Back to Journals » OncoTargets and Therapy » Volume 8
Association between EHBP1 rs721048(A>G) polymorphism and prostate cancer susceptibility: a meta-analysis of 17 studies involving 150,678 subjects
Authors Ao X, Liu Y, Bai X, Qu X, Xu Z, Hu G, Chen M, Wu H
Received 5 March 2015
Accepted for publication 18 May 2015
Published 7 July 2015 Volume 2015:8 Pages 1671—1680
DOI https://doi.org/10.2147/OTT.S84034
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Jianmin Xu
Xiang Ao,1,* Ying Liu,1,* Xiao-Yan Bai,1 Xinjian Qu,2 Zhaowei Xu,1 Gaolei Hu,1 Min Chen,1 Huijian Wu1,2
1Laboratory of Molecular Medicine & Pharmacy, School of Life Science and Biotechnology, Dalian University of Technology, Dalian, 2Laboratory of Molecular Medicine & Pharmacy, School of Life Science and Medicine, Dalian University of Technology, Panjin, Liaoning, People’s Republic of China
*These authors have contributed equally to this work
Background: EHBP1 rs721048(A) was first identified as a prostate cancer (PCa) risk in Caucasians by genome-wide association study, but subsequent replication studies involving Caucasian and other ethnicities did not produce consistent results. The aim of this study was to obtain a more definite association between rs721048(A) and PCa risk.
Methods: We comprehensively searched several databases updated to September 2014, including PubMed, Web of Science, EBSCO, and Google Scholar. Two authors independently screened and reviewed the eligibility of each study. The quality of the included studies was assessed by the Newcastle–Ottawa scale. The association of rs721048(A) and PCa risk was assessed by pooling odds ratios (ORs) with 95% confidence intervals (CIs).
Results: A total of 17 studies, including 48,135 cases and 102,543 controls, published between 2008 and 2014 were included in the meta-analysis. Overall, the pooled analysis demonstrated that rs721048(A) was significantly associated with the risk of PCa under the allele model (OR=1.14, 95% CI=1.11–1.17, P=0.000). Subgroup analysis based on ethnicity revealed a significant association between rs721048(A) and PCa in Caucasian (OR=1.14, 95% CI=1.11–1.16, P=0.000), African descent (OR=1.11, 95% CI=1.01–1.23, P=0.025), and Asian (OR=1.35, 95% CI=1.12–1.64, P=0.002).
Conclusion: Our results provided strong evidence that rs721048(A) could be a risk factor for PCa.
Keywords: EHBP1, rs721048, meta-analysis, prostate cancer
Introduction
Prostate cancer (PCa) is a common malignant tumor of the urinary system and the most frequently diagnosed nonskin cancer in men, with over 659,000 cases in developed countries.1–4 The incidence of PCa has increased as a result of increased awareness and screening.5 The highest PCa incidence rates occur in developed countries, such as Australia and New Zealand, Western and Northern Europe, and Northern America, and the highest mortality rates are among those of African descent in the Caribbean region, whereas the lowest incidence and mortality rates occur in South-Central Asia.6 Several risk factors, such as androgen, obesity, diet, smoking, alcohol, race, family history, and advancing age, are associated with the occurrence of PCa. Among them, race, family history of the disease, and advancing age are the only established risk factors for PCa,7 indicating genetics as a key factor in the risk of PCa. However, the detailed pathogenesis of this disease is still unclear. Therefore, more researches are now in progress to look for new risk factors for PCa. Genome-wide association study (GWAS), also known as whole genome association study (WGAS), is widely used in the study of genetic epidemiology. At present, more than 50 susceptibility loci associated with PCa risk have been identified by GWAS. Most of them are located in the introns and noncoding regions of the genes. Among them, eight loci were identified in the Japanese,8,9 one locus was identified in African-Americans,10 and two loci were identified in the Chinese,11 while other susceptibility loci were identified in Europeans.12–22
EH domain protein 2 (EHD2) binding protein 1 (EHBP1) was first identified and cloned in 3T3-L1 adipocytes, containing a calponin homology domain and five NPF motifs.23 EHBP1 connects clathrin-mediated endocytosis to the actin cytoskeleton and plays an important role in insulin-stimulated GLUT4 movements and hexose transportation by interacting with EHD2 protein through its NPF motifs.23,24 Upregulation of EHBP1 in colorectal cancer is related to the survival of colorectal cancer patients. Moreover, EHBP1 can also mediate the anti-invasive effect of atorvastatin in PTEN-expressing PCa cells.25
rs721048 is an A/G variation, which is located within one of the introns of the EHBP1 gene on chromosome 2p15, and rs721048(A) was first identified as a risk factor for PCa in Caucasians.17 Subsequently, extensive replication studies examined the association between rs721048(A>G) polymorphism and PCa risk in Caucasians and other ethnicities, such as those of African descent, Asians, Latinos, Native Hawaiians, and those of Japanese descent, but these studies yielded inconsistent results. Only seven studies yielded similar trends with GWAS analysis, whereas the other nine studies showed no association of rs721048(A) with the risk of PCa. These inconsistent results may have resulted from the different ethnicities in these replication studies. Among them, eight studies assessed the association of rs721048(A) with PCa risk in Caucasians. Four studies yielded similar results to those obtained with GWAS analysis,20,26–28 whereas the other four studies yielded opposite results.29–32 The first replication study conducted by Waters et al showed an association of rs721048(A) with PCa risk in Latinos, but not in Caucasians, those of African descent, of Japanese descent, and Native Hawaiians.29 Another GWAS conducted by Gudmundsson et al identified four new variants associated with PCa risk. Moreover, one replication study further confirmed that rs721048(A) is a risk factor for PCa in Caucasians.20 Six studies evaluated the association of rs721048(A) with PCa risk in African descent,29,33–37 but only one study yielded positive result.34 Three studies evaluated the association of rs721048(A) with PCa risk in Asians,38–40 but only one yielded positive result.40 Na et al demonstrated that rs721048(A) is a risk factor for PCa and demonstrated that rs721048(A) is associated with high-grade PCa (Gleason score ≥8) in Chinese.40 Another cause of the inconsistent results could be the small sample size of subjects. There were only 443 samples in Zheng et al’s,38 472 in Wang et al’s,36 1,176 in Nguyen Bentzon et al’s,32 and 1,746 in Xu et al’s35 study. In addition, publication bias may exist. These three points are likely to be the cause of the inconsistent results.41 Therefore, we wanted to confirm whether rs721048(A) is a risk factor for PCa.
rs721048(A) was first identified as a risk factor for PCa in Caucasians, but subsequent replication studies in Caucasians yielded inconsistent results and the replication studies in other ethnicities (such as Asian and African descent) also yielded inconsistent results. Thus, we performed a pooled meta-analysis to obtain a more precise prediction for the association between rs721048A>G polymorphism and PCa risk. To the best of our knowledge, our study was the first comprehensive meta-analysis to assess the association of rs721048(A) with PCa risk. In addition, we also evaluated the association between rs721048(A) and PCa risk in different ethnicities in subgroup analysis.
