Back to Journals » Journal of Inflammation Research » Volume 18
Association Between Maternal IBD-Related Antibodies and Early Life Gut Inflammatory Status and Microbiota Composition: Insights from Cord Blood
Authors Kim T ![]() , Nicoletti P, Estevinho MM, Tarassishin L
, Nicoletti P, Estevinho MM, Tarassishin L ![]() , Picker M, Debebe A, Nguyen I, Dutra-Clarke R, Young C, Stone J, Bar-Gil Shitrit A
, Picker M, Debebe A, Nguyen I, Dutra-Clarke R, Young C, Stone J, Bar-Gil Shitrit A ![]() , Agrawal M, Colombel JF, Torres J, Ng SC, Zhang L, McGovern DPB, Peter I
, Agrawal M, Colombel JF, Torres J, Ng SC, Zhang L, McGovern DPB, Peter I
Received 5 August 2025
Accepted for publication 7 October 2025
Published 18 November 2025 Volume 2025:18 Pages 16009—16022
DOI https://doi.org/10.2147/JIR.S557581
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Nadia Andrea Andreani
Taegyu Kim,1 Paola Nicoletti,1 Maria Manuela Estevinho,2,3 Leonid Tarassishin,1 Mellissa Picker,1 Anketse Debebe,1 Isabella Nguyen,1 Rhiannon Dutra-Clarke,4 Courtney Young,4 Joanne Stone,5 Ariella Bar-Gil Shitrit,6 Manasi Agrawal,7 Jean-Frederic Colombel,7 Joana Torres,7,8 Siew C Ng,9– 11 Lin Zhang,9,12 Dermot P B McGovern,4 Inga Peter1
1Department of Genetics and Genomic Sciences, Icahn School of Medicine at Mount Sinai, New York, NY, USA; 2Department of Gastroenterology, Unidade Local de Saúde Gaia Espinho, Vila Nova de Gaia, Portugal; 3Department of Biomedicine, University of Porto, Porto, Portugal; 4F. Widjaja Inflammatory Bowel Disease Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA; 5Department of Obstetrics, Icahn School of Medicine at Mount Sinai, New York, NY, USA; 6Shaare Zedek Medical Center, The Hebrew University of Jerusalem, Jerusalem, Israel; 7Department of Medicine, Icahn School of Medicine at Mount Sinai, New York, NY, USA; 8Division of Gastroenterology, Hospital da Luz & Hospital Beatriz Ângelo, Loures, Portugal; 9Microbiota I-Center (MagIC), Hong Kong SAR, People’s Republic of China; 10Department of Medicine and Therapeutics, The Chinese University of Hong Kong, Hong Kong SAR, People’s Republic of China; 11New Cornerstone Science Laboratory, The Chinese University of Hong Kong, Hong Kong SAR, People’s Republic of China; 12Department of Anaesthesia and Intensive Care, The Chinese University of Hong Kong, Hong Kong SAR, People’s Republic of China
Correspondence: Inga Peter, Department of Genetics and Genomic Sciences, Icahn School of Medicine at Mount Sinai, 1425 Madison Ave, New York, NY, 10029, USA, Tel +1-212-659-6881, Email [email protected]
Purpose: Antibodies in peripheral blood are used to aid in the diagnosis of inflammatory bowel disease (IBD), but their presence in neonatal cord blood and potential effects on early life development remain unknown.
Methods: We measured anti-CBir1, ANCA, anti-OmpC, ASCA IgA, and ASCA IgG levels in the cord blood of babies born to 78 mothers with or without IBD. Their association with fecal calprotectin (FC), and microbiota composition, characterized by 16S rRNA sequencing, was assessed throughout pregnancy and during the first three and five years of life, respectively, using linear mixed-effects models.
Results: Antibodies were detected in cord blood, with significantly higher levels of anti-CBir1 and ASCA IgG in babies born to mothers with Crohn’s disease (p = 0.002) and higher abundance of ANCA and anti-OmpC in babies of mothers with ulcerative colitis (p = 0.002), compared to controls. ASCA IgG levels positively correlated with babies’ FC (p = 0.006). Both ANCA and anti-OmpC were positively correlated with the abundance of Romboutsia spp. Clostridia, and Pseudomonas spp. while Blautia spp. Citrobacter spp. Clostridiaceae, and Varibaculum spp. were positively correlated with either ANCA or anti-OmpC levels. Higher abundance of Lactobacillus spp. was associated with higher ASCA IgG and anti-CBir1 levels (all multiple testing-adjusted q< 0.1).
Conclusion: Our findings suggest that maternal IBD-associated antibodies cross the placenta barrier and may be associated with intestinal inflammation and imbalanced microbiota colonization. Whether these serological profiles negatively influence the priming of the baby’s immune system or IBD risk later in life remains to be determined.
Keywords: anti-CBir1, ANCA, anti-OmpC, ASCA, inflammatory bowel disease, pregnancy
Introduction
The maternal immune health and gut microbiota composition have been shown to play a crucial role in the offspring’s immune system development both in utero and after birth.1 During pregnancy, the immune-modulating factors – including maternal antibodies and cytokines – are transmitted from the mother to the fetus via the placenta and cord blood. Maternal antibodies provide passive immunity and immediate protection, while cytokines influence how neonatal immune cells are primed to respond to future immune challenges,1–3 such as infections and allergens.4,5 At birth, vaginal seeding is recognized as an essential mechanism through which newborns acquire initial microbial colonization.6 However, by four months of age, most gut colonizers are sourced from the maternal gut microbiome rather than vaginal flora.7 The mother–baby immune-related interactions continue with breastfeeding, which facilitates the transfer of gut microbiome-induced maternal antibodies to the neonatal intestine, along with secretory IgA and other bioactive factors.8–10
Therefore, maternal pre-existing health conditions can affect the infant’s immune system development, mediating long-lasting health outcomes,11 including the predisposition to immune-mediated diseases later in life.12,13 Our team has shown that babies born to mothers with inflammatory bowel disease (IBD), a chronic inflammatory condition of the gastrointestinal tract that may be caused by loss of mucosal tolerance to the commensal microbiota in genetically susceptible individuals,14,15 have lower gut bacterial diversity, resulting in abnormal imprinting of the intestinal immune system in germ-free mice colonized with these infants’ stool.16 Moreover, we demonstrated higher levels of fecal calprotectin (FC) – a well-established biomarker of the presence and degree of intestinal inflammation17 and a predictor of future IBD risk18 - in unaffected offspring of IBD mothers up to at least 3 years of age.19 These findings suggest that maternal IBD may negatively affect fetal–maternal interactions.
Serological antibodies in the peripheral blood, such as anti-flagellin CBir1 (anti-CBir1), anti-outer membrane porin C (anti-OmpC), anti-neutrophil cytoplasmic antibody (ANCA), and anti-Saccharomyces cerevisiae antibody (ASCA), recognize a range of antigens, including microbial proteins (eg, CBir1, OmpC), host-derived cytoplasmic proteins (eg, ANCA), and oligosaccharide antigens (eg, ASCA). These antibodies have been used to aid in the differential diagnosis of IBD and its subtypes, Crohn’s disease (CD) and ulcerative colitis (UC).20–22 High levels of anti-CBir1 indicate a distinct immune response in CD.23 ANCA antibodies are more prevalent in UC, being detected in 60–80% of UC patients but only in 5–20% of CD patients.24–26 ASCA is more frequently detected in CD than UC.27,28 A combination of positive ASCA IgA and IgG with negative ANCA is strongly associated with CD and has prognostic value for disease progression.29,30 While these markers aid in differentiating IBD from non-IBD and CD from UC, their transmission from mother to fetus and their impact on the factors driving immune development during early life remain poorly explored.
