Back to Journals » Infection and Drug Resistance » Volume 12
Association between biofilm formation, structure and antibiotic resistance in Staphylococcus epidermidis isolated from neonatal septicemia in southwest Iran
Authors Farajzadeh Sheikh A, Asareh Zadegan Dezfuli A ![]() , Navidifar T
, Navidifar T ![]() , Fard SS, Dehdashtian M
, Fard SS, Dehdashtian M
Received 12 February 2019
Accepted for publication 7 May 2019
Published 27 June 2019 Volume 2019:12 Pages 1771—1782
DOI https://doi.org/10.2147/IDR.S204432
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Ahmad Farajzadeh Sheikh,1,2 Aram Asareh Zadegan Dezfuli,2 Tahereh Navidifar,2 Shahla Samei Fard,2 Masood Dehdashtian3
1Infectious and Tropical Diseases Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 2Department of Microbiology, Faculty of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran; 3Neonatology Ward Imam Khomeini Teaching Hospital, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran
Background: Staphylococcus epidermidis has emerged as the pathogen from neonatal septicemia. Antibiotic resistance and the capability of biofilm formation make these infections much harder to treat. Hence, the aim of this study was to investigate the association between biofilm formation, structure and antibiotic resistance in S. epidermidis isolated from neonatal septicemia.
Methods: Overall, 65 S. epidermidis isolates were recovered from blood cultures of neonatal septicemia. Antibiotic resistance pattern and the biofilm production were determined using phenotypic methods. The presence of ica operon, the bhp, the aap genes and SCCmec types were screened using PCR.
Results: Most S.epidermidis isolates were resistant to erythromycin, while all isolates were sensitive to linezolid and vancomycin. Fifty-three percent of S.epidermidis isolates were resistant to methicillin. SCCmec types II was found commonly among methicillin-resistant S. epidermidis (MRSE) strains. The biofilm formation was observed in 65% of S.epidermidis isolates and the majority have polysaccharide matrix. icaA and icaD genes were found in 40% and 19% of isolates. Twenty-three isolates (62%) produced dissolvable polysaccharide intercellular adhesion (PIA)-dependent biofilms in SM after growth in TSB with NaCl and 14 (37%) isolates produced dissolvable protein-dependent biofilms in PK after growth in TSB with glucose. Three isolates (62%) produced dissolvable polysaccharide intercellular adhesion.
Conclusion: Our data indicate the high rates of antibiotic resistance and the capability of biofilm formation among S. epidermidis isolates. Hence, the transmission of these strains can cause an increased risk of serious nosocomial infections.
Keywords: S. epidermidis, antibiotic resistance, biofilm formation
Introduction
Septicemia is one of the leading causes of neonatal mortality and morbidity worldwide. According to the World Health Organization report (WHO), more than 3 million newborns suffer from septicemia globally.1 According to a previous study in Iran, the rate of neonatal septicemia has been reported between 12% and 16.7%.2 In the last two decades, coagulase-negative staphylococci (CoNS) group especially Staphylococcus epidermidis, a normal flora of the skin, has emerged as a common cause of septicemia in the neonatal intensive care units (NICUs) especially in late-onset sepsis (LOS).3 Neonatal sepsis can be considered either as early onset sepsis (EOS) occurring in the first 72 hours of age or LOS which follows after 72 hours of age.4 LOS has been well recognized to be associated with prematurity, invasive interventions like intravascular catheterization, failure in early enteral feeding, prolonged antibiotic treatment, andhospitalization.5 The presence of S. epidermidis on human skin may allow S. epidermidis to form biofilm over medical implants and easily invade the bloodstream through indwelling catheters.6 The microorganisms attach to surfaces and produce extracellular polysaccharides, resulting in the formation of a biofilm.7 Biofilms are a serious problem for public health because of the increased resistance of biofilm-associated organisms to antimicrobial agents by slow diffusion of conventional antibiotics through the extracellular polymeric substance and the potential for these organisms to cause infections in patients with indwelling medical devices.7,8 The biofilm formation and the antibiotic resistance of the S. epidermidis isolates can be one of the important reasons for prolonging the period of treatment of infants.9 In S. epidermidis, the expression of several genes is associated with the biofilm formation, including icaABCD locus that encodes the polysaccharide intercellular adhesion (PIA), the bhp gene that encodes a cell wall surface anchor protein and the aap gene that encodes an accumulation-associated protein.10 Otto et al indicated that in strains that lack the ica locus, biofilm formation is due to the presence of aap gene, which enables bacteria to bind to various matrix proteins.11 Recently, increasing resistance of S. epidermidis strains to glycopeptide agents and methicillin has spurred high interests in understanding molecular mechanisms of antibiotic resistance.6,11 Similar to Staphylococcus aureus, the mechanism of methicillin resistance is mediated by the mecA gene which encodes penicillin binding protein 2a (PBP 2a) with reduced affinity for beta-lactam antibiotics.12 It is believed that CoNS acts as an important reservoir of resistance-associated mobile genetic elements, which can be transferred between staphylococcal species. The mecA gene is located on a mobile genetic element called the staphylococcal cassette chromosome mec (SCCmec). Moreover, SCCmec compromises two main components: the ccr gene complexccrand the mec gene complex. According to the combination of ccr allotypes with the mec gene complex, 11 types (I–XI) SCCmec have already been reported.13 In Iran, there are few published literature on the association between biofilm formation, structure, antibiotic resistance as well as SCCmec typing in S. epidermidis isolated. Hence, the aim of this study was to investigate the association between biofilm formation, structure and antibiotic resistance in S. epidermidis isolated from neonatal septicemia.