Materials and methods
Search strategy
The meta-analysis was performed according to the PRISMA guidelines.42 We searched the articles using the following terms and keywords: “EHD2-binding protein 1” or “EHBP1” or “rs721048” and “prostate cancer” in academic databases, including PubMed, EBSCO, Web of Science, and Google Scholar (last search was dated September 21, 2014). Additionally, reference lists were hand searched from retrieved articles and reviews. Articles were limited to English language. Two investigators (Xiang Ao and Ying Liu) read the abstracts of the retrieved articles to check all relevant studies independently. After that, all investigators further reviewed the full-text articles to identify all appropriate articles upon inclusion and exclusion criteria. Disagreements were solved by discussion among all authors, until an agreement was reached by all parties.
Inclusion and exclusion criteria
To be eligible for inclusion in this meta-analysis, the included studies must meet all the following inclusion criteria: 1) articles about the association between EHBP1 rs721048(A>G) polymorphism and PCa risk; 2) case-control studies or cohort studies; 3) studies providing sufficient published data of odds ratio (OR) with 95% confidence interval (CI). Studies were excluded due to the following reasons: 1) reviews; 2) case-only studies; 3) studies about the association between rs721048(A) and PCa progression; 4) studies about the association between rs721048(A) and Gleason grade and stage; 5) the genotype distributions in the controls deviated from Hardy–Weinberg equilibrium (P≤0.05).
Data extraction
Data extraction was performed by two investigators (Xiang Ao and Ying Liu) independently. The following data were extracted: first author’s name, publication year, sample size, country and ethnicity of case-controls, source of control, genotyping method and OR (95% CI). The quality of the included studies was assessed by the Newcastle–Ottawa scale (NOS) (http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp), including selection of groups, comparability of groups, and ascertainment of exposure. The range of NOS score was from 0 to 10 stars. Studies with NOS score above 5 stars were included in the final analysis.
Statistical analysis
Data were analyzed using Stata 12.0. The strength of association between rs721048(A) and PCa risk was measured by OR and 95% CI. Heterogeneity of the included studies was examined by Cochran’s Q test. If no heterogeneity was detected in the studies (P>0.1), the fixed-effect model was chosen to calculate the pooled OR and 95% CI. In contrast, if heterogeneity was detected in the studies (P≤0.1), the random effect model was adopted. I2 statistical test was used to calculate the degree of heterogeneity (I2=0%–25%, no heterogeneity; I2=25%–50%, moderate heterogeneity; I2=50%–75%, large heterogeneity; I2=75%–100%, extreme heterogeneity). For further subgroup analysis, similar method was used to assess the association between rs721048(A) and the risk of PCa in different ethnicities. In the subgroup analysis, different ethnicities or subsets in the study were recognized as individual studies. One-way sensitivity analysis was carried out to evaluate the stability of the results. Cumulative meta-analysis was performed to reflect the trends of the results induced by publication year and sample size. All included studies were ranked according to publication year or sample size. These analyses were performed by adding one new study at a time. Publication bias was examined by the Egger’s test and Begg’s funnel plot. z<1.96 or P>0.05, indicated no publication bias. Otherwise, publication bias was considered.
Results
Characteristics of eligible studies
The process of including and excluding articles with specific reasons is described in Figure 1. A total of 182 articles were obtained through literature search, each of which was relevant to the search terms. Among them, 65 articles were duplicated. After reading the abstracts of the remaining 117 articles, 77 articles were excluded because they did not meet our specified criteria. The full text of the remaining 40 articles was further checked. The reasons for exclusion are listed in Figure 1. Only 17 articles, published between 2008 and 2014, were retained for the meta-analysis because they met our criteria. These articles included a total of 48,135 cases and 102,543 controls. The main characteristics of the 17 eligible studies are summarized in Table 1. GWAS analysis conducted by Gudmundsson et al initially identified the association of rs721048A>G polymorphism with PCa risk.17 The other 16 articles were replication studies at the population level. Overall, 17 studies were conducted in five different regions, including Europe, USA, UK, Australia, and People’s Republic of China. On the level of ethnicity, the 17 included studies were classified according to previous descriptions:43,44 nine studies were of Caucasians (White), four were Asians, six were African descent, one was Japanese descent, one was a Native Hawaiian, and one was the Latino population. Six studies had a NOS score of 8 stars, whereas the other eleven studies had a NOS score of 7 stars. Since most studies only provided an adjusted OR with 95% CI under the allele model, without providing the details of the genotypes in the cases and controls, we extracted the adjusted OR with 95% CI under the allele model in our meta-analysis.
