Back to Journals » Clinical Interventions in Aging » Volume 20
Association of Decay Accelerating Factor (CD55) Positive Extracellular Vesicles with Advanced Age and Blood Glucose Levels in Elderly Individuals
Authors Yue X, Liang S, Zhang H, Dai T, Wu J ![]()
Received 24 April 2025
Accepted for publication 3 September 2025
Published 12 September 2025 Volume 2025:20 Pages 1549—1560
DOI https://doi.org/10.2147/CIA.S536479
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Zhi-Ying Wu
Xinyang Yue,1,* Shuang Liang,2,* Haiyue Zhang,2 Tenglong Dai,1 Jun Wu1
1Thrombosis Research Center, Peking University Fourth School of Clinical Medicine, Beijing, People’s Republic of China; 2Thrombosis Research Center, Beijing Jishuitan Hospital Affiliated to Capital Medical University, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jun Wu, Thrombosis Research Center, Peking University Fourth School of Clinical Medicine, Beijing, People’s Republic of China, Email [email protected]
Background: Extracellular vesicles (EVs) play a pivotal role in driving aging, serving as vehicles for the transmission of aging-related signals. Decay-accelerating factors (DAFs), also known as CD55, a crucial complement regulator, exhibits age-associated alterations, yet its expression on circulating EVs remains poorly characterized. This study aims to investigate differences in plasma EVs and CD55 positive EVs concentrations in younger and older adults, and explore their correlations with age and blood glucose levels.
Methods: In this study, 40 older adults (≥ 65 years) undergoing routine physical examinations at Beijing Jishuitan Hospital and 40 gender-matched younger adults (< 65 years) were enrolled prospectively. Blood glucose levels were measured using the standard glucose oxidase method. Circulating EVs concentrations were quantified by flow cytometry, including circulating total EVs, endothelial EVs (EEVs), platelet EVs (PEVs), red blood cell EVs (REVs), lymphocyte EVs (LEVs), monocyte EVs (MEVs), neutrophil EVs (NEVs), white blood cell EVs (WEVs), and CD55-positive EVs.
Results: Compared to younger adults, older adults exhibited significantly lower concentrations of LEVs, MEVs, NEVs, and WEVs (p < 0.05). Concentrations of CD55 positive EVs, including CD55+EEVs, CD55+PEVs, CD55+LEVs, CD55+MEVs, CD55+NEVs, and CD55+WEVs, were markedly reduced in older adults (p < 0.05). Notably, CD55+PEVs showed the strongest negative correlation with age (r = - 0.6228, p < 0.001). Furthermore, significant inverse correlations were found between blood glucose levels and concentrations of CD55+EEVs (p = 0.001), CD55+PEVs (p = 0.006), CD55+LEVs (p < 0.001), CD55+MEVs (p = 0.001), and CD55+WEVs (p = 0.003).
Conclusion: Our findings indicate significant differences in plasma EVs subtypes between older and younger adults and demonstrate robust negative correlations between CD55+ EVs subtypes and both aging and elevated glucose levels. Thus, these results suggest CD55+ EVs as a critical contributor to metabolic disorders in aging, offering potential risk assessment and monitoring strategies in older populations.
Keywords: extracellular vesicles, decay accelerating factor, aging, flow cytometry, complement regulation
Introduction
The complement system plays a crucial role in both innate and adaptive immune responses. Disorders or dysfunctions of this system can have widespread effects, with one of the most notable changes being the inflammatory response.1 Complement regulatory factors are critical for maintaining the stability of the complement system. Research has shown that mutations in regulatory factors I and H result in secondary C3 deficiencies.2 Moreover, complement regulatory factors also play a significant role in aging and age-related diseases. A combination of locally synthesized complement regulatory factors (eg, FD, FHL-1) contributes to the onset of age-related macular degeneration (AMD).3 Recent studies have shown that complement-mediated synaptic pruning is a key factor in synaptic loss in Alzheimer’s disease (AD), with the deposition of complement effectors and regulatory proteins on synapses being a major contributing factor.4 The decay-accelerating factor (DAF), also known as CD55, is a critical membrane regulator of complement activation. CD55 was first isolated from human red blood cells by Hoffman in 1969.5 Since then, CD55 has been widely studied. CD55 has been associated with age-related diseases. Several studies have indicated that the activity of CD55 is significantly diminished with age,6,7 and this loss of function has been associated with diabetes.8 Moreover, gene mutations in the temporal cortex of the brain may alter the transcriptional activity of the CD55 gene, thereby impacting the complement system and potentially contributing to the development of AD.9 Another study found that the expression of CD55 in the exosome of astrocytes in AD was significantly reduced, leading to complement system dysfunction and neuronal damage.10 Therefore, CD55 may emerge as a potential target in the pathophysiological processes of aging and age-related diseases, offering new insights into the mechanisms underlying human aging.
Furthermore, Membrane-bound CD55 is considered the primary form of CD55,11 and its localization has been identified on the surface of extracellular vesicles (EVs).12
EVs are cell-derived membrane-bound structures, which can be classified into exosomes and microparticles based on their size.13 EVs are present in various bodily fluids, including blood, urine, and cerebrospinal fluid, and participate in a wide range of pathological and physiological processes. EVs primarily exert their effects by carrying proteins, lipids, and nucleic acids derived from parent cells, and also through proteins expressed on their membrane surface.14 The relationship between EVs and aging has been extensively explored. In an in vitro experiment, inducing aging in retinal pigment epithelial cell lines led to the release of a greater number and larger volumes of EVs, which transmitted aging signals to neighboring cells and may help elucidate the pathogenesis of AMD.15 Aging is typically accompanied by increased EVs release, and both EVs and the “cargo” they carry play important roles in the aging process. It has been reported that EVs released by senescent cells contain miRNAs that induce senescence in neighboring cells and inhibit their proliferation, thereby accelerating overall aging.16 EVs derived from healthy, non-aging cells have been shown to exert anti-aging effects, potentially offering therapeutic benefits.17 These vesicles can mitigate oxidative damage within the body, thereby contributing to cellular homeostasis and longevity.18 Conversely, EVs also exhibit pro-aging effects; Research has shown that EVs secreted by aging bone marrow-derived macrophages are implicated in transmitting aging-related signals throughout the organism. This process may play a pivotal role in inducing functional impairments commonly associated with aging.19 In addition, proteomic studies on EVs have shown that many proteins carried by EVs are significantly influenced by age, suggesting that EVs may serve as effective biomarkers for the aging process. Notably, some of these proteins are not affected by gender, indicating that certain EVs-associated proteins could act as universal markers of aging in both men and women.20
How age affects the release of circulating EVs and their subtypes remains unclear. Given the association of CD55 with age-related diseases and its expression on the surface of EVs, it is crucial to explore the role of age in modulating EVs and CD55-positive EVs under physiological conditions.