Methods and materials
Ethics
This research was conducted according to the Helsinki Declaration. This study was approved by the Research Ethics Committee of the Ahvaz Jundishapur University of Medical Sciences (No: IR.AJUMS.REC.1395. 332), Ahvaz, Iran, after submission of the preliminary proposal, and necessary permission for sample collection was granted. The study was accepted by the Imam Khomeini hospital data protection authority. After having read the information letter concerning the study, all respondents were asked for oral and written consent to participate. We emphasized that participation was voluntary and that parents could withdraw from the research at any time.
Sample collection
A total of 130 nonduplicate CoNS isolates were consecutively collected from April 2016 to February 2017 from blood cultures of 521 neonates with suspected septicemia hospitalized in the NICU, Imam Khomeini hospital. At least 1 mL of blood was collected from the peripheral vein of a neonate with suspected sepsis before starting antibiotics, either by a needle or a new cannula, using aseptic methods. The inclusion criteria of septicemia caused by CoNS were as follows: clinical signs of sepsis in a neonate older than 3 days of age, positive monomicrobial blood culture and elevated CRP >10 mg/L within 2 days of blood culture. Alternatively, CoNS blood cultures growing more than one organism were considered as contaminants.
Bacterial isolation
First, the isolates were subcultured on blood agar (EMD Millipore, Billerica, MA, USA) and then, the single colony was inoculated on Mannitol salt agar at 37°C for 24 hours (EMD Millipore) to identify S. epidermidis colonies. Then, suspicious colonies were subjected to biochemical tests, including gram staining, catalase, tube-coagulase, DNase and novobiocin susceptibility test (MAST Diagnostics, Merseyside, UK).14 The isolates were confirmed as S. epidermidis using the amplification of the sesC gene.15 S. epidermidis RP62A strain was used as positive control and distilled water as a negative control in all PCR reactions.
Investigation of susceptibility to antimicrobial agents
Antibiotic susceptibility testing was performed for 17 drugs covering all the nine antimicrobial categories comprising aminoglycosides, ansamycins, fluoroquinolones, folate pathway inhibitors, tetracyclines, glycopeptides, oxazolidinones, macrolides and incosamidesl were determined using the disc diffusion susceptibility test according to clinical and laboratory standards institute (CLSI) guidelines. Commercial antibiotic discs of rifampin (5 μg), linezolid (30 μg), ciprofloxacin (5 μg), levofloxacin (5 μg), minocyclin (30 μg), doxycycline (30 μg), tetracycline(10 μg), gentamycin (10 μg), tobramycin (10 μg), amikacin(1 μg), erythromycin(15 μg), azithromycin (15 μg), clarithromycin (15 μg),clindamycin (2 μg) trimethoprim/sulfamethoxazole (1.25/23.75 μg),(MAST Diagnostics, Merseyside UK) were used in disc diffusion test. Then, MDR/XDR/PDR phenotype of these isolates was established according to the results obtained from the disc diffusion test. Briefly, multidrug-resistance (MDR) was defined as resistance to at least three or more different classes, extensive drug-resistance (XDR) was defined as resistance to at least one agent in all but two or fewer antimicrobial categories and pan drug-resistance (PDR) was defined as resistance to all agents in all antimicrobial categories.16,17
Screening for vancomycin resistance
Resistance to vancomycin (MAST Diagnostics) was prepared by Mueller–Hinton agar (Merck, Germany) containing vancomycin 6 μg/mL and 4% NaCl. The plates were incubated at 37°C for 24 hours according to CLSI guidelines. Any visible growth after 24 hours was considered vancomycin resistant.
Screening for methicillin resistance
All of the S. epidermidis strains were tested for susceptibility to methicillin using a cefoxitin (30 µg) disc. Results were interpreted according to the criteria established by the CLSI. The presence of the mecA gene was evaluated using PCR amplification, as previously described by Shrestha et al.17 S. aureus ATCC 29213 strains were used as a positive control and distilled water as negative control.