Cord blood provides a unique opportunity to study the direct maternal transfer of IBD-associated antibodies to the baby, which can potentially influence immune development, modulate gut microbiome colonization, and impact inflammatory pathways during early life, a critical period for establishing long-term immune homeostasis.31 A recent study established that antibodies like ASCA are not just passive markers but are active initiators of the inflammatory cascade,32 raising a question of what microbial triggers might be responsible for producing these pathogenic antibodies. Therefore, this study aimed to investigate the levels of the top IBD-associated antibodies (anti-CBir1, anti-OmpC, ANCA, and ASCA) in cord blood from pregnancies of women with and without IBD and determine their association with the intestinal inflammatory status and gut microbiota composition of both mother and infant to identify potential relationships that may influence early gut colonization. A better understanding of feto-maternal interactions may help shed light on the potential pathways that promote the risk transmission of immune-mediated diseases, such as IBD, and provide valuable insights to inform early intervention strategies and promote long-term health.
Materials and Methods
Participant Characteristics
This study was conducted with 17 UC cases, 11 CD cases, and 50 controls in a subset of the MECONIUM (Exploring Mechanisms of Disease Transmission In Utero through the Microbiome) cohort16 enrolled at two sites. Specifically, pregnant women with IBD, and age and race/ethnicity-matched controls were recruited on a rolling basis between 2015 and 2021 at Mount Sinai Hospital in New York City, the United States (US; 13 UC, 11 CD, and 28 controls) and between 2019 and 2021 at the medical and IBD clinics at the Prince of Wales Hospital in Hong Kong, China (HKG; 4 UC and 22 controls) after signing an informed consent. Exclusion criteria included the inability to provide consent, HIV/AIDS, other autoimmune diseases, fetal chromosomal or structural abnormalities, and active infection during pregnancy.
The information about age at enrollment, race/ethnicity, smoking status, baseline body mass index (BMI), and IBD-related drug usage during pregnancy (eg, aminosalicylates, steroids, immunomodulators [IMM], and anti-tumor necrosis factor [anti-TNF-α]) was collected for mothers. Disease activity was assessed prospectively by Physician Global Assessment and evaluated as either active or in remission during each trimester. Harvey–Bradshaw Index (HBI) and the Mayo score (without an endoscopic score) were used to assess disease activity in CD and UC, respectively. CD was considered in remission if HBI was <5, while UC was in remission if the partial Mayo score was <2.
In the offspring, the collected data included gestational age at delivery, delivery mode (vaginal or cesarean), sex, exposure to antibiotics, and feeding behavior (exclusive breastfeeding, exclusive formula feeding, or mixed feeding). Infant feeding practices were surveyed at one week, two weeks, one month, two months, and three months postpartum using two dichotomous (yes/no) questions: “Was your child breastfed during this period?” and “Did you breastfeed exclusively during this period?”
This study was approved by the Institutional Review Board of the Icahn School of Medicine at Mount Sinai (HS#14-00554) and Joint Chinese University of Hong Kong – New Territories East Cluster Clinical Research Ethics Committee (CREC no. 2016.547).
Cord Blood Sample Collection and Antibody Response Analysis
Venous umbilical cord blood was collected at the time of delivery using a non-EDTA blood collection tube for the US samples. The tubes were left standing for 30 minutes to allow clotting, and then centrifuged at 1520 × g for 15 minutes at 4°C to collect the serum samples. Serum samples were stored at −20°C until the analysis. Umbilical cord blood (for buffy coat, serum, and plasma [EDTA]) was also collected at the HKG site. Cold chain shipping couriers were employed for the shipment of the HKG samples to Mount Sinai Hospital. Sample shipments complied with International Air Transport Association (IATA) Dangerous Goods Regulations for biological substances (Class 6, Toxic [Poisonous] and Infectious Substances) and dry ice (Class 9, Miscellaneous Dangerous Goods).
Patient sera were shipped on ice using a next-day air service and analyzed in a blinded manner at Cedars-Sinai Medical Center for the expression of several antibodies by enzyme-linked immunosorbent assay (ELISA) following previously described protocols.33–37 The analysis measured both ASCA IgG and ASCA IgA, as well as ANCA, anti-OmpC, and anti-CBir1 (for which IgG and IgA isotypes were not separately measured). In short, test sera were analyzed alongside control reference sera from Cedars-Sinai’s MIRIAD Biobank, including negative and positive controls. ANCA expression was assessed by fixed neutrophil ELISA with methanol-fixed human neutrophils, while ASCA, anti-OmpC, and anti-CBir1 expressions were determined by reactivity to Saccharomyces cerevisiae mannan, E. coli OmpC, and CBir1 flagellin, respectively, on high-binding polystyrene microtiter plates. The assays involved antigen coating, blocking with 0.5% BSA/PBS, incubation with diluted sera, and detection with specific secondary antibodies, followed by pNPP substrate. Plates were read on an Emax microplate reader (Molecular Devices, CA, USA) when positive control optical density (OD) reached 1.00–2.00 OD above blanks. Raw OD values were normalized to ELISA units (EU) relative to positive controls (100 EU), with seropositivity defined by assay-specific cutoffs (ANCA >30 EU, ASCA IgA >20 EU, ASCA IgG >40 EU, anti-OmpC >23 EU, anti-CBir1 >25 EU), which were established with peripheral blood during assay development. These thresholds were typically defined as the mean plus two standard deviations of seroreactivity scores from a cohort of clinically well-characterized CD and UC patient sera for each assay.
Fecal Calprotectin (FC) Analysis
During pregnancy, stool samples were collected at home with an ALPCO Diagnostics EasySampler Stool Collection kit (ALPCO Life Science, Salem, NH, USA) at the 1st (T1), 2nd (T2), and 3rd (T3) trimesters. Following delivery, the babies’ diapers or stool samples were collected at 1W (week), 2W, 1M (month), 2M, 3M, 1Y (year), 1.5Y, 2Y, 3Y, 4Y, and 5Y. Samples were shipped on ice to the lab and stored at −20°C or −80°C until further analysis. The FC concentration from both mothers and babies was determined using a CALPROLAB Calprotectin ELISA kit (Svar Life Science, Lysaker, Norway), as described elsewhere.19 For FC, data up to 3 years of age were used.
Stool Microbiome Analysis
A subset of stool samples (maternal samples n = 108, baby samples n = 401) were profiled using bacterial 16S rRNA gene sequencing with HiSeq paired-end 250 × 2 fast mode (Illumina, San Diego, CA, USA). The fecal DNA extraction and dual-barcoded 16S V4 PCR amplicon preparation was performed as previously described.16 For sequence analysis, mothur was employed38 using a protocol similar to the MiSeq SOP.39 In short, paired-end reads were merged, and sequences with one or more ambiguous bases or a homopolymer longer than eight nucleotides were removed. Sequences were aligned to the SILVA v.138 reference file,40 and chimeric sequences were then identified and discarded using VSEARCH.41 A Bayesian classifier and the SILVA v.138 reference database were used to classify the sequences.42 Sequences that did not classify as Bacteria or Archaea were discarded. The number of sequences was normalized before sequence clustering into operational taxonomic units (OTUs) at a 3% similarity threshold. Then, the numbers of OTUs and Shannon diversity index were calculated for the microbiota alpha diversity, and Bray-Curtis dissimilarity was calculated for the microbiota beta diversity.
Statistical Analysis
Fisher’s exact test and Wilcoxon rank-sum test were employed to evaluate statistical differences between any two groups. Principal Component Analysis (PCA) was conducted to visualize the variation in serological marker levels across the samples, along with the loadings of each serological marker on the principal components. Multiple logistic regression analyses were performed to generate receiver operating characteristic (ROC) curves to assess predictive variables for maternal IBD diagnoses. These analyses were performed using GraphPad Prism, version 10.2.3 (GraphPad, San Diego, CA, USA). To assess the significant differences in serological marker levels between groups of samples, an analysis of similarities (ANOSIM) was performed using the R package vegan,43 and confidence ellipses were calculated by the R package MASS.44
Permutational Multivariate Analysis of Variance (PERMANOVA) was conducted using the adonis2 function from the R package vegan43 to partition the variance in the baby stool microbiota data. The Bray-Curtis dissimilarity matrix was used as the response variable. The model tested the marginal effects of delivery mode, prenatal antibiotic usage, maternal IBD status, exclusive breastfeeding in the first 90 days, geographic location, and time point (baby age) as predictor variables. To account for repeated measures, a linear mixed-effects model was employed using the R package lme4,45 with subject ID as a random effect and fixed effects drawn from disease type, time point, BMI, anti-TNF-α usage, and geographic location. FC values were log-transformed prior to analyses that assumed linear relationships, including Pearson’s correlation and linear mixed-effects model analysis. Principal coordinates analysis (PCoA) and analysis of molecular variance (AMOVA) were conducted using mothur.38 Multiple testing correction using false discovery rate was applied with a significance threshold of q < 0.1.46
Results
Participant Characteristics
The characteristics of the 17 UC, 11 CD, and 50 control subjects are summarized in Table 1. Aside from a higher proportion of mothers in the CD group reporting Jewish ancestry compared to controls (55% vs 12%, p = 0.01), no other significant differences were observed between the groups (p > 0.05).
