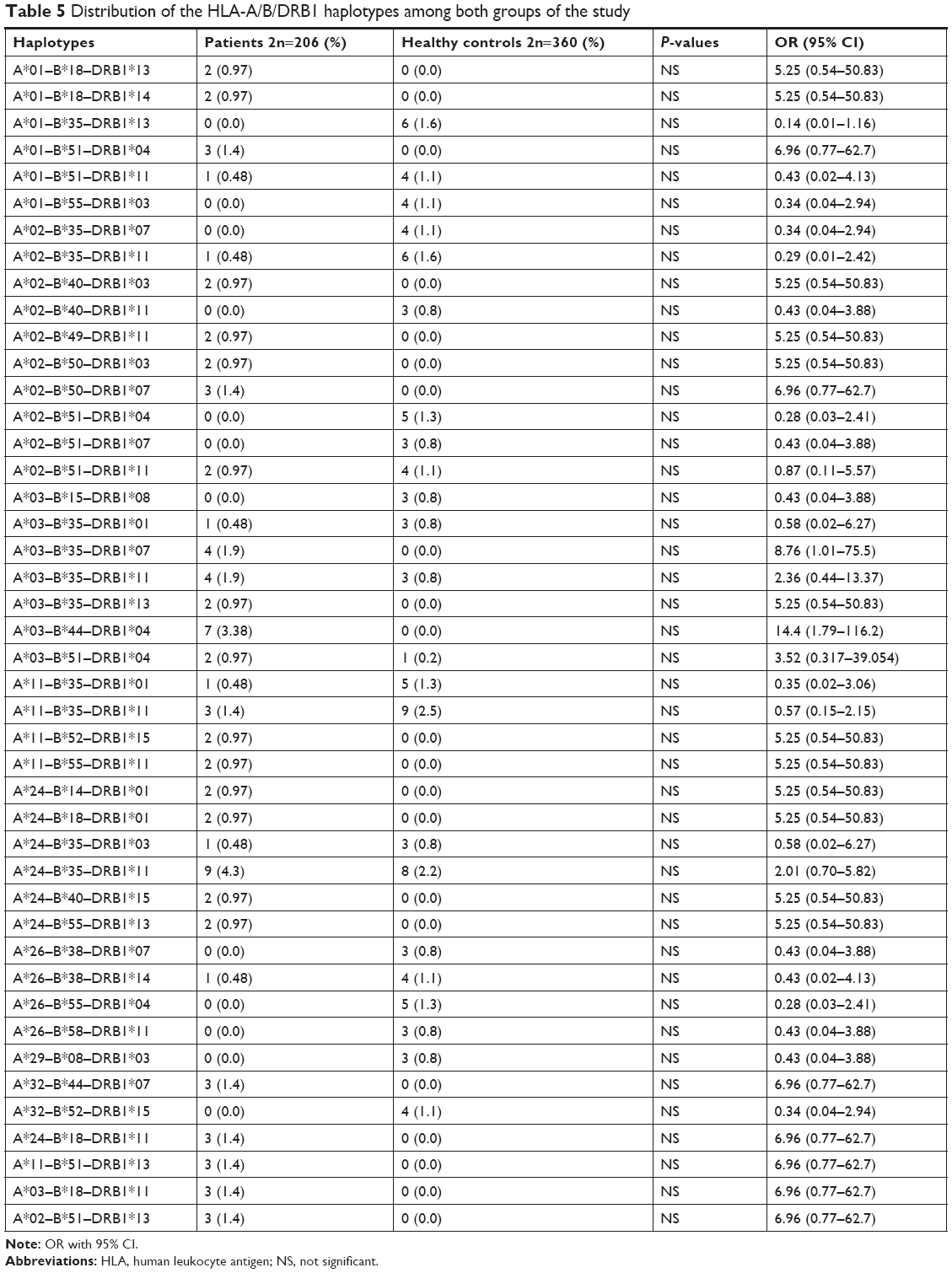 | Table 5 Distribution of the HLA-A/B/DRB1 haplotypes among both groups of the study |
Discussion
Although the exact underlying mechanism of ASD has not been clarified yet, genetic and environmental factors are supposed to participate in the onset of this disorder. In recent times, both genetic and environmental interdependent researches suggest a fundamental role for immune-related genes and immune responses to environmental stimuli in ASD pathogenesis.11 Class I HLA molecules in addition to their established role in immune response regulation are known to take part in neuronal signaling and activity-dependent changes in synaptic connectivity12 as well as neuronal plasticity,13 all of which supposed to be disrupted in ASD patients.14 Moreover, disturbances in HLA expression in the developing brain resulted from mutations and/or immune deregulation have been suggested to change the brain connectivity and function in an ASD distinctive manner.15
In the present study, we reported no significant differences in allele and haplotype frequencies of HLA-DRB1, HLA-A, and HLA-B between ASD patients and healthy subjects. Previous studies reported the existence of immune dysregulation in ASD patients. The occurrence of abnormal immune function during susceptible and critical course of neurodevelopment has been suggested as an underlying cause of neurological dysfunction detected in ASD.3 Such neurological dysfunction has strongly been associated with HLA alleles/haplotypes.4 Among HLA-DRB alleles, we only demonstrated over-presentation of DRB1*01 in ASD patients compared with controls but 95% CI values include OR =1, which disapproves the results. HLA-DRB1*01 has been previously shown to be associated with rheumatoid arthritis in Syrian population16 and with ASD in Macedonia.17 Although we could not detect any association between DRB1*04 allele and ASD, the role of this allele in ASD susceptibility has been shown in multiple studies. For instance, a previous study has demonstrated significant over-presentation of the DRB1*0401, DR01*0404, DRB1*0101, and DRB1*0701 alleles in ASD patients compared with normal subjects.7 Torres et al18 endorsed the association of the HLA-DRB1*04 allele with ASD and also discovered the lower frequency of DRB1*13 and DRB1*14 alleles in ASD patients compared with healthy subjects, which implies a possible protective effect. Associations between the DRB1*04 allele and ASD have been further approved in Lee et al19 and Johnson et al20 studies. More recently, HLA-DRB1 gene has been suggested to be associated with ASD in Han Chinese.21 Besides, DRB1*04, DRB1*11, and DRB1*14 have been shown to influence the intelligence and neuropsychology tests in ASD patients.21 Although we detected higher frequencies of A*03, A*24, and B*18 and lower frequencies of A*23, A*26, and A*30 in ASD patients compared with healthy controls, the results did not remain significant after multiple-comparison correction. HLA-A*24 and -B*18 have been previously shown to be susceptibility