Back to Journals » International Journal of Women's Health » Volume 18
Association Between Hemostatic Profiles and Hypertensive Disorders of Pregnancy: A Multi-Omics Mendelian Randomization Study
Authors Li Y, Long X, Zhang Y, Tu S, Luo W, Wang X, Zeng L, Ye F, Deng X, Zhang Z
Received 13 February 2026
Accepted for publication 19 May 2026
Published 15 June 2026 Volume 2026:18 603459
DOI https://doi.org/10.2147/IJWH.S603459
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Vinay Kumar
Yixuan Li,* Xianglin Long,* Yeshen Zhang,* Shan Tu, Wenzhi Luo, Xiaoyan Wang, Lixiong Zeng, Fei Ye, Xu Deng, Zhihui Zhang
Department of Cardiology, The Third Xiangya Hospital of Central South University, Changsha, 410013, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhihui Zhang, Department of Cardiology, The Third Xiangya Hospital of Central South University, Changsha, 410013, People’s Republic of China, Email [email protected] Xu Deng, Department of Cardiology, The Third Xiangya Hospital of Central South University, Changsha, 410013, People’s Republic of China, Email [email protected]
Background: Hypertensive disorders of pregnancy (HDPs) are important contributors to maternal mortality worldwide. Activation of the coagulation cascade is a hallmark of preeclampsia, but its causal role remains unclear. This study aimed to investigate the potential causal relationship between hemostatic profiles and the risk of HDPs.
Methods: To investigate the potential causal associations between hemostatic profiles and the risk of HDPs, two-sample Mendelian randomization (MR) analysis was conducted utilizing genome-wide association studies summary statistics. Subsequently, we performed proteome-wide MR analysis to further assess the causal relationship of circulating hemostatic factors on the risk of HDPs. In addition, heterogeneity test, horizontal pleiotropy test, Steiger filtering, Bonferroni correction, and Bayesian colocalization were carried out to evaluate the robustness of the findings.
Results: Our two-sample MR analysis identified nominally significant associations between genetically predicted higher vWF (OR 1.27, 95% CI 1.06– 1.51), FVIII (OR 2.21, 95% CI 1.05– 4.66), and lower Protein C (OR: 0.91, 95% CI: 0.83– 0.99) with a higher risk of preeclampsia. The proteome-wide MR analysis further implicated nominally significant associations for circulating levels of FII, FV, and vWF with preeclampsia risk. Sensitivity analyses showed no evidence of widespread horizontal pleiotropy or heterogeneity, and Steiger filtering confirmed the causal direction. However, these associations did not reach statistical significance after Bonferroni correction and Bayesian colocalization analysis indicated low posterior probabilities for a shared causal variant.
Conclusion: Our findings provide suggestive genetic evidence implicating several hemostatic factors, particularly vWF, in the pathophysiology of preeclampsia. These results suggest the potential involvement of the hemostatic system in the development of HDPs and warrant further investigation to validate these exploratory findings, given the lack of robustness after multiple testing correction. The infographic illustrates the Mendelian randomization study design. The top section shows exposures (hemostatic profiles and circulating proteins from GWAS and deCODE data) and outcomes (HDPs, gestational hypertension, and preeclampsia from FinnGen database). The bottom section shows the coagulation cascade (intrinsic, extrinsic, and common pathways) and key genetically predicted factors (vWF, FII, FV, FVIII as risk factors; Protein C as a protective factor), with MR analysis linking them to preeclampsia risk.Mendelian randomization design for hemostatic factors and preeclampsia risk. Exposures: hemostatic profiles and circulating proteins. Outcomes: HDPs, gestational hypertension, preeclampsia. Key factors: vWF, FII, FV, FVIII, Protein C.
Keywords: hypertensive disorders of pregnancy, coagulation factors, preeclampsia, Mendelian randomization
Introduction
Hypertensive disorders of pregnancy (HDPs) are one of the most common pregnancy complications that lead to short- and long-term fetal and maternal health concerns, mainly including gestational hypertension, preeclampsia, and eclampsia.1,2 The global prevalence of HDPs is approximately 116 per 100 000 women of childbearing age and has increased over time.3 Notably, 14% of all maternal deaths globally are attributed to HDPs, especially preeclampsia.4,5 However, the only definitive treatment of preeclampsia is delivery, and there is currently no effective pharmacological treatment for preeclampsia.6 It is possible for preeclampsia to persist after delivery or to develop de novo in some cases during the postpartum period.7 In addition, although many theories have been established to elucidate the pathophysiology of preeclampsia, the etiology of preeclampsia has not been fully elucidated. Therefore, there is a need to further understand the etiology of preeclampsia to promote the development of pharmacological treatment or prophylactic strategies to improve perinatal outcomes for both mother and fetus.
Previous studies have demonstrated that platelet and coagulation cascade activation are the prominent features of preeclampsia.8–10 Accordingly, low-dose aspirin is widely recommended for pregnant women at high risk for preeclampsia.11 A previous meta-analysis found that both aspirin and low-molecular-weight heparin (LMWH) reduced the risk of preeclampsia, but the certainty of the evidence was low.12 In addition, the mechanisms by which aspirin and LMWH prevent preeclampsia remain unclear, and proposed mechanisms are largely speculative and based primarily on in vitro research.13,14 Notably, it is also unknown whether platelet and coagulation cascade activation causally contribute to the etiology of preeclampsia. Therefore, elucidating the associations of platelet and coagulation cascade activation with preeclampsia would facilitate the prevention and treatment of preeclampsia.
Mendelian randomization (MR) analysis has emerged as a powerful method that utilizes genetic variants as instrumental variables (IVs) to investigate causal relationships between exposures and outcomes, which helps to reduce confounding bias and reverse causation.15 In the present study, we conducted a two-sample MR analysis to evaluate the potential causal relationships between hemostatic profiles and HDPs using genome-wide association studies (GWAS) summary statistics. Subsequently, we performed a proteome-wide MR analysis to explore whether circulating hemostatic factors were causally associated with HDPs based on a large-scale protein GWAS. Furthermore, several sensitivity analyses, including Bayesian colocalization, were conducted to evaluate the robustness of the associations between circulating hemostatic factors and HDPs.