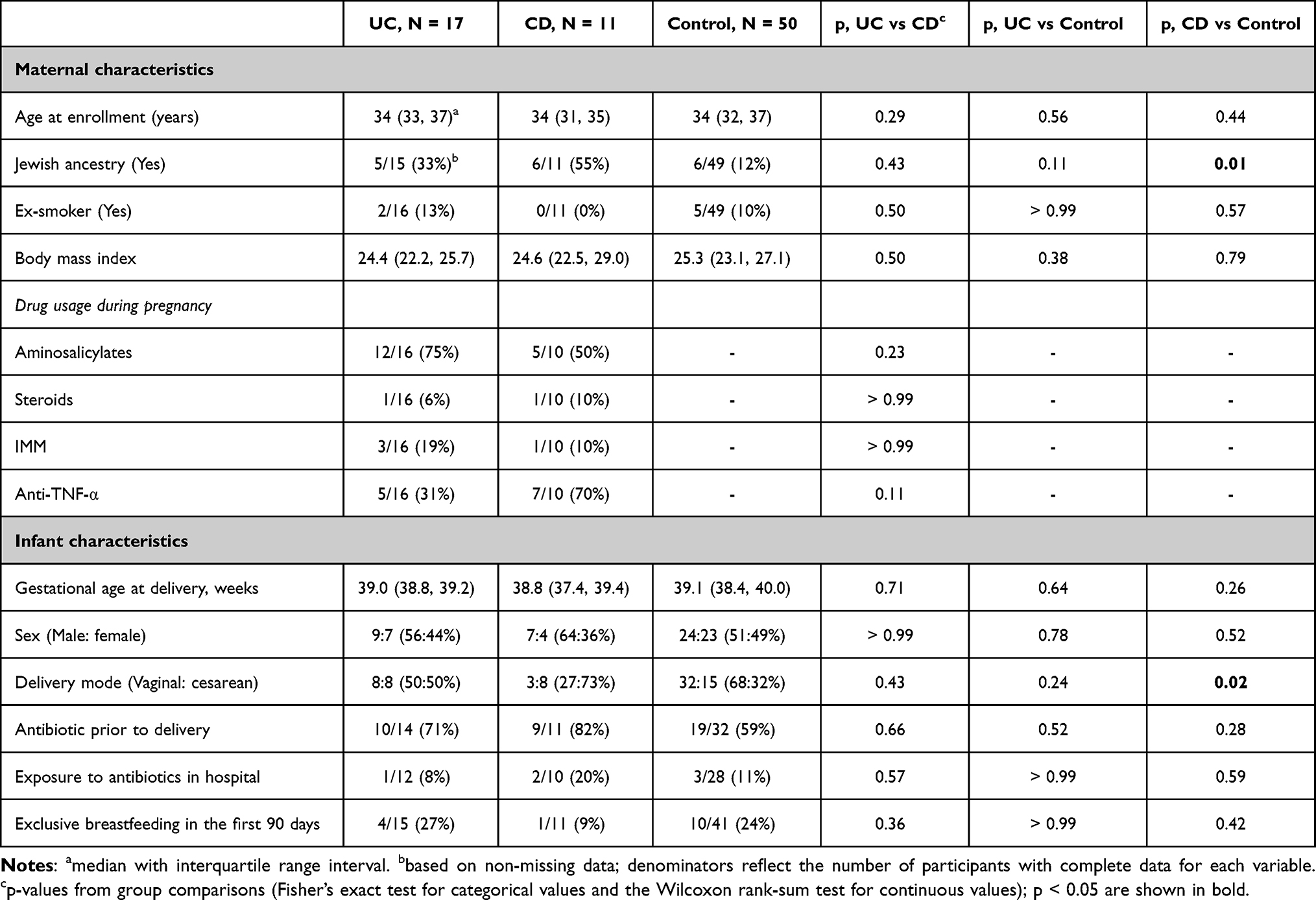 |
Table 1 Participant Characteristics |
Infant sex, exposure to antibiotics in the hospital, and feeding behavior up to 90 days did not differ significantly between groups, although a higher proportion of CD mothers underwent cesarean delivery compared to controls (73% vs 32%, p = 0.02).
Serological Marker Levels in Cord Blood
Serological antibodies were detectable in cord blood samples (Figure 1). With the standard seropositivity threshold established for peripheral blood during assay development at the laboratory, anti-CBir1 was positive in 0% of UC patients, 27% of CD patients, and 2% of controls, while ANCA was positive in 29% of UC patients, 9% of CD patients, and 2% of controls (Table 2). The seropositivity rates of the UC group were distinct from the CD and the control groups (both p < 0.0001); however, the CD group was not distinct from the control group (p = 0.29). This suggests that the standard peripheral blood positivity cutoff may be overly stringent for cord blood samples, and the placental transmissivity of ASCA is lower than that of anti-CBir1 and ANCA. Indeed, when the analysis was repeated using a halved peripheral blood threshold, an exploratory and provisional value chosen due to the lack of established cord blood reference ranges, a significant difference emerged between the CD and control groups (p = 0.0006), and all inter-group comparisons reached statistical significance.
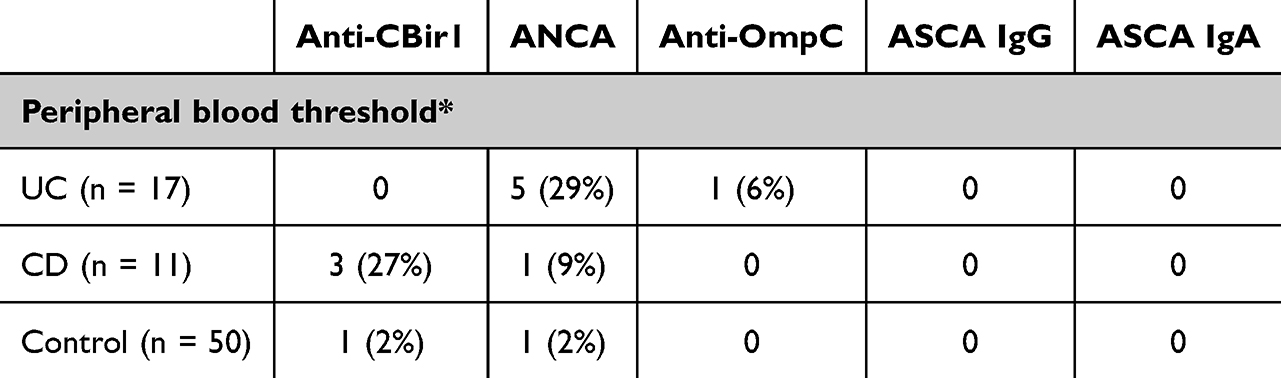 |
Table 2 Number of Seropositivity Cases and Percentages in Parentheses in Each Group |
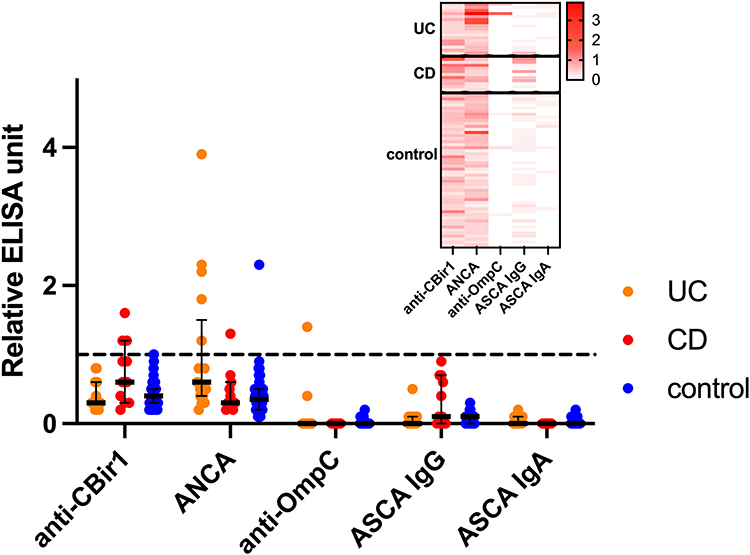 |
Figure 1 Relative ELISA units of five serological biomarker levels in cord blood with respect to the seropositivity threshold used in peripheral blood. The threshold used in peripheral blood is shown as a black dashed line. A heatmap shows the relative ELISA units of each biomarker in each individual. |
Anti-OmpC and ASCA IgA were not detected or detected at a very low level in most samples. Anti-OmpC was positive based on the standard seropositivity threshold in one UC sample. ASCA IgA was detected in 10 out of 17 UC samples, 4 out of 11 CD samples, and 15 out of 50 control samples at a level lower than the threshold. The detected signals were weaker compared to ASCA IgG, which was present in all cord blood samples.
