Back to Journals » Infection and Drug Resistance » Volume 12
Association Between Biofilm Formation, Structure, and the Expression Levels of Genes Related to biofilm formation and Biofilm-Specific Resistance of Acinetobacter baumannii Strains Isolated from Burn Infection in Ahvaz, Iran
Authors Amin M, Navidifar T ![]() , Shooshtari FS
, Shooshtari FS ![]() , Rashno M, Savari M, Jahangirmehr F, Arshadi M
, Rashno M, Savari M, Jahangirmehr F, Arshadi M
Received 8 September 2019
Accepted for publication 15 November 2019
Published 12 December 2019 Volume 2019:12 Pages 3867—3881
DOI https://doi.org/10.2147/IDR.S228981
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Mansour Amin,1,2 Tahereh Navidifar,2,3 Farkhondeh Saleh Shooshtari,2,4 Mohammad Rashno,3,5 Mohammad Savari,1,2 Fatemeh Jahangirmehr,6 Mania Arshadi1
1Infectious and Tropical Diseases Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 3Cellular and Molecular Research Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 4Infertility Research and Treatment Center of Jahad Daneshgahi, Ahvaz, Khuzestan, Iran; 5Department of Immunology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 6Pain Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Correspondence: Tahereh Navidifar
Cellular and Molecular Research Center, Ahvaz Jundishapur University of Medical Sciences, Golestan Street, Ahvaz, Iran
Tel +989307292597
Email [email protected]
Background: The ability of biofilm formation is an effective way for Acinetobacter baumannii survival from stressed conditions. This present study was aimed to evaluate the association between biofilm formation, structure, the expression levels of genes related to biofilm formation and biofilm-specific resistance of A. baumannii strains isolated from burn infections in Ahvaz, Iran.
Methods: In this study, we assessed the antibiotic susceptibilities, ERIC-PCR typing, capacity of biofilm formation and biofilm structure of 64 A. baumannii isolates collected from burn infections. The distribution and the expression levels of genes involved in the biofilm formation including bap, ompA, abaI, pgaA and csuE were assessed by PCR and real-time PCR, respectively.
Results: We classified A. baumannii isolates in 14 clonal types of ERIC-PCR. Most A. baumannii isolates were resistant to all antibiotics tested except to tigecycline and colistin and had the biofilm formation capability but with different capacities. There was a significant inverse relationship between resistance to antibiotic agents and biofilm formation. The biofilm matrix of 50 strains consisted of polysaccharides together with DNA or proteins. The genes involved in the biofilm formation were detected in both biofilm-forming and non-biofilm forming; however, the expression levels of these genes were higher in biofilm producers compared with non-producers.
Conclusion: The biofilm cells exhibited dramatically decreased susceptibility to antibiotic agents; hence, they have great significance for public health. Therefore, the determination of antibiotic susceptibilities in biofilm and planktonic mode, molecular typing, and capacity of biofilm formation in clinical setting is essential.
Keywords: Acinetobacter baumannii, biofilm formation, gene expression, biofilm structure
Introduction
Burn infections are considered as one of the most important complications of thermal injuries. Burn infections may be a life-threatening cause if bacteria have invaded into the tissue layers beneath the dermis. In addition, the invasion of bacteria may lead to bacteremia, sepsis, and multiple-organ dysfunction syndrome.1
In addition to the nature of the burn infections, the quantities of microorganisms colonizing these infections are also important. Gram-negative bacteria due to their extensive antibiotic resistance have accounted as the main pathogens of the burn infections. Acinetobacter baumannii is recognized as one of the leading pathogens in burn units’ worldwide.2
In recent years, the extensive emergence of multi- and pan drug-resistant A. baumannii strains has revealed this organism’s ability to quickly adapt to environmental changes.3
The biofilm formation is one of the hallmark characteristics of opportunistic pathogens such as A. baumannii. Biofilm is a community of bacterial cells attached to biotic or abiotic surfaces where these cells have intimate contacts with each other. A. baumannii can produce a wide variety of virulence factors in the biofilm mode that contributes in the various steps of the attachment of biofilm cells to the biotic or abiotic surfaces.4
The biofilm-associated protein (Bap) is one of the key factors in the initial attachment and maturation of A. baumannii biofilm, so that can affect both the biofilm thickness and bio-volume.5 Also, the major outer membrane protein (OmpA) has an important role in the attachment of bacterial cells on the abiotic surfaces and human alveolar epithelial cells in the biofilm communities.6
The production of pili is essential for the initial steps of biofilm formation. The pili of A. baumannii are encoded by the csuA/BABCDE operon and the inactivation of the csuE gene resulted in the abolition of both pili production and biofilm formation.7 One of the important polysaccharides in the biofilm communities is poly-β-(1-6)-N-acetylglucosamine (PNAG) that its synthesis is controlled by the pgaABCD locus.8 On the other hand, the production of the acyl-homoserine lactone molecules as the product of the autoinducer synthase abaI gene, is essential for quorum sensing and subsequently the later stages of the biofilm formation.9 Overall, the presence and the expression levels of genes involved in the biofilm formation have determined the capacity of the biofilm formation.
One of the main characteristics of bacteria in the biofilm mode is greater tolerance to antibiotics rather than planktonic mode, so that bacteria in the biofilm mode can tolerate 100 to 1000 folds higher concentrations of antimicrobial agents than the planktonic mode.10 Many studies in Iran and other countries reported the capacity of biofilm formation in A. baumannii strains, as well as the frequencies of genes related to it.11–15 However, the associations between the expression levels of genes related to biofilm formation and the capacity of biofilm formation, as well as the structural properties of biofilm matrix are less studied. Hence, this study was aimed to evaluate the association between the biofilm formation, structure, the expression levels of genes related to biofilm formation and biofilm-specific resistance of A. baumannii strains isolated from burn infections in Ahvaz, Iran.
Methods
Study Design and Bacterial Strains
A total of 64 A. baumannii isolates were collected between August and December 2018 from clinical samples of burn infections of hospitalized patients in the intensive care unit (ICU) of Taleghani Hospital in Ahvaz, Iran. The study design was approved by the Research Ethics Committee of Ahvaz Jundishapur University of Medical Sciences, Iran (AJUMS.REC.1396.333). These isolates were transferred to Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences and cultured on MacConkey agar (Merck, Germany). All of these isolates were confirmed as A. baumannii by some biochemical tests, including sugar fermentation, motility, citrate utilization, urease, oxidative/fermentative glucose (O/F) test, catalase, oxidase and growth ability at 37°C and 42°C16 and the amplification of the blaOXA51-like gene,17 according to the primer set listed in Table 1. The A. baumannii strain ATCC19606 was used as the reference strain.