This study aimed to explore the impact of age on EVs release by enrolling two cohorts of healthy donors with a broad age range, revealing significant age-related differences in EVs subtypes and CD55-positive EVs subpopulations. We analyzed CD55 expression across various EVs subtypes and compared their concentrations between younger and older adult groups. These findings may provide insights into the mechanisms underlying immune dysfunction and complement regulation during aging, and serve as a basis for future diagnostic and therapeutic strategies.
Materials and Methods
Study Subjects and Blood Collection
In this study, we recruited 80 individuals who underwent physical examinations at Beijing Jishuitan Hospital between June and December 2024. The individuals were divided into two groups: the older adult group (age ≥ 65, n = 40) and the younger adult group (age < 65, n = 40). All individuals had generally normal laboratory test results and no history of surgery or major illnesses. The sample processing procedure referred to previously published methods.21,22 Briefly, peripheral venous blood samples were collected from these individuals and transferred into vacuum anticoagulant tubes containing 3.2% citrate (BD Biosciences, USA). The blood samples were centrifuged at 2500g for 15 minutes at room temperature (RT), repeated twice, and the upper plasma was transferred to 1.5mL Eppendorf tubes to obtain platelet-free plasma (PFP). The prepared PFP was stored in a −80°C freezer for subsequent detection of plasma EVs. Informed consent was obtained from all participants, and the study was approved by the Ethics Committee of Beijing Jishuitan Hospital [K2024531 - 00], strictly adhering to the Helsinki Declaration.
Reagents
Brilliant Violet 421™ anti-CD41 (Biolegend, 303730), PE-Cyanine7 anti-CD55 (Biolegend, 311314), APC-Fire™750 anti-CD45 (Biolegend, 304062), FITC anti-CD14 (Biolegend, 325604), PerCP-Cyanine5.5 anti-CD144 (Biolegend, 348510), APC-Cyanine7 anti-CD235a (Biolegend, 349116), Brilliant Violet 421™ anti-CD66b (Biolegend, 392916), PerCP-Cyanine5.5 anti-CD3 (Biolegend, 317336), Trucount Absolute Counting Tubes IVD (BD Biosciences, 340334, USA), Annexin V Binding Buffer (Abcam, ab14084, USA), SPHEROTM Flow Cytometry Nano Fluorescent Size Standard Kit (Spherotech, NFPPS – 52-4K).
Flow Cytometry
Circulating EVs were detected using flow cytometry with a BD FACSCanto II, following the established method in our laboratory.23 Briefly, using the SPHEROTM Flow Cytometry Nano Fluorescent Size Standard Kit, fluorescent beads of varying sizes (0.22μm - 1.33μm) were added to and mixed with 400μL PBS before being loaded onto the machine. The EVs gates were then determined using 0.22 μm and 0.88 μm fluorescent beads (Supplemental Figure 1). PFP was thawed in a 37°C water bath, and different subtypes of EVs were labeled. Each sample was divided into two tubes. In the first tube, 8 μL of PFP was added to an absolute counting tube, along with 1 μL of BV421 anti-CD41 antibody, PerCP Cy5.5 anti-CD144 antibody, APC Cy7 anti-CD235A antibody, and PE-Cy7 anti-CD55 antibody. The total volume of the solution was adjusted to 50 μL using 2 × Annexin V binding buffer. After shaking and mixing, the solution was incubated in the dark at RT for 20 minutes. After incubation, 250 μL of Annexin V binding buffer was added and incubated in the dark at RT for an additional 10 minutes. Subsequently, EVs detection was performed on the machine. CD41 positivity represents platelet-derived extracellular vesicles (PEVs), CD144 positivity represents endothelial cell-derived extracellular vesicles (EEVs), CD235a positivity represents red blood cell-derived extracellular vesicles (REVs), and double positivity for different subtypes of EVs and CD55 represents a subset of EVs that are CD55 positive.
For the second tube, the procedure for the second tube is similar to that of the first tube, with the only difference being the fluorescent antibodies used. Add 8 μL of PFP and 1 μL of BV421 anti-CD66b antibody, PerCP Cy5.5 anti-CD3 antibody, APC-Fire™750 anti-CD45 antibody, FITC anti-CD14 antibody, and PE-Cy7 anti-CD55 antibody to an absolute counting tube. Adjust the total volume of the solution to 50 μL using 2 × Annexin V binding buffer, shake well, and incubate in the dark at RT for 20 minutes. After incubation is complete, 250 μL of Annexin V binding buffer was added and incubated in the dark at RT for 10 minutes. Subsequently, EVs detection was performed on the machine. CD66b positivity represents neutrophil-derived extracellular vesicles (NEVs), CD3 positivity represents lymphocyte-derived extracellular vesicles (LEVs), CD14 positivity represents monocyte-derived extracellular vesicles (MEVs), CD45 positivity represents white blood cell-derived (WEVs) (Supplemental Figure 2).
The circulating EVs concentration was determined according to the formula: events/µL = (Positive events × Counting bead concentration) / (PFP volume × Number of Counting Beads).
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 9 and IBM SPSS 24 (SPSS Inc., USA) software. Non-parametric tests were used to evaluate differences in hematological parameters and EVs concentrations between groups, with data presented as median (25th, 75th percentile) and analyzed using the Mann–Whitney U-test. Spearman correlation analysis was used to assess the correlation between CD55-positive EVs and age and blood glucose concentration. A P-value of < 0.05 was considered statistically significant for all data.