alleles for type 1 diabetes, which is considered as an autoimmune disease.22 There is no common allele between our study and Al-Hakbany et al’s11 study of ASD patients conducted in Saudi Arabia. They have shown significant association of HLA-A*01, HLA-A*02, HLA-B*07, and DRB1*1104 as well as haplotype A*02–B*07 with ASD. In addition, the A*01–B*07–DRB1*0701–DQB1*0602 and A*31–B*51–DRB1*0103–DQB1*0302 haplotypes were positively associated with ASD among Saudi patients.11 Another study conducted in Egypt has revealed higher frequency of HLA-DRB1*11 allele and lower frequency of HLA-DRB1*03 allele in ASD patients compared with controls.23 In the present study, although the frequency of DRB1*03 allele was lower in ASD patients than in controls, it did not reach the level of significance. Such inconsistency between the results of different studies regarding the HLA alleles and ASD has been noted previously,11 which might be explained by HLA allelic heterogeneity or distinctive underlying mechanisms for ASD in different populations, which might include the interaction between HLA allele and various infectious agents or environmental allergens in each area.11 The inconsistencies between our results and the previously conducted studies might be explained by differences in ethnic background, role of other genetic factors that alter HLA function, or existence of population-based environmental factors that modulate the effect of genetic factors.
Furthermore, we could not find significant association between any HLA haplotypes and ASD in our population. The haplotypes A*01–B*35–DRB1*13, A*02–B*50–DRB1*07, A*32–B*44–DRB1*07, A*24–B*18–DRB1*11, A*11–B*51–DRB1*13, A*03–B*18–DRB1*11, A*02–B*51–DRB1*13, and A*03–B*35–DRB1*07 were just seen in ASD patients and not in healthy controls. However, none of them reached the level of significance after multiple-comparison correction. Previously, one HLA extended haplotype containing the complement C4B null allele in the class III region, DRB1*04 in the class II region, and A*02 and B*44 alleles in the class I region has been shown to be associated with ASD in different populations.24–26
In brief, although HLA haplotypes might exert a pathogenic role in neurodevelopmental process in ASD patients, particularly after microbial infection, we could not suggest any HLA haplotypes as markers of genetic susceptibility to ASD in Iranian population. The specific function of HLA in ASD pathogenesis should be explored in future studies. Moreover, the function of the gene products should be considered in future studies, because even if there are no changes in the nucleotide sequences of genes, abnormal function of the proteins would still result in defects.
Besides, there are some limitations in our study including technical limitation (low-resolution HLA typing), lack of HLA-C and DQ typing, and relatively small size. Consequently, further researches are needed to approve these results and to draw a clear conclusion in this regard.
Disclosure
The authors report no conflicts of interest in this work.