Screening for of SCCmec elements
The presence of SCCmec genes in S. epidermidis strains were checked using PCR amplification, as previously explained by Moosavian et al.18 Five MRSA strains, NCTC10442 (SCCmec I), NCTC N315 (SCCmec II), NCTC 85/2082 (SCCmec III), NCTC CA05 (SCCmec IVa), and JCSC3624 (SCCmec V) were used as a positive control and distilled water as negative control.
Biofilm formation in 96-well microtiter plate
The biofilm formation capacity of isolates was evaluated using the crystal violet staining method. First, these isolates were inoculated in Mueller–Hinton agar at 37°C overnight. Then, these isolates were adjusted to 0.5 McFarland (~1.5×108 CFU/mL) with normal saline (0.85% NaCl). A 10-μL aliquot of each suspension was then diluted 1:200 in 190 μL of tryptic soy broth (TSB) containing 1% glucose in 96-well polystyrene microtiter plates. Following incubation at 37°C overnight, the plates were washed three times with PBS, fixed by adding 200 μL of methanol into each well, and stained with 200 μL of 0.1% crystal violet (CV) for 20 minutes. The plates were again washed three times to remove excess stain, and the remaining CV was solubilized by incubating with 200 μL of 95% ethanol for 10 minutes. The optical density at 570 nm (OD570) of each well was measured by the ELISA plate reader (μQuant; BioTek Instruments, Winooski, VT, USA), to evaluate the biofilm formation capacity. S. epidermidis ATCC 35984 and TSB broth were used as positive and negative controls (ODc) for the biofilm formation, respectively. The results were interpreted according to the criteria suggested by Zhang et al. Briefly, the isolates were classified into the several groups about the biofilm formation capacity: OD570≤ODc=no biofilm producer; ODc<OD570≤2×ODc=weak biofilm producer; 2×ODc<OD570≤4×ODc=moderate biofilm producer; and 4×ODc<OD570=strong biofilm producer.19 All experiments were performed in triplicate.
Congo red agar test
Congo red agar (CRA) medium was composed of brain heart infusion broth (BHI, Oxoid; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with NaCl 1.5%, sucrose 5% and 0.08% Congo red.20 The inoculated CRA plates were incubated at 37°C overnight. Then, isolates were interpreted according to their colony phenotypes. Black colonies were indicative of biofilm production, while red colonies were considered as the nonbiofilm producers. S. epidermidis ATCC 35984 and S. epidermidis ATCC 12228 strains were used as biofilm-positive and biofilm-negative controls, respectively.
Biochemical characterization of the biofilm matrix
To determine the biochemical characterization of the biofilm matrix, first the biofilm formation was induced as described above. Then, all wells were washed with PBS and treated for 1 hour at37°C either with a solution of 10 mM sodium metaperiodate in 50 mM sodium acetate buffer (pH 4.5) to disrupt the extracellular polysaccharides, or with 100 µg/ml of proteinase K (SinaClon, Tehran, Iran) in 20 mM Tris (pH 7.5) and 100 mM NaCl to disrupt the biofilm extracellular protein. Subsequently, the biofilms were washed, fixed and stained with CV, and the optical absorbance (570 nm) measured as described above.21
Detection of genes involved in biofilm formation
All S. epidermidis isolates were screened using PCR for the presence of the ica operon (icaA, icaB, icaC, icaD), the bhp and the aap genes using primers and conditions as previously described. S. epidermidis RP62A was positive control for ica operon and S. epidermidis 1457 Δica was positive control for aap and bhp genes.22
Results
In this study, 215 out of 512 neonates with suspected septicemia had positive blood cultures. Of these 215 cases, 135(62.79%) CoNS strains were isolated from blood cultures. Moreover, of the 135 CoNS isolates, 65 (48.14%) isolates were confirmed as S. epidermidis using biochemical tests and PCR. Demographic and clinical features at the time of presentation of neonatal had septicemia confirmed by S. epidermidis are shown in Table 1. The resistance to methicillin in S. epidermidis was recognized by cefoxitin disk in 39 (60%) isolates and the amplification of mecA gene in 35(53%) isolates. The distribution of SCCmec types in the 35
 |
Table 1 Demographic and clinical features at the time of presentation of neonatal septicemia |
Methicillin-resistant S. epidermidis (MRSE) isolates showed that SCCmec types II and III dominated among the tested isolates (Table 2). Of the 35 MRSE isolates, 16 (45%) carried type II SCCmec, 11 (31%) carried type III SCCmec and 4(11%) carried type I and IV SCCmec respectively. None of the tested isolates had type IV and two isolates were untypeable by the routine PCR assays used (Table 2). According to antibiogram results, the majority of S. epidermidis were resistance to the antibiotics used (Figure 1). The maximum resistance was found to erythromycin (81%), clindamycin and amikacin (52%), gentamicin (46%), ciprofloxacin (44%), tobramycin (33%), tetracycline (24%), clarithromycin (16%), rifampin and minocycline (15% each), azithromycin and trimethoprim-sulfamethoxazole (13%), doxycycline (12%), levofloxacin (10%). Also, all isolates were sensitive to linezolid and vancomycin.