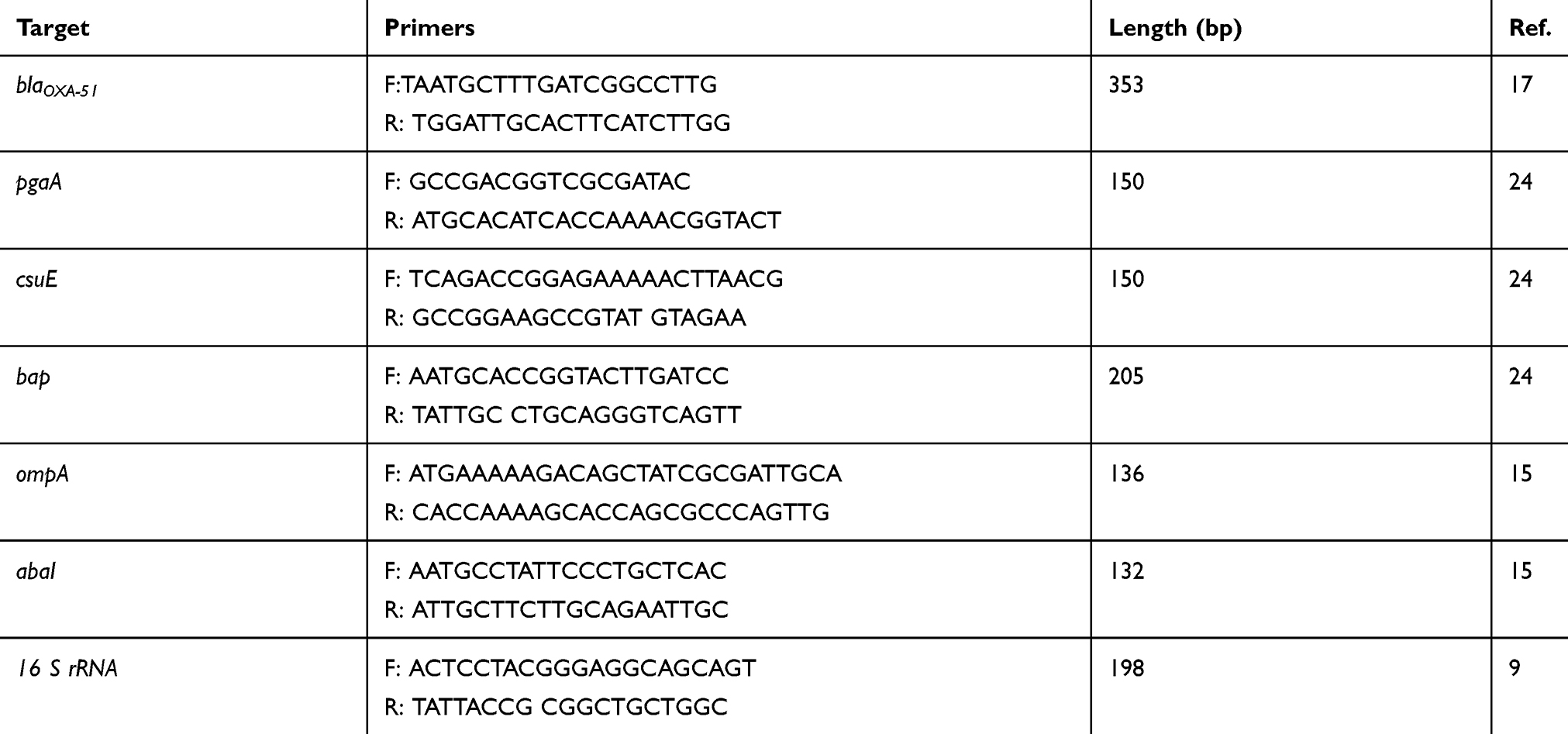 |
Table 1 Primers Used in This Study |
Antibiotic Susceptibility Testing
The minimum inhibitory concentrations (MICs) of a quinolone (levofloxacin), an aminoglycoside (amikacin), a carbapenem (meropenem), a glycylcycline (tigecycline), a fourth-generation cephalosporin (cefepime) and a lipopeptide (colistin) were performed using broth microdilution method and their results were interpreted according to the Clinical and Laboratory Standards Institute guidelines (CLSL, 2018).18 All of these antibiotics were purchased from Sigma Aldrich, USA. Briefly, for levofloxacin, amikacin, meropenem, colistin and cefepime, the MICs of greater than or equal to 8 µg/mL, 64 µg/mL, 8 µg/mL, 4 µg/mL and 32 µg/mL were considered as the resistance breakpoints, respectively. In addition, for tigecycline, a MIC of greater than or equal to 8 µg/mL was proposed as the resistance breakpoint according to the criteria suggested by Jones et al.19
Molecular Typing by ERIC-PCR
The clonal relatedness of the A. baumannii strains was evaluated by enterobacterial repetitive intergenic consensus PCR (ERIC-PCR).20 This reaction was performed using primers ERIC-F (5′-ATGTAAGCTCCTGGGGATTCAC-3′) and ERIC-R (5′-AAGTAAGTGACTGGGGTG AGCG-3′) in a final volume of 20 μL containing 2 U of Taq DNA polymerase, 1.5 mM MgCl2, 200 μM dNTPs, 0.5 μM of each primer (forward and reverse), 10x PCR buffer, 2 μL template DNA (100 pg concentration) and nuclease-free water up to a final volume of 20μL. This amplification condition was as follows: one cycle initial denaturation at 94°C for 5 mins; 35 cycles of 94°C for 1 min, 52°C for 1 min, 72°C for 1.5 mins, and a final extension cycle at 72°C for 15 mins. The comparison of ERIC-PCR banding patterns was performed using Gel Compare II software version 6.6 (Applied Math, Sint-Martens-Latem, Belgium). The relatedness of each ERIC-PCR pattern was determined using Dice coefficient/unweighted pair-group method with arithmetic mean [UPGMA] analysis, with 1% position tolerance. A. baumannii strains with a similarity of 85% were considered as a clonal type.
Biofilm Formation Determination and Quantification
The biofilm formation capability of A. baumannii isolates was assessed using the crystal violet (CV) staining method, as previously described.21 Briefly, an overnight culture of each isolate was prepared in Muller Hinton broth and adjusted to a turbidity of 0.5 McFarland with normal saline (0.85% NaCl). A 10μL aliquot of each suspension was diluted 1:20 in 190 μL of Mueller Hinton broth containing 2.5% glucose in the 96-well polystyrene microtiter plates. After incubation at 37°C for an overnight, the plates were rinsed with PBS three times. The adherent cells were fixed with absolute methanol for 10 mins and stained with 200 μL of 0.1% CV for 20 mins at room temperature. Again, the plates were washed with PBS three times, and then air-dried. Subsequently, the unbound CV was removed by adding 200 μL of a mixture of ethanol:acetone (1:5) for 20 mins and the optical density (OD) at 570 nm was measured using an UV visible spectrophotometer (UV-1601, SHIMADZU). Also, A. baumannii strain ATCC19606 and Muller Hinton Broth were used as positive and negative controls for the biofilm formation, respectively. The results were interpreted according to the criteria suggested by Zhang et al.22
Determination of Biochemical Properties of the Biofilm Matrix
To characterize the biochemical properties of the biofilm matrix, first, the 24 hrs old biofilms of A. baumannii isolates were established in the sterile 96-well polystyrene microtiter plates and washed with PBS for three times. The biofilms were then treated for 1 hr at 37°C either with a solution of 10 mM sodium metaperiodate (NaIO4) in 50 mM sodium acetate buffer for the disruption of the extracellular polysaccharides, with 100 µg/mL of proteinase K for the disruption of the extracellular proteins, or with 100 µg/mL of DNAseI in 150 mM of NaCl and 1 mM CaCl2 for the disruption of the extracellular DNAs. Afterward treatments, the biofilms were washed with PBS, stained with 0.1% CV and the OD at 570 nm measured, as described above. Since the biofilm matrix for each isolate can be a combination structure from polysaccharide, protein and/or DNA, the composition of the biofilm matrix after removing biofilm with specific treatments of sodium metaperiodate (NaIO4), proteinase K and DNAseI was described as + = ≤30%; ++ = 30–70% and +++ = ≥70%.23
Biofilm Antibiotic Susceptibility Testing
The minimum biofilm eradication concentration (MBEC) values of levofloxacin, amikacin, meropenem, tigecycline, colistin and cefepime in A. baumannii isolates were measured using the broth microdilution method. First, the isolates were cultivated in the sterile 96-well polystyrene microtiter plates for an overnight at 37°C to allow for the biofilm formation. The biofilms were then exposed to the concentrations of 2 to 4096 µg/mL of levofloxacin, 4 to 8192 µg/mL of amikacin, 2 to 4096 µg/mL of meropenem, 0.5 to 2048 µg/mL of tigecycline and 16 to 16,384 µg/mL of cefepime for an overnight at 37°C. Then, the wells were washed with sterile PBS three times, and incubated with Muller Hinton Broth (Merck, Germany) for an overnight at 37°C. The MBEC was proposed as any viable cell was not recovered from the biofilm material or i.e. OD570 was less than 0.1. All tests were repeated in triplicate.21
Molecular Identification of the Genes Encoding Biofilm Formation and Efflux Pumps
The genomic DNA of these isolates were extracted using the boiling method, as previously described.17 The uniplex PCR assays were performed for the molecular identifications of the bap, pgaA, abaI, ompA and csuE genes and efflux pumps, including adeB, adeJ and adeG in a final volume of 25 μL.15,24 The sequences and sizes of the primers used in this study are shown in Table 1.
The amplification mixture consisted of 1U Taq DNA polymerase (Cinaclone, Iran), 1.5 mM MgCl2, 200 μM dNTPs, 0.35 μM of each primer, 10x PCR buffer, 5 μL of template DNA and distilled water up to a final volume of 25 μL. The amplification process was performed in a Mastercycler Nexus Thermal Cycler Gradient (Eppendorf, Hamburg, Germany) with one cycle of initial denaturation at 94°C for 3 mins, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 56°C for the bap, ompA and csuE genes, 59°C for the abaI and ompA genes and 55°C for the adeB, adeJ and adeG genes for 30 s, extension at 72°C for 30 s, and a final extension cycle at 72°C for 10 mins. The PCR products were visualized on 2% agarose gel stained with safe stain.