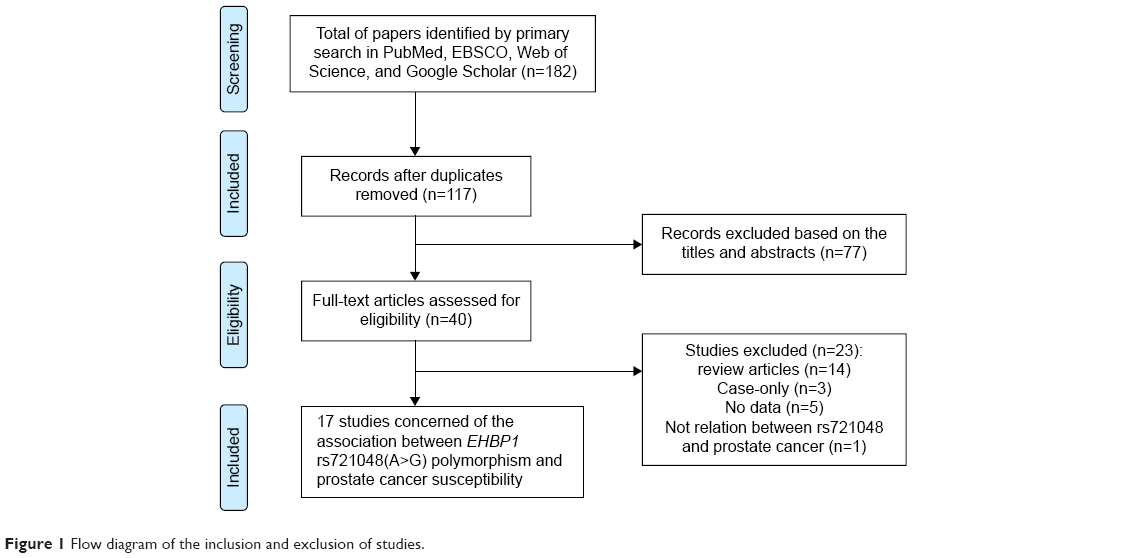 | Figure 1 Flow diagram of the inclusion and exclusion of studies. |
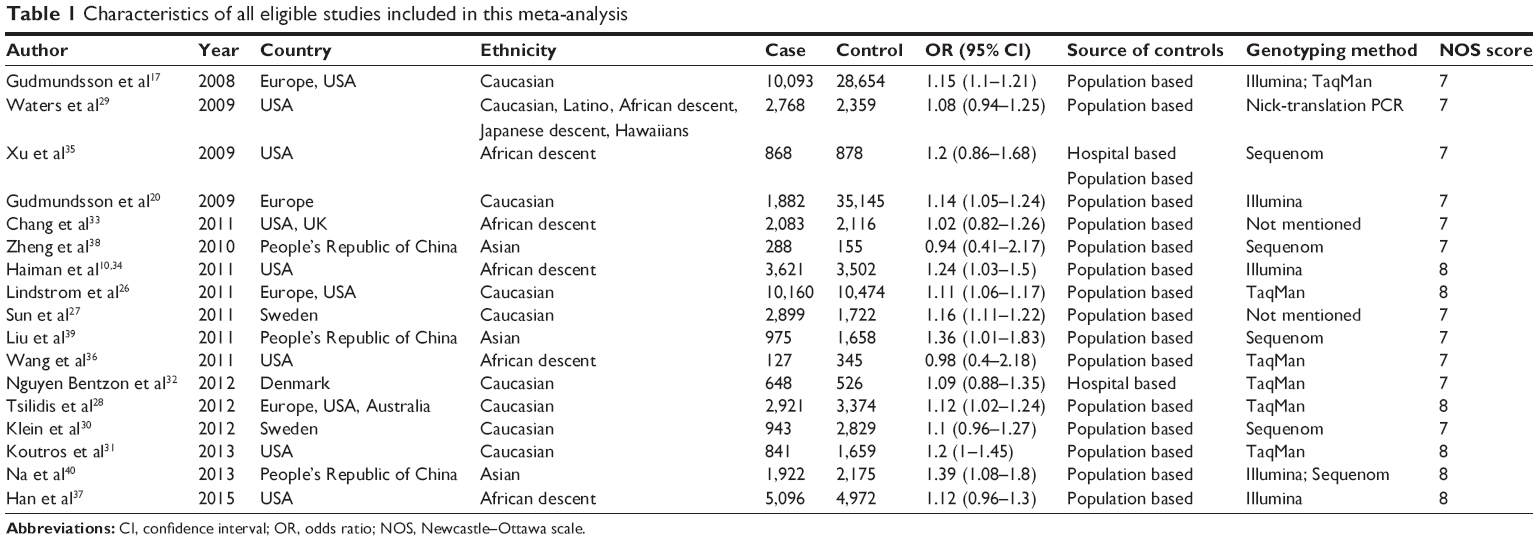 | Table 1 Characteristics of all eligible studies included in this meta-analysis |
Overall analysis of data and subgroup analysis
A total of 48,135 cases and 102,543 controls were included in this meta-analysis. Given that each of the 17 studies only provided one adjusted OR with 95% CI under the allele model, we calculated the pooled OR under the allele model to assess the association between rs721048A>G polymorphism and PCa risk. The heterogeneity among the 17 studies was measured by Cochran’s Q test. No heterogeneity was observed among these studies (I2=0.0%, P=0.909) (Figure 2). Therefore, the meta-analysis was performed in a fixed-effect model. A highly significant association was observed between EHBP1 rs721048A>G polymorphism and PCa risk in the allele model (OR=1.14, 95% CI=1.11–1.17) (Figure 2). Considering that rs721048(A) was a risk factor for PCa in Caucasian as revealed by GWAS analysis, we performed a subgroup analysis according to ethnicity to examine whether rs721048(A) is also a risk factor in other ethnicities. A significant association between rs721048(A) and the risk of PCa was observed in the subgroups of Caucasian (OR=1.14, 95% CI=1.11–1.16), African descent (OR=1.11, 95% CI=1.01–1.23), and Asian population (OR=1.35, 95% CI=1.12–1.64) under the allele model. In addition, no heterogeneity was observed (Figure 3).