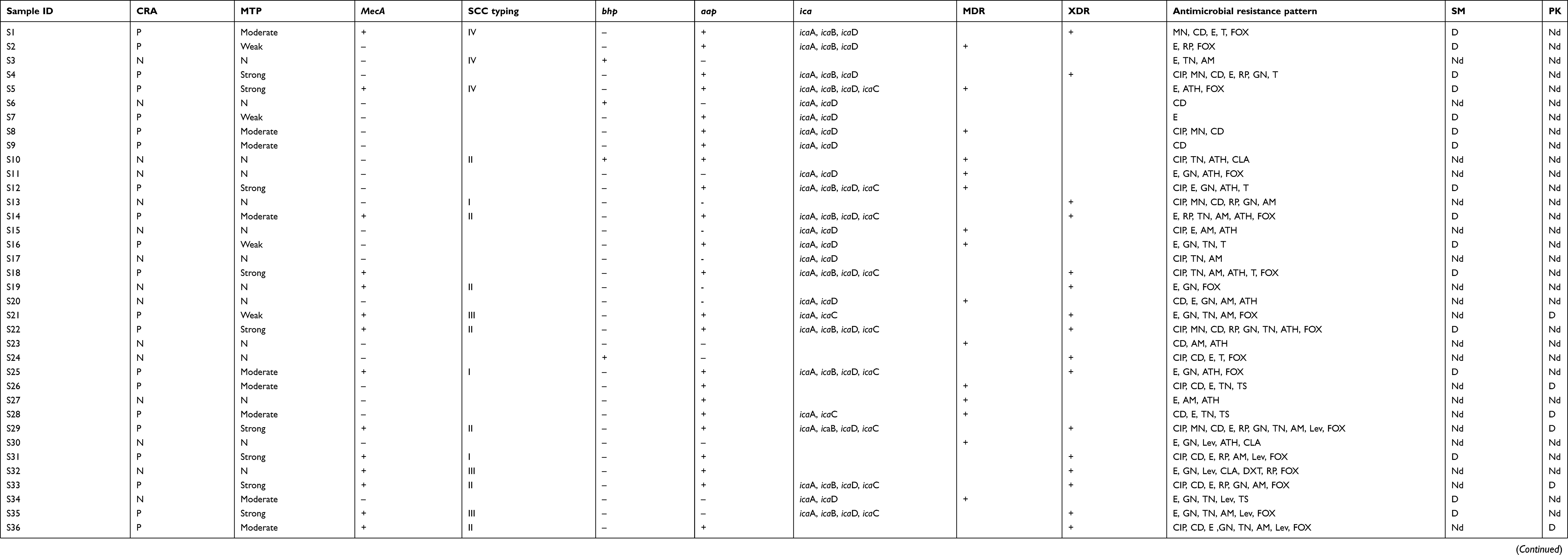 |
Table 2 Biofilm formation structure antibiotic resistance and biofilm related genes in all S. epidermidis isolat |
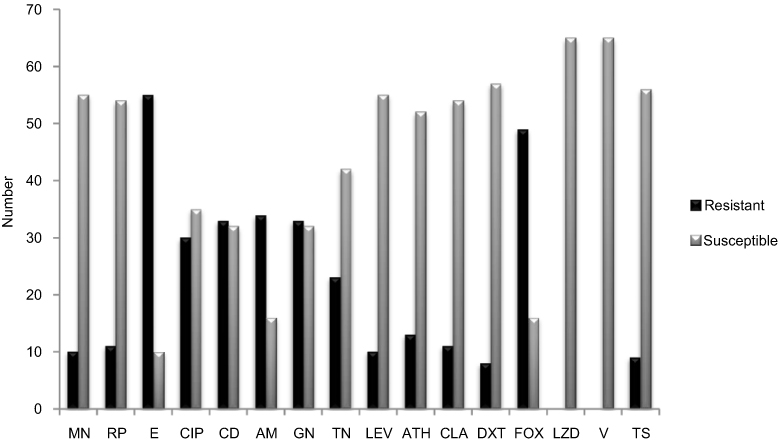 |
Figure 1 Antibiotic resistance rate among all 65 S.epidermidis isolated. Abbreviations: AM, amikacin; GN, gentamicin; FOX, cefoxitin; CIP, ciprofloxacin; E, erythromycin; T, tetracycline; TS, sulfamethoxazole-trimethoprim; ATH, azithromycin; CLA, clarithromycin; LZD, linezolid; RP, rifampin; DXT, doxycycline; MN, minocycline; TN, tobramycin; CD, clindamycin; Lecv, levofloxacin; V, vancomycin. |
Twenty-one (32.30%) of the S. epidermidis isolates were MDR and 37 (56%) were XDR. None of our isolates were PDR. The resistance rates to ciprofloxacin, Erythromycin, gentamicin, and clindamycin were found to be extremely high among the MRSE isolates compared to those that were methicillin-susceptible S. epidermidis (MSSE).