Maternal Disease Classification
Serological marker abundances in cord blood samples were distinctive based on the maternal IBD status (Figure 2A; ANOSIM test for separation between UC vs CD [p = 0.025], UC vs control [p = 0.002], and CD vs control [p = 0.002]). ASCA IgG and anti-CBir1 drove the differences with CD, while anti-OmpC and ANCA levels differentiated the UC group from others.
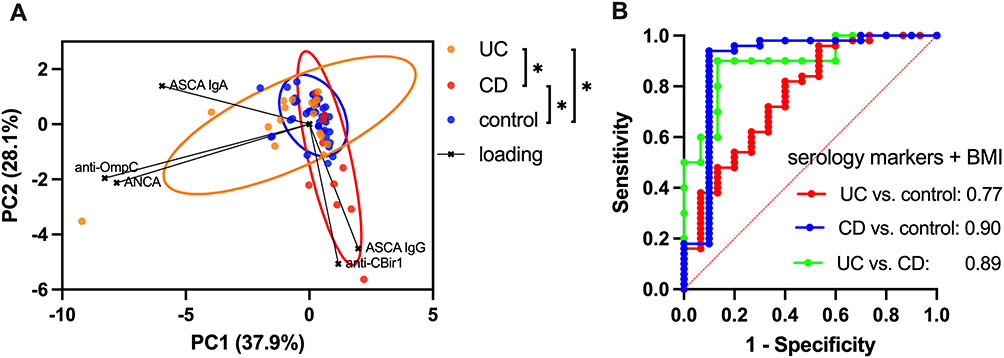 |
Figure 2 (A) Principal component analysis (PCA) of cord blood serological marker levels; asterisks (*) indicate significant group differences as determined by analysis of similarity (ANOSIM; p < 0.05) and 90% confidence ellipses are shown, (B) Receiver operating characteristic (ROC) curves for different disease types (UC, ulcerative colitis; CD, Crohn’s disease, and control), derived from logistic regression models using serology marker levels and maternal BMI. The areas under the curves (AUCs) are displayed in each plot. Detailed test results with sensitivity, specificity, negative/positive predictive values, and accuracy with and without BMI are shown in Table S1. |
Logistic regression analysis also showed that those markers can have predictive value for maternal disease subtypes (Figure 2B). Sensitivity for detecting true UC and CD cases compared to control groups was 29.4% and 54.6%, respectively, while specificity was high for both at 98.0%, with corresponding area under the curves (AUCs) of 0.77 and 0.90 (Table S1). In all three group comparisons among UC, CD, and control, the negative predictive value, positive predictive value, and overall accuracy were all equal to or greater than 80.0%. When the logistic regression model included the maternal BMI, to account for possible hemodilution of serologic markers, sensitivity for detecting true UC and CD cases compared to the control group increased to 33.3% and 70.0%, while specificity remained the same at 98.0% (Table S1).
Within IBD mothers, those treated with any IBD-directed drug exhibited higher cord blood marker levels overall, particularly anti-TNF-α (Figure S1). In the case of UC, the anti-TNF-α-treated group showed significantly higher ANCA levels (p = 0.02). For CD, the anti-TNF-α-treated group demonstrated elevated anti-CBir1 levels with borderline statistical significance (p = 0.08). The median ANCA level appeared to be higher in UC mothers with active disease during pregnancy, although the difference was not statistically significant (p = 0.50), likely due to the limited sample size (5 active, 8 inactive, 4 unknown; Figure S1).
Maternal BMI measured in early pregnancy among IBD mothers was negatively correlated with all serological biomarker levels in cord blood, both in UC and CD groups (rho ranged from −0.16 to −0.68), with the statistical significance shown between BMI and anti-OmpC in the CD group (p = 0.04) (Figure S2). The correlation between BMI and serological biomarker levels in the control group was not statistically significant (rho ranged from −0.02 to 0.27, all p > 0.06; Figure S2).
Fecal Calprotectin (FC) and Serological Markers
Cord blood serology marker levels showed weak positive correlations with maternal FC levels during pregnancy, except for ASCA IgA (Figure 3). Although individual serological markers were not significantly associated with maternal FC levels, a composite z-score combining all five markers was significantly associated with maternal FC after adjusting for time in a multiple linear regression model (p = 0.019). In the baby samples, there was a significant overall association between ASCA IgG and FC levels, after adjustment for time point (p = 0.006), and after adjustment for time point and maternal IBD status (p = 0.05). Pearson’s correlation analysis between ASCA IgG and FC levels at individual time points further supported a positive relationship, with statistically significant associations detected at 2 weeks, 3 months, and 1 year (p = 0.04, 0.04, and 0.02, respectively; Figure 3).
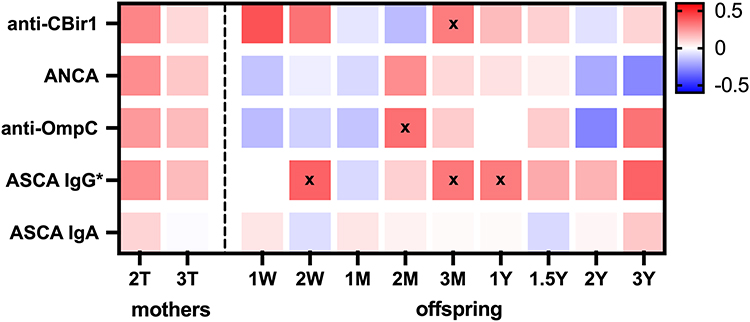 |
Figure 3 Pearson’s correlation coefficient heatmap between log-transformed fecal calprotectin levels and cord blood serological markers in mothers (2nd trimester [2T] and 3rd trimester [3T]) and their babies at multiple time points (W: weeks, M: months, Y: years). An asterisk (*) indicates a statistically significant association in baby samples across multiple time points, based on a mixed-effects model adjusted for time and maternal disease status. “x” marks denote time points with statistically significant positive correlations (p < 0.05). |
Stool Microbiome and Taxa Abundance
To assess associations between cord blood serological biomarker levels and maternal stool microbiota alpha diversity or taxa abundance throughout pregnancy, linear mixed-effects models were used while adjusting for disease subtype, time point, BMI, anti-TNF-α therapy, and geographic location (Figure 4A). For taxa abundance, the analysis included the top 105 bacterial taxa with an average relative abundance >0.05% and revealed a significant negative association between anti-CBir1 antibody levels and the following taxa: Streptococcus spp., Romboutsia spp., Enterobacteriaceae, Actinomyces spp., Granulicatella spp., and Lactobacillales. Conversely, anti-CBir1 levels showed a positive association with Butyricimonas spp., Bacteroidales, Hungatella spp., Ruminococcaceae, Tannerellaceae, Ruminococcaceae UBA1819, and Parabacteroides spp. (all q < 0.1). Additionally, ANCA levels were positively associated with the number of observed OTUs (p = 0.03) and with Barnesiella spp. abundance (q = 0.01).