Results
Basic Characteristics and Clinical Parameters in the Older Adult and Younger Adult Groups
The demographic and basic laboratory parameters of all study subjects are shown in Table 1. The gender distribution between the two groups was comparable. The platelet count in the older adult group was significantly lower than that in the younger adult group [216 (191–249) vs 252 (224–282) ×109/L, p < 0.001]. The platelet crit in the older adult group was lower than that in the younger adult group [0.22 (0.19–0.25) vs 0.26 (0.22–0.28) %, p < 0.001]. Furthermore, the red blood cell count [4.62 (4.27–4.92) vs 4.73 (4.48–5.21) ×1012/L, p = 0.049] and lymphocyte count [1.79 (1.56–2.11) vs 1.99 (1.73–2.24) ×109/L, p = 0.044] were significantly lower in the older adult group compared to the younger adult group. When comparing coagulation parameters between the two groups, D-dimer [0.33 (0.24–0.43) vs 0.22 (0.14–0.32) mg/l, p = 0.001] and fibrin degradation products (FDP) [1.92 (1.6–2.9) vs 1.39 (1.1–1.9) μg/mL, p < 0.001] were significantly higher in the older adult group compared to the younger adult group, whereas activated partial thromboplastin time (APTT) in the older adult group was lower than that in the younger adult group [26.6 (25.6–27.2) vs 27.8 (26.5–28.8) S, p < 0.001]. We also compared blood glucose levels between the older adult and younger adult groups. The results indicated that blood glucose was higher in the older adult group than in the younger adult group [5.5 (5.1–5.8) vs 5 (4.7–5.3) mmol/l, p < 0.001], with a statistically significant difference.
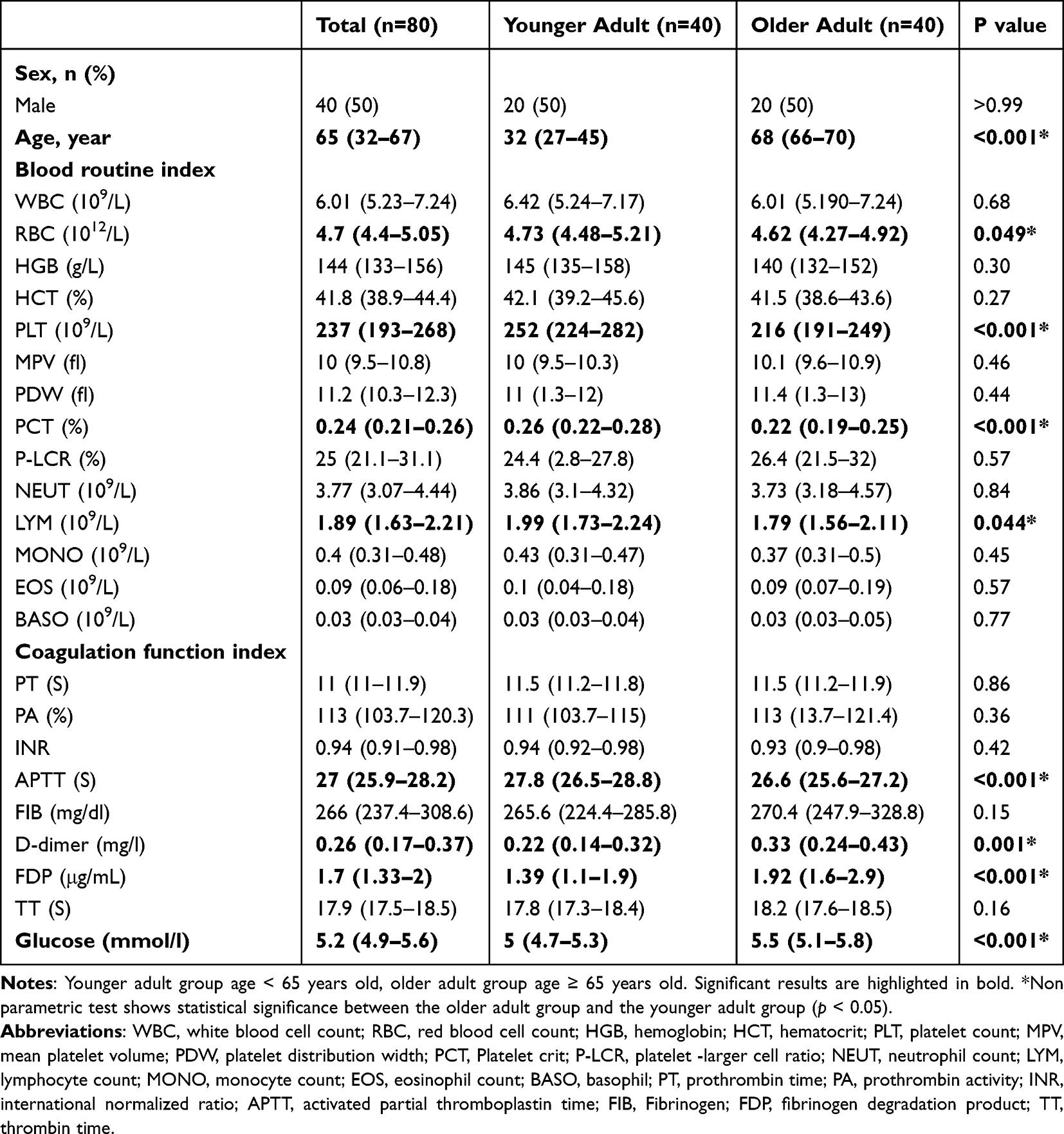 |
Table 1 Basic Clinical Parameters of the Older Adult and Younger Adult |
Concentration of Circulating EVs in Older Adult and Younger Adult Groups
We first measured the concentrations of circulating total EVs and several EVs subtypes, including EEVs, PEVs, REVs, LEVs, MEVs, NEVs, and WEVs, in the older adult and younger adult groups using flow cytometry (Figure 1). Our data indicate that the total circulating EVs concentration in the older adult group is higher than in the younger adult group (Figure 1A), though the difference is not statistically significant. The concentration of EEVs in the older adult group was lower than in the younger adult group, but the difference was not statistically significant (Figure 1B).