In the CRA assay, 32(49%) isolates were considered as producing biofilm and produced black colonies whereas 33 (50%) isolates produced red colonies. The biofilm production analysis by MTP method differentiated isolates into strong, moderate, weak, and nonbiofilm-forming according to the OD values at 570 nm. Again, 37of the isolates 17 (45%) were categorized as strong biofilm-formers; 12 isolates (32%) were moderate; 8(21%) were weak biofilm-formers (48%); and 28 (43%) could not form any detectable biofilm. The OD570 values for the reference strain (ATCC 35,984) and negative control were 0.416±0.048 and 0.074±0.010, respectively. The composition of the biofilm matrix of 23(62%) S. epidermidis isolates were polysaccharide and 14 (37%) isolates were protein (Table 2). In the 17 strong biofilm-formers, 13(76%) isolates were XDR and 4(23%) isolates were MDR. Among the 12 moderate biofilm-formers, 6(50%) isolates were XDR and 5(41%) isolates were MDR. Also, of the eight weak biofilm-formers, 4(50%) isolates were XDR and 2(25%) isolates were MDR (Table 2). The biofilm producers were more resistant to all antibiotics used to expect linezolid and vancomycin than biofilm nonproducers. The correlation of biofilm-forming capacity and antibiotic resistance is summarized in the (Table 3) (P<0.001). The distribution of SCCmec types among S. epidermidis isolates of biofilm producers and nonbiofilm producers is shown in Figure 2. According to these results, SCCmec type II was commonly found among biofilm producers Figure 3. The presence of icaA, icaB, icaC, icaD, bhp, and aap genes was confirmed by PCR among 40(61%), 19(29%), 12(8%), 35(53%), 10(15%), and 36(55%) isolates, respectively (Table 2). Among 37 biofilm producing strains harbored icaA and icaB, icaC, and icaD genes respectively and (81%) and (24%) of strains were positive for aap and bhp genes. Furthermore, in nonbiofilm producing strains, 21%, 17%, and 30% of the isolates carried icaA, icaB, and icaD genes. icaC was not detected in nonbiofilm producing strains. Table 2 summarizes the antibiotic resistant, adhesions analysis of the genetic factors and biofilm production in S. epidermidis isolates. We assessed the relationship between biofilm formation capacity and genes involved in this process (Table 4). Out of 17 strong biofilm-formers, 88% strains were positive for the icaA and icaD genes while 60%, 52%, 60%, and 11% were positive for the icaB, icaC, aap, and bhp genes respectively. Also, among 12 moderate biofilm-formers, 10 (83%), 5 (41%), 4 (33%), 8 (66%), 1 (8%), and 11 (91%) strains were positive for the icaA,caB, icaC, icaD, aap, and bhp. Alternatively, out of eight weak biofilm-formers, 2(25%) strains were positive for the icaB and icaC genes, followed by the icaA 7(78%), icaD 6(75%), aap 6(75%), and weak biofilm-formers did not harbor bhp gene. There is a significant association between the biofilm formation and genes encoding ica operon, and aap gene (P<0.001). The presence of all genes involved in biofilm formation among biofilm positive strains was significantly higher than nonbiofilm-producing strains (Figure 4).