 |
Figure 4 Sparse plots showing statistically significant beta coefficients from linear mixed-effects models assessing associations between cord blood serological biomarker levels and (A) maternal stool alpha diversity or taxa abundance, adjusted for disease type, time point, BMI, anti-TNF-α usage, and geographic location; and (B) baby stool alpha diversity or taxa abundance, adjusted for delivery mode, perinatal antibiotic usage, time point, and geographic location. The red columns show positive associations, and blue columns show negative associations. For alpha diversity, the results with p < 0.05 are shown. Taxa with an average abundance >0.05% were tested (105 for mothers and 92 for babies), and results are shown only for associations with a multiple testing-adjusted q < 0.1. |
In babies, we examined the association between cord blood serological biomarkers and gut microbiome alpha diversity and the relative abundance of the top 92 bacterial taxa with an average relative abundance >0.05%, while adjusting for delivery mode, prenatal antibiotic usage, time point, and geographic location (Figure 4B). These covariates were selected based on the variance explained in the PERMANOVA analysis (Table S2), as inclusion of additional covariates in the model led to singularity errors. Individual bacterial taxa, Prevotella spp., Turicibacter spp., and Lactobacillus spp., were positively associated with anti-CBir1 levels (all q < 0.1). Lactobacillus spp. and Roseburia spp. were positively associated with ASCA IgG and ASCA IgA levels, respectively (both q < 0.1). Furthermore, positive associations with ANCA and/or anti-OmpC levels were found for several genera, including Blautia spp., Romboutsia spp., Citrobacter spp., Pseudomonas spp., and Varibaculum spp., as well as for the summed abundances of bacteria within Clostridiaceae family and Clostridia class (all q < 0.1; Figure 4B).
To compare cord blood markers with gut microbiota alpha and beta diversity, we focused on anti-CBir1 and ANCA, the two most frequently detected antibodies. Subjects were stratified into four groups based on high versus low levels of each marker to enable group-level comparisons. A provisional threshold set at 50% of the standard peripheral blood seropositivity cutoff was used to define “high” levels. Using this criterion, 25 subjects were classified into the high anti-CBir1 group (including 7 of the 11 CD cases), and 25 subjects into the high ANCA group (including 12 of the 17 UC cases). Maternal stool microbiota composition (beta diversity) significantly differed between the high-ANCA group and the both-low group, both at 2T and 3T, based on the AMOVA test (p = 0.012 and 0.031, respectively; Figure S3). Also, the high-ANCA group showed a higher number of observed OTUs (alpha diversity) compared to the both-low group at T3, although the difference was borderline significant (p = 0.053; Figure S4).
The baby stool microbiota showed significant differences in both alpha and beta diversity between the high-ANCA group and the both-low group at 1, 2, and 3 months of age. AMOVA testing revealed significantly different microbial community composition between the two groups at these time points (p = 0.004, <0.001, and 0.003, respectively; Figure S5). However, by 1 year of age, the difference was no longer significant (F = 1.2, p = 0.23). Similarly, Shannon diversity was significantly lower in the high-ANCA group compared to the both-low group at 1, 2, and 3 months (p = 0.003, 0.006, and 0.016, respectively; Figure S6A), but this difference was not observed after 1 year (Figure S6B).
Discussion
Our study demonstrates, for the first time, that IBD-associated microbial antibodies – predominantly IgG isotypes such as ASCA, anti-CBir1, ANCA, and anti-OmpC - are detectable in cord blood, with higher levels observed in offspring of women living with IBD. Antibody profiles also differ according to maternal disease type, distinguishing between UC and CD. Overall, serological antibody levels showed a positive relationship with maternal FC levels during pregnancy, whereas ASCA IgG levels were positively correlated with the offspring FC levels throughout first months of life. In addition, anti-CBir1, ANCA, and anti-OmpC levels were associated with taxa abundances in the gut microbiota of mothers and their offspring. Our results suggest that pregnant women with IBD have elevated biomarkers of immune response due to mucosal barrier dysfunction and microbial translocation. The transplacental passage of these biomarkers to their babies may be associated with subsequent changes in intestinal health, bacterial diversity, and early life immune system development.
Cord blood is a readily accessible sample obtained at birth that can be a valuable resource for risk stratification. Previous studies have identified specific cytokine-related markers that were predictive of subsequent bronchopulmonary dysplasia development in premature infants47 or durability of infant vaccine responses.48 Maternal IgG antibodies cross the placenta starting in the second trimester of gestation and can be detected in infants up to 6 months of age, while IgA antibodies cross the placenta only negligibly due to their lack of binding to the neonatal Fc receptor and their larger size, particularly in the dimeric form.5,49,50 In our study, antibody profiles were predictive of maternal IBD diagnosis and differed by IBD type: ASCA IgG and anti-CBir1 were elevated in the CD group, while anti-OmpC and ANCA were higher in the UC group. This is in line with the reports in the peripheral blood showing elevated ASCA and anti-CBir1 in CD patients, and ANCA in UC patients.23–28 However, the positivity of anti-OmpC has been shown to be more frequent in CD (30–60%) compared to UC (5–24%),51,52 whereas in the cord blood, we detected them exclusively in mothers with UC, a finding that warrants further validation and raises the question of potential disease-specific placental transfer mechanisms.
Alongside IBD diagnosis, maternal medication, particularly anti-TNF-α therapy, and BMI during pregnancy seemed to be related to cord blood antibody transmission. Notably, we showed that treatment with TNF-α inhibitors was associated with increased levels of several serological biomarkers, with a significant difference observed in the ANCA levels between treated and untreated UC patients. While TNF-α inhibitors are typically used to suppress inflammation, our finding of elevated cord blood ANCA levels in the anti-TNF-α-treated group may reflect paradoxical immune activation, or an induced autoantibody response, as has been observed in other TNF-induced autoimmune phenomena.53 Alternatively, these findings may reflect underlying disease severity not fully controlled by TNF-α inhibitors. Out of the five UC mothers and seven CD mothers treated with anti-TNF-α, only one in each group had active disease during pregnancy, precluding a stratified analysis to discern the effects of medication from those of disease activity.
While TNF-α inhibitors have been shown to be transferred across the placenta and be detectable in infants up to 6 months after birth,49 it remains to be determined whether these drugs directly suppress the infant’s own immune system54 or indirectly alter it by increasing the amount of maternal antibodies transferred across the placenta. In contrast, maternal baseline BMI showed a trend towards a negative relationship with biomarker levels, which could be attributed to physiological changes in pregnancy, including hemodilution. While the direct relationship between BMI and IBD-associated antibody levels during pregnancy has not been reported, one study observed a higher, though not statistically significant, BMI and obesity rate in the low anti-CBir1 group compared to the high anti-CBir1 group.55 These observations emphasize the importance of considering the role of both disease-related and maternal physiological factors in immune “inheritance.”
Maternal gut inflammation may also affect the antibody transmission to the baby by modulating placental barrier permeability or Fc receptor expression, thereby enhancing antibody transfer. FC is a well-established intestinal inflammatory marker and a marker of subclinical immune activation.19 Our study showed that maternal FC levels during pregnancy were positively associated with cord blood antibody levels, with a similar relationship reported in the peripheral blood.56 Subsequently, cord blood ASCA IgG levels were also positively correlated with infant FC levels, suggesting that maternal gut inflammation may have an effect on neonatal gut health, which in turn may contribute to the priming of the immune system and susceptibility to immune-mediated diseases later in life.57 Further studies are needed to elucidate the mechanistic pathways between maternal ASCA IgG and future disease susceptibility in the baby.