 |
Figure 1 Evaluate the differences in circulating EVs and their subtypes concentrations between two groups using flow cytometry. (A) EVs. (B) EEVs. (C) PEVs (p = 0.02). (D) REVs. (E) LEVs (p < 0.001). (F) MEVs (p = 0.02). (G) NEVs (p = 0.002). (H) WEVs. (p = 0.001). *(p < 0.05); **(p < 0.01); ***(p < 0.001). p < 0.05 is statistically significant. The data is presented as the medians and (25th, 75th percentile). Younger adult group age < 65 years old, older adult group age ≥ 65 years old. Abbreviations: EVs, extracellular vesicles; EEVs, endothelial cell derived extracellular vesicles; PEVs, platelet derived extracellular vesicles; REVs, red blood cell derived extracellular vesicles; LEVs, lymphocyte derived extracellular vesicles; MEVs, monocyte derived extracellular vesicles; NEVs, neutrophil derived extracellular vesicles; WEVs, white blood cell derived extracellular vesicles. |
Interestingly, the concentration of PEVs was significantly higher in the older adult group than in the younger adult group (Figure 1C), despite significantly lower platelet counts in the older adult group (Table 1), with the difference being statistically significant (p = 0.02). Similarly, the concentration of REVs in the older adult group was lower than that in the younger adult group, without a statistically significant difference (Figure 1D). Among all EVs subtypes, LEVs showed the most significant reduction in the older adult group compared with the younger adult group (p < 0.001, Figure 1E). MEVs (p = 0.02, Figure 1F), NEVs (p = 0.002, Figure 1G), and WEVs (p = 0.001, Figure 1H) concentrations were also significantly lower in the older adult group than in the younger adult group. These data suggest that age has a greater impact on EVs derived from immune cells.
Concentration of Circulating CD55 Positive EVs in Elder and Younger Adult Groups
We next analyzed the concentrations of circulating total CD55+EVs and their subtypes in both the younger adult and older adult groups using flow cytometry, as shown in Figure 2. The total concentration of CD55+EVs was significantly higher in the younger adult group than in the older adult group (p = 0.02, Figure 2A), suggesting a general age-associated decline in CD55+EVs levels. For endothelial-derived EVs, the concentration of CD55+EEVs was markedly higher in the younger adult group than in the older adult group (p < 0.001, Figure 2B). In contrast to the total PEVs trend observed earlier, the concentration of CD55+PEVs was significantly higher in the younger adult group (p < 0.001, Figure 2C), indicating a possible reduction in CD55 expression on platelet-derived EVs with age. Moreover, the concentration of CD55+REVs was lower in the older adult group than in the younger adult group, but the difference was not statistically significant (Figure 2D). And the CD55+LEVs showed a significantly higher concentration in the younger adult group (p < 0.001, Figure 2E), indicating that aging may diminish CD55+LEVs release or expression. Similarly, the concentration of CD55+MEVs were significantly higher in the younger adult group compared to the older adult group (p < 0.001, Figure 2F). No statistically significant difference was observed in CD55+NEVs levels between the two groups (Figure 2G). Finally, CD55+WEVs were significantly elevated in the younger adult group (p < 0.001, Figure 2H), consistent with the trend observed across most CD55 positive immune cell–derived EVs.
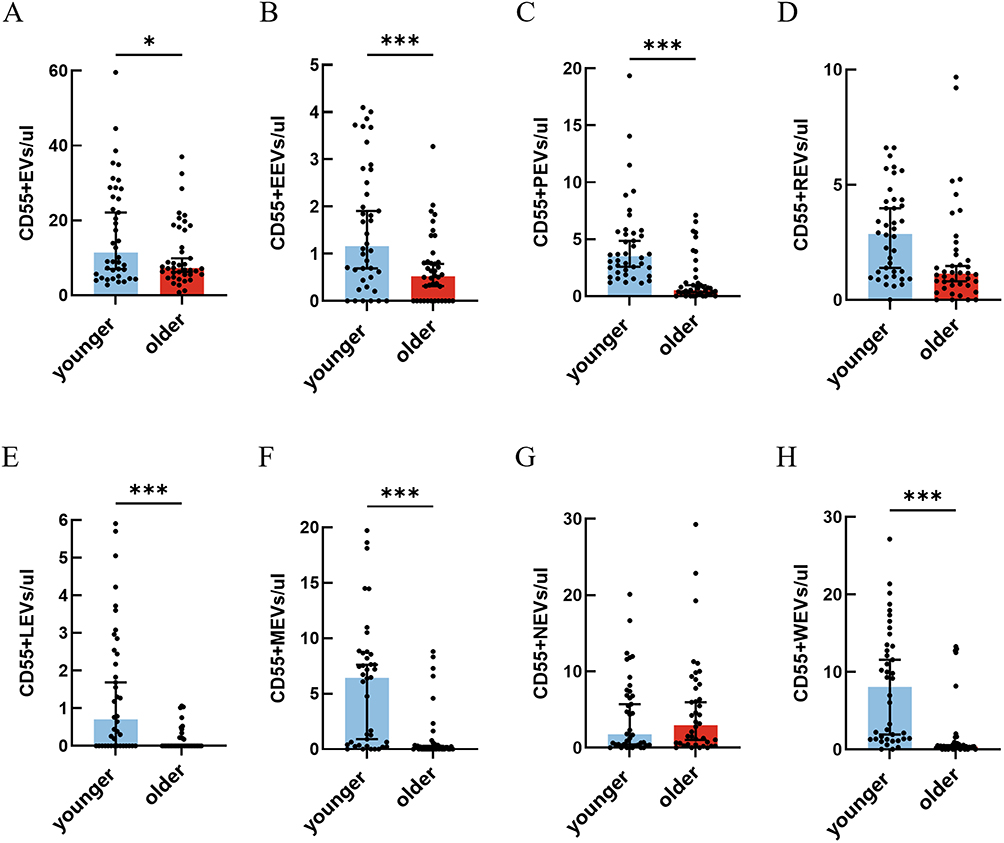 |
Figure 2 Evaluate the differences in circulating CD55 positive EVs and their subtypes concentrations between two groups using flow cytometry. (A) CD55+EVs (p = 0.02). (B) CD55+EEVs (p < 0.001). (C) CD55+PEVs (p < 0.001). (D) CD55+REVs. (E) CD55+LEVs (p < 0.001). (F) CD55+MEVs (p < 0.001). (G) CD55+NEVs. (H) CD55+WEVs (p < 0.001). *(p < 0.05); ***(p < 0.001). p < 0.05 is statistically significant. The data is presented as the medians and (25th, 75th percentile). Younger adult group age < 65 years old, older adult group age ≥ 65 years old. Abbreviations: CD55, decay-accelerating factor (also known CD55); EVs, extracellular vesicles; EEVs, endothelial cell derived extracellular vesicles; PEVs, platelet derived extracellular vesicles; REVs, red blood cell derived extracellular vesicles; LEVs, lymphocyte derived extracellular vesicles; MEVs, monocyte derived extracellular vesicles; NEVs, neutrophil derived extracellular vesicles; WEVs, white blood cell derived extracellular vesicles. |