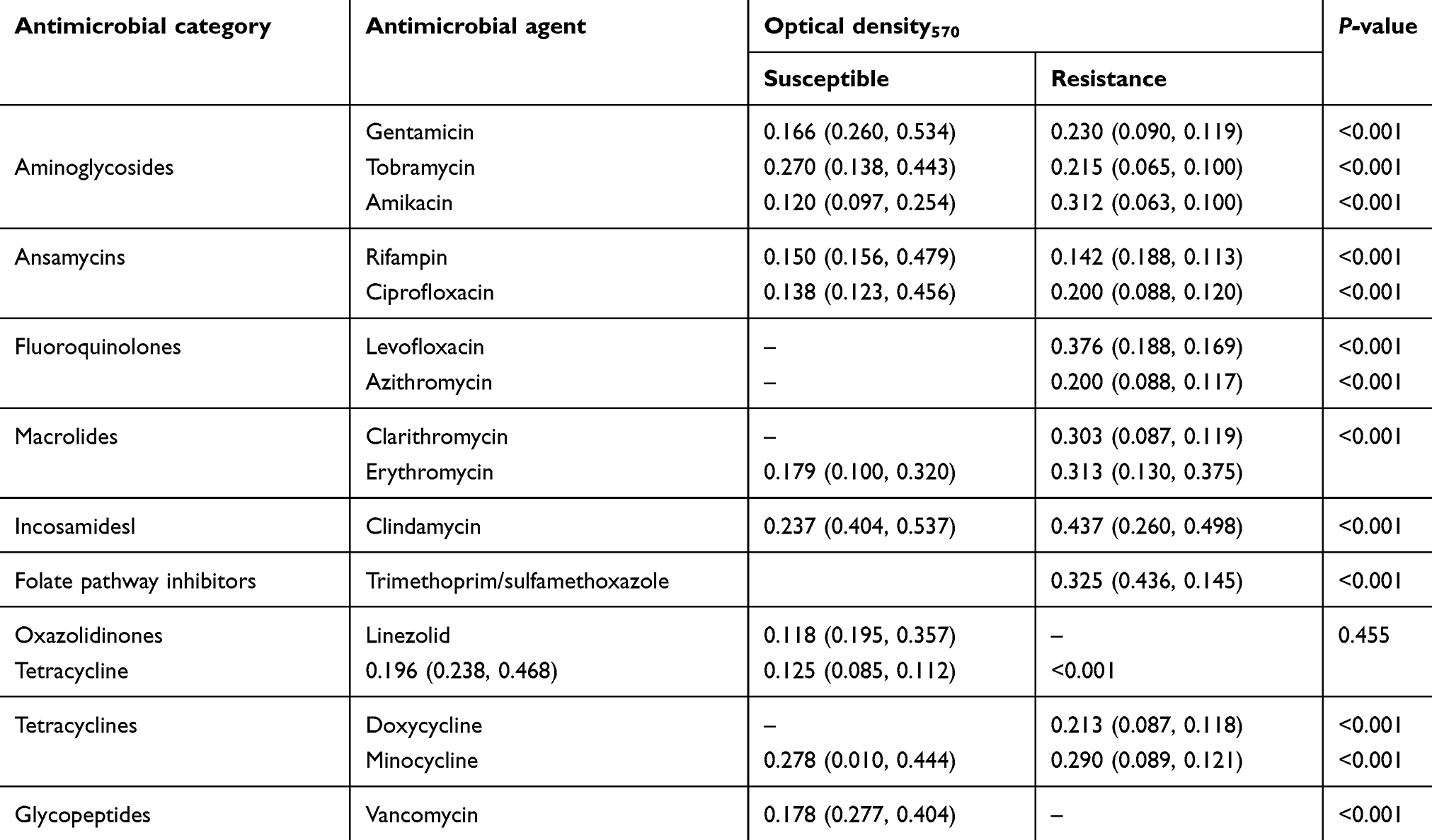 |
Table 3 Correlation between the level of biofilm formation and antibiotic resistance in 65 S. epidermidis isolates |
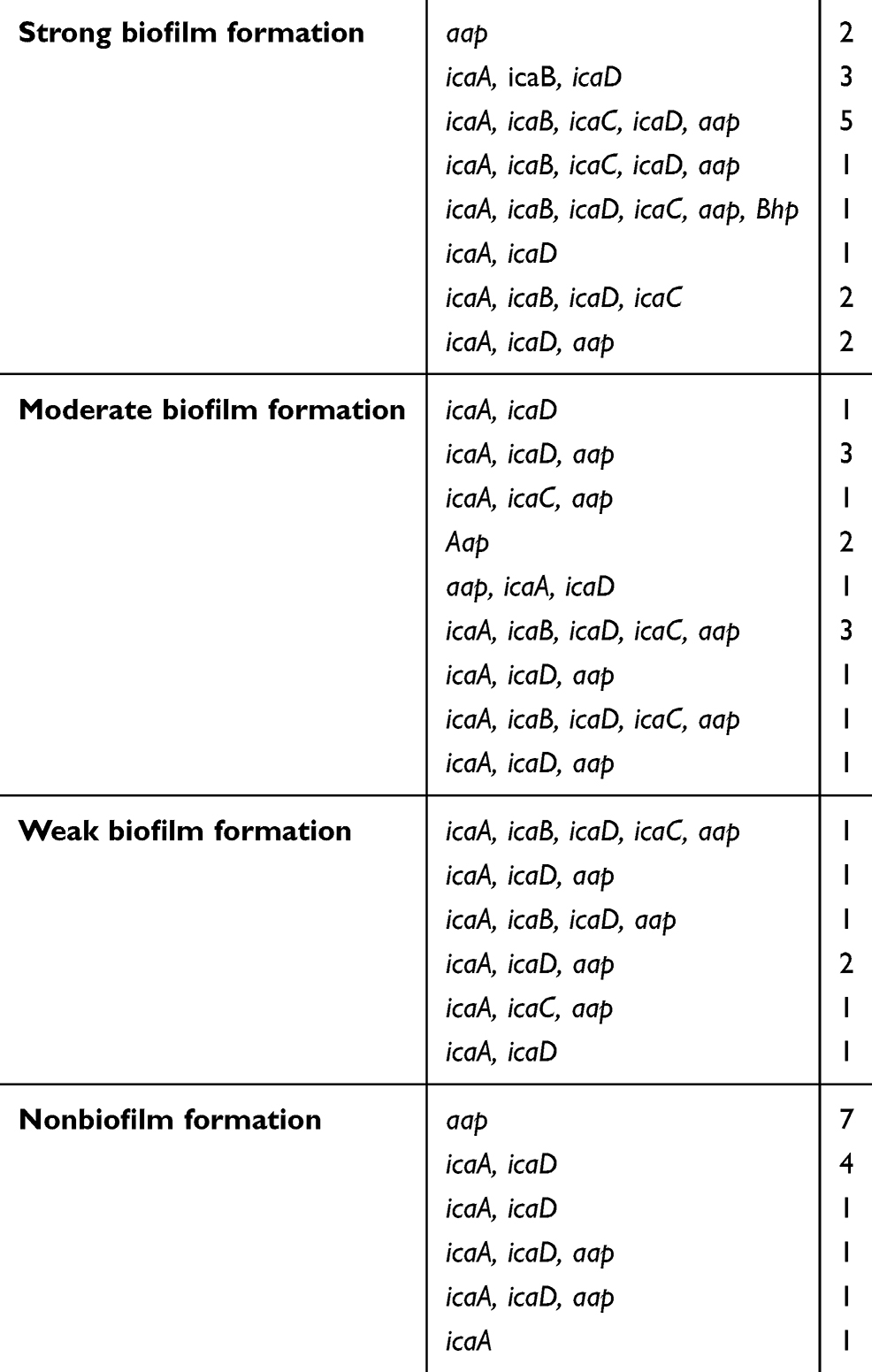 |
Table 4 The pattern of the genes that make up biofilms is based on the biofilm capacity |
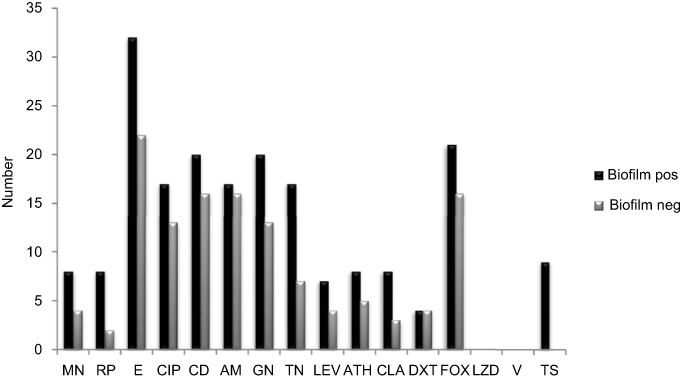 |
Figure 2 Comparison of antimicrobial susceptibilities between biofilm and nonbiofilm producing S. epidermidis isolated. Abbreviations: AM, amikacin; GN, gentamicin; FOX, cefoxitin; CIP, ciprofloxacin; E, erythromycin; T, tetracycline; TS, sulfamethoxazole-trimethoprim; ATH, azithromycin; CLA, clarithromycin; LZD, linezolid; RP, rifampin; DXT, doxycycline; MN, minocycline; TN, tobramycin; CD, clindamycin; Lecv, levofloxacin; V, vancomycin; Biofilm pos, biofilm positive; Biofilm neg, biofilm negative. |
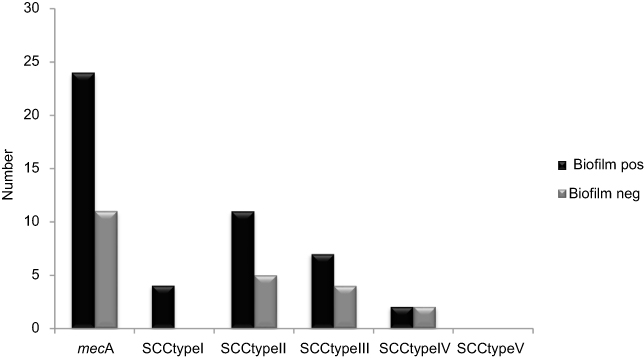 |
Figure 3 Distribution of SCC typing in S. epidermidis biofilm producer and nonbiofilm producer. Abbreviations: mecA, methicillin-resistant gene; SCC, staphylococcal cassette chromosome; Biofilm pos, biofilm positive; Biofilm neg, biofilm negative. |
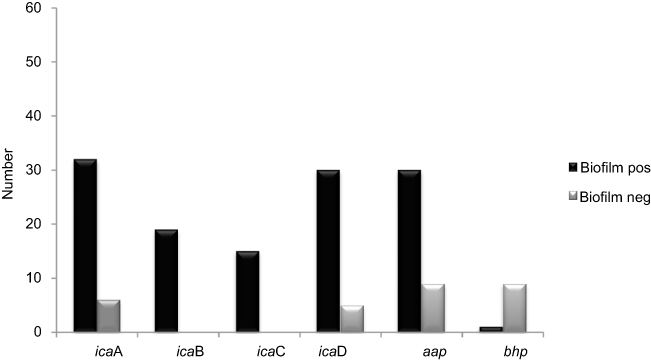 |
Figure 4 Distribution of biofilm formation genes among biofilm producer and nonbiofilm producer S. epidermidis isolated. Abbreviations: ica, intercellular adhesion; aap, accumulation-associated protein; bhp, biofilm associated protein; Biofilm pos, biofilm positive; Biofilm neg, biofilm negative. |
Discussion
Bacteria present in human skin thus have the opportunity to control cell behaviors below the surface. Examples of beneficial functions induced by specific skin bacteria include the capacity23 Also, a prior study has described that S.epidermidis induces the secretion of antimicrobial peptide, which increased the capacity of cell lysates to inhibit the growth of group A