Quantitative Real-Time PCR Assay
First, the RNA extraction and genomic DNA removal of all A. baumannii strains were performed using an RNeasy Mini kit (Qiagen, Tokyo, Japan). The quality and integrity of the total RNA were evaluated with the NanoDrop spectrophotometer (Thermo Fisher Scientific, MA, USA) and agarose gel electrophoresis. The final concentration of each one of the RNA extracts was adjusted to 400 ng/μL. The RNA was reverse transcribed to cDNA using PrimeScript™ 1st strand cDNA Synthesis Kit (Qiagen Tokyo, Japan) according to the manufacturer’s procedure and all of the cDNAs were stored at −20°C. Real-time PCR amplification reaction was prepared in a final volume 20 µL, with 400 ng cDNA, 10 µL RealQ Plus Master Mix Green (Amliqon, Denmark), 0.5 µL each of forward and reverse primers (10 nM each) and RNase- and DNase-free water up to the final volume 20 µL. The Primer sequences used for the genes related to the biofilm formation, including bap, abaI ompA, csuE and pgaA gene and efflux pumps, including adeB, adeJ and adeG are shown in Table 1.24 The 16rRNA gene was used as an internal control for the normalization of gene expression levels. Real-time PCR was performed using a Step One Real-Time PCR System (Applied Biosystems, CA, USA) as follows: on cycle of initial denaturation at 95°C for 15 mins, 40 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 30 s.
The relative expression levels of each gene in the clinical strains are shown as the fold changes compared with the transcript levels of those in A. baumannii strain ATCC 19606 and were calculated using the 2−ΔΔCt method.
Statistical Analysis
The descriptive statistical tests were used for the evaluation of OD values of the biofilm formation, the frequencies of genes encoding biofilm and susceptibility antibiotic pattern. Spearman’s rank (rS) correlation was used for the comparison of the biofilm formation capacity with susceptibility and resistance to antibiotic agents, and Pearson correlation coefficient (PCC) was used for the determination of correlation between MIC and MBEC values. The mRNA expression analysis were performed using Student’s t test and one-way analysis of variance (ANOVA), followed by the Tukey's multiple comparison test. In all analyses, a two-sided significance level of <0.01 was considered statistically significant.
Results
Bacterial Strains, Antibiotic Susceptibility and Molecular Typing
A total of 64 non-duplicate isolates were confirmed as A. baumannii using the amplification of the blaOXA-51 gene. These samples were collected from the different clinical samples, including bronchial lavage 17 (26.56%), wound 15 (23.43%), tracheal secretion 11 (17.18%), urine 11 (17.18%), and blood 10 (15.62%). The antibiotic susceptibility of these isolates in planktonic mode was determined using the broth microdilution method and according to these results, 56.25% of the A. baumannii isolates were MDR. Moreover, the majority of the isolates were resistant to amikacin (79.7%; 51 isolates), followed by meropenem (75%; 48 isolates), levofloxacin (60.9%; 39 isolates), cefepime (53.1%; 24 strains) and tigecycline (25%; 16 isolates), while all isolates were sensitive to colistin. Also, according to the analysis of ERIC-PCR band patterns, these 64 A. baumannii isolates were clustered into 14 clonal types and 11 single type of ERIC-PCR. Figure 1 describes the dendrogram obtained from ERIC-PCR analysis of these isolates. The correlation between the MICs of each one of antibiotics and ERIC-PCR types is shown in Table 2. According to these results, all strains belonging to a same clone type had similar antibiotic resistance pattern, i.e. there was a significant association (p< 0.05) between the clone types and antibiotic resistance.
 |
Table 2 MICs of Tested Antibiotics and ERIC-PCR Types for Acinetobacter baumannii Strains |
 |
Figure 1 Dendrogram of ERIC-PCR analysis of 64 Acinetobacter baumannii strains. |
Biofilm Formation Capacity and Its Structure Characterizes
The biofilm-forming capacity of these 64 A. baumannii isolates was evaluated using CV staining method. The OD570 values for A. baumannii ACC19606 (as positive control) and Muller Hinton broth (as negative control) were 0.452 ± 0.052 and 0.085 ± 0.002, respectively. The OD570 values for the clinical strains ranged from 0.125± 0.056 to 1.745 ± 0.054. Overall, 55 isolates were biofilm producers among which, 15 (23.4%) isolates were strong producers, 13 (20.3%) were moderate producers and 27 (42.2%) were weak producers. Also, 13 (20.3%) isolates showed the biofilm formation more robust than A. baumannii ATCC19606. The correlation between biofilm-forming capacity and ERIC-PCR types is shown in Table 3. According to these results, all strains belonging to a same clone type had similar biofilm-forming capacity.
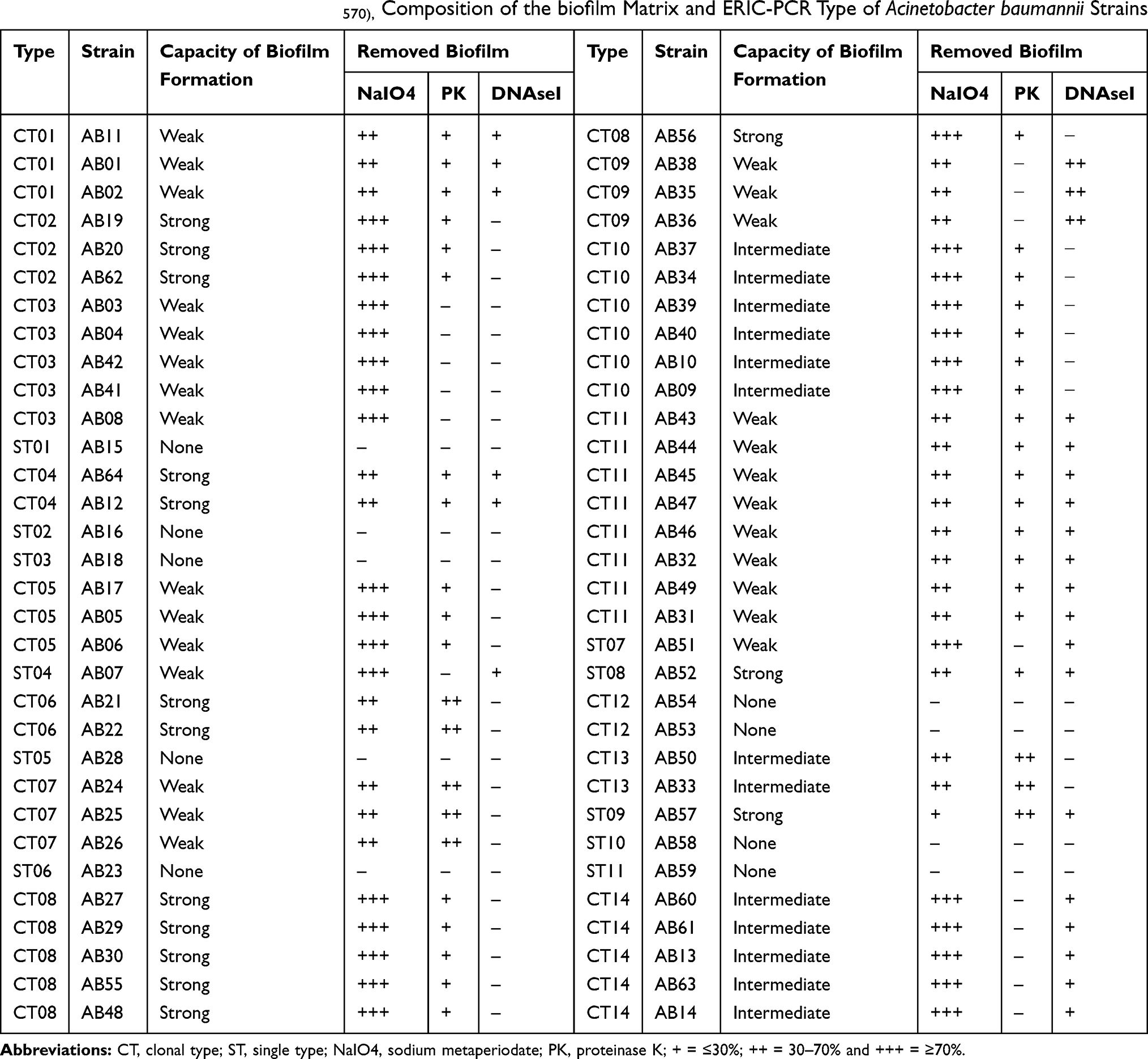 |
Table 3 Biofilm-Forming Capacity (OD570), Composition of the biofilm Matrix and ERIC-PCR Type of Acinetobacter baumannii Strains |
On the other hand, the structure characterizes of the biofilm matrix were recognized by several treatments. According to these treatments, the biofilm matrix in 5 (7.8%) A. baumannii isolates was exclusively composited from polysaccharides, in 15 (23.4%) isolates was composited from the combination of proteins, polysaccharides and DNA, in 25 (39.06%) isolates were composited from the combination of proteins and polysaccharides and in 10 (15.62%) isolates was composited from the combination of DNA and polysaccharides. The composition of the biofilm matrix for each strain is shown in Table 3. According to these results, all strains belonging to a same clone type had similar structure characterizes in biofilm matrix.