Correlation Between CD55 Positive EVs and Age
Several subtypes of CD55+EVs exhibited a negative correlation with age in the enrolled population (Figure 3). Specifically, the concentration of CD55+EEVs was negatively correlated with age (r = –0.3176, p = 0.004, Figure 3A). CD55+PEVs displayed the strongest negative correlation among all subtypes (r = –0.6228, p < 0.001, Figure 3B). Similarly, CD55+LEVs (r = –0.3324, p = 0.003, Figure 3C), CD55+MEVs (r = –0.3916, p < 0.001, Figure 3D), and CD55+WEVs (r = –0.4805, p < 0.001, Figure 3E) also demonstrated significant negative correlations with age. In contrast, no significant age-related correlation was observed for CD55+REVs, CD55+NEVs, or the total CD55+EVs population (data not shown).
 |
Figure 3 The relationship between age and CD55 positive EVs. (A) CD55+EEVs was negatively correlated with age (p = 0.004, r = −0.3176). (B) CD55+PEVs was negatively correlated with age (p < 0.001, r = −0.6228). (C) CD55+LEVs was negatively correlated with age (p = 0.003, r = −0.3324). (D) CD55+MEVs was negatively correlated with age (p < 0.001, r = −0.3916). (E) CD55+MEVs was negatively correlated with age (p < 0.001, r = −0.4805). Abbreviations: CD55, decay-accelerating factor (also known CD55); EEVs, endothelial cell-derived extracellular vesicles; PEVs, platelet derived extracellular vesicles; LEVs, lymphocyte derived extracellular vesicles; MEVs, monocyte derived extracellular vesicles; WEVs, white blood cell derived extracellular vesicles. |
Correlation Between CD55 Positive EVs and Glucose
Besides age, we also analyzed the relationship between CD55 positive EVs and blood glucose (Figure 4), as increasing evidence suggests that CD55 is downregulated in the pathogenesis of diabetes.24 Our data indicate that as the concentration of CD55 positive EVs increases, blood glucose levels tend to decrease. A negative correlation was found between blood glucose concentration and CD55 positive EVs concentration. The correlations between CD55+EVs, CD55+REVs, and CD55+NEVs with blood glucose were not statistically significant (data not shown). Additionally, CD55+EEVs (r = –0.3731, p = 0.001, Figure 4A), CD55+PEVs (r = –0.3246, p = 0.006, Figure 4B), CD55+LEVs (r = –0.4092, p < 0.001, Figure 4C), CD55+MEVs (r = –0.3786, p = 0.001, Figure 4D), and CD55+WEVs (r = –0.3474, p = 0.003, Figure 4E) exhibited a strong negative correlation with blood glucose concentration.
 |
Figure 4 The relationship between GLU and CD55 positive EVs. (A) CD55+EEVs was negatively correlated with GLU (p = 0.001, r = −0.3731). (B) CD55+PEVs was negatively correlated with GLU (p = 0.006, r = −0.3246). C. CD55+LEVs was negatively correlated with GLU (p < 0.001, r = −0.4092). (D) CD55+MEVs was negatively correlated with GLU (p = 0.001, r = −0.3786). (E) CD55+WEVs was negatively correlated with GLU (p = 0.003, r = −0.3474). Abbreviations: CD55, decay-accelerating factor (also known CD55); EEVs, endothelial cell-derived extracellular vesicles; PEVs, platelet derived extracellular vesicles; LEVs, lymphocyte derived extracellular vesicles; MEVs, monocyte derived extracellular vesicles; WEVs, white blood cell derived extracellular vesicles; GLU, glucose. |
Discussion
Our data indicate differences in circulating EVs and their subtypes between older adult and younger adult individuals. Our data show a decreasing trend in the concentration of EVs in the older adult population. As shown in Figures 1 and 2, only the concentrations of circulating total EVs, PEVs, and CD55+NEVs were higher in the older adult group than in the younger adult group, while the concentrations of other EVs were higher in the younger adult group. We further analyzed the correlation between EVs concentrations and age, as well as EVs and blood glucose. As age increases, the concentration of some CD55+EVs decreases, accompanied by an increase in blood glucose levels, suggesting that CD55+EVs may play a role in the pathogenesis of diabetes in the older adult.