Microbial taxa can have a bidirectional association with antibody levels. Microbes shape immunity by stimulating antibody response; conversely, the antibodies can influence microbes by targeting bacteria for immune clearance. Cord blood may capture shared immunological and microbial exposures of the mother and the fetus, offering insights into the early development of the infant microbiome. Previous studies have shown that prenatal inflammatory signals can influence neonatal immune tolerance and microbial interactions.58 We observed a significant relationship between serological antibodies and the abundance of specific bacterial taxa in both the mothers and the babies. For instance, in mothers, anti-CBir1 levels were negatively associated with the Streptococcus spp. and the Enterobacteriaceae family, which are often considered potentially proinflammatory.59 Conversely, anti-CBir1 levels were positively associated with Butyricimonas spp. and the Ruminococcaceae family, which includes essential butyrate producers.60 A positive association was also observed with Bacteroidales, which are generally beneficial anaerobic bacteria. In babies, both ANCA and anti-OmpC were positively correlated with the abundance of Romboutsia spp., a commensal gut inhabitant whose overgrowth could contribute to gut inflammation,61 as well as Clostridia and Pseudomonas spp., both of which contain opportunistic pathogens.62,63 Interestingly, higher abundance of Lactobacillus spp. was associated with higher ASCA IgG and anti-CBir1 levels. While generally considered beneficial, increased Lactobacillus abundance has also been noted in other diseases, leading researchers to question whether it is a cause or a result of the pro-inflammatory environment.64 Given that antibody-microbiota interactions may influence gut barrier function and immune tolerance, future studies should explore how specific antibody-microbiota signatures during early life inform the priming of the immune system in the offspring of IBD patients.
This is the first study that integrates cord blood antibody levels with serial FC and gut microbiota profiling in both mothers and babies, to identify potential inflammatory signals of disease transmission from pregnant women with IBD. However, several limitations must be acknowledged. While the stool samples were collected throughout pregnancy and during the first five years of life, no peripheral blood was available to assess the correlation with cord blood antibody levels in mothers and babies or to verify the familial concordance of these serological markers, as has been demonstrated in previous work.65 Also, despite the prospective study design, the identified associations do not establish causal relationships. Moreover, the modest sample size and lack of CD cases from the HKG site may limit the generalizability of our findings. Although we observed several meaningful trends, our subgroup analyses were particularly affected. For instance, comparisons based on disease activity were likely underpowered, and we cannot rule out clinically meaningful differences in antibody levels between mothers with active UC versus those in remission. The results from these specific analyses should, therefore, be interpreted with caution and viewed as exploratory. Also, the relationship between the cord blood antibodies and taxa abundance in the baby stool during the first five years of life was studied with adjustment for time point, geographic location, delivery mode, and prenatal antibiotic use. We acknowledge that other confounders, such as maternal IBD status and other health conditions, diet, and environmental exposures, could affect both cord blood antibody transfer and gut microbiota composition and should be incorporated in future studies. Additionally, our results should be validated in larger cohorts to allow a better understanding of the effects of medications and other prenatal risk factors on IBD transmission. Furthermore, given the absence of established seropositivity thresholds in cord blood, our findings should be validated in larger cohorts to define cord blood-specific reference ranges.
Conclusions
In summary, our findings suggest that maternal IBD-associated antibodies cross the placenta barrier and may contribute to intestinal inflammation and imbalanced microbiota colonization during early life. Whether these serological profiles negatively influence the priming of the baby’s immune system or increase IBD risk in the offspring remains to be determined in the longer longitudinal follow-up.
Abbreviations
AMOVA, Analysis of Molecular Variance; ANCA, Anti-neutrophil cytoplasmic antibody; ANOSIM, Analysis of Similarities; anti-CBir1, Anti-flagellin CBir1; anti-OmpC, Anti-outer membrane porin C; anti-TNF-α, Anti-tumor necrosis factor-alpha; ASCA, Anti-Saccharomyces cerevisiae antibody; AUCs, Areas Under the Curves; BMI, Body Mass Index; CD, Crohn’s Disease; ELISA, Enzyme-linked Immunosorbent Assay; EU, ELISA Units; FC, Fecal Calprotectin; HBI, Harvey–Bradshaw Index; HKG, Hong Kong; IBD, Inflammatory Bowel Disease; IgA, Immunoglobulin A; IgG, Immunoglobulin G; IMM, Immunomodulators; OD, Optical Density; OTUs, Operational Taxonomic Units; PCA, Principal Component Analysis; PCoA, Principal Coordinates Analysis; ROC, Receiver Operating Characteristic; UC, Ulcerative Colitis.
Data Sharing Statement
Data from this study can be obtained by making a reasonable request to the corresponding author.
Ethics Approval and Consent to Participate
This study was approved by the Institutional Review Board of the Icahn School of Medicine at Mount Sinai (HS#14-00554) as well as the Joint Chinese University of Hong Kong – New Territories East Cluster Clinical Research Ethics Committee (2016.547). This research complies with ethics regulations and infants’ written informed consent were provided by their parents. This study was conducted in accordance with the principles of the Declaration of Helsinki.
Consent for Publication
All patient data included in this manuscript has been de-identified, and therefore, this section is not applicable.
Acknowledgments
The authors would like to thank all study participants, recruiting physicians, as well as CALPRO AS for providing ELISA kits. This paper has been uploaded to bioRxiv as a preprint: https://doi.org/10.1101/2025.08.04.668517.
Funding
This study was supported by the International Organization for the Study of Inflammatory Bowel Disease (to I.P. and J.F.C.), the Crohn’s and Colitis Foundation (to I.P., J.T., J.F.C.), National Institutes of Health’s National Institute of Diabetes and Digestive and Kidney Diseases (DK125906, to I.P., J.F.C.), and the National Institute of Diabetes and Digestive and Kidney Diseases (K23DK129762-02, to M.A., and U01DK062413, to DPBM). This work was also supported in part through the computational and data resources and staff expertise provided by Scientific Computing and Data at the Icahn School of Medicine at Mount Sinai and supported by the Clinical and Translational Science Awards (CTSA) grant UL1TR004419 from the National Center for Advancing Translational Sciences. Research reported in this publication was also supported by the Office of Research Infrastructure of the National Institutes of Health under award numbers S10OD026880 and S10OD030463. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. This work was also supported in part by The Leona M. and Harry B. Helmsley Charitable Trust and SCN was supported by the New Cornerstone Science Foundation through the New Cornerstone Investigator Program (grant no. NCI202346). SCN and LZ are affiliated with MagIC and partially supported by InnoHK, The Government of Hong Kong, Special Administrative Region of the People’s Republic of China.