Streptococcus and S.aureus. Nevertheless, S.epidermidis biofilm-associated infections are increasing the use of indwelling or implanted medical devices. These often can spread into the bloodstream and cause nosocomial sepsis.23,24 In this study, the prevalence of septicemia associated with CoNS in the neonatal population during one year was 62.8%. In agreement with our study, the incidence of septicemia associated with CoNS evaluated in other studies ranged from 34% to 48%.25,26 The incidence of LOS increases up to 50% in the immature preterm infants and very low birth weight (VLBW) infants have a high danger of septicemia associated with CoNS.27 In our study, the clinical isolate of S. epidermidis was more common in neonatal have immature preterm infants and in very low birth weight (<1500 g) (Table 1).
In the present study, the high prevalence of MRSE in the NICUhighlighted the importance of a suitable choice of antibiotic therapy and to develop treatments against S. epidermidis infections. Earlier studies have shown that the proportion of MRSE in Iran, with an incidence of 31% to 89%.28,29 Due to the wide variation seen in the different reports in Iran it could be in part due to different populations and geographical locations, and the quality of hospital sampling carried out. Four of the MRSE isolated appeared susceptible to cefoxitin but were negative to harbor mecA. Such strains are perhapsvery heterogeneous in their expression of methicillin resistance. Also a previous study reported the complete absence of five major SCCmec types and mecA gene as well as the gene product of PBP2a in isolates which were phenotypically MRSA suggesting a probability of hyper production of β-lactamase as a cause of the phenomenon.30 In agreement with this study, another study has shown most S. epidermidis isolates were resistant to aminoglycoside, fluoroquinolone, and