Aging leads to numerous changes in many components of the immune system, a phenomenon known as “immunosenescence.” Indeed, older adult individuals exhibit increased susceptibility to various diseases, including cancer, autoimmune disorders, and other chronic inflammatory conditions.25 This is due to a specific series of events, including genomic instability, epigenetic alterations, mitochondrial dysfunction, stem cell exhaustion, and cellular senescence, all of which affect cellular physiology.26 Senescent cells in the aging microenvironment secrete factors known as the senescence-associated secretory phenotype (SASP). SASP plays a dual role in the aging microenvironment. It not only recruits immune cells to fight bacterial or viral invasions, but also induces the transmission of senescent signals, contributing to inflammation.27 EVs, as components of the SASP, play a significant role in the senescent microenvironment. DNA damage response (DDR) is the primary factor driving changes in EVs within the senescent microenvironment. During DDR, EVs production depends on functional p53 and p65. Inhibition of p53 can reduce the secretion of CD63+EVs, which play a key role in cell senescence and apoptosis. Inhibition of p65 can also prevent the transmission of senescent signals between mesenchymal stem cells via EVs.28,29 The senescent microenvironment affects EVs production, and the EVs produced can, in turn, influence the pathological state of the senescent microenvironment. The impact of aging on EVs is also reflected in their “cargo”. The heterogeneity of EVs cargo determines the diversity of EVs functions in the aging microenvironment, enabling EVs to carry specific markers, proteins, lipids, noncoding RNAs, and to up- or downregulate the expression of these molecules during aging.30,31
Moreover, the effects of senescence on cell metabolism, intercellular communication, and immune system function have been confirmed in various studies. The major aspects of aging include persistent low-grade inflammation, decreased ability to fight infection or cancer, impaired responses to new antigens, increased incidence of autoimmunity, and impaired wound repair.32 The decrease in CD55+EVs in the older adult population may contribute to these age-related changes. The reduction in CD55 levels and the diminished antagonistic effect on the complement pathway greatly increase the older adult’s susceptibility to diseases. Moreover, it has been found that EVs purified from patients with Parkinson’s disease are rich in markers of systemic inflammation, such as CRP and TNF-α. The reduction in CD55+EVs in the older adult may lead to a loss of anti-inflammatory function, which could contribute to the pro-inflammatory EVs phenotype observed in Parkinson’s disease patients.33 A decrease in regenerative capacity is also a hallmark of aging.32 Although the expected cellular senescence trajectory involves cell elimination, excessive accumulation of senescent cells in older adult individuals disrupts the programmed cell death pathway, resulting in a decline in regenerative capacity.34 The weakened regenerative capacity in this environment leads to the downregulation of certain molecules in the SASP. Thus, it is unsurprising that the number of EVs decreases in the aging microenvironment, as observed in our experiments, where both the total EVs and CD55+EVs involved in anti-inflammatory responses were reduced in the older adult population. Moreover, as reported in most studies, both the number of EVs and the contents they carry tend to decrease with age. Natha et al found that in aging fibroblast-derived EVs, half to two-thirds of the proteins were downregulated.35 However, Rosalia et al reported opposite findings, showing that the number of EVs in the follicular fluid of older women was higher than that in younger women.36 This discrepancy may be due to the organ-specific nature of follicular fluid. By contrast, EVs in plasma are derived from a wide range of organs and cell types, which may provide a broader perspective on age-related changes in the EVs profile across the entire organism.
The latest evidence indicates that EVs play a critical role in the pathophysiological mechanisms of diabetes and its complications.37 CD55 is a complement regulatory protein that has been linked to the progression of diabetes. B. Aydin Özgur et al investigated CD55 expression on the surface of lymphocytes in patients with diabetes and found that CD55 expression was significantly lower in patients with diabetic nephropathy, retinopathy, and cardiovascular disease compared to healthy controls.38 Similarly, MA Xiwen et al reported a significant reduction in CD55 expression on the surface of leukocytes in diabetes patients, especially those with macroangiopathy.39 However, most previous studies have focused on CD55 expression on the cell surface, rather than on the surface of EVs. Our study helps to fill this gap and contributes to a deeper understanding of the potential role of CD55 in glucose metabolism.
In our study, we measured CD55 expression on the surface of circulating EVs and compared the differences between older adult individuals and younger adult individuals. We observed a negative correlation between CD55-positive EVs and blood glucose levels. The decrease in CD55-positive EVs corresponded with an increase in blood glucose concentration, which parallels the reduction of CD55 expression on the cell surface in diabetes patients. These findings provide valuable insight into the pathophysiological mechanisms of diabetes. Future research should further explore the CD55-positive EVs in controlling blood glucose and whether it can become a new therapeutic target in diabetes.
Conclusion
Our cohort study revealed a significant reduction in the concentrations of EVs subtypes in the plasma of older adults compared to younger adults, highlighting potential age-related changes in the circulating EVs profile. These findings may indicate age-related functional changes in EVs and reflect alterations in cellular status and the microenvironment during the aging process. Importantly, we identified a negative correlation between CD55-positive EVs and blood glucose concentrations, suggesting a potential link between these EVs and glucose metabolism. Dysregulation of CD55’s complement regulatory function may impair the body’s regulation of blood glucose by increasing the vulnerability of pancreatic β cells to complement-mediated damage. Our observation offers a new perspective on the pathophysiological mechanisms of metabolic diseases, especially diabetes.
Furthermore, expanding the sample size and conducting multicenter studies will help to further validate and strengthen our conclusions. Meanwhile, investigating other bioactive components carried by EVs, such as nucleic acids and lipids, may provide a more comprehensive understanding of their roles in aging. This study lays the groundwork for exploring the potential involvement of CD55-positive EVs in age-related diseases such as Alzheimer’s and Parkinson’s disease. Future research may further elucidate their diagnostic and therapeutic value in the aging process.