Methods
Study Design
The overall design of this MR analysis is presented in Figure 1. This study is based on publicly available data, and ethical approval and patient consent had already been received in each data source. We reported the results following the Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization (STROBE-MR) reporting guideline (Supplemental Table S1).16
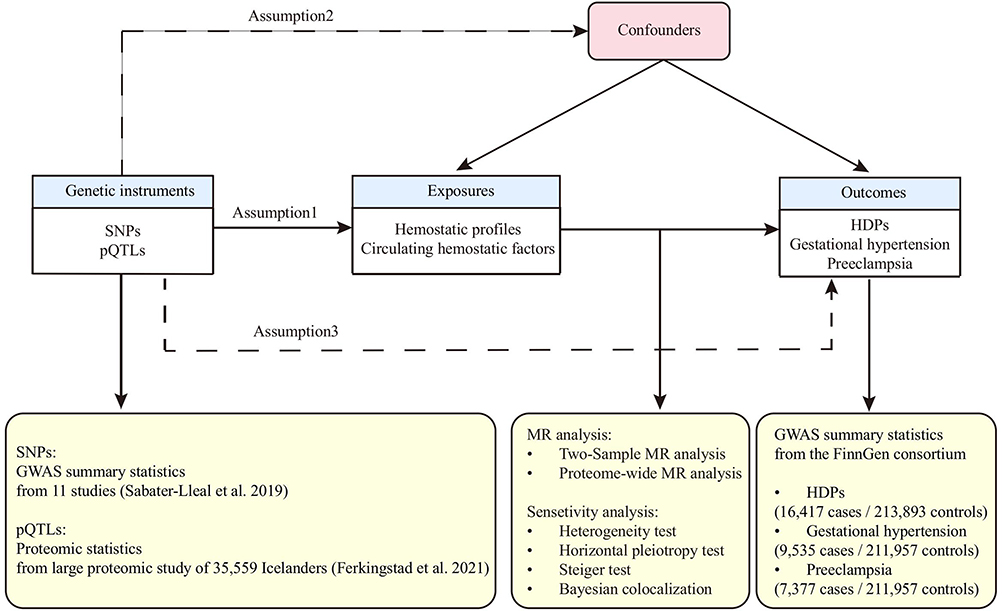 |
Figure 1 Study design of the two-sample Mendelian randomization analysis. The diagram illustrates the framework and three core assumptions of Mendelian randomization. Data sources for genetic instruments (SNPs for hemostatic profiles and pQTLs for circulating proteins), outcomes (HDPs from the FinnGen consortium), and the key analytical steps are detailed. Abbreviations: HDPs, hypertensive disorders of pregnancy; pQTL, protein quantitative trait locus. |
Genetic Instrumental Variable Selection for Hemostatic Profiles
We selected GWAS summary statistics of hemostatic profiles by systematically reviewing the literature to identify IVs for two-sample MR analysis. The selected hemostatic profiles include von Willebrand factor (vWF), a thrombospondin type 1 motif, member 13 (ADAMTS13), factor VII (FVII), prothrombin time (PT), factor VIII (FVIII), factor XI (FXI), activated partial thromboplastin time (APTT), factor X (FX), endogenous thrombin potential (ETP), protein C, and D-dimer, and the detailed information of hemostatic profiles is provided in Supplemental Table S2.17–26 Specifically, for studies involving multi-ethnic cohorts, we utilized summary statistics restricted to individuals of European ancestry to minimize population stratification bias.
To obtain valid IVs for MR analysis, IVs of hemostatic profiles were selected according to the following criteria: (1) SNPs are significantly correlated with specific hemostatic profiles (P < 5×10−8); (2) excluding rare SNPs (minor allele frequency < 0.01); (3) excluding SNP associated with other potential confounders (body mass index, lipid, thyroid disease, cardiovascular disease, and metabolic diseases, etc.) by searching PhenoScanner; (4) excluding SNP located in the major histocompatibility complex human leukocyte antigen region (chr6:28.4–33.4Mb); (5) excluding SNP was palindromic with intermediate allele frequencies. The F-statistic was calculated for each SNP to assess the strength of the instruments, with a threshold of F > 10 generally considered to indicate sufficient power to avoid weak instrument bias. The details of the included IVs of specific hemostatic profiles, their associations with HDPs are shown in Supplemental Tables S3 and S4.
Proteomic Data Source of Circulating Hemostatic Factors
The proteomic data for circulating hemostatic factors were available from a large-scale protein quantitative trait loci (pQTL) study of 35,559 Icelanders (https://www.decode.com/summarydata/).27 Proteomic data in this pQTL study was measured by a multiplexed, modified aptamer-based binding assay (SOMAscan version 4), and protein levels were rank-inverse normal transformed by age and gender. More details of this proteomic study can be found in the original publication.27 In the present study, the following criteria were used to identify valid pQTLs for specific circulating hemostatic factors: (1) SNPs are significantly correlated with specific circulating hemostatic factors (P < 5×10−8); (2) uncorrelated SNPs (clumped at correlation threshold r2 < 0.01); (3) excluding rare SNPs (minor allele frequency < 0.01); (4) SNPs with F statistic > 10 (R2=2×EAF×(1-EAF)×beta;2 F=R2×(N−2)/(1−R2)); (5) excluding the SNPs within the major histocompatibility complex human leukocyte antigen region (chr6:28.4–33.4Mb) due to complex linkage disequilibrium (LD) structure in this region. Further, the valid pQTLs were classified as cis- or trans-pQTLs according to the following criteria: a pQTL located within 1 Mb of the transcription start site of the protein-coding gene was defined as cis-pQTL, whereas a pQTL located outside this region was defined as trans-pQTL.
GWAS Summary Statistics of Hypertensive Disorders of Pregnancy
GWAS summary statistics for HDPs (HDPs as a whole and its 2 subtypes of gestational hypertension and preeclampsia) were extracted from the FinnGen consortium (Release 10, https://r10.finngen.fi/).28 The specific phenotypes utilized were “Hypertensive disorders of pregnancy” (O15_HYPTENSPREG), “Gestational hypertension” (O15_GESTAT_HYPERT), and “Preeclampsia” (O15_PREECLAMPS). The FinnGen consortium included 16,417 cases and 213,893 controls for HDPs, 9,535 cases and 211,957 controls for gestational hypertension, and 7,377 cases and 211,957 controls for preeclampsia, with a median age at first event between 28.74 and 29.74 years. These phenotypes in the FinnGen consortium were defined in accordance with the International Classification of Diseases code. It is important to note that the control group in the FinnGen consortium consists of all female participants in the cohort and is not restricted to pregnant women or those with a history of normotensive pregnancy.
Statistical Analysis
Mendelian Randomization Analyses
In two-sample MR analyses, we evaluated the causal associations between hemostatic factors and HDPs applying the random-effects inverse-variance weighted (IVW) method as the primary statistical model. To assess the robustness of the IVW estimates, we performed supplementary analyses employing the weighted median, MR-Egger, and MR-PRESSO methods for all exposures with at least three valid IVs. In cases of discordant results, we prioritized the IVW estimates, given their superior statistical power under balanced pleiotropy.
In proteome-wide MR analyses, both cis-pQTLs and trans-pQTLs were employed as IVs. However, we prioritized cis-pQTLs for primary analyses, as trans-pQTLs are more susceptible to violating key MR assumptions. Similarly, we designated the IVW method as the primary analytical approach, with weighted median, simple/weighted mode, and MR-Egger regression as supplementary analyses.
Sensitivity Analyses
We performed several sensitivity analyses to evaluate the reliability of our findings. The MR-Egger intercept and the MR-PRESSO global test were used to investigate potential horizontal pleiotropy. Heterogeneity across IVs was quantified using Cochran’s Q-test and visualized with funnel plots. Subsequently, we conducted leave-one-out analyses to assess the influence of individual SNPs. Finally, Steiger filtering was used to verify the assumed causal direction.