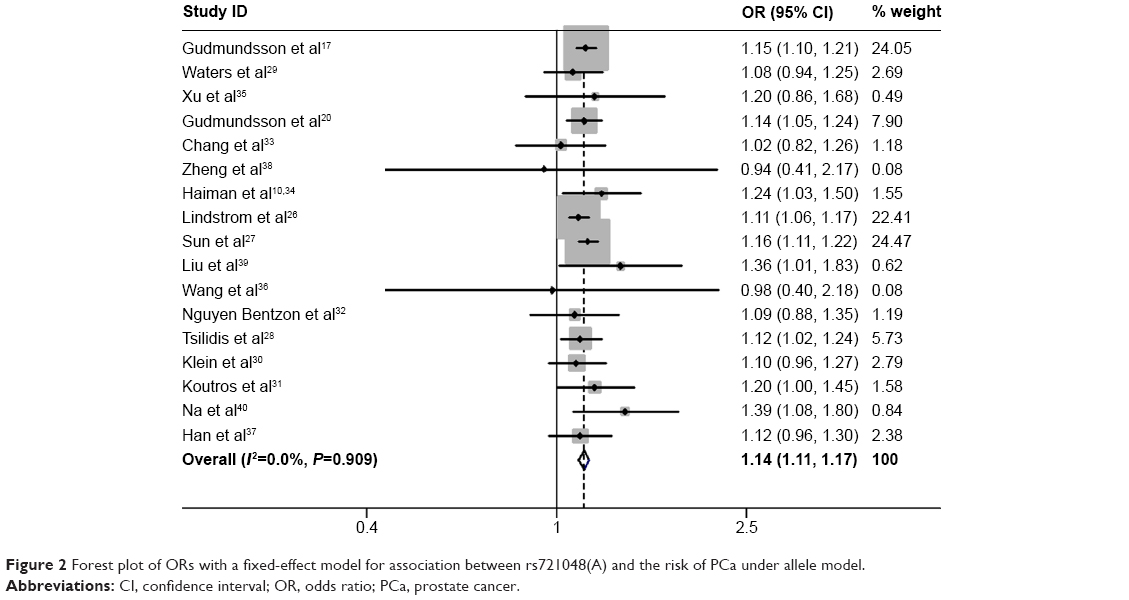 | Figure 2 Forest plot of ORs with a fixed-effect model for association between rs721048(A) and the risk of PCa under allele model. |
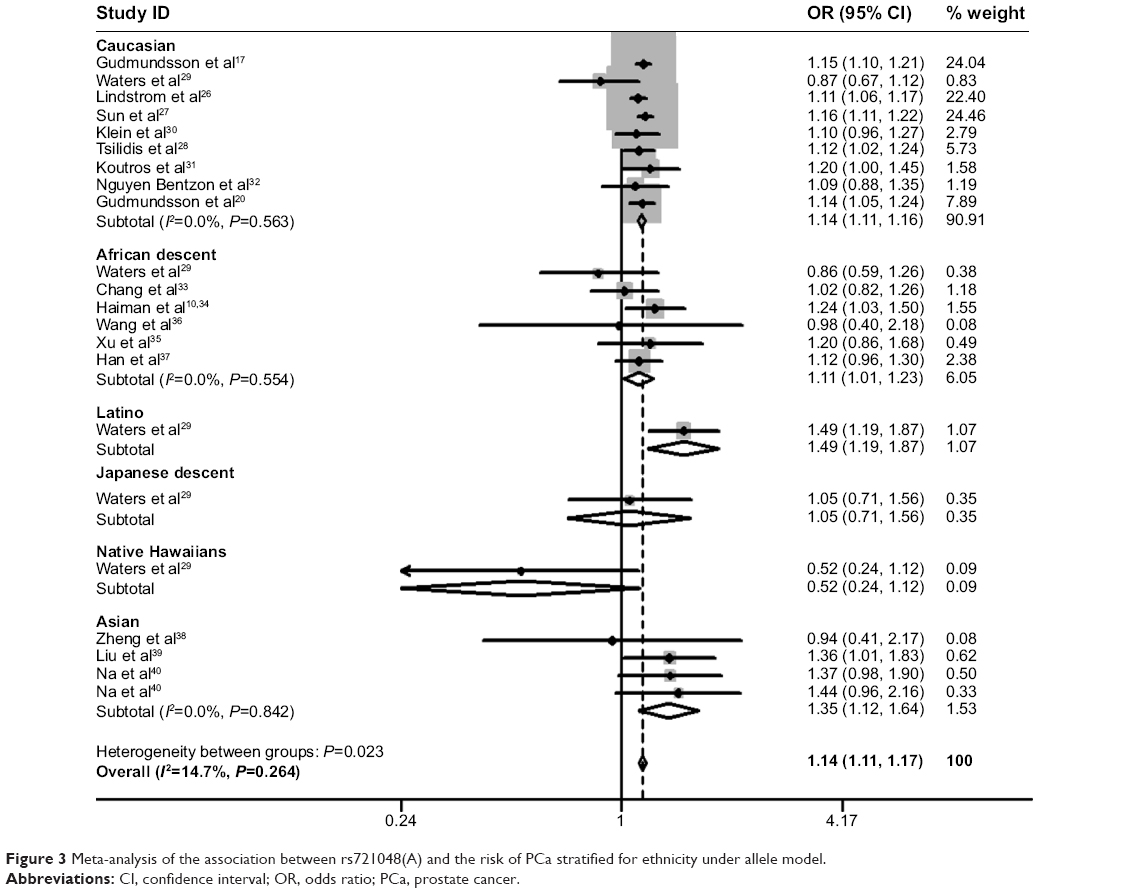 | Figure 3 Meta-analysis of the association between rs721048(A) and the risk of PCa stratified for ethnicity under allele model. |
Sensitivity analysis
In order to evaluate the stability of the results, one-way sensitivity analysis was performed. As shown in Figure 4, when one study was omitted in sequence, no obvious change was observed for the pooled OR and 95% CI, further confirming that rs721048(A) polymorphism was a risk factor for PCa.
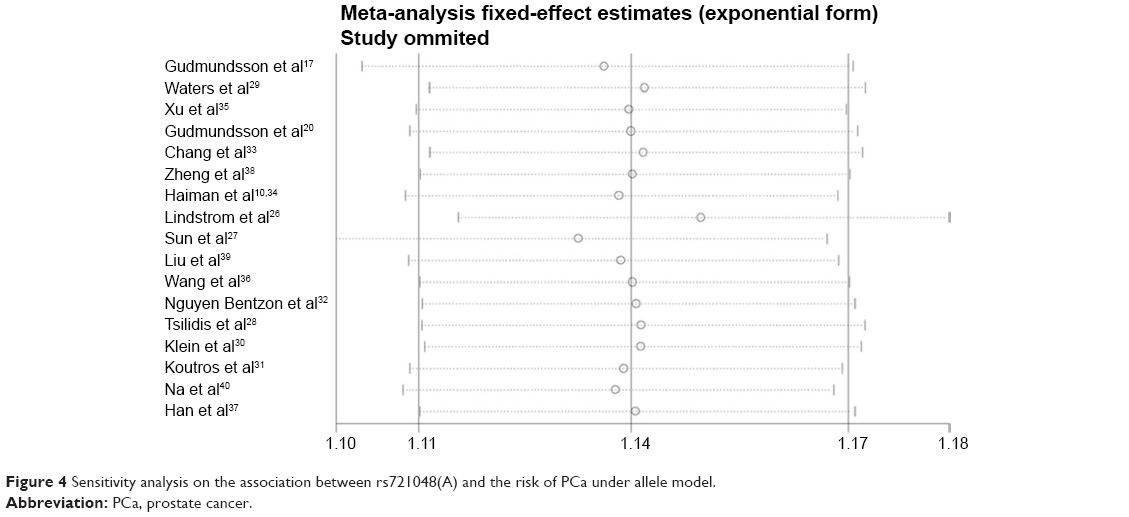 | Figure 4 Sensitivity analysis on the association between rs721048(A) and the risk of PCa under allele model. |
Cumulative meta-analysis
Cumulative meta-analyses were conducted to elucidate the variation in ORs and 95% CIs and to find out at which point of the analysis the risk between rs721048(A) polymorphism and PCa would become statistically significant through sorting studies according to publication year or sample size. As shown in Figure 5, rs721048(A) was further confirmed as a significant risk factor for PCa, and this association was not significantly affected by the changes in publication years. Cumulative meta-analysis by sample size showed that the pooled result began to reach statistical significance after the fifth study included (a total number of 2,772 samples), and the range of CIs became more defined with the increased population sizes in the included studies (Figure 6).