Abbreviations
AD, Alzheimer’s disease; AMD, Age-related macular degeneration; APTT, Activated partial thromboplastin time; BASO, Basophils; DAFs, Decay-accelerating factors; DDR, DNA damage response; EEVs, Endothelial extracellular vesicles; EOS, Eosinophils; EVs, Extracellular vesicles; FDP, Fibrin degradation products; FIB, Fibrinogen; GLU, Glucose; HCT, Hematocrit; HGB, Hemoglobin; INR, International normalized ratio; LEVs, Lymphocyte extracellular vesicles; LYM, Lymphocytes; MEVs, Monocyte extracellular vesicles; MONO, Monocytes; MPV, Mean platelet volume; NEUT, Neutrophils; NEVs, Neutrophil extracellular vesicles; PA, Procoagulant activity; PCT, Platelet crit; PEVs, Platelet extracellular vesicles; PFP, Platelet-free plasma; PLT, Platelet count; P-LCR, Platelet-large cell ratio; PT, Prothrombin time; RBC, Red blood cells; REVs, Red blood cell extracellular vesicles; RT, Room temperature; SASP, Senescence-associated secretory phenotype; TT, Thrombin time; WBC, White blood cells; WEVs, White blood cell extracellular vesicles.
Data Sharing Statement
Data are available from the corresponding author upon request and with appropriate authorization.
Ethics Approval and Consent to Participate
The study protocol was approved by Ethics Committee of Beijing Jishuitan Hospital, Capital Medical University [K2024531 – 00].
Author Contributions
Xinyang Yue and Shuang Liang should be regarded as co-first authors. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by Beijing Municipal Natural Science Foundation (L246006), Beijing Natural Science Foundation (L254086), Beijing Municipal Health Commission High-Level Public Health Technical Talent Development Project, Natural Science Foundation Cultivation Project (ZR-202412) of Beijing Jishuitan Hospital Affiliated to Capital Medical University and Wu Jieping Clinical Research Fund.
Disclosure
The authors declare no competing interests.
References
1. Conigliaro P, Triggianese P, Ballanti E. et al. Complement, infection, and autoimmunity. Curr Opin Rheumatol. 2019;31(5):532–541. doi:10.1097/bor.0000000000000633
2. Falcão DA, Isaac L, Isaac L. Clinical aspects and molecular basis of primary deficiencies of complement component C3 and its regulatory proteins factor I and factor H. Scand J Immunol. 2006;63(3):155–168. doi:10.1111/j.1365-3083.2006.01729.x
3. Armento A, Ueffing M, Clark SJ. The complement system in age-related macular degeneration. Cell Mol Life Sci. 2021;78(10):4487–4505. doi:10.1007/s00018-021-03796-9
4. Wen L, Bi D, Shen Y. Complement-mediated synapse loss in Alzheimer’s disease: mechanisms and involvement of risk factors. Trends Neurosci. 2024;47(2):135–149. doi:10.1016/j.tins.2023.11.010
5. Hoffman EM. Inhibition of complement by a substance isolated from human erythrocytes. I. Extraction from human erythrocyte stromata. Immunochemistry. 1969;6(3):391–403. doi:10.1016/0019-2791(69)90296-1
6. Gwamaka M, Fried M, Domingo G, Duffy PE. Early and extensive CD55 loss from red blood cells supports a causal role in malarial anaemia. Malar J. 2011;10(1):386. doi:10.1186/1475-2875-10-386
7. Fuchino Y, Okada N, Funabashi K, et al. Expression of decay-accelerating factor on CD8-positive lymphocytes as an index of aging and of host defense function. Nihon Ronen Igakkai Zasshi. 1996;33(8):580–585. doi:10.3143/geriatrics.33.580
8. Moin ASM, Nandakumar M, Diboun I, et al. Hypoglycemia-induced changes in complement pathways in type 2 diabetes. Atheroscler Plus. 2021;46:35–45. doi:10.1016/j.athplu.2021.11.002
9. Helgadottir HT, Lundin P, Wallén Arzt E, Lindström AK, Graff C, Eriksson M. Somatic mutation that affects transcription factor binding upstream of CD55 in the temporal cortex of a late-onset Alzheimer disease patient. Hum Mol Genet. 2019;28(16):2675–2685. doi:10.1093/hmg/ddz085
10. Goetzl EJ, Schwartz JB, Abner EL, Jicha GA, Kapogiannis D. High complement levels in astrocyte-derived exosomes of Alzheimer disease. Ann Neurol. 2018;83(3):544–552. doi:10.1002/ana.25172
11. Vainer ED, Meir K, Furman M, Semenenko I, Konikoff F, Vainer GW. Characterization of novel CD55 isoforms expression in normal and neoplastic tissues. Tissue Antigens. 2013;82(1):26–34. doi:10.1111/tan.12138
12. Brennan K, Iversen KF, Blanco-Fernández A, Lund T, Plesner T, Mc Gee MM. Extracellular Vesicles Isolated from Plasma of Multiple Myeloma Patients Treated with Daratumumab Express CD38, PD-L1, and the Complement Inhibitory Proteins CD55 and CD59. Cells. 2022;11(21):3365. doi:10.3390/cells11213365
13. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
14. Urabe F, Kosaka N, Ito K, Kimura T, Egawa S, Ochiya T. Extracellular vesicles as biomarkers and therapeutic targets for cancer. Am J Physiol Cell Physiol. 2020;318(1):C29–c39. doi:10.1152/ajpcell.00280.2019
15. Fernandes R, Correia D, Martins B, Malva J, Girão H. Modeling senescence in ARPE19 cells to investigate the role of small extracellular vesicles in age-related macular degeneration. Acta Ophthalmologica. 2024;103:17344. doi:10.1111/aos.17344
16. Ganesh BH, Padinjarathil H, Rajendran RL, Ramani P, Gangadaran P, Ahn B-C. The Role of Extracellular Vesicles in Aging and Age-Related Disorders. Antioxidants. 2025;14(2):177. doi:10.3390/antiox14020177
17. Mas-Bargues C, Sanz-Ros J, Román-Domínguez A, et al. Extracellular Vesicles from Healthy Cells Improves Cell Function and Stemness in Premature Senescent Stem Cells by miR-302b and HIF-1α Activation. Biomolecules. 2020;10(6):957. doi:10.3390/biom10060957