Colocalization Analysis
For the exposure-outcome pairs showing significant associations in the proteome-wide MR analyses, we conducted Bayesian colocalization analysis using the summary statistics for the specific cis-pQTLs and the corresponding HDP GWAS. This analysis aimed to test whether identified associations were driven by the same causal variant. This method evaluates five competing hypotheses (H0–H4) regarding the genetic architecture of the two traits within a given genomic region. H0: No SNP in the given genomic region is associated with two traits. H1/H2: SNPs are associated with only one trait. H3: Both traits are associated in the region, but distinct causal variants were for two traits. H4: Both traits are associated due to the same causal variant. A posterior probability for a shared causal variant (PPH4) of less than 0.8 is considered to indicate the absence of strong support for colocalization.
Statistical Software and Significance Thresholds
Two-sample MR analyses were conducted primarily using the TwoSampleMR package in R software (version 4.2.1). Sensitivity analyses were supplemented by the MRPRESSO package for outlier detection and correction. Bayesian colocalization analyses were performed with the coloc package. All reported P-values are two-sided. Given the multiple exposures tested, we considered a Bonferroni-corrected significance threshold to account for multiple testing. For the analysis of 11 hemostatic profiles, the corrected threshold was P < 0.0017 (0.05 / 30 tests), and for the 12 circulating proteins, it was P < 0.0014 (0.05 / 36 tests). Associations with a nominal P-value < 0.05 but above the corrected threshold were considered suggestive.
Results
Genetic Instrumental Variable for Hemostatic Profiles
For the first stage of our analysis, we selected genetic instruments for 11 clinically relevant hemostatic profiles from previously published GWAS (details in Supplementary Table S2). FX was subsequently analyzed only in the proteome-wide MR analysis. After rigorous screening, a final set of SNPs was selected for each profile, and the F-statistics of selected IVs were much greater than 10 (Supplementary Table S3), suggesting that those SNPs were strong instrumental variables. To assess potential violations of the MR assumptions, we examined the direct associations of the selected IVs with the outcomes. None of the instruments for the hemostatic profiles showed a significant association with HDPs or its subtypes (Supplementary Table S4), which provides some evidence against widespread horizontal pleiotropy.
In the proteome-wide MR analyses, we curated pQTLs associated with hemostatic factors including coagulation factors (FII, FV, FVII, FX, FXI, FXIII), vWF, ADAMTS13, fibrinogen gamma chain (FGG), plasminogen activator inhibitor (PAI), protein C, and tissue plasminogen activator (tPA) from a pQTL study involving 35,559 Icelandic individuals. After applying the screening criteria mentioned earlier to remove unqualified SNPs, we retained 170 cis-pQTLs and 350 trans-pQTLs for 12 circulating proteins as instruments for the proteome-wide MR analysis.
Associations of Hemostatic Profiles with Hypertensive Disorders of Pregnancy
To investigate the potential causal associations between hemostatic profiles and HDPs, we first performed comprehensive two-sample MR analyses using GWAS summary statistics for 11 hemostatic factors and HDPs. As shown in Figure 2 and Supplementary Table S5, genetically predicted higher vWF levels were nominally associated with an increased risk of preeclampsia. Specifically, per 1-standard deviation (SD) increase in vWF, the OR for preeclampsia was 1.27 (95% CI: 1.06–1.51, P = 0.008) in the primary IVW analysis. This finding was consistent across sensitivity analyses using the weighted median (OR: 1.26, 95% CI: 1.04–1.52, P = 0.016) and MR-PRESSO methods (OR: 1.27, 95% CI: 1.06–1.51, P = 0.022). However, it is important to note that this association was nominally significant and did not survive the strict Bonferroni-corrected threshold (P < 0.0017). No significant causal effects of vWF were observed on gestational hypertension or HDPs overall in the primary IVW analysis, although MR-PRESSO suggested a potential association with HDPs that may be influenced by outlier variants (OR: 1.31, P = 0.011).
 |
Figure 2 Forest plot of MR estimates for the effects of hemostatic profiles on HDPs and its subtypes. Odds ratios (ORs) are for the genetically predicted risk of (A) HDPs, (B) Gestational Hypertension, and (C) Preeclampsia per standard deviation increase in each hemostatic profile. Estimates were calculated using the inverse-variance weighted (IVW) method. P values for heterogeneity (Cochran’s Q) and pleiotropy (MR-Egger intercept) are shown. |
In addition, higher genetically predicted FVIII levels were associated with a higher risk of preeclampsia in both the IVW (per 1-SD increase, OR: 2.21, 95% CI: 1.05–4.66, P = 0.037) and weighted median analyses (OR: 2.59, 95% CI: 1.09–6.17, P = 0.032). We also identified a potential protective effect of higher Protein C levels on preeclampsia risk (IVW OR: 0.91, 95% CI: 0.83–0.99, P = 0.036). Similar to vWF, these associations did not persist after correction for multiple testing. No significant causal effects were detected for the other hemostatic profiles examined (ADAMTS13, APTT, FXI, PT, FVII, EPT, D-dimer) on any of the HDP outcomes (Figure 2 and Supplementary Table S5).
The robustness of these findings was confirmed through several sensitivity analyses. There was no evidence of horizontal pleiotropy from the MR-Egger intercept or MR-PRESSO global tests, nor was there significant heterogeneity according to Cochran’s Q tests (Table 1). Leave-one-out analyses indicated that the causal estimates were not driven by any single influential SNP, particularly for the association between vWF and preeclampsia (Supplementary Figures S1 to S3). Finally, Steiger filtering confirmed the assumed causal direction from hemostatic profiles to preeclampsia for all significant associations, minimizing the likelihood of reverse causation (Supplementary Table S6).