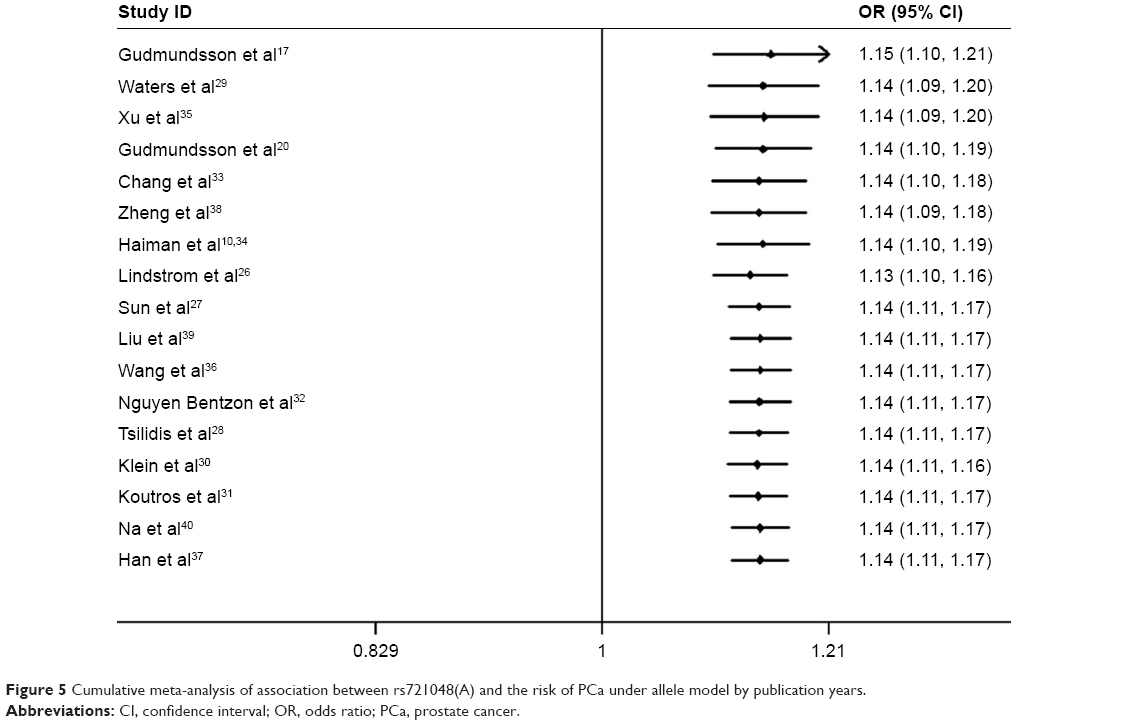 | Figure 5 Cumulative meta-analysis of association between rs721048(A) and the risk of PCa under allele model by publication years. |
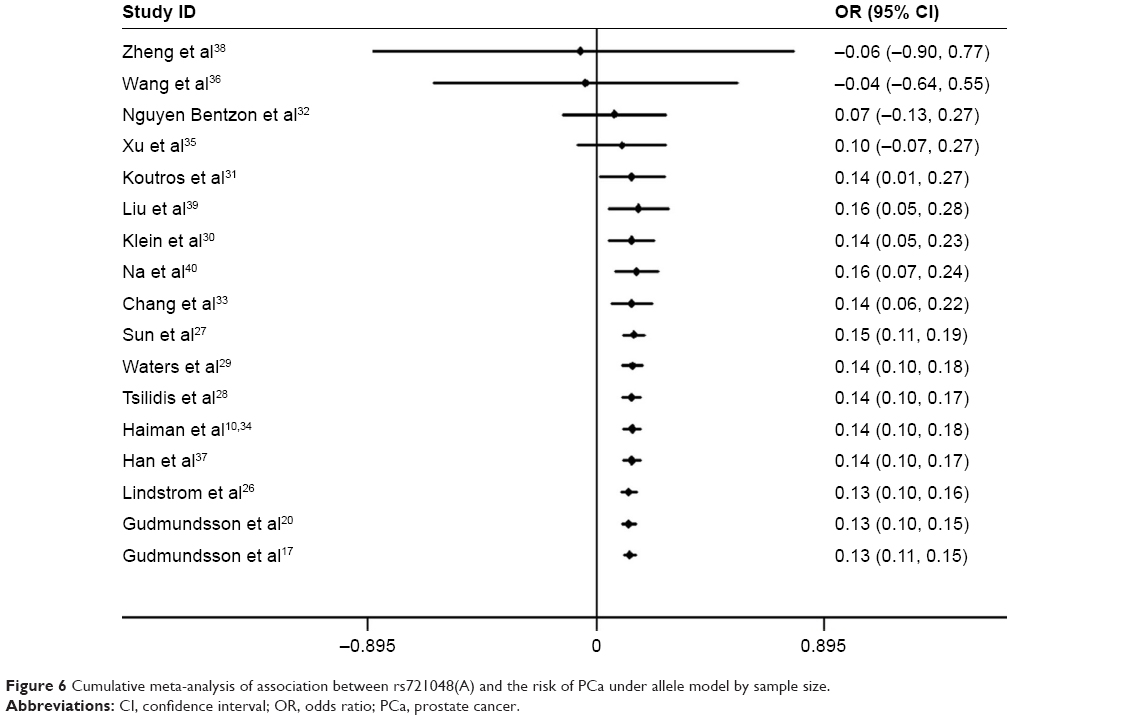 | Figure 6 Cumulative meta-analysis of association between rs721048(A) and the risk of PCa under allele model by sample size. |
Publication bias
In the meta-analysis, publication bias is the most important contributing factor for bias, which can lead to false positive results. The potential publication bias of the 17 studies was determined by Begg’s funnel plot and Egger’s test. The values of individual ORs were within the range of the standard deviation in the funnel plot (Figure 7) and, therefore, no obvious asymmetry and significant publication bias were observed (z=0.21, P=0.837). Similar results were obtained from Egger’s test and no evidence of publication bias was observed (P=0.803, 95% CI=-0.569 to 0.723) (Figure 8).