18. Sanz-Ros J, Mas-Bargues C, Romero-García N, Huete-Acevedo J, Dromant M, Borrás C. Therapeutic Potential of Extracellular Vesicles in Aging and Age-Related Diseases. Int J Mol Sci. 2022;23(23):14632. doi:10.3390/ijms232314632
19. Hou J, Chen KX, He C, et al. Aged bone marrow macrophages drive systemic aging and age-related dysfunction via extracellular vesicle-mediated induction of paracrine senescence. Nat Aging. 2024;4(11):1562–1581. doi:10.1038/s43587-024-00694-0
20. Huang Y, Feng J, Xu J, et al. Associations of age and sex with characteristics of extracellular vesicles and protein-enriched fractions of blood plasma. Aging Cell. 2025;24(1). doi:10.1111/acel.14356
21. Chen Y, Liu J, Su Y, et al. Annexin V and tissue factor+microparticles as biomarkers for predicting deep vein thrombosis in patients after joint arthroplasty. Clin. Chim. Acta. 2022;536:169–179. doi:10.1016/j.cca.2022.09.011
22. Zhang W, Su Y, Liu L, et al. Fibrinolysis Index as a new predictor of deep vein thrombosis after traumatic lower extremity fractures. Clinica Chimica Act. 2020;511:1. doi:10.1016/j.cca.2020.10.018
23. Zang X, Su Y, Zhang W, et al. Hepatocyte-derived Microparticles as Novel Biomarkers for the Diagnosis of Deep Venous Thrombosis in Trauma Patients. Clin Appl Thromb Hemost. 2023;29:10760296231153400. doi:10.1177/10760296231153400
24. Arredondo-Damián JG, Martínez-Soto JM, Molina-Pelayo FA, et al. Systematic review and bioinformatics analysis of plasma and serum extracellular vesicles proteome in type 2 diabetes. Heliyon. 2024;10(3):e25537. doi:10.1016/j.heliyon.2024.e25537
25. Ikeda H, Togashi Y. Aging, cancer, and antitumor immunity. Int J Clin Oncol. 2022;27(2):316–322. doi:10.1007/s10147-021-01913-z
26. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–1217. doi:10.1016/j.cell.2013.05.039
27. Ovadya Y, Landsberger T, Leins H, et al. Impaired immune surveillance accelerates accumulation of senescent cells and aging. Nat Commun. 2018;9(1):5435. doi:10.1038/s41467-018-07825-3
28. Tesei A, Arienti C, Bossi G, et al. TP53 drives abscopal effect by secretion of senescence-associated molecular signals in non-small cell lung cancer. J Exp Clin Cancer Res. 2021;40(1):89. doi:10.1186/s13046-021-01883-0
29. Mato-Basalo R, Morente-López M, Arntz OJ, van de Loo FAJ, Fafián-Labora J, Arufe MC. Therapeutic Potential for Regulation of the Nuclear Factor Kappa-B Transcription Factor p65 to Prevent Cellular Senescence and Activation of Pro-Inflammatory in Mesenchymal Stem Cells. Int J Mol Sci. 2021;22(7):3367. doi:10.3390/ijms22073367
30. Khayrullin A, Krishnan P, Martinez-Nater L, et al. Very Long-Chain C24:1 Ceramide Is Increased in Serum Extracellular Vesicles with Aging and Can Induce Senescence in Bone-Derived Mesenchymal Stem Cells. Cells. 2019;8(1):37. doi:10.3390/cells8010037
31. Pienimaeki-Roemer A, Konovalova T, Musri MM, Sigruener A, Boettcher A, Meister G. Transcriptomic profiling of platelet senescence and platelet extracellular vesicles. Transfusion. 2017;57(1):144–156. doi:10.1111/trf.13896
32. Goronzy JJ, Weyand CM. Understanding immunosenescence to improve responses to vaccines. Nat Immunol. 2013;14(5):428–436. doi:10.1038/ni.2588
33. Picca A, Guerra F, Calvani R, et al. Mitochondrial Signatures in Circulating Extracellular Vesicles of Older Adults with Parkinson’s Disease: results from the EXosomes in PArkiNson’s Disease (EXPAND) Study. J Clin Med. 2020;9(2):504. doi:10.3390/jcm9020504
34. Masaldan S, Clatworthy SAS, Gamell C, et al. Iron accumulation in senescent cells is coupled with impaired ferritinophagy and inhibition of ferroptosis. Redox Biol. 2018;14:100–115. doi:10.1016/j.redox.2017.08.015
35. Basisty N, Kale A, Jeon OH, et al. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol. 2020;18(1):e3000599. doi:10.1371/journal.pbio.3000599
36. Battaglia R, Musumeci P, Ragusa M, et al. Ovarian aging increases small extracellular vesicle CD81(+) release in human follicular fluid and influences miRNA profiles. Aging. 2020;12(12):12324–12341. doi:10.18632/aging.103441
37. Ashrafizadeh M, Kumar AP, Aref AR, Zarrabi A, Mostafavi E. Exosomes as Promising Nanostructures in Diabetes Mellitus: from Insulin Sensitivity to Ameliorating Diabetic Complications. Int J Nanomed. 2022;17:1229–1253. doi:10.2147/ijn.S350250
38. Aydin Ozgur B, Coskunpinar E, Bilgic Gazioglu S, et al. Effects of Complement Regulators and Chemokine Receptors in Type 2 Diabetes. Immunol Invest. 2021;50(5):478–491. doi:10.1080/08820139.2020.1778022
39. Ma XW, Chang ZW, Qin MZ, Sun Y, Huang HL, He Y. Decreased expression of complement regulatory proteins, CD55 and CD59, on peripheral blood leucocytes in patients with type 2 diabetes and macrovascular diseases. Chin Med J. 2021;122:2123–2128.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Extracellular Vesicles in Skin: Biological Function and Therapeutic Potential

Yuan S, Jin M, Zhang Y, Zhang M, Yuan M, Ding X, Wang J
International Journal of Nanomedicine 2025, 20:11211-11233
Published Date: 13 September 2025
The Dual Role of Extracellular Vesicles in Aging and Age-Related Diseases: Pathophysiology and Therapeutic Potential
Zhu Y, Fang X, Zhang S, Liao Y, Lin H, Chen P, Yang M, Huang J, Wang X
International Journal of Nanomedicine 2026, 21:589123
Published Date: 4 March 2026