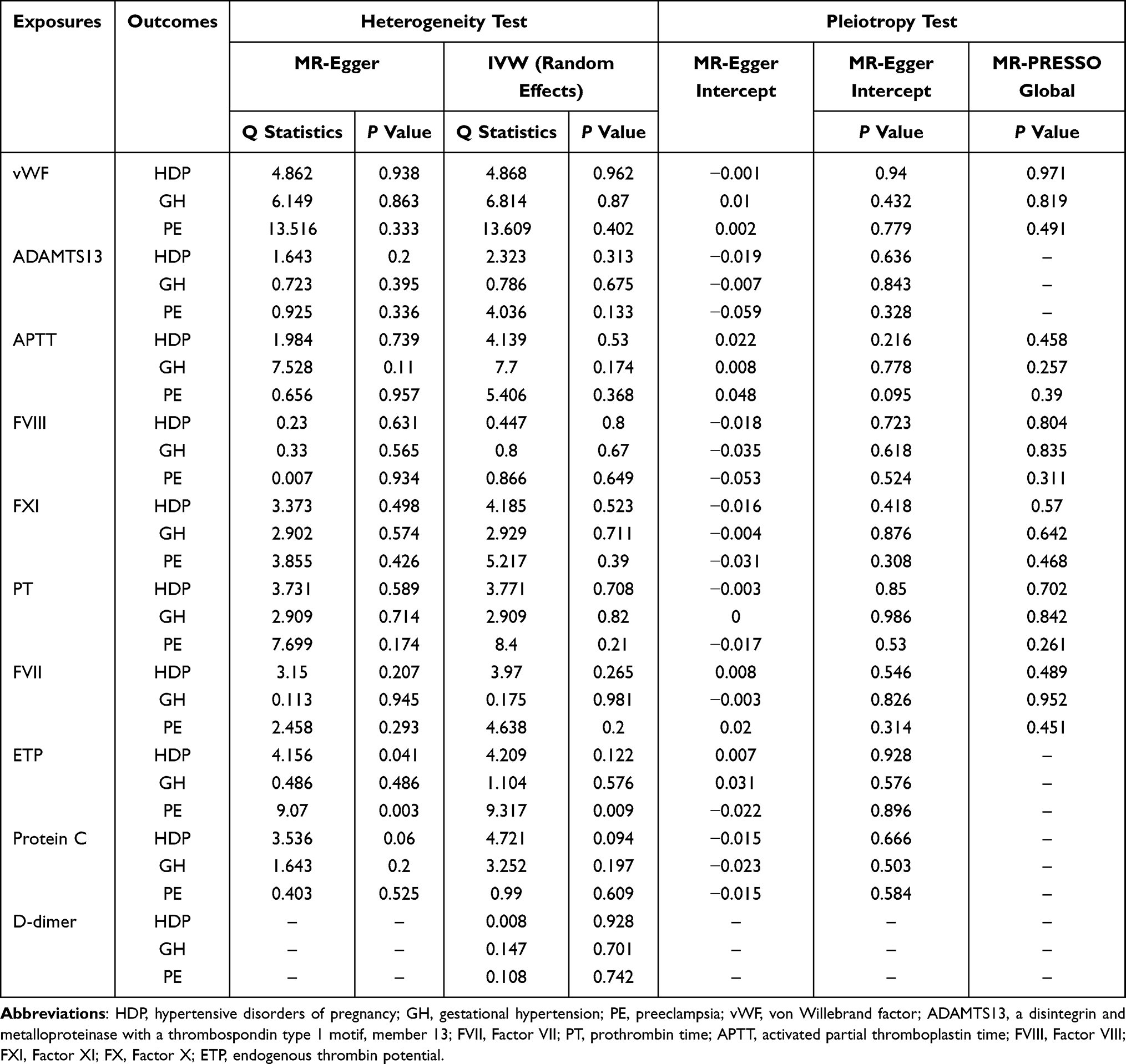 |
Table 1 MR Heterogeneity and Pleiotropy in Genetically Predicted Coagulation Factors and the Risk of Hypertensive Disorders of Pregnancy and Its Subtypes |
Associations of Circulating Hemostatic Factors with Hypertensive Disorders of Pregnancy
To further investigate the molecular drivers of HDPs, we performed a proteome-wide MR analysis using cis-pQTLs as genetic instruments. This analysis provided suggestive evidence consistent with the GWAS-based results for vWF, showing that higher genetically predicted circulating vWF levels were nominally associated with an increased risk of preeclampsia (IVW OR: 1.25, 95% CI: 1.00–1.57, P = 0.047) (Figure 3, Supplementary Figures S4 and S7). Furthermore, we identified novel nominal associations for two additional coagulation factors: higher circulating levels of both FII (IVW OR: 1.39, 95% CI: 1.01–1.91, P = 0.043) and FV (IVW OR: 1.12, 95% CI: 1.03–1.22, P = 0.010) were associated with an increased risk of preeclampsia (Supplementary Figures S5 and S6). None of these associations met the Bonferroni-corrected significance threshold. The detailed statistics for each genetic instrument used in this analysis are provided in Supplementary Table S8.
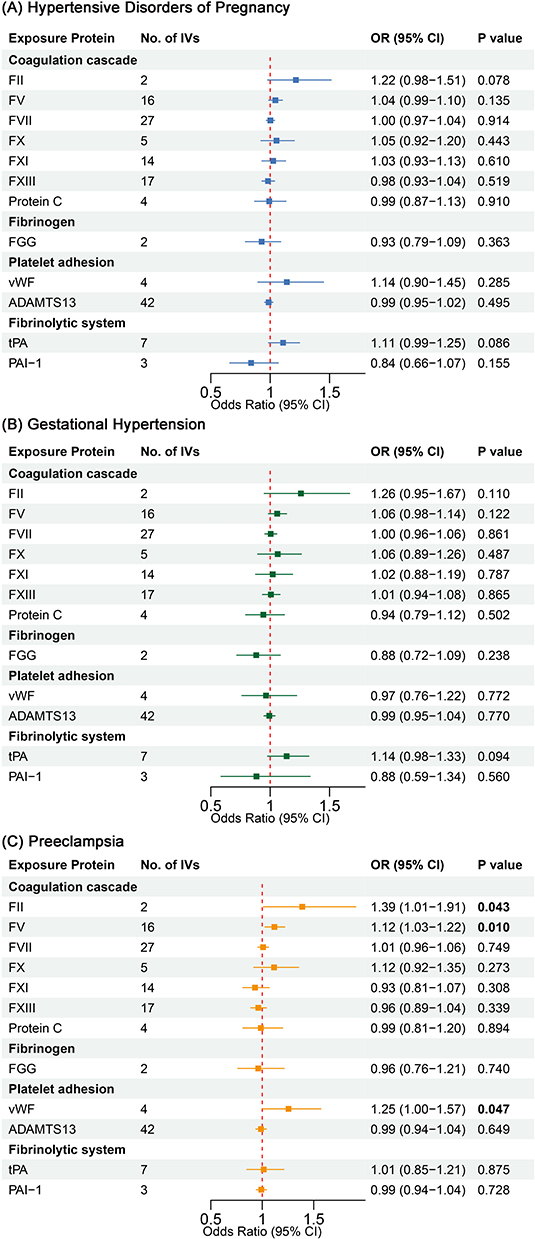 |
Figure 3 Forest plot of MR estimates for the effects of circulating hemostatic proteins on HDPs and its subtypes. ORs represent the genetically predicted risk of (A) HDPs, (B) Gestational Hypertension, and (C) Preeclampsia per standard deviation increase in protein level, estimated by the IVW method using cis-pQTLs as instruments. |
Additionally, using trans-pQTLs of circulating hemostatic factors, we identified nominal associations between circulating FGG levels and both hypertensive disorders of pregnancy and preeclampsia in IVW analyses (Supplementary Table S9). However, these associations may be susceptible to confounding and weak instrument bias due to the inherent limitations of trans-pQTLs.
The robustness of these findings was confirmed through comprehensive sensitivity analyses. Cochran’s Q tests revealed no significant heterogeneity in the instrumental variable estimates, indicating consistent effect sizes across genetic variants. MR-Egger intercept and MR-PRESSO global tests provided no evidence of horizontal pleiotropy that might bias our causal estimates (Table 2). Furthermore, Steiger filtering corroborated the directionality of the causal effects, effectively supporting that changes in circulating coagulation factor levels were not driven by the diseases (Supplementary Table S10).