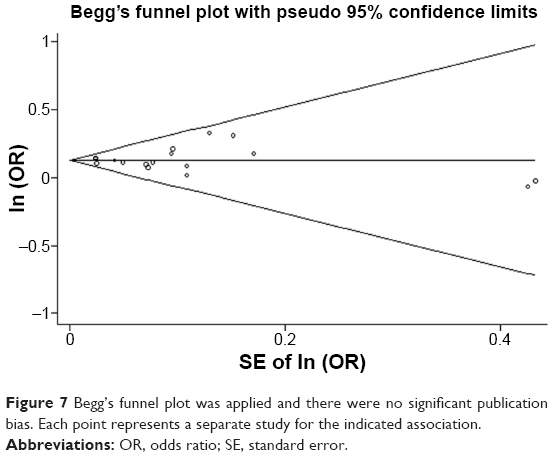 | Figure 7 Begg’s funnel plot was applied and there were no significant publication bias. Each point represents a separate study for the indicated association. |
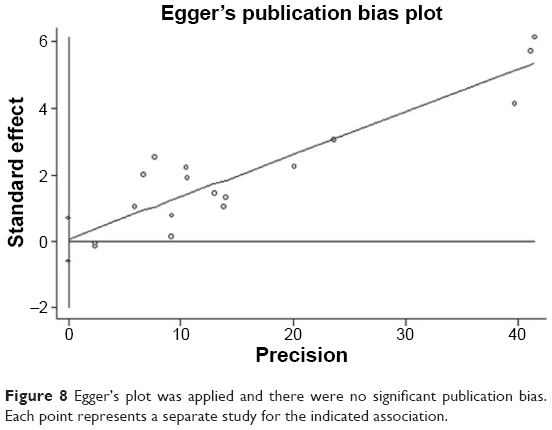 | Figure 8 Egger’s plot was applied and there were no significant publication bias. Each point represents a separate study for the indicated association. |
Discussion
Currently, more than 50 susceptibility loci associated with the risk of PCa have been identified in Europeans, the Japanese, the Chinese, and African-Americans by GWAS, suggesting that genetic factors could be important factors affecting the risk of PCa. In these susceptibility loci, the polymorphism of rs721048A>G in EHBP1 gene has attracted a lot of attention. Extensive research has investigated the association between rs721048(A) and the risk of PCa, but these investigations failed to produce consistent results. The lack of consistency among these results may be due to the small sample sizes of the studies, different ethnicities or publication bias. Therefore, we performed a pooled meta-analysis to determine whether rs721048(A) is a risk factor for PCa, and try to find out the underlying cause of these inconsistent results.
To the best of our knowledge, our study was the first comprehensive meta-analysis to demonstrate the significant association between rs721048(A) and the risk of PCa in the population of the overall study. The meta-analysis included 17 studies with a total of 48,135 cases and 102,543 controls. The results confirmed that rs721048A>G polymorphism was significantly associated with PCa risk. The results from subgroup analysis, which were based on ethnicity, showed that the association was significant in Caucasians, those of African descent, and the Asian population. Sensitivity analysis showed that the results of our meta-analysis were stable and reliable. We also examined the heterogeneity in our study given that heterogeneity is a potential factor that may affect the accuracy of the results. Fortunately, no statistically significant heterogeneity was detected in the meta-analysis. In addition, no publication bias was detected, indicating that the eligible studies were unbiased. The association of rs721048(A) with PCa risk was further confirmed by cumulative meta-analyses. When five studies were included, the analysis results reached statistical significance and became stable with further increases in sample size.
The EHBP1 gene encodes the EHBP1 protein, which is known to associate with PCa risk in humans.17 However, the detailed mechanisms of EHBP1 in the initiation and progression of PCa are still poorly characterized. It has been reported that EHBP1 is essential for the anti-invasive effect of atorvastatin in PTEN-expressing PCa cells. EHBP1 connects the statin-induced P2X7 signaling with aggressive PCa by interacting with P-Rex1 in response to atorvastatin and by regulating the invasiveness of PTEN-positive PCa cells.25 However its role in PTEN-negative PCa cells has not been reported. PTEN is a phosphatase with both protein and lipid phosphatase activities, and it functions as a tumor suppressor. PTEN acts as a negative regulator of the PI3K/AKT signaling pathway by dephosphorylating phosphatidylinositol (3,4,5)-trisphosphate, which is a potent activator of AKT.45 EHBP1 may affect PI3K/AKT signaling pathway through direct interaction with PTEN and subsequent modulation of cellular processes, such as cell metabolism, proliferation, migration, and cell cycle. In addition, EHBP1 is also involved in insulin-stimulated rapid receptor trafficking and the translocation of GLUT4 in 3T3-L1 adipocytes,23 suggesting a possible role for EHBP1 in mediating the translocation of proteins in PCa. The above evidence appeared to support a possible involvement of the EHBP1 gene in P2X7 signaling and invasiveness regulation of PCa cells in humans.