Relationships Between Biofilm Formation and Antibiotic Resistance
Our results revealed that all strong biofilm-formers were non-MDR strains, while all weak biofilm-formers were MDR strains. Also, nine strains were non biofilm-formers, which consisted of one non-MDR strain and eight MDR strains. Hence, it is suggested that non-MDR A. baumannii strains tended to form stronger biofilms than MDR strains, as was confirmed by statistical analyses (rs = −0.715, P < 0.001), i.e. non-MDR strains had greater possibility to produce strong biofilm than MDR strains.
On the other hand, we evaluated the association between the biofilm-forming capacities and antibiotic resistance. The results revealed that apart from colistin, for each antibiotic, susceptible isolates could form more robust biofilms than moderate and resistant strains, indicating a significant negative correlation between the capacity of biofilm formation and resistance phenotypes to all antibiotics except to colistin (rs = −0.258 to −0.708, P < 0.001; Table 4). For colistin, no significant correlation was observed (rs = −0.030, P = 0.813; Table 4).
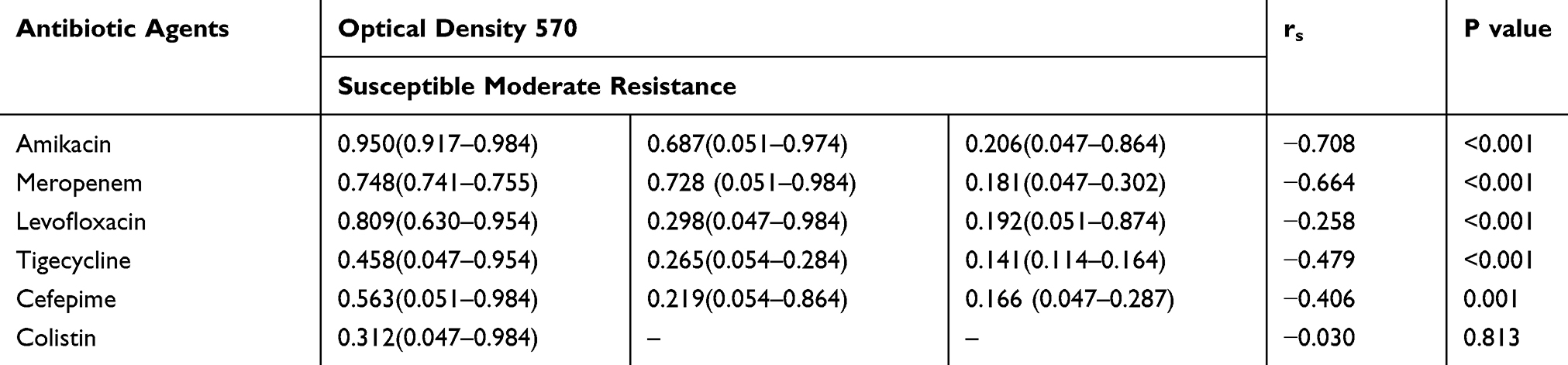 |
Table 4 Association Between the Biofilm-Forming Capacities and Antibiotic Resistance |
Relationship Between MBECs and MICs
The MIC values of meropenem, levofloxacin, cefepime, tigecycline, amikacin and colistin of these isolates ranged from 1 to 1024 μg/mL, 1 to 512 μg/mL, 0.5 to 512 μg/mL, 0.25 to 32 μg/mL, 8 to 512 μg/mL and 0.25 to 1 μg/mL, respectively. As expected, the MBECs of these antibiotics were higher than their respective MICs, followed by 32 to 4096 μg/mL for amikacin, 8 to 4096 μg/mL for levofloxacin and cefepime, 16 to 4096 μg/mL for meropenem, 8 to 512 μg/mL for tigecycline and 4 to 16 μg/mL for colistin.
With an analysis of MBEC and MIC values of these antibiotics, we found an increase of 2- to 16-fold higher MBEC values rather than MIC values for amikacin, 4- to 32-fold for cefepime, colistin and meropenem, 8- to 32-fold for levofloxacin and 8- to 64-fold for tigecycline. By Pearson correlation coefficients (PCC) analysis of the MIC and MBEC values of each antibiotic, a significant positive relationship (P < 0.001) was demonstrated between the MIC and MBEC values for all of these antibiotics (PCC = 0.686 for tigecycline, PCC = 0.853 for cefepime, PCC = 0.885 for meropenem, PCC = 0.928 for levofloxacin and PCC = 0.799 for amikacin P < 0.001) except to colistin (PCC = 0.138; P = 0.280)
Distribution of the Genes Involved in Biofilm Formation and Efflux Pumps by PCR
The amplification of the genes encoding biofilm formation was performed by PCR. According to these results, all isolates (biofilm producers or no producers) harbored the ompA and abaI genes. Also, the majority of the isolates were positive for the amplification of pgaA gene (85.93%; 55 strains), followed by csuE gene (60.93%; 39 strains) and bap (48.43%; 31 strains). The presence of csuE gene was confirmed among all strong biofilm producers, 11 out of 13 intermediate producers, 3 out of 27 weak producers as well as 5 out of 9 no producers, while the bap and pgaA genes were detected only among biofilm producers. Moreover, the pgaA gene were recognized only among biofilm producers (55 strains), while the presence of bap gene was confirmed among all strong biofilm producers, 10 out of 13 intermediate producers and 6 out of 27 weak producers. Table 5 shows the association between the biofilm formation capacity, the clonal types and gene profiles. According to these results, all biofilm strong producers had the gene profile of bap, csuE, ompA, pgaA,and abaI. Also, almost all of the strains belonging to a clonal type had similar gene profiles; however, the strains belonging to each one of the clonal types of CT10 and CT14 had several gene profiles, as described in Table 5.
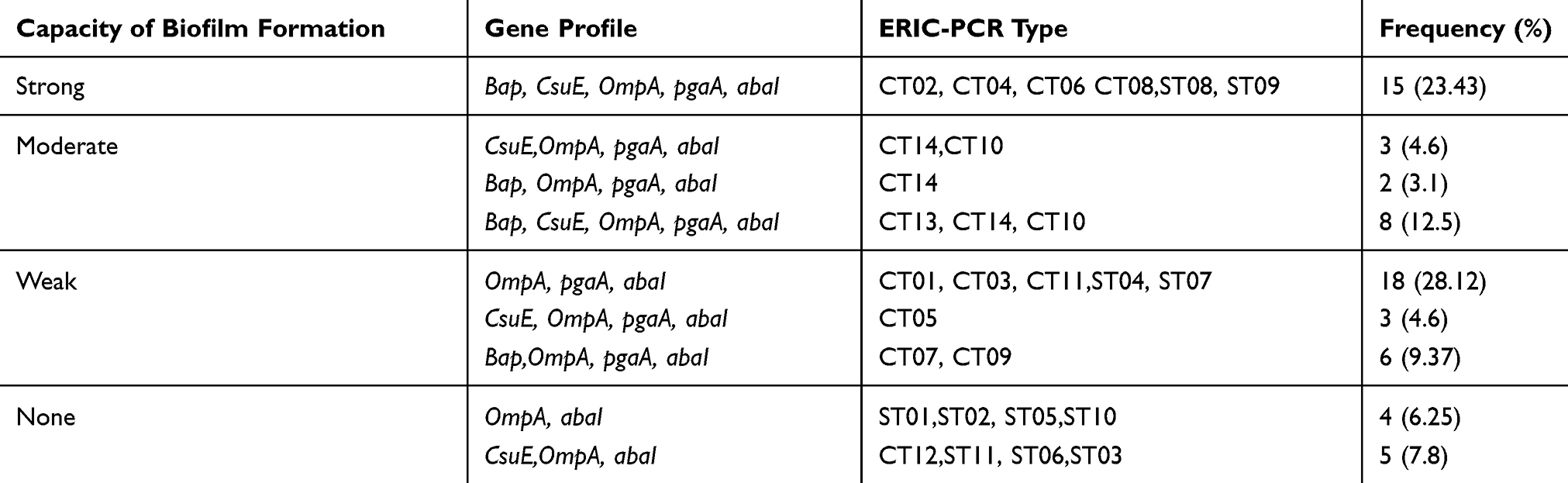 |
Table 5 Association Between the Biofilm Formation Capacity, the Clonal Types and Gene Profiles |
On the other hand, the adeB, adeJ and adeG genes were detected among 92.18% (n=59), 98.43% (n=63) and 89.06% (n=57) of strains, respectively.