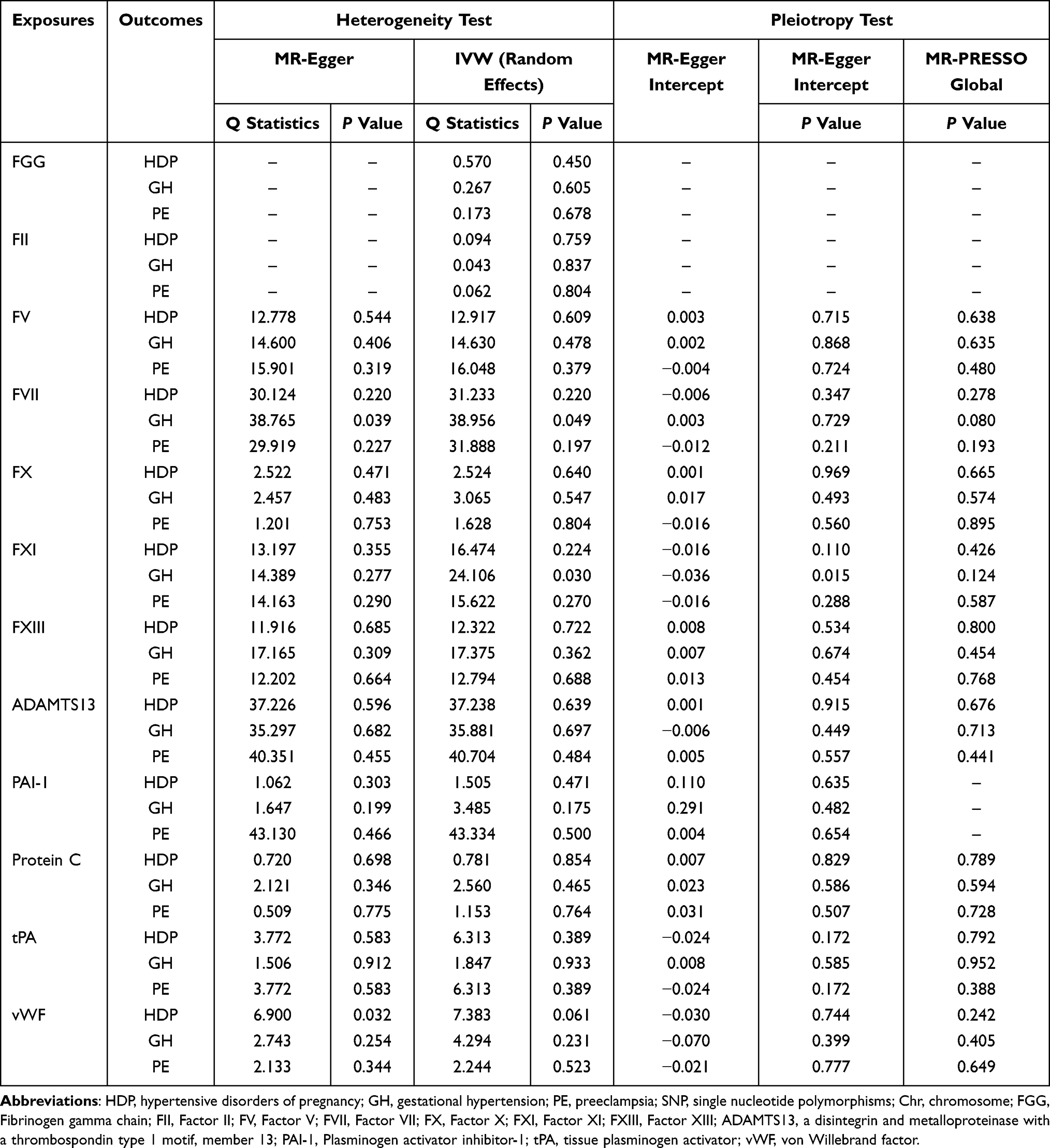 |
Table 2 MR Heterogeneity and Pleiotropy in Genetically Predicted Circulating Hemostatic Factors and the Risk of Hypertensive Disorders of Pregnancy and Its Subtypes |
Colocalization Analysis
To investigate whether the identified nominal MR associations were driven by shared causal variants, we performed Bayesian colocalization analysis for the vWF, FII, and FV loci using summary statistics from the cis-pQTL and HDP GWASs. The analysis revealed little evidence for a single shared causal variant (Hypothesis 4), with very low posterior probabilities across all three protein-phenotype pairs (PPH4: vWF=0.028, FII=0.034, FV=0.011). Instead, the results favored a model where the traits are associated with distinct causal variants within the same genomic region (Hypothesis 3) (PPH3: vWF=0.36, FII=0.38, FV=0.36) (Figure 4 and Supplementary Table S11). These findings suggest that while there are genetic signals for both hemostatic factors and preeclampsia in these loci, the MR associations may be driven by distinct variants in linkage disequilibrium rather than a single shared genetic effect, indicating a complex genetic architecture.
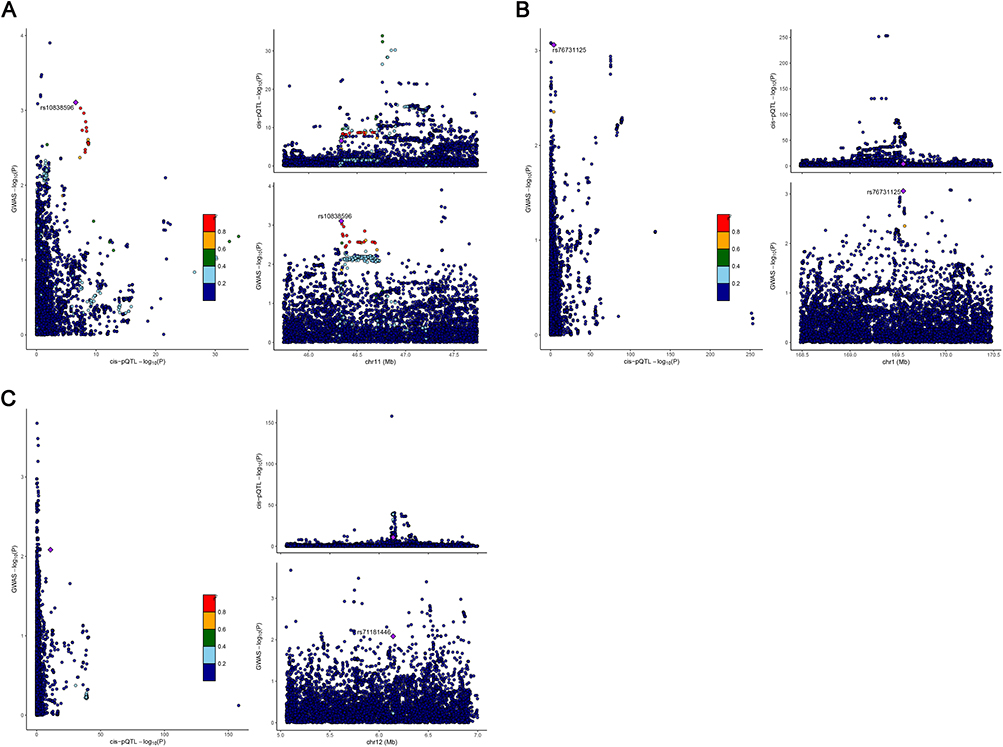 |
Figure 4 Locus-specific colocalization analyses for circulating hemostatic proteins and preeclampsia risk. Visualization of genetic associations at the encoding gene loci for (A) FII, (B) FV, and (C) vWF. For each panel, the left plot (LocusCompare) shows the -log10 P-values for the cis-pQTL (x-axis) versus the preeclampsia GWAS (y-axis). The right plots (LocusZoom) show regional association signals for the cis-pQTL (top) and preeclampsia GWAS (bottom), plotted against chromosomal position (x-axis). Each point represents a SNP, colored by its linkage disequilibrium (LD, r2) with the lead preeclampsia-associated SNP (purple diamond). |
Discussion
In the present study, two-sample MR analysis provided suggestive genetic evidence associating genetically predicted levels of vWF, FVIII, and Protein C with the risk of preeclampsia. Subsequently, proteome-wide MR analysis further indicated that genetically predicted circulating levels of FII, FV, and vWF were nominally associated with preeclampsia risk, utilizing IVs linked to pQTLs to provide inferences based on protein levels. However, it is important to note that subsequent colocalization analyses yielded low posterior probabilities for a single shared causal variant. This suggests that the observed associations may arise from a complex genetic architecture involving distinct functional variants in linkage disequilibrium rather than a single shared genetic pathway. Despite this complexity and the fact that results did not survive rigorous multiple-testing correction, our findings suggest a potential role for the hemostatic system in preeclampsia, which warrants further mechanistic investigation.