Several features of our study are worth highlighting. First, the sample size of the analysis (a total of 48,135 cases and 102,543 controls) was large enough to minimize error, significantly increasing the statistical power of the analysis. Moreover, no heterogeneity was observed in the pooled meta-analysis and subgroup analysis, suggesting a high level of homogeneity in the population of the overall study. The adjusted OR with 95% CI for each study was used to calculate the pooled effect, which increased the accuracy of the estimate. Furthermore, our meta-analysis suggested that rs721048(A), which was initially identified as a PCa risk in Caucasians, was also associated with PCa risk in the overall population, especially in Caucasians, Asians, and those of African descent. Finally, cumulative meta-analysis, sensitivity analysis, and publication bias results indicated that the results were stable and accurate in the overall study, which comprises of different ethnic populations.
Some potential limitations should be noted in this study. 1) Since our data were extracted from published articles that just included OR with 95% CI under the allele model, and only the allele model was used to assess the association of rs721048(A) with PCa risk. Thus, the distributions of gene in other genetic models are required for further evaluation. 2) We analyzed the association of rs721048(A) with the risk of PCa, not considering SNP–SNP and SNP–environment interactions, or other environmental factors.46 3) The association between SNP rs721048(A) and PCa grade or stage has been confirmed in only four of the 17 included articles.17,26,33,40 Thus, the meta-analysis examining the association of rs721048(A) with PCa grade or stage was not conducted. 4) Our results showed that rs721048(A) was significantly associated with the risk of PCa in the overall population. However, as only one study was included in each subgroup (Latinos, Native Hawaiians, and Japanese descent), the pooled result was not observed in these three subgroups. Thus, further studies of Latinos, Native Hawaiians, and those of Japanese descent and the other ethnicities need to be conducted in the future.
In conclusion, the data from our meta-analysis demonstrated that EHBP1 rs721048(A) polymorphism is a risk factor for PCa in the overall population of the study. Although this SNP was initially identified as a risk factor for PCa in Caucasians, subsequent replication studies yielded inconsistent results. Our meta-analysis strongly demonstrated that rs721048(A) is the risk factor for PCa in Caucasians. A lot of replication studies involving Asian and African descent subjects yielded inconsistent results. The pooled analysis that we performed clearly demonstrated that rs721048(A) is a risk factor for PCa in the overall study population, including Caucasians, Asian, and those of African descent. Our finding provided an important basis for the early screening and clinical treatment of PCa. In addition, the functions and mechanisms of EHBP1 and rs721048A>G polymorphism in PCa also need to be elucidated in the future.
Acknowledgments
We are grateful to Dr Alan K Chang for his valuable contribution in the preparation of this manuscript. This study was supported by grants (31171353, 31271500 to HW) from the National Natural Science Foundation of China and grants (973 Program 2011CB504201 to HW) from the Ministry of Science and Technology of the People’ s Republic of China.
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel RD. American society of clinical oncology quality care symposium 2012: re-engineering your practice to deliver quality and value. J Oncol Pract. 2013;9(3):160–161. | ||
Nelson AW, Tilley WD, Neal DE, Carroll JS. Estrogen receptor beta in prostate cancer: friend or foe? Endocr Relat Cancer. 2014;21(4):T219–T234. | ||
Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127(12):2893–2917. | ||
Hu B, Yang H, Yang H. Diagnostic value of urine prostate cancer antigen 3 test using a cutoff value of 35 mug/L in patients with prostate cancer. Tumour Biol. 2014;35(9):8573–8580. | ||
Patel AR, Klein EA. Risk factors for prostate cancer. Nat Clin Pract Urol. 2009;6(2):87–95. | ||
Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61(2):69–90. | ||
Hsing AW, Chokkalingam AP. Prostate cancer epidemiology. Front Biosci. 2006;11:1388–1413. | ||
Takata R, Akamatsu S, Kubo M, et al. Genome-wide association study identifies five new susceptibility loci for prostate cancer in the Japanese population. Nat Genet. 2010;42(9):751–754. | ||
Akamatsu S, Takata R, Haiman CA, et al. Common variants at 11q12, 10q26 and 3p11.2 are associated with prostate cancer susceptibility in Japanese. Nat Genet. 2012;44(4):426–429,S421. | ||
Haiman CA, Chen GK, Blot WJ, et al. Genome-wide association study of prostate cancer in men of African ancestry identifies a susceptibility locus at 17q21. Nat Genet. 2011;43(6):570–573. | ||
Xu J, Mo Z, Ye D, et al. Genome-wide association study in Chinese men identifies two new prostate cancer risk loci at 9q31.2 and 19q13.4. Nat Genet. 2012;44(11):1231–1235. | ||
Amundadottir LT, Sulem P, Gudmundsson J, et al. A common variant associated with prostate cancer in European and African populations. Nat Genet. 2006;38(6):652–658. | ||
Gudmundsson J, Sulem P, Manolescu A, et al. Genome-wide association study identifies a second prostate cancer susceptibility variant at 8q24. Nat Genet. 2007;39(5):631–637. | ||
Gudmundsson J, Sulem P, Steinthorsdottir V, et al. Two variants on chromosome 17 confer prostate cancer risk, and the one in TCF2 protects against type 2 diabetes. Nat Genet. 2007;39(8):977–983. | ||
Yeager M, Orr N, Hayes RB, et al. Genome-wide association study of prostate cancer identifies a second risk locus at 8q24. Nat Genet. 2007;39(5):645–649. | ||
Eeles RA, Kote-Jarai Z, Giles GG, et al; UK Genetic Prostate Cancer Study Collaborators, British Association of Urological Surgeons’ Section of Oncology, UK ProtecT Study Collaborators. Multiple newly identified loci associated with prostate cancer susceptibility. Nat Genet. 2008;40(3):316–321. | ||