Expression Levels of the Genes Involved in Biofilm Formation and Efflux Pumps by Real-Time PCR
To determine the expression levels of the genes involved in biofilm formation and efflux pumps, we analyzed the rate of expression of each one of the genes involved in biofilm formation in 64 A. baumannii strains as compared to those in A. baumannii ATCC19606 strain by real-time PCR. Table 6 shows the expression means of the genes involved in biofilm formation with regard to the biofilm formation capacity. According to these results, the expression means of all of these genes were significantly higher among strong biofilm producers than moderate or weak producers (p<0.01).
 |
Table 6 Association Between the Expression Levels of Genes Involved in Biofilm Formation and the biofilm formation Capacity |
As mentioned above, the ompA and abaI genes were detected among all isolates; however, the expression level means of these two genes were less among biofilm no-producers than biofilm producers (Table 6). Overall, there was a significant difference between the expression levels of all of the genes involved in biofilm formation and the biofilm formation capacity (p<0.01).
The expression levels of mRNA of the adeB, adeJ and adeG genes with regard to non-susceptible to antibiotics are described in Table 7. According to the results, the increased levels of adeB gene compared to that of A. baumannii ATCC 19606 were observed among 32 (50%) isolates ranged from 1.62- to 85.63-fold. The increased level of adeB was obvious among 23 (43.4%), 20 (44.4%), 25 (46.3%), 23 (60.5%) and 25 (50%) isolates of non-susceptible to amikacin, cefepime, levofloxacin, tigecycline and meropenem, respectively, while the level of adeB enhanced among 9 (81.8%), 12 (63.2%), 7 (70%), 9 (34.6%) and 7 (50%) isolates of susceptible to amikacin, cefepime, levofloxacin, tigecycline and meropenem, respectively.
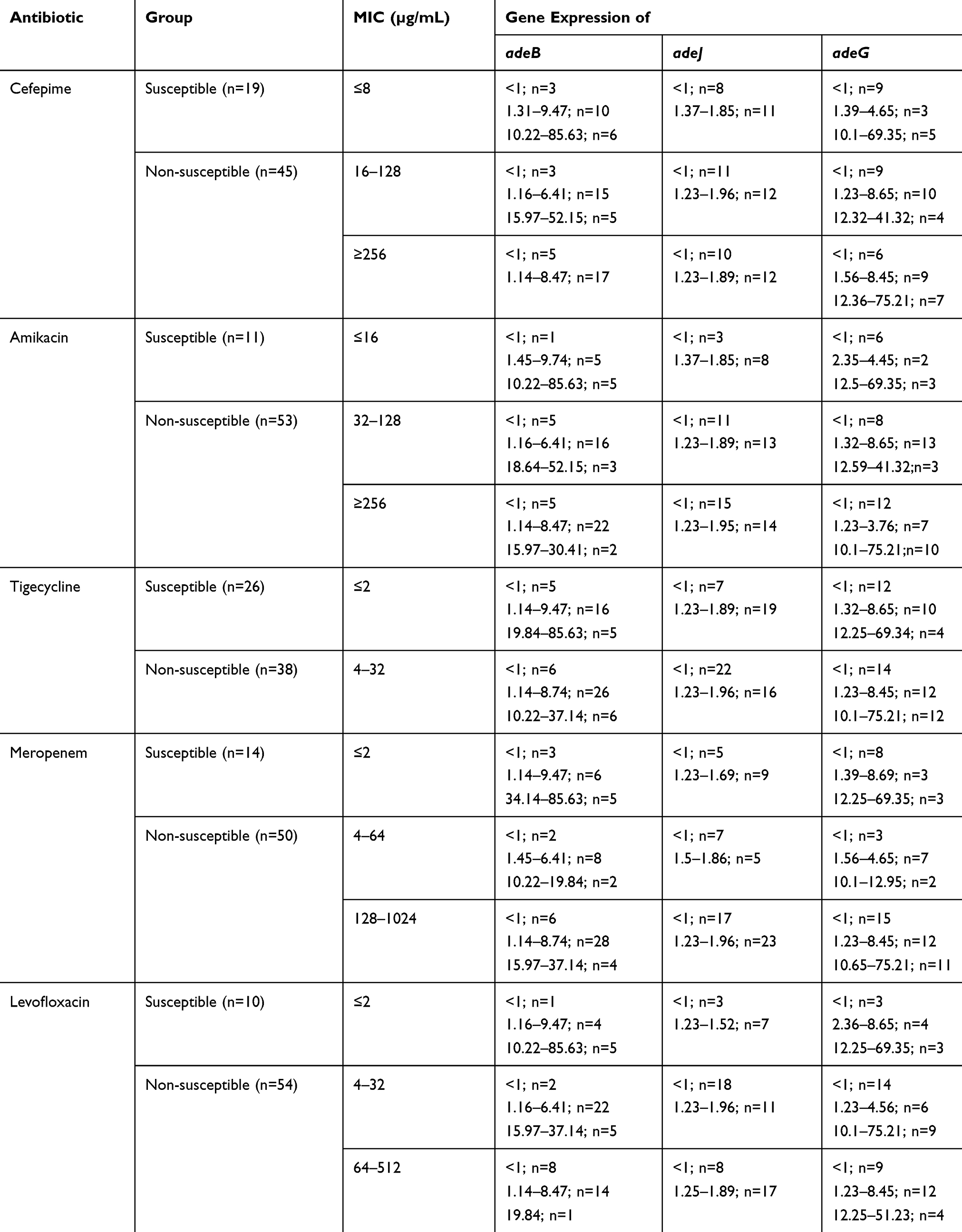 |
Table 7 Expression Levels of mRNA of the adeB, adeJ and adeG Genes with Regard to Non-Susceptible to Antibiotics |
The increased levels of adeJ gene compared to that of A. baumannii ATCC 19606 were observed among 16 (25%) isolates ranged from 1.5- to 1.96-fold. The increased level of adeB was obvious among 15 (28.3%), 14 (10.5%), 16 (26.6%), 7 (18.4%) and 15 (30%) isolates of non-susceptible to amikacin, cefepime, levofloxacin, tigecycline and meropenem, respectively, while the level of adeB enhanced among 1 (9.1%), 2 (31.1%), 0.9 (34.6%) and 1 (7.1%) isolates of susceptible to amikacin, cefepime, levofloxacin, tigecycline and meropenem, respectively.
The increased levels of adeG gene compared to that of A. baumannii ATCC 19606 were observed among 32 (50%) isolates ranged from 1.56- to 75.21-fold. The increased level of adeB was obvious among 27 (50.9%), 25 (55.6%), 25 (46.3%), 20 (52.8%) and 27 (54%) isolates of non-susceptible to amikacin, cefepime, levofloxacin, tigecycline and meropenem, respectively, while the level of adeB enhanced among 5 (45.5%), 7 (36.8%), 7 (70%), 12 (46.2%) and 5 (35.7%) isolates of susceptible to amikacin, cefepime, levofloxacin, tigecycline and meropenem, respectively.
Statistical analysis revealed the significant association between the increased level of adeB gene and non-susceptibility to amikacin (p=0.043). However, no statistically significant association was found between overexpression of adeB, adeJ or adeG gene and non-susceptibility to other antibiotics.
The expression levels of mRNA of the adeB, adeJ and adeG genes with regard to the capacity of biofilm formation are described in Table 8. According to the results, the increased levels of adeB gene compared to that of A. baumannii ATCC 19606 were indicated among 4 (44.4%), 15 (55.8%), 3 (23.1%), and 10 (66.7%) isolates of non-biofilm producers, weak, intermediate, and strong producers, respectively. The increased levels of adeG gene compared to that of A. baumannii ATCC 19606 were indicated among 4 (44.4%), 15 (55.8%), 6 (46.2%), and 7 (46.7%) isolates of non-biofilm producers, weak, intermediated, and strong producers, respectively. The increased levels of adeJ gene compared to that of A. baumannii ATCC 19606 were indicated among 5 (55.6%), 5 (18.5%), 6 (46.2%), and 7 (46.7%) isolates of non-biofilm producers, weak, and intermediate producers, respectively, while the level of adeB enhanced among 22 (81.5%) weak producers and all strong producers.