Preeclampsia is characterized by an excessive maternal inflammatory response to pregnancy, with generalized endothelial dysfunction as a core feature.29,30 Accumulating evidence indicates that extensive cross-talk between the coagulation cascade and inflammation plays a critical role in preeclampsia.8,31 Our MR analyses point towards a potential role for circulating vWF in preeclampsia, which aligns with its known biological functions. vWF not only serves as a marker of endothelial cells activation and damage, but also plays a direct role in regulating inflammatory responses and thrombosis.32 Moreover, the interaction between vWF and the complement system is also involved in the development of preeclampsia.33 In an in vitro study conducted by Youssef et al, significantly high deposition of vWF was observed in endothelial cells exposed to activated plasma from early-onset severe preeclampsia.8 Furthermore, increasing observational studies identified elevated plasma vWF levels as a risk factor for preeclampsia, which further supported our result.8,34–36 Importantly, vWF acts as a carrier molecule for FVIII, preventing its plasma clearance; consequently, elevated plasma levels of the vWF-FVIII complex contribute to thrombosis.32 Consistent with previous observational studies, our two-sample MR analysis also unveiled a nominal genetic association between FVIII and preeclampsia, which further reinforces the potential impact of the vWF-FVIII axis on this condition.34,37 However, the magnitude of these genetic associations should be interpreted with caution, given the potential for biological variability, population heterogeneity, and the inherent limitation of using lifelong genetic proxies to represent the dynamic physiological processes of pregnancy. Notably, preeclampsia is a multifactorial disorder involving immune, placental, and vascular systems. In this context, vWF represents one component within a broader pathophysiological network, and its role should not be interpreted in isolation from other contributing factors such as inflammation, placental dysfunction, and endothelial activation. Future studies integrating multi-omics approaches are needed to further elucidate how these systems interact to influence the risk and progression of preeclampsia.
Although we did not observe a significant genetic association between ADAMTS13 and HDPs, it is valuable to contextualize these findings within the broader spectrum of pregnancy-associated thrombotic microangiopathies (TMAs). Plasma ADAMTS13 is a metalloprotease responsible for cleaving ultra-large vWF multimers.38,39 The balance between ADAMTS13 and vWF is crucial for regulating thrombosis and inflammation. While preeclampsia does not exhibit the profound ADAMTS13 deficiency characteristic of thrombotic thrombocytopenic purpura (TTP), numerous studies have observed moderately reduced ADAMTS13 activity and elevated vWF antigen in preeclampsia patients compared to healthy controls.34,36,40 This suggests an overlap in pathophysiology involving endothelial activation and microvascular thrombosis. The discrepancy between these observational findings and our null MR results may be attributed to the limited number of strong genetic instruments for ADAMTS13 included in our study. Alternatively, it is possible that the reduction in ADAMTS13 activity observed in clinical settings is a secondary consequence of endothelial injury and high vWF consumption during the active phase of the disease, rather than a primary genetic driver initiating the condition. Future studies are needed to further investigate the interplay between ADAMTS13, vWF, and endothelial dysfunction in preeclampsia.
Of note, our colocalization analysis further revealed that the posterior probability of a shared causal variant was low across all tested protein-phenotype pairs, with relatively higher PPH3 values favoring distinct variants within the same loci. This finding raises the possibility that the causal associations may be partially influenced by linkage disequilibrium rather than reflecting a shared biological mechanism. Therefore, our MR results should be interpreted with caution. Further studies, including experimental models, are warranted to clarify the underlying mechanisms and to validate these findings.
There is currently no effective pharmacological treatment for preeclampsia, and aspirin remains the only preventive drug recommended by clinical guidelines, although its mechanism of action in this context is still incompletely understood.41 Aspirin is well established to inhibit platelet aggregation and promote vasodilation, while vWF plays a critical role in platelet adhesion and thrombus formation upon endothelial activation.42 The nominal genetic association between vWF and preeclampsia observed in our MR analysis provides a biological rationale that aligns with the known pharmacology of aspirin.
Our previous network meta-analysis suggested that both LMWH and aspirin may reduce the risk of preeclampsia, but the certainty of the evidence was low.12 The present study offers genetic evidence that several hemostatic factors, particularly vWF, FII and FV, are nominally associated with preeclampsia. The role of vWF in platelet adhesion and aggregation directly aligns with the antiplatelet action of aspirin. Meanwhile, FII and FV are key components of the thrombin generation pathway, which is specifically inhibited by LMWH.43 These genetic findings therefore provide a biological rationale and genetic evidence that is consistent with the observed clinical effects of aspirin and LMWH. It should be noted that MR estimates reflect lifelong genetic influences rather than the effects of acute pharmacological interventions. Thus, while our findings provide a biological rationale that aligns with the observed clinical effects of aspirin and LMWH, they do not constitute direct evidence for the efficacy of these drugs in clinical practice.
Previous observational studies only showed correlation, rather than causality, between coagulation factors and HDPs, and were affected by reverse causality and confounders. To the best of our knowledge, this is the first study to systematically investigate the causal association between hemostatic profiles and HDPs utilizing multi-omics MR method which provides estimates less prone to environmental confounding. Our study observed a nominal genetic association between elevated vWF levels and preeclampsia. While our colocalization analysis indicates that these associations likely arise from distinct functional variants in linkage disequilibrium rather than a single shared causal variant, the convergence of evidence from multiple analyses supports vWF as a potentially important factor in the disease. These findings offer insights into the etiology of preeclampsia and suggest that vWF may be a candidate for future investigation.