Disclosure
S.C.N. has served as an advisory board member for Pfizer, Ferring, Janssen and Abbvie and received honoraria as a speaker for Ferring, Tillotts, Menarini, Janssen, Abbvie and Takeda; has received research grants through her affiliated institutions from Olympus, Ferring and Abbvie; is a founder member, non-executive Board director and non-executive scientific advisor of GenieBiome Ltd and its holding company which is non-remunerative; is a shareholder of GenieBiome Holdings Ltd; is a founder member, non-executive Board director and non-executive scientific advisor of MicroSigX Biotech Diagnostic Limited and its holding company which is non-remunerative; is a shareholder of MicroSigX Diagnostic Holdings Limited; and receives patent royalties through her affiliated institutions. Paola Nicoletti reports grants from NIH, during the conduct of the study; personal fees from CHIESI FARMACEUTICI, outside the submitted work; Ariella Shitrit reports personal fees from Takeda, personal fees from AbbVie, personal fees from Janssen, personal fees from Lilly, outside the submitted work; Jean-Frederic Colombel reports grants, personal fees from AbbVie, personal fees from Amgen, personal fees from AnaptysBio, personal fees from Astellas, personal fees from Boehringer Ingelheim, grants from Bristol Myers Squibb, personal fees from candidrx, personal fees from Celgene, personal fees from Celltrion, personal fees from Clearview, personal fees from Curogen, personal fees from Eli Lilly, personal fees from Ferring Pharmaceuticals, personal fees from Glaxo Smith Kline, personal fees from Roche, grants from anssen Pharmaceuticals, personal fees from Merck, personal fees from Novartis, personal fees from Otsuka Pharmaceutical, personal fees from Owkin, personal fees from Pfizer, personal fees from Protagonist Therapeutics, personal fees from Sanofi, personal fees from Sun Pharma, grants from Takeda, personal fees from Teva, personal fees from TiGenix, other from Intestinal Biotech Development, outside the submitted work; Dermot McGovern reports grants, personal fees from Prometheus Laboratories, personal fees from MERCK, personal fees from TAKEDA, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Jennewein MF, Abu-Raya B, Jiang Y, Alter G, Marchant A. Transfer of maternal immunity and programming of the newborn immune system. Semin Immunopathol. 2017;39(6):605–613. doi:10.1007/s00281-017-0653-x
2. Gomez de Aguero M, Ganal-Vonarburg SC, Fuhrer T, et al. The maternal microbiota drives early postnatal innate immune development. Science. 2016;351(6279):1296–1302. doi:10.1126/science.aad2571
3. Langel SN, Blasi M, Permar SR. Maternal immune protection against infectious diseases. Cell Host Microbe. 2022;30(5):660–674. doi:10.1016/j.chom.2022.04.007
4. Sabic D, Koenig JM. A perfect storm: fetal inflammation and the developing immune system. Pediatr Res. 2020;87(2):319–326. doi:10.1038/s41390-019-0582-6
5. Palmeira P, Quinello C, Silveira-Lessa AL, Zago CA, Carneiro-Sampaio M. IgG placental transfer in healthy and pathological pregnancies. Clin Dev Immunol. 2012;2012(1):985646. doi:10.1155/2012/985646
6. Martino C, Dilmore AH, Burcham ZM, Metcalf JL, Jeste D, Knight R. Microbiota succession throughout life from the cradle to the grave. Nat Rev Microbiol. 2022;20(12):707–720. doi:10.1038/s41579-022-00768-z
7. Ferretti P, Pasolli E, Tett A, et al. Mother-to-infant microbial transmission from different body sites shapes the developing infant gut microbiome. Cell Host Microbe. 2018;24(1):133–145e5. doi:10.1016/j.chom.2018.06.005
8. Sanidad KZ, Amir M, Ananthanarayanan A, et al. Maternal gut microbiome-induced IgG regulates neonatal gut microbiome and immunity. Sci Immunol. 2022;7(72):eabh3816. doi:10.1126/sciimmunol.abh3816
9. Verhasselt V, Tellier J, Carsetti R, Tepekule B. Antibodies in breast milk: pro-bodies designed for healthy newborn development. Immunol Rev. 2024;328(1):192–204. doi:10.1111/imr.13411
10. Guo J, Ren C, Han X, Huang W, You Y, Zhan J. Role of IgA in the early-life establishment of the gut microbiota and immunity: implications for constructing a healthy start. Gut Microbes. 2021;13(1):1–21. doi:10.1080/19490976.2021.1908101
11. Romano-Keeler J, Weitkamp JH. Maternal influences on fetal microbial colonization and immune development. Pediatr Res. 2015;77(1–2):189–195. doi:10.1038/pr.2014.163
12. Knight DJ, Girling KJ. Gut flora in health and disease. Lancet. 2003;361(9371):1831. doi:10.1016/S0140-6736(03)12489-0
13. Gensollen T, Iyer SS, Kasper DL, Blumberg RS. How colonization by microbiota in early life shapes the immune system. Science. 2016;352(6285):539–544. doi:10.1126/science.aad9378
14. Torres J, Mehandru S, Colombel JF, Peyrin-Biroulet L. Crohn’s disease. Lancet. 2017;389(10080):1741–1755. doi:10.1016/S0140-6736(16)31711-1
15. Ungaro R, Mehandru S, Allen PB, Peyrin-Biroulet L, Colombel JF. Ulcerative colitis. Lancet. 2017;389(10080):1756–1770. doi:10.1016/S0140-6736(16)32126-2
16. Torres J, Hu J, Seki A, et al. Infants born to mothers with IBD present with altered gut microbiome that transfers abnormalities of the adaptive immune system to germ-free mice. Gut. 2020;69(1):42–51. doi:10.1136/gutjnl-2018-317855
17. Oord T, Hornung N. Fecal calprotectin in healthy children. Scand J Clin Lab Invest. 2014;74(3):254–258. doi:10.3109/00365513.2013.879732
18. Klingberg E, Strid H, Stahl A, et al. A longitudinal study of fecal calprotectin and the development of inflammatory bowel disease in ankylosing spondylitis. Arthritis Res Ther. 2017;19(1):21. doi:10.1186/s13075-017-1223-2
19. Kim ES, Tarassishin L, Eisele C, et al. Longitudinal changes in fecal calprotectin levels among pregnant women with and without inflammatory bowel disease and their babies. Gastroenterology. 2021;160(4):1118–1130e3. doi:10.1053/j.gastro.2020.11.050
20. Millman G. Neonatal immunity. Arch Dischildhood. 2005;90(4):F352–f352. doi:10.1136/adc.2004.068759
21. Bourgonje AR, Vogl T, Segal E, Weersma RK. Antibody signatures in inflammatory bowel disease: current developments and future applications. Trends Mol Med. 2022;28(8):693–705. doi:10.1016/j.molmed.2022.05.004
22. Prideaux L, De Cruz P, Ng SC, Kamm MA. Serological antibodies in inflammatory bowel disease: a systematic review. Inflamm Bowel Dis. 2012;18(7):1340–1355. doi:10.1002/ibd.21903
23. Targan SR, Landers CJ, Yang H, et al. Antibodies to CBir1 flagellin define a unique response that is associated independently with complicated Crohn’s disease. Gastroenterology. 2005;128(7):2020–2028. doi:10.1053/j.gastro.2005.03.046
24. Seibold F, Weber P, Schoning A, Mork H, Goppel S, Scheurlen M. Neutrophil antibodies (pANCA) in chronic liver disease and inflammatory bowel disease: do they react with different antigens? Eur J Gastroenterol Hepatol. 1996;8(11):1095–1100. doi:10.1097/00042737-199611000-00012
25. Linskens RK, Mallant-Hent RC, Groothuismink ZM, et al. Evaluation of serological markers to differentiate between ulcerative colitis and Crohn’s disease: pANCA, ASCA and agglutinating antibodies to anaerobic coccoid rods. Eur J Gastroenterol Hepatol. 2002;14(9):1013–1018. doi:10.1097/00042737-200209000-00013
26. Roozendaal C, Kallenberg C. Are anti-neutrophil cytoplasmic antibodies (ANCA) clinically useful in inflammatory bowel disease (IBD)? Clin Exp Immunol. 1999;116(2):206–213. doi:10.1046/j.1365-2249.1999.00905.x
27. Sendid B, Quinton J-F, Charrier G, et al. Anti-saccharomyces cerevisiaemannan antibodies in familial crohn’s disease. Official J Am College Gastroenterol. 1998;93(8):1306–1310. doi:10.1111/j.1572-0241.1998.00415.x