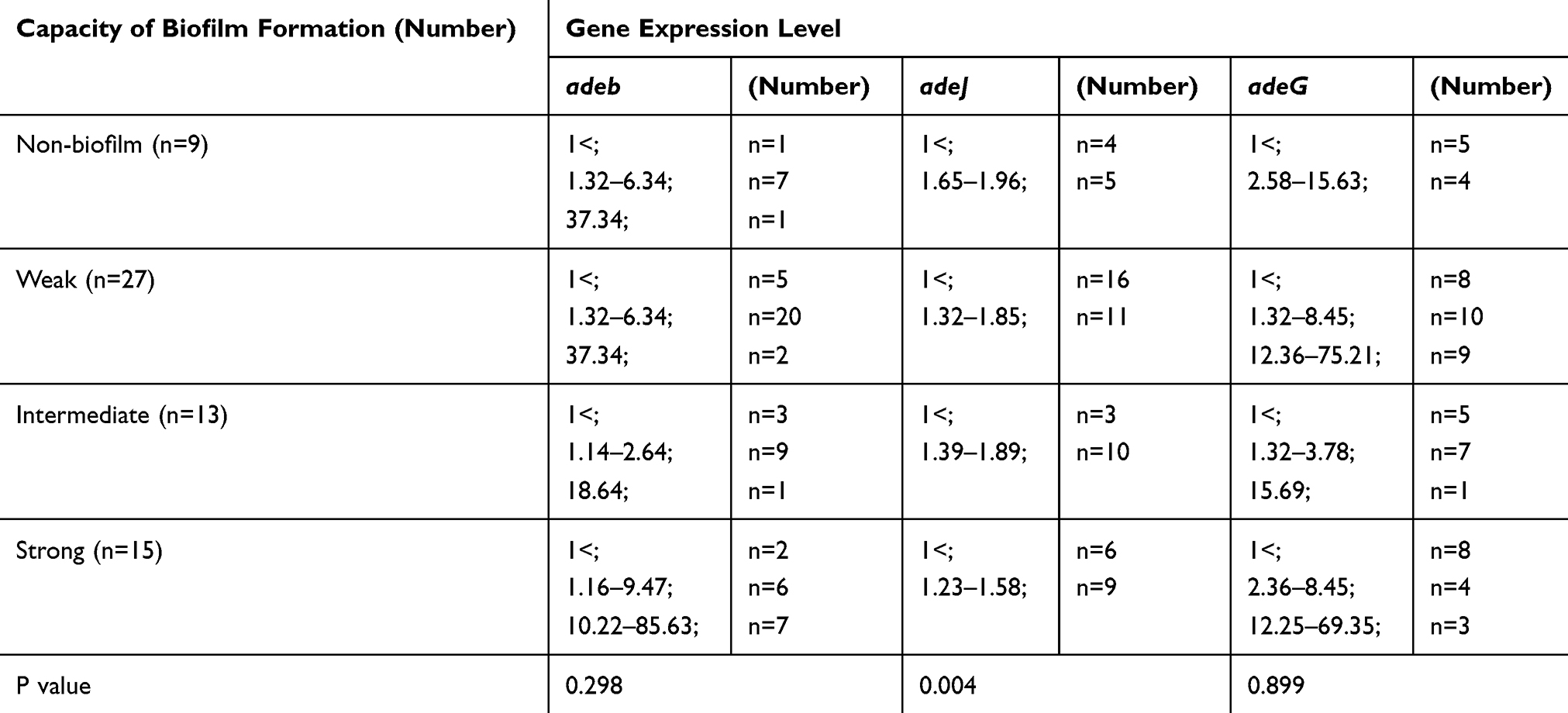 |
Table 8 Expression Levels of mRNA of the adeB, adeJ and adeG Genes with Regard to Capacity of Biofilm Formation |
Statistical analysis showed the significant association between the increased level of adeJ gene and the capacity of biofilm formation (p=0.004). However, no statistically significant association was found between overexpression of adeB or adeG gene and the capacity of biofilm formation.
Discussion
A. baumannii as one of the main causes of burn infections is responsible for an extensive range of serious infections.25 The up-regulation of innate resistance mechanisms such as overexpression of efflux pumps and the acquisition of foreign genetic determinants such as plasmids is critical characteristics for the survival of A. baumannii during environmental pressures such as hospital environments.26 The growing global emergence of A. baumannii strains resistant to all β-lactam agents, highlights the potential of this microorganism to adapt rapidly to selective environmental stresses. MDR A. baumannii is recognized to be among the most difficult antibiotic-resistant gram-negative bacilli to control and treat.27
The results of our study showed that 56.25% of A. baumannii isolates were MDR. The high prevalence of MDR A. baumannii isolates was also reported from some studies in Iran and other countries.28–33 The multidrug antibiotic resistance can decrease the efficacy of the common antibiotics used in the clinical setting especially in the infections caused by carbapenem-resistant strains. During the recent two decades, the carbapenems have recommended as the first-line antibiotics for the treatment of A. baumannii infections. However, unfortunately, increasing resistance to carbapenems has been reported worldwide among A. baumannii strains.26 In our study, the antibiotic susceptibility test results showed that the majority of these isolates were resistant to meropenem (75%). In agreement with ours, the high rates of carbapenem-resistant A. baumannii isolates were reported from other studies.28–33
Also, our results showed that the antibiotic resistance rates to amikacin, cefepime and levofloxacin among MDR A. baumannii strains were more than 50%. In similar to our work, Mirnejad et al,34 Huang et al35 and Taherikalani et al36 also, reported the high prevalence of the resistance to these antibiotic agents among A. baumannii isolates.
Tigecycline and colistin are the only treatment options for infections caused by extensively drug resistant (XDR) or MDR A. baumannii.27 However, these two antibiotics have some undesirable side effects, such as nephrotoxicity and neurotoxicity. Nevertheless, carbapenems together with tigecycline or colistin are recommended as the best therapeutic approaches for the treatment of MDR A. baumannii infections. Moreover, these combination therapies are efficacious and have lower toxicity than monotherapy with tigecycline or colistin.37
The antibiotic susceptibility test results showed that the majority of A. baumannii isolates were susceptible to tigecycline and all isolates were sensitive to colistin which are in agreement with other reports obtained from previous studies in Iran and other countries.38–41 Hence, these results suggest that tigecycline and colistin are still the most effective antibiotic agents against MDR A. baumannii strains.
The biofilm matrix can considerably protect bacteria from both the immune system cells and antibiotic agents.42 In our study, most A. baumannii strains had the ability of biofilm production but with different capacities. As mentioned above, we found a significant inverse relationship between the capacity of biofilm formation and resistance to all antibiotic agents except to colistin (p<0.001), i.e. the biofilm density in sensitive strains was more than biofilm density in resistant strains. In consistent with our study, some researchers also21,43,44 demonstrated that the sensitive strains tended to produce stronger biofilms than the resistant strains whereas some others45,46 showed that MDR strains had more capability for the biofilm production than sensitive strains. It seems that the biofilm formation acts as a mechanism for bacteria to get a better survival when they are exposed to antibiotic agents, especially in the strains which their antibiotic resistance levels are not high enough.21 The molecular mechanisms govern on this process are not clear yet; however, Gallant et al47 showed that the expression of the β-lactamase gene blaTEM−1 in a clinical strain of P. aeruginosa inhibited the biofilm formation by distributing cell wall; indicating a genetic association between the biofilm formation and antibiotic resistance. Nevertheless, the exploration of the genetic links between biofilm formation and antibiotic resistance mechanisms than blaTEM−1 is required to fully elucidate the possible causes involved in this process.
The biofilm cells exhibit dramatically decreased susceptibility to antibiotic agents; hence, they have great significance for public health. In this present study, the MIC and MBEC values of antibiotic agents were determined using broth microdilution method. As expected, we found a significant increase in MBECs as compared to MICs for all of the antibiotic agents. These findings are also consistent with the findings of Li et al21 indicating biofilm cells exhibit enhanced antibiotic resistance as compared to planktonic cells. The enhancement of biofilm-specific resistance can be explained by several factors, including the exopolysaccharide matrix of biofilm, different growth rates and nutrient gradients within the biofilm, mutational resistance, the up-regulation of efflux pumps, persister phenomenon and intrinsic characteristics of bacteria cells in biofilm mode.10,48 Moreover, the persister cells as the dormant variants within bacterial biofilms have more tolerant to most antibiotics than planktonic bacteria that may cause a relapse of infection.48
Bacteria molecular typing is nowadays an integral part of the public health microbiology researches that is able to differentiate epidemiological relates from unrelated isolates belonging to a same bacterial species. Molecular typing is used to elucidate the source and route of transmission of microorganisms causing outbreaks of infectious diseases, the bacterial population structures and microbial genetic diversity in different environments. A variety of typing methods have been developed to classify and compare the genetic relatedness of bacterial isolates in epidemiological investigations49 among which, ERIC-PCR is a plausible, easy and fast strategy that does not require the specialized equipment and reagents.50 Our results showed that strains belonging to a clonal type had similar properties such as the composition of biofilm matrix, antibiotic resistance and gene profile, indicating the importance of molecular typing methods such as ERIC-PCR for the identification of clonal types of A.baumannii in infectious diseases.
In A. baumannii, the genes encoding ompA, bap, pgaA, csuE and abaI are known to be the key factors in the biofilm formation. Our results revealed that the gene profile of bap, csuE, ompA, pgaA, abaI was presented in all biofilm strong producers and some moderate producers. Also, the gene pattern of csuE, ompA, pgaA, abaI was found in some biofilm weak producers and some moderate producers. These findings suggest that in addition to the presence of these genes, their expression levels also play the important role in the determination of the capacity of biofilm formation. On the other hand, some genes involved in the biofilm formation were detected in non-biofilm forming strains; however, the expression levels of these genes were higher in biofilm producers as compared with non-producers, which is in agreement with Wang et al14 findings. Moreover, they indicated the higher expression levels of the Bap1, AbaI and Csu A/B genes in the biofilm-forming strains compared to the matched non-biofilm-forming strains.