Limitations
Nonetheless, several limitations need to be acknowledged. First, this study focused on people of European ancestry, which limits the generalizability of our MR findings. Second, HDPs are female-specific conditions, yet the existing GWASs for coagulation factors were conducted in mixed-sex populations and did not provide sex-stratified data. This may introduce bias if the genetic regulation of hemostatic factors differs by sex. Third, the definition of HDP outcomes relied on ICD codes from electronic health records, which may be subject to phenotype misclassification. Furthermore, the control group in the FinnGen consortium included all female participants rather than being restricted to women with documented normotensive pregnancies. This broad control definition could potentially dilute the effect estimates and bias our results toward the null. Fourth, sensitivity analyses such as Steiger filtering and MR-PRESSO are more sensitive to gross violations of MR assumptions, while subtle residual pleiotropy may remain undetected. Therefore, although we applied rigorous sensitivity analyses, the possibility of horizontal pleiotropy or directional bias cannot be completely excluded. Finally, it is important to emphasize that while several associations were nominally significant, they did not survive a strict Bonferroni correction for multiple testing. This may be partly attributed to the limited sample sizes of the exposure GWASs, which could result in insufficient statistical power to detect weaker associations. Therefore, our findings should be interpreted as suggestive and require validation in larger-scale studies with more precise phenotyping.
Conclusion
In conclusion, this multi-omics Mendelian randomization study provides suggestive genetic evidence associated with several hemostatic factors, particularly vWF, in the pathophysiology of preeclampsia. Our findings suggest the potential involvement of the hemostatic system in the development of this disorder. These results suggest that vWF may be a candidate for further research. However, given the lack of robustness after multiple testing correction and absence of colocalization support, these findings should be interpreted with caution and require validation in future studies.
Data Sharing Statement
The original contributions made in this research are available within the article; any further questions should be addressed to the corresponding author.
Ethics Approval
This study used publicly available anonymized GWAS summary statistics. Under Article 32 of “Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects” (China, February 18, 2023), ethical review is not required for research using anonymized information data that does not cause harm, involve sensitive personal information, or have commercial interests. This study meets these criteria, and therefore formal Institutional Review Board approval was not required.
Acknowledgments
This study utilized publicly available summary data, and the authors gratefully acknowledge the FinnGen consortium, the deCODE study, and the various consortia responsible for the hemostatic profile GWAS data.
Funding
This work was supported by the Fundamental Research Funds for the Central Universities of Central South University (Grant No. 2025ZZTS0309 to Yeshen Zhang), the Scientific Research and Innovation Project of postgraduates in Hunan Province (Grant No. CX20250422 to Yeshen Zhang), the Natural Science Foundation of Hunan Province (Grant No. 2023JJ30819 to Xu Deng), and the National Natural Science Foundation of China (Grant No. 82570448 to Zhihui Zhang).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Say L, Chou D, Gemmill A, et al. Global causes of maternal death: a WHO systematic analysis. Lancet Glob Health. 2014;2(6):e323–15. doi:10.1016/s2214-109x(14)70227-x
2. Wu P, Green M, Myers JE. Hypertensive disorders of pregnancy. BMJ. 2023;381:e071653. doi:10.1136/bmj-2022-071653
3. Jiang L, Tang K, Magee LA, et al. A global view of hypertensive disorders and diabetes mellitus during pregnancy. Nat Rev Endocrinol. 2022;18:760–775. doi:10.1038/s41574-022-00734-y
4. Vousden N, Lawley E, Seed PT, et al. Incidence of eclampsia and related complications across 10 low- and middle-resource geographical regions: secondary analysis of a cluster randomised controlled trial. PLoS Med. 2019;16:e1002775. doi:10.1371/journal.pmed.1002775
5. McCance DR, Holmes VA, Maresh MJ, et al. Vitamins C and E for prevention of pre-eclampsia in women with type 1 diabetes (DAPIT): a randomised placebo-controlled trial. Lancet. 2010;376:259–266. doi:10.1016/s0140-6736(10)60630-7
6. Ives CW, Sinkey R, Rajapreyar I, Tita ATN, Oparil S. Preeclampsia-Pathophysiology and Clinical Presentations: JACC State-of-the-Art Review. J Am Coll Cardiol. 2020;76:1690–1702. doi:10.1016/j.jacc.2020.08.014
7. Rana S, Lemoine E, Granger JP, Karumanchi SA. Preeclampsia: pathophysiology, Challenges, and Perspectives. Circ Res. 2019;124:1094–1112. doi:10.1161/circresaha.118.313276
8. Youssef L, Miranda J, Blasco M, et al. Complement and coagulation cascades activation is the main pathophysiological pathway in early-onset severe preeclampsia revealed by maternal proteomics. Sci Rep. 2021;11:3048. doi:10.1038/s41598-021-82733-z
9. Burton GJ, Redman CW, Roberts JM, Moffett A. Pre-eclampsia: pathophysiology and clinical implications. BMJ. 2019;366:l2381. doi:10.1136/bmj.l2381
10. Chaiworapongsa T, Chaemsaithong P, Yeo L, Romero R. Pre-eclampsia part 1: current understanding of its pathophysiology. Nat Rev Nephrol. 2014;10:466–480. doi:10.1038/nrneph.2014.102
11. Davidson KW, Barry MJ, Mangione CM, et al. Aspirin Use to Prevent Preeclampsia and Related Morbidity and Mortality: US Preventive Services Task Force Recommendation Statement. JAMA. 2021;326:1186–1191. doi:10.1001/jama.2021.14781
12. Liu YH, Zhang YS, Chen JY, et al. Comparative effectiveness of prophylactic strategies for preeclampsia: a network meta-analysis of randomized controlled trials. Am J Obstet Gynecol. 2023;228:535–546. doi:10.1016/j.ajog.2022.10.014
13. Rolnik DL, Nicolaides KH, Poon LC. Prevention of preeclampsia with aspirin. Am J Obstet Gynecol. 2022;226:S1108–s1119. doi:10.1016/j.ajog.2020.08.045
14. Cruz-Lemini M, Vázquez JC, Ullmo J, Llurba E. Low-molecular-weight heparin for prevention of preeclampsia and other placenta-mediated complications: a systematic review and meta-analysis. Am J Obstet Gynecol. 2022;226:S1126–S1144. doi:10.1016/j.ajog.2020.11.006