28. Zhou G, Song Y, Yang W, et al. ASCA, ANCA, ALCA and many more: are they useful in the diagnosis of inflammatory bowel disease? Dig Dis. 2016;34(1–2):90–97. doi:10.1159/000442934
29. Quinton JF, Sendid B, Reumaux D, et al. Anti-Saccharomyces cerevisiae mannan antibodies combined with antineutrophil cytoplasmic autoantibodies in inflammatory bowel disease: prevalence and diagnostic role. Gut. 1998;42(6):788–791. doi:10.1136/gut.42.6.788
30. Dubinsky MC, Ofman JJ, Urman M, Targan SR, Seidman EG. Clinical utility of serodiagnostic testing in suspected pediatric inflammatory bowel disease. Am J Gastroenterol. 2001;96(3):758–765. doi:10.1111/j.1572-0241.2001.03618.x
31. Sereme Y, Toumi E, Saifi E, Faury H, Skurnik D. Maternal immune factors involved in the prevention or facilitation of neonatal bacterial infections. Cell Immunol. 2024;395-396:104796. doi:10.1016/j.cellimm.2023.104796
32. Gaifem J, Rodrigues CS, Petralia F, et al. A unique serum IgG glycosylation signature predicts development of Crohn’s disease and is associated with pathogenic antibodies to mannose glycan. Nat Immunol. 2024;25(9):1692–1703. doi:10.1038/s41590-024-01916-8
33. Landers CJ, Cohavy O, Misra R, et al. Selected loss of tolerance evidenced by Crohn’s disease-associated immune responses to auto- and microbial antigens. Gastroenterology. 2002;123(3):689–699. doi:10.1053/gast.2002.35379
34. Lodes MJ, Cong Y, Elson CO, et al. Bacterial flagellin is a dominant antigen in Crohn disease. J Clin Invest. 2004;113(9):1296–1306. doi:10.1172/JCI20295
35. Saxon A, Shanahan F, Landers C, Ganz T, Targan S. A distinct subset of antineutrophil cytoplasmic antibodies is associated with inflammatory bowel disease. J Allergy Clin Immunol. 1990;86(2):202–210. doi:10.1016/s0091-6749(05)80067-3
36. Papadakis KA, Yang H, Ippoliti A, et al. Anti-flagellin (CBir1) phenotypic and genetic Crohn’s disease associations. Inflamm Bowel Dis. 2007;13(5):524–530. doi:10.1002/ibd.20106
37. Sutton CL, Yang H, Li Z, Rotter JI, Targan SR, Braun J. Familial expression of anti-saccharomyces cerevisiae mannan antibodies in affected and unaffected relatives of patients with crohn’s disease. Gut. 2000;46(1):58–63. doi:10.1136/gut.46.1.58
38. Schloss PD, Westcott SL, Ryabin T, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol. 2009;75(23):7537–7541. doi:10.1128/AEM.01541-09
39. Kozich JJ, Westcott SL, Baxter NT, Highlander SK, Schloss PD. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl Environ Microbiol. 2013;79(17):5112–5120. doi:10.1128/AEM.01043-13
40. Quast C, Pruesse E, Yilmaz P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2012;41(D1):D590–D596. doi:10.1093/nar/gks1219
41. Rognes T, Flouri T, Nichols B, Quince C, Mahé F. VSEARCH: a versatile open source tool for metagenomics. PeerJ. 2016;4(e2584). doi:10.7717/peerj.2584
42. Wang Q, Garrity GM, Tiedje JM, Cole JR. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 2007;73(16):5261–5267. doi:10.1128/AEM.00062-07
43. Dixon P. VEGAN, a package of R functions for community ecology. J Veg Sci. 2003;14(6):927–930. doi:10.1111/j.1654-1103.2003.tb02228.x
44. Ripley B, Venables B, Bates DM, et al. Package ‘mass’. Cran r. 2013;538(113–120):822. doi:10.32614/CRAN.package.MASS
45. Bates D, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. Journal of Statistical Software. 2015;67:1–48. doi:10.18637/jss.v067.i01
46. Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc. 1995;57(1):289–300. doi:10.1111/j.2517-6161.1995.tb02031.x
47. Tian XY, Zhang XD, Li QL, Shen Y, Zheng J. Biological markers in cord blood for prediction of bronchopulmonary dysplasia in premature infants. Clin Exp Obstet Gynecol. 2014;41(3):313–318. doi:10.12891/ceog16292014X
48. Baloh CH, Venturi GM, Fischer BM, et al. Biomarkers detected in cord blood predict vaccine responses in young infants. Front Immunol. 2023;14:1152538. doi:10.3389/fimmu.2023.1152538
49. Mahadevan U, Wolf DC, Dubinsky M, et al. Placental transfer of anti-tumor necrosis factor agents in pregnant patients with inflammatory bowel disease. Clin Gastroenterol Hepatol. 2013;11(3):286–92;quize24. doi:10.1016/j.cgh.2012.11.011
50. Kubiak JM, Murphy EA, Yee J, et al. Severe acute respiratory syndrome coronavirus 2 serology levels in pregnant women and their neonates. Am J Obstet Gynecol. 2021;225(1):73e1–73e7. doi:10.1016/j.ajog.2021.01.016
51. Mitsuyama K, Niwa M, Takedatsu H, et al. Antibody markers in the diagnosis of inflammatory bowel disease. World J Gastroenterol. 2016;22(3):1304–1310. doi:10.3748/wjg.v22.i3.1304
52. Kohoutova D, Drahosova M, Moravkova P, Rejchrt S, Bures J. Anti-Outer membrane protein C and anti-glycoprotein 2 antibodies in inflammatory bowel disease and their association with complicated forms of Crohn’s disease. BMC Gastroenterol. 2014;14:190. doi:10.1186/s12876-014-0190-1
53. De Bandt M, Sibilia J, Le Loet X, et al. Systemic lupus erythematosus induced by anti-tumour necrosis factor alpha therapy: a French national survey. Arthritis Res Ther. 2005;7(3):R545–51. doi:10.1186/ar1715
54. Ghalandari N, Kemper E, Crijns IH, et al. Analysing cord blood levels of TNF inhibitors to validate the EULAR points to consider for TNF inhibitor use during pregnancy. Ann Rheum Dis. 2022;81(3):402–405. doi:10.1136/annrheumdis-2021-221036
55. Ahmed Z, Lysek M, Zhang N, Malik TA. Association between serological markers and crohn’s disease activity. J Clin Med Res. 2020;12(1):6–12. doi:10.14740/jocmr4016
56. Smids C, Horjus Talabur Horje CS, Groenen MJM, van Koolwijk EHM, Wahab PJ, van Lochem EG. The value of serum antibodies in differentiating inflammatory bowel disease, predicting disease activity and disease course in the newly diagnosed patient. Scand J Gastroenterol. 2017;52(10):1104–1112. doi:10.1080/00365521.2017.1344875
57. Renz H, Brandtzaeg P, Hornef M. The impact of perinatal immune development on mucosal homeostasis and chronic inflammation. Nat Rev Immunol. 2012;12(1):9–23. doi:10.1038/nri3112
58. Gao Y, O’Hely M, Quinn TP, et al. Maternal gut microbiota during pregnancy and the composition of immune cells in infancy. Front Immunol. 2022;13:986340. doi:10.3389/fimmu.2022.986340
59. Baldelli V, Scaldaferri F, Putignani L, Del Chierico F. The role of enterobacteriaceae in gut microbiota dysbiosis in inflammatory bowel diseases. Microorganisms. 2021;9(4):697. doi:10.3390/microorganisms9040697
60. Louis P, Flint HJ. Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol. 2017;19(1):29–41. doi:10.1111/1462-2920.13589
61. Wang HG, Zhang MN, Wen X, et al. Cepharanthine ameliorates dextran sulphate sodium-induced colitis through modulating gut microbiota. Microb Biotechnol. 2022;15(8):2208–2222. doi:10.1111/1751-7915.14059
62. Gellatly SL, Hancock RE. Pseudomonas aeruginosa: new insights into pathogenesis and host defenses. Pathog Dis. 2013;67(3):159–173. doi:10.1111/2049-632X.12033
63. Nitzan O, Elias M, Chazan B, Raz R, Saliba W. Clostridium difficile and inflammatory bowel disease: role in pathogenesis and implications in treatment. World J Gastroenterol. 2013;19(43):7577–7585. doi:10.3748/wjg.v19.i43.7577
64. Huang R, Wu F, Zhou Q, et al. Lactobacillus and intestinal diseases: mechanisms of action and clinical applications. Microbiol Res. 2022;260:127019. doi:10.1016/j.micres.2022.127019
65. Poulain D, Sendid B, Fajardy I, Danze PM, Colombel JF. Mother to child transmission of anti-S cerevisiae mannan antibodies (ASCA) in non-IBD families. Gut. 2000;47(6):870–871. doi:10.1136/gut.47.6.870a
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.