The importance of efflux-mediated resistance in the development of resistance to several classes of antibiotics is indicated in several reports.51–54 Our results indicated that the role of RND-type efflux pump of AdeABC in conferring to amikacin, as showed in the studies conducted by Magnet et al55 and Marchand56 et al who indicated the adeB mutants had more susceptibility (8 to 32-fold) to aminoglycoside agents than their parent strains; however, the expression level of AdeABC efflux pump was not measured in their studies. In this present study, no statistically significant association was found between overexpression of adeB, adeJ or adeG gene and non-susceptibility to other antibiotics. However, in contrast with our findings, some researchers revealed the high expression of the AdeABC efflux pump is closely associated with meropenem and tigecycline resistance,57–59 suggesting still more researches are needful for elucidating the association between the AdeABC efflux pump and resistance to meropenem and tigecycline.
On the other hand, the efflux pumps especially the resistance-nodulation-cell division (RND) family confer low-level multidrug resistance and have several roles during the process of biofilm formation in A. baumannii. Moreover, these pumps have extruded actively the autoinducers associated with quorum sensing, and harmful molecules such as antibiotics and metabolic intermediates.24 In this present study, there was a significant association between the overexpression of adeJ and the capacity of biofilm formation, so that all strong biofilm producers and 81.5% of intermediate producers showed the overexpression of the adeJ. However, similar to our study, no research has not showed the association between the expression of efflux pumps and the capacity of biofilm formation.
Conclusion
In this study, the frequencies of genes involved in biofilm formation were high. There was a significant inverse relationship between resistance to antibiotic agents and biofilm formation. Also, a significant increase in MBECs was showed as compared to their respective MICs. The genes involved in the biofilm formation were detected in both biofilm-forming and non-biofilm-forming strains; however, their expression levels were higher in biofilm producers as compared with non-producers.
Acknowledgments
This study was a part of Ph.D. thesis of Tahereh Navidifar which has been approved as a research (Grant No. CMRC-9614) and was financially supported by Cellular and Molecular Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran and thank all of them. The Health Research Institute, Infectious and Tropical Diseases Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran for financial support (Grant No. CMRC-9614).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Rowan MP, Cancio LC, Elster EA, et al. Burn wound healing and treatment: review and advancements. Crit Care. 2015;19:243. doi:10.1186/s13054-015-0961-2
2. Church D, Elsayed S, Reid O, Winston B, Lindsay R. Burn wound infections. Clin Microbiol Rev. 2006;19(2):403–434. doi:10.1128/CMR.19.2.403-434.2006
3. Dedeić-Ljubović A, Granov Đ, Hukić M. Emergence of extensive drug-resistant (XDR) Acinetobacter baumanniiin the Clinical Center University of Sarajevo, Bosnia and Herzegovina. Med Glas (Zenica). 2015;12(2):169–176. doi:10.17392/809-15
4. Gaddy JA, Actis LA. Regulation of Acinetobacter baumannii biofilm formation. Future Microbiol. 2009;4(3):273–278. doi:10.2217/fmb.09.5
5. Brossard KA, Campagnari AA. The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect Immun. 2012;80(1):228–233. doi:10.1128/IAI.05913-11
6. Gaddy JA, Tomaras AP, Actis LA. The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect Immun. 2009;77(8):3150–3160. doi:10.1128/IAI.00096-09
7. Tomaras AP, Dorsey CW, Edelmann RE, Actis LA. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: involvement of a novel chaperone-usher pili assembly system. Microbiology. 2003;149(pt2):3473–3484. doi:10.1099/mic.0.26541-0
8. Choi AH, Slamti L, Avci FY, Pier GB, Maira-Litrán T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J Bacteriol. 2009;191(19):5953–5963. doi:10.1128/JB.00647-09
9. Bhargava N, Sharma P, Capalash N. Quorum sensing in Acinetobacter: an emerging pathogen. Crit Rev Microbiol. 2010;36(4):349–360. doi:10.3109/1040841X.2010.512269
10. Stewart PS. Antimicrobial tolerance in biofilms. Microbiol Spectr. 2015;3. doi:10.1128/microbiolspec.MB-0010-2014
11. Fallah A, Rezaee MA, Hasani A, Barhaghi MHS, Kafil HS. Frequency of bap and cpaA virulence genes in drug resistant clinical isolates of Acinetobacter baumannii and their role in biofilm formation. Iran J Basic Med Sci. 2017;20(8):849–855.
12. Monfared AM, Rezaei A, Poursina F, Faghri J. Detection of genes involved in biofilm formation in MDR and XDR Acinetobacter baumannii isolated from human clinical specimens in Isfahan, Iran. Arch Clin Infect Dis. 2019;14(2):e85766.
13. Badmasti F, Siadat SD, Bouzari S, Ajdary S, Shahcheraghi F. Molecular detection of genes related to biofilm formation in multidrug-resistant Acinetobacter baumannii isolated from clinical settings. J Med Microbiol. 2015;64(Pt 5):559–564. doi:10.1099/jmm.0.000058
14. Wang YC, Huang TW, Yang YS, et al. Biofilm formation is not associated with worse outcome in Acinetobacter baumannii bacteraemic pneumonia. Sci Rep. 2018;8(1):7289. doi:10.1038/s41598-018-25661-9
15. He X, Lu F, Yuan F, et al. Biofilm formation caused by clinical Acinetobacter baumannii isolates is associated with overexpression of the AdeFGH efflux pump. Antimicrob Agents Chemother. 2015;59(8):4817–4825. doi:10.1128/AAC.00877-15
16. Hall GS. Non-fermenting and miscellaneous gram-negative bacilli. In: Mahon CR, Lehman DC, Manuselis G, editors. Textbook of Diagnostic Microbiology. Maryland Heights (MO): Saunders/Elsevier; 2011:482–501.
17. Turton JF, Woodford N, Glover J, Yarde S, Kaufmann ME, Pitt TL. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J Clin Microbiol. 2006;44(8):2974–2976. doi:10.1128/JCM.01021-06
18. CLSI M100-S28. Performance standards for antimicrobial susceptibility testing; Twenty-eight informational supplement. 2018.
19. Jones RN, Ferraro MJ, Reller LB, Schreckenberger PC, Swenson JM, Sader HS. Multicenter studies of tigecycline disk diffusion susceptibility results for Acinetobacter spp. J Clin Microbiol. 2007;45(1):227–230. doi:10.1128/JCM.01588-06
20. Ferreira AE, Marchetti DP, Cunha GR, et al. Molecular characterization of clinical multiresistant isolates of Acinetobacter sp. from hospitals in Porto Alegre, State of Rio Grande do Sul, Brazil. Rev Soc Bras Med Trop. 2011;44(6):725–730. doi:10.1590/S0037-86822011000600014
21. Qi L, Li H, Zhang C, et al. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in Acinetobacter baumannii. Front Microbiol. 2016;7:483. doi:10.3389/fmicb.2016.00483
22. Zhang D, Xia J, Xu Y, et al. Biological features of biofilm-forming ability of Acinetobacter baumannii strains derived from 121 elderly patients with hospital-acquired pneumonia. Clin Exp Med. 2016;16(1):73–80. doi:10.1007/s10238-014-0333-2
23. Gutiérrez D, Ruas-Madiedo P, Martínez B, Rodríguez A, García P. Effective removal of staphylococcal biofilms by the endolysin LysH5. PLoS One. 2014;9(9):e107307. doi:10.1371/journal.pone.0107307
24. Navidifar T, Amin M, Rashno M. Effects of sub-inhibitory concentrations of meropenem and tigecycline on the expression of genes regulating pili, efflux pumps and virulence factors involved in biofilm formation by Acinetobacter baumannii. Infect Drug Resist. 2019;12:1099–1111. doi:10.2147/IDR.S199993
25. Almasaudi SB. Acinetobacter spp. as nosocomial pathogens: epidemiology and resistance features. Saudi J Biol Sci. 2018;25(3):586–596. doi:10.1016/j.sjbs.2016.02.009
26. Asif M, Alvi IA, Rehman SU. Insight into Acinetobacter baumannii: pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities. Infect Drug Resist. 2018;11:1249–1260. doi:10.2147/IDR.S166750
27. Gonzalez-Villoria AM, Valverde-Garduno V. Antibiotic-resistant Acinetobacter baumannii increasing success remains a challenge as a nosocomial pathogen. J Pathog. 2016;2016:7318075. doi:10.1155/2016/7318075
28. Tavakol M, Momtaz H, Mohajeri P, Shokoohizadeh L, Tajbakhsh E. Genotyping and distribution of putative virulence factors and antibiotic resistance genes of Acinetobacter baumannii strains isolated from raw meat. Antimicrob Resist Infect Control. 2018;7:120. doi:10.1186/s13756-018-0405-2
29. Koeleman JGM, Stoof J, Van Der Bijl MW, Vandenbroucke-Grauls CMJE, Savelkoul PHM. Identification of epidemic strains of Acinetobacter baumannii by integrase gene PCR. J Clin Microbiol. 2001;39(1):8–13. doi:10.1128/JCM.39.1.8-13.2001
30. Fallah F, Noori M, Hashemi A, et al. Prevalence of blaNDM, blaPER, blaVEB, blaIMP, and blaVIM Genes among Acinetobacter baumannii Isolated from Two Hospitals of Tehran, Iran. Scientifica (Cairo). 2014;2014:245162.