Gudmundsson J, Sulem P, Rafnar T, et al. Common sequence variants on 2p15 and Xp11.22 confer susceptibility to prostate cancer. Nat Genet. 2008;40(3):281–283. | ||
Thomas G, Jacobs KB, Yeager M, et al. Multiple loci identified in a genome-wide association study of prostate cancer. Nat Genet. 2008;40(3):310–315. | ||
Eeles RA, Kote-Jarai Z, Al Olama AA, et al; UK Genetic Prostate Cancer Study Collaborators/British Association of Urological Surgeons’ Section of Oncology, UK ProtecT Study Collaborators, PRACTICAL Consortium. Identification of seven new prostate cancer susceptibility loci through a genome-wide association study. Nat Genet. 2009;41(10):1116–1121. | ||
Gudmundsson J, Sulem P, Gudbjartsson DF, et al. Genome-wide association and replication studies identify four variants associated with prostate cancer susceptibility. Nat Genet. 2009;41(10): 1122–1126. | ||
Yeager M, Chatterjee N, Ciampa J, et al. Identification of a new prostate cancer susceptibility locus on chromosome 8q24. Nat Genet. 2009;41(10):1055–1057. | ||
Kote-Jarai Z, Olama AA, Giles GG, et al; UK Genetic Prostate Cancer Study Collaborators/British Association of Urological Surgeons’ Section of Oncology, UK ProtecT Study Collaborators, The Australian Prostate Cancer BioResource, PRACTICAL Consortium. Seven prostate cancer susceptibility loci identified by a multi-stage genome-wide association study. Nat Genet. 2011;43(8):785–791. | ||
Guilherme A, Soriano NA, Furcinitti PS, Czech MP. Role of EHD1 and EHBP1 in perinuclear sorting and insulin-regulated GLUT4 recycling in 3T3-L1 adipocytes. J Biol Chem. 2004;279(38):40062–40075. | ||
Guilherme A, Soriano NA, Bose S, et al. EHD2 and the novel EH domain binding protein EHBP1 couple endocytosis to the actin cytoskeleton. J Biol Chem. 2004;279(11):10593–10605. | ||
Ghalali A, Wiklund F, Zheng H, Stenius U, Hogberg J. Atorvastatin prevents ATP-driven invasiveness via P2X7 and EHBP1 signaling in PTEN-expressing prostate cancer cells. Carcinogenesis. 2014;35(7):1547–1555. | ||
Lindstrom S, Schumacher F, Siddiq A, et al. Characterizing associations and SNP-environment interactions for GWAS-identified prostate cancer risk markers – results from BPC3. PLoS One. 2011;6(2):e17142. | ||
Sun J, Kader AK, Hsu FC, et al. Inherited genetic markers discovered to date are able to identify a significant number of men at considerably elevated risk for prostate cancer. Prostate. 2011;71(4):421–430. | ||
Tsilidis KK, Travis RC, Appleby PN, et al. Interactions between genome-wide significant genetic variants and circulating concentrations of insulin-like growth factor 1, sex hormones, and binding proteins in relation to prostate cancer risk in the National Cancer Institute Breast and Prostate Cancer Cohort Consortium. Am J Epidemiol. 2012;175(9):926–935. | ||
Waters KM, Le Marchand L, Kolonel LN, et al. Generalizability of associations from prostate cancer genome-wide association studies in multiple populations. Cancer Epidemiol Biomarkers Prev. 2009;18(4):1285–1289. | ||
Klein RJ, Hallden C, Gupta A, et al. Evaluation of multiple risk-associated single nucleotide polymorphisms versus prostate-specific antigen at baseline to predict prostate cancer in unscreened men. Eur Urol. 2012;61(3):471–477. | ||
Koutros S, Berndt SI, Hughes Barry K, et al. Genetic susceptibility loci, pesticide exposure and prostate cancer risk. PLoS One. 2013;8(4):e58195. | ||
Nguyen Bentzon D, Nyegaard M, Børglum A, Ørntoft T, Borre M, Dalsgaard Sørensen K. Replication of prostate cancer risk variants in a Danish case-control association study. Open J Urol. 2012;2(2): 45–54. | ||
Chang BL, Spangler E, Gallagher S, et al. Validation of genome-wide prostate cancer associations in men of African descent. Cancer Epidemiol Biomarkers Prev. 2011;20(1):23–32. | ||
Haiman CA, Chen GK, Blot WJ, et al. Characterizing genetic risk at known prostate cancer susceptibility loci in African Americans. PLoS Genet. 2011;7(5):e1001387. | ||
Xu J, Kibel AS, Hu JJ, et al. Prostate cancer risk associated loci in African Americans. Cancer Epidemiol Biomarkers Prev. 2009;18(7):2145–2149. | ||
Wang Y, Ray AM, Johnson EK, Zuhlke KA, Cooney KA, Lange EM. Evidence for an association between prostate cancer and chromosome 8q24 and 10q11 genetic variants in African American men: the Flint Men’s Health Study. Prostate. 2011;71(3):225–231. | ||
Han Y, Signorello LB, Strom SS, et al; PRACTICAL Consortium, ELLIPSE GAME-ON Consortium. Generalizability of established prostate cancer risk variants in men of African ancestry. Int J Cancer. 2015;136(5):1210–1217. | ||
Zheng SL, Hsing AW, Sun J, et al. Association of 17 prostate cancer susceptibility loci with prostate cancer risk in Chinese men. Prostate. 2010;70(4):425–432. | ||
Liu F, Hsing AW, Wang X, et al. Systematic confirmation study of reported prostate cancer risk-associated single nucleotide polymorphisms in Chinese men. Cancer Sci. 2011;102(10):1916–1920. | ||
Na R, Liu F, Zhang P, et al. Evaluation of reported prostate cancer risk-associated SNPs from genome-wide association studies of various racial populations in Chinese men. Prostate. 2013;73(15):1623–1635. | ||
Zhang Q, Zheng M, Qi XL, Liu F, Mao ZJ, Zhang DH. Effect of NQO1 C609T polymorphism on prostate cancer risk: a meta-analysis. Onco Targets Ther. 2014;7:907–914. | ||
Moher D, Liberati A, Tetzlaff J, Altman DG, Group P. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med. 2009;6(7):e1000097. | ||
Kolonel LN, Henderson BE, Hankin JH, et al. A multiethnic cohort in Hawaii and Los Angeles: baseline characteristics. Am J Epidemiol. 2000;151(4):346–357. | ||
Lou Y, Li R, Xiong L, et al. NAD(P)H: quinone oxidoreductase 1 (NQO1) C609T polymorphism and lung cancer risk: a meta-analysis. Tumour Biol. 2013;34(6):3967–3979. | ||
Yang C, Li S, Wang M, et al. PTEN suppresses the oncogenic function of AIB1 through decreasing its protein stability via mechanism involving Fbw7 alpha. Mol Cancer. 2013;12:21. | ||
Pan H, Niu W, He L, et al. Contributory role of five common polymorphisms of RAGE and APE1 genes in lung cancer among Han Chinese. PLoS One. 2013;8(7):e69018. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.