References
Safari MR, Ghafouri-Fard S, Noroozi R, et al. FOXP3 gene variations and susceptibility to autism: A case-control study. Gene. 2017;596:119–122. | ||
Hamedani SY, Gharesouran J, Noroozi R, et al. Ras-like without CAAX 2 (RIT2): a susceptibility gene for autism spectrum disorder. Metab Brain Dis. 2017;32(3):751–755. | ||
Ashwood P, Wills S, Van de Water J. The immune response in autism: a new frontier for autism research. J Leukoc Biol. 2006;80(1):1–15. | ||
Torres AR, Westover JB, Rosenspire AJ. HLA immune function genes in Autism. Autism Res Treat. 2012;2012:959073–13. | ||
Polleux F, Lauder JM. Toward a developmental neurobiology of autism. Ment Retard Dev Disabil Res Rev. 2004;10(4):303–317. | ||
Burger RA, Warren RP. Possible immunogenetic basis for autism. Ment Retard Dev Disabil Res Rev. 1998;4(2):137–141. | ||
Warren RP, Odell JD, Warren WL, et al. Strong association of the third hypervariable region of HLA-DR beta 1 with autism. J Neuroimmunol. 1996;67(2):97–102. | ||
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®). American Psychiatric Pub; 2013. | ||
Rutter M, Le Couteur A, Lord C. Autism Diagnostic Interview-Revised. Vol. 29. Los Angeles, CA: Western Psychological Services; 2003:30. | ||
Noroozi R, Taheri M, Movafagh A, et al. Glutamate receptor, metabotropic 7 (GRM7) gene variations and susceptibility to autism: A case-control study. Autism Res. 2016;9(11):1161–1168. | ||
Al-Hakbany M, Awadallah S, Al-Ayadhi L. The Relationship of HLA Class I and II Alleles and Haplotypes with Autism: A Case Control Study. Autism Res Treat. 2014;2014(5):1–6. | ||
Corriveau RA, Huh GS, Shatz CJ. Regulation of class I MHC gene expression in the developing and mature CNS by neural activity. Neuron. 1998;21(3):505–520. | ||
Shatz CJ. MHC class I: an unexpected role in neuronal plasticity. Neuron. 2009;64(1):40–45. | ||
Bourgeron T. From the genetic architecture to synaptic plasticity in autism spectrum disorder. Nat Rev Neurosci. 2015;16(9):551–563. | ||
Needleman LA, McAllister AK. The major histocompatibility complex and autism spectrum disorder. Dev Neurobiol. 2012;72(10):1288–1301. | ||
Mourad J, Monem F. HLA-DRB1 allele association with rheumatoid arthritis susceptibility and severity in Syria. Rev Bras Reumatol. 2013;53(1):47–56. | ||
Trajkovski V, Spiroski M. DNA typing of HLA-A, -C, -B, and -DRB1 in the children with autism in the Republic of Macedonia. Bratisl Lek Listy. 2015;116(1):14–19. | ||
Torres AR, Maciulis A, Stubbs EG, Cutler A, Odell D. The transmission disequilibrium test suggests that HLA-DR4 and DR13 are linked to autism spectrum disorder. Hum Immunol. 2002;63(4):311–316. | ||
Lee LC, Zachary AA, Leffell MS, et al. HLA-DR4 in families with autism. Pediatr Neurol. 2006;35(5):303–307. | ||
Johnson WG, Buyske S, Mars AE, et al. HLA-DR4 as a risk allele for autism acting in mothers of probands possibly during pregnancy. Arch Pediatr Adolesc Med. 2009;163(6):542–546. | ||
Chien YL, Wu YY, Chen CH, et al. Association of HLA-DRB1 alleles and neuropsychological function in autism. Psychiatr Genet. 2012;22(1):46–49. | ||
Gough SC, Simmonds MJ. The HLA region and autoimmune disease: Associations and mechanisms of action. Curr Genomics. 2007;8(7):453–465. | ||
Mostafa GA, Shehab AA, Al-Ayadhi LY. The link between some alleles on human leukocyte antigen system and autism in children. J Neuroimmunol. 2013;255(1–2):70–74. | ||
Warren RP, Singh VK, Cole P, et al. Possible association of the extended MHC haplotype B44-SC30-DR4 with autism. Immunogenetics. 1992;36(4):203–207. | ||
Daniels WW, Warren RP, Odell JD, et al. Increased frequency of the extended or ancestral haplotype B44-SC30-DR4 in autism. Neuropsychobiology. 1995;32(3):120–123. | ||
Odell D, Maciulis A, Cutler A, et al. Confirmation of the association of the C4B null allelle in autism. Hum Immunol. 2005;66(2):140–145. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.