31. Aksoy MD, Çavuşlu Ş, Tuğrul HM. Investigation of metallo beta lactamases and oxacilinases in carbapenem resistant Acinetobacter baumannii strains isolated from inpatients. Balkan Med J. 2015;32(1):79–83. doi:10.5152/balkanmedj.2015.15302
32. Mlynarcik P, Roderova M, Kolar M. Primer evaluation for PCR and its application for detection of carbapenemases in enterobacteriaceae. Jundishapur J Microbiol. 2016;9(1):e29314. doi:10.5812/jjm
33. Moura A, Henriques I, Ribeiro R, Correia A. Prevalence and characterization of integrons from bacteria isolated from a slaughterhouse wastewater treatment plant. J Antimicrob Chemother. 2007;60(6):1243–1250. doi:10.1093/jac/dkm340
34. Mirnejad R, Mostofi S, Masjedian F. Antibiotic resistance and carriage class 1and 2 integrons in clinical isolates of Acinetobacter baumannii from Tehran,Iran. Asian Pac J Trop Biomed. 2013;3(2):140–145. doi:10.1016/S2221-1691(13)60038-6
35. Taherikalani M, Fatolahzadeh B, Emaneini M, Soroush S, Feizabadi MM. Distribution of different carbapenem resistant clones of Acinetobacter baumannii in Tehran hospitals. New Microbiol. 2009;32(3):265–271.
36. Najar Peerayeh S, Karmostaji A. Molecular identification of resistance determinants, integrons and genetic relatedness of extensively drug resistant Acinetobacter baumannii isolated from hospitals in Tehran, Iran. Jundishapur J Microbiol. 2015;8(7):e27021. doi:10.5812/jjm
37. Michail G, Labrou M, Pitiriga V, et al. Activity of tigecycline in combination with colistin, meropenem, rifampin, or gentamicin against KPC-producing Enterobacteriaceae in a murine thigh infection model. Antimicrob Agents Chemother. 2013;57(12):6028–6033. doi:10.1128/AAC.00891-13
38. Garza-González E, Llaca-Díaz JM, Bosques-Padilla FJ, González GM. Prevalence of multidrug-resistant bacteria at a tertiary-care teaching hospital in Mexico: special focus on Acinetobacter baumannii. Chemotherapy. 2010;56(4):275–279. doi:10.1159/000319903
39. Baadani AM, Thawadi SI, El-Khizzi NA, Omrani AS. Prevalence of colistin and tigecycline resistance in Acinetobacter baumannii clinical isolates from 2 hospitals in Riyadh Region over a 2-year period. Saudi Med J. 2013;34(3):248–253.
40. Bahador A, Taheri M, Pourakbari B, et al. Emergence of rifampicin, tigecycline, and colistin-resistant Acinetobacter baumannii in Iran; spreading of MDR strains of novel international clone variants. Microb Drug Resist. 2013;19(5):397–406. doi:10.1089/mdr.2012.0233
41. Hasan B, Perveen K, Olsen B, Zahra R. Emergence of carbapenem-resistant Acinetobacter baumannii in hospitals in Pakistan. J Med Microbiol. 2014;63(1):50–55. doi:10.1099/jmm.0.063925-0
42. Rabin N, Zheng Y, Opoku-Temeng C, Du Y, Bonsu E, Sintim HO. Agents that inhibit bacterial biofilm formation. Future Med Chem. 2015;7(5):647–671. doi:10.4155/fmc.15.7
43. Rodríguez-Baño J, Martí S, Soto S, et al. Biofilm formation in Acinetobacter baumannii: associated features and clinical implications. Clin Microbiol Infect. 2008;14(3):276–278. doi:10.1111/j.1469-0691.2007.01916.x
44. Li J, Nation RL, Owen RJ, Wong S, Spelman D, Franklin C. Antibiograms of multidrug-resistant clinical Acinetobacter baumannii: promising therapeutic options for treatment of infection with colistin-resistant strains. Clin Infect Dis. 2007;45(5):594–598. doi:10.1086/520658
45. Gurung J, Khyriem AB, Banik A, Lyngdoh WV, Choudhury B, Bhattacharyya P. Association of biofilm production with multidrug resistance among clinical isolates of Acinetobacter baumannii and Pseudomonas aeruginosa from intensive care unit. Indian J Crit Care Med. 2013;17(4):214–218. doi:10.4103/0972-5229.118416
46. Abdi-Ali A, Hendiani S, Mohammadi P, Gharavi S. Assessment of biofilm formation and resistance to imipenem and ciprofloxacin among clinical isolates of Acinetobacter baumannii in Tehran. Jundishapur J Microbiol. 2014;7(1):e8606. doi:10.5812/jjm
47. Gallant CV, Daniels C, Leung JM, et al. Common beta-lactamases inhibit bacterial biofilm formation. Mol Microbiol. 2005;58(4):1012–1024. doi:10.1111/j.1365-2958.2005.04892.x
48. Macià MD, Rojo-Molinero E, Oliver A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin Microbiol Infect. 2014;20(10):981–990. doi:10.1111/1469-0691.12651
49. Ranjbar R, Karami A, Farshad S, Giammanco GM, Mammina C. Typing methods used in the molecular epidemiology of microbial pathogens: a how-to guide. New Microbiol. 2014;37(1):1–15.
50. Aljindan R, Alsamman K, Elhadi N. ERIC-PCR genotyping of Acinetobacter baumannii isolated from different clinical specimens. Saudi J Med Med Sci. 2018;6(1):13–17. doi:10.4103/sjmms.sjmms_138_16
51. Coyne S, Courvalin P, Périchon B. Efflux-mediated antibiotic resistance in Acinetobacter spp. Antimicrob Agents Chemother. 2011;55:947–953. doi:10.1128/AAC.01388-10
52. Sheikhalizadeh V, Hasani A, Ahangarzadeh Rezaee M, et al. Comprehensive study to investigate the role of various aminoglycoside resistance mechanisms in clinical isolates of Acinetobacter baumannii. J Infect Chemother. 2017;23(2):74–79. doi:10.1016/j.jiac.2016.09.012
53. Zhang Y, Li Z, He X, et al. Overproduction of efflux pumps caused reduced susceptibility to carbapenem under consecutive imipenem-selected stress in Acinetobacter baumannii. Infect Drug Resist. 2018;11(3):457–467. doi:10.2147/IDR.S151423
54. Wieczorek P, Sacha P, Hauschild T, Zórawski M, Krawczyk M, Tryniszewska E. Multidrug resistant Acinetobacter baumannii– the role of AdeABC (RND family) efflux pump in resistance to antibiotics. Folia Histochem Cytobiol. 2008;46(3):257–267. doi:10.2478/v10042-008-0056-x
55. Magnet S, Courvalin P, Lambert T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob Agents Chemother. 2001;45(12):3375–3380. doi:10.1128/AAC.45.12.3375-3380.2001
56. Marchand I, Damier-Piolle L, Courvalin P, Lambert T. Expression of the RND-type efflux pump AdeABC in Acinetobacter baumannii is regulated by the AdeRS two-component system. Antimicrob Agents Chemother. 2004;48(9):3298–3304. doi:10.1128/AAC.48.9.3298-3304.2004
57. Dou Q, Zou M, Li J, Wang H, Hu Y, Liu W. AdeABC efflux pump and resistance of Acinetobacter baumannii against carbapenem. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2017;42(4):426–433. doi:10.11817/j.issn.1672-7347.2017.04.010
58. Ardehali SH, Azimi T, Fallah F, Owrang M, Aghamohammadi N, Azimi L. Role of efflux pumps in reduced susceptibility to tigecycline in Acinetobacter baumannii. New Microbes New Infect. 2019;30:100547. doi:10.1016/j.nmni.2019.100547
59. Li H, Wang X, Zhang Y, et al. The role of RND efflux pump and global regulators in tigecycline resistance in clinical Acinetobacter baumannii isolates. Future Microbiol. 2015;10(3):337–346. doi:10.2217/fmb.15.7
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.