15. Emdin CA, Khera AV, Kathiresan S. Mendelian Randomization. JAMA. 2017;318:1925–1926. doi:10.1001/jama.2017.17219
16. Skrivankova VW, Richmond RC, Woolf BAR, et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: the STROBE-MR Statement. JAMA. 2021;326:1614–1621. doi:10.1001/jama.2021.18236
17. Smith NL, Huffman JE, Strachan DP, et al. Genetic predictors of fibrin D-dimer levels in healthy adults. Circulation. 2011;123:1864–1872. doi:10.1161/circulationaha.110.009480
18. Tang W, Basu S, Kong X, et al. Genome-wide association study identifies novel loci for plasma levels of protein C: the ARIC study. Blood. 2010;116:5032–5036. doi:10.1182/blood-2010-05-283739
19. Rocanin-Arjo A, Cohen W, Carcaillon L, et al. A meta-analysis of genome-wide association studies identifies ORM1 as a novel gene controlling thrombin generation potential. Blood. 2014;123:777–785. doi:10.1182/blood-2013-10-529628
20. Sun BB, Maranville JC, Peters JE, et al. Genomic atlas of the human plasma proteome. Nature. 2018;558:73–79. doi:10.1038/s41586-018-0175-2
21. Suhre K, Arnold M, Bhagwat AM, et al. Connecting genetic risk to disease end points through the human blood plasma proteome. Nat Commun. 2017;8:14357. doi:10.1038/ncomms14357
22. Tang W, Schwienbacher C, Lopez LM, et al. Genetic associations for activated partial thromboplastin time and prothrombin time, their gene expression profiles, and risk of coronary artery disease. Am J Hum Genet. 2012;91:152–162. doi:10.1016/j.ajhg.2012.05.009
23. Goldstein JA, Weinstock JS, Bastarache LA, et al. LabWAS: novel findings and study design recommendations from a meta-analysis of clinical labs in two independent biobanks. PLoS Genet. 2020;16:e1009077. doi:10.1371/journal.pgen.1009077
24. de Vries PS, Sabater-Lleal M, Huffman JE, et al. A genome-wide association study identifies new loci for factor VII and implicates factor VII in ischemic stroke etiology. Blood. 2019;133:967–977. doi:10.1182/blood-2018-05-849240
25. Ma Q, Jacobi PM, Emmer BT, et al. Genetic variants in ADAMTS13 as well as smoking are major determinants of plasma ADAMTS13 levels. Blood Adv. 2017;1:1037–1046. doi:10.1182/bloodadvances.2017005629
26. Sabater-Lleal M, Huffman JE, de Vries PS, et al. Genome-Wide Association Transethnic Meta-Analyses Identifies Novel Associations Regulating Coagulation Factor VIII and von Willebrand Factor Plasma Levels. Circulation. 2019;139:620–635. doi:10.1161/circulationaha.118.034532
27. Ferkingstad E, Sulem P, Atlason BA, et al. Large-scale integration of the plasma proteome with genetics and disease. Nat Genet. 2021;53:1712–1721. doi:10.1038/s41588-021-00978-w
28. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613:508–518. doi:10.1038/s41586-022-05473-8
29. Redman CW, Sacks GP, Sargent IL. Preeclampsia: an excessive maternal inflammatory response to pregnancy. Am J Clin Exp Obstet Gynecol. 1999;180:499–506.
30. Szpera-Gozdziewicz A, Breborowicz GH. Endothelial dysfunction in the pathogenesis of pre-eclampsia. Front Biosci. 2014;19:734–746.
31. Levi M, van der Poll T. Inflammation and coagulation. Crit Care Med. 2010;38:S26–S34. doi:10.1097/CCM.0b013e3181c98d21
32. Atiq F, O’Donnell JS. Novel functions for von Willebrand factor. Blood. 2024;144:1247–1256. doi:10.1182/blood.2023021915
33. Gardikioti A, Venou T-M, Gavriilaki E, et al. Molecular Advances in Preeclampsia and HELLP Syndrome. Int J Mol Sci. 2022;23:3851. doi:10.3390/ijms23073851
34. Alpoim PN, Gomes KB, Godoi LC, et al. ADAMTS13, FVIII, von Willebrand factor, ABO blood group assessment in preeclampsia. Clin Chim Acta. 2011;412:2162–2166. doi:10.1016/j.cca.2011.07.030
35. Molvarec A, Rigó J, Bõze T, et al. Increased plasma von Willebrand factor antigen levels but normal von Willebrand factor cleaving protease (ADAMTS13) activity in preeclampsia. Thromb Haemost. 2009;101:305–311.
36. Ahmad A, Mustafa G, Mazari N, Naveed MA. Comparison of thrombomodulin, vWF, and ADAMTS13 levels between preeclampsia and normal pregnancy. J Pak Med Assoc. 2024;74:38–42. doi:10.47391/JPMA.7537
37. Witsenburg CPJ, Rosendaal FR, Middeldorp JM, Van der Meer FJM, Scherjon SA. Factor VIII levels and the risk of pre-eclampsia, HELLP syndrome, pregnancy related hypertension and severe intrauterine growth retardation. Thromb Res. 2005;115:387–392.
38. Zheng XL. Structure-function and regulation of ADAMTS-13 protease. J Thromb Haemost. 2013;11(Suppl 1):11–23. doi:10.1111/jth.12221
39. J-f D, Moake JL, Nolasco L, et al. ADAMTS-13 rapidly cleaves newly secreted ultralarge von Willebrand factor multimers on the endothelial surface under flowing conditions. Blood. 2002;100:4033–4039.
40. Stepanian A, Cohen-Moatti M, Sanglier T, et al. Von Willebrand factor and ADAMTS13: a candidate couple for preeclampsia pathophysiology. Arteriosclerosis Thrombosis Vasc Biol. 2011;31:1703–1709. doi:10.1161/ATVBAHA.111.223610
41. Chappell LC, Cluver CA, Kingdom J, Tong S. Pre-eclampsia. Lancet. 2021;398:341–354. doi:10.1016/S0140-6736(20)32335-7
42. Neubauer K, Zieger B. Endothelial cells and coagulation. Cell Tissue Res. 2022;387:391–398. doi:10.1007/s00441-021-03471-2
43. Versteeg HH, Heemskerk JWM, Levi M, Reitsma PH. New fundamentals in hemostasis. Physiol Rev. 2013;93:327–358. doi:10.1152/physrev.00016.2011
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Relationship of Placental and Serum Lipoprotein-Associated Phospholipase A2 Levels with Hypertensive Disorders of Pregnancy
Wang J, Dong X, Wu HY, Bu WH, Cong R, Wang X, Shang LX, Jiang W
International Journal of Women's Health 2022, 14:797-804
Published Date: 17 June 2022
Reviewing Accuracy of First Trimester Screening for Preeclampsia Using Maternal Factors and Biomarkers
Malone SL, Haj Yahya R, Kane SC
International Journal of Women's Health 2022, 14:1371-1384
Published Date: 19 September 2022
Genetic Insights Into Lipid Traits and Lipid-Modifying Drug Targets in Pregnancy Complications: A Two-Sample Mendelian Randomization Study
Shao H, Xu C, Zhang C, Li L, Wu P, Chen Z, Guan R
International Journal of Women's Health 2025, 17:221-234
Published Date: 31 January 2025
Investigating Serum and Placental Levels of IGF-1 and IGF-1R in Preeclampsia Patients and Their Clinical Implications
Su J, Huang X, Meng S, Wang S
International Journal of Women's Health 2025, 17:729-738
Published Date: 13 March 2025
Endometriosis Severity and Risk of Preeclampsia: A Combined Mendelian Randomization and Observational Study
Zu Y, Xie Y, Zhang H, Chen L, Yan S, Wang Z, Fang Z, Lin S, Yan J
International Journal of Women's Health 2025, 17:923-935
Published Date: 27 March 2025