macrolide agents.31 In our study, 56.3% of S. epidermidis isolates were XDR (≥3 antibiotic classes). Considering the presence of XDR S. epidermidis in NICUcan cause infection and would be more complicated to treat. In our study, the majority of the S. epidermidis isolated had the ability of biofilm production but with different capacities. The mechanisms responsible for antimicrobial resistance in organisms producing biofilms may be one or more of the following, such as the poor diffusion of the antimicrobial penetration through the biofilm extracellular matrix, the different growth rate of biofilm organisms, etc. Thus, the ability to form biofilm could be an effective strategy to enhance the survival and persistence under stressed conditions like host invasion or antibiotic treatment.32–35 Confirming these, in our study; we found a significant correlation between the capacity of biofilm formation and antibiotic resistance (P<0.001). In other words, the biofilm density in S. epidermidis resistance strains was more than susceptible strains. Inconsistent with our study, some researchers demonstrated that the resistant isolates were stronger producers of biofilm than the susceptible isolates.16 Together, in this study, the evaluation and comparison of biofilm formation between nonMDR and MDR/XDR have shown that the majority of the MDR/XDR isolates have a significantly higher capacity to form biofilms compared to nonMDR isolated. In S. epidermidis isolates, the ica operon appears to play an essential role in biofilm formation. As found by other authors, our data indicate that the prevalence of the ica operon in S. epidermidis isolates from neonatal septicemia was 61.50%.36,37 In our study, even though a significant difference was found among biofilm producers and nonproducers in the bap gene (24% vs 2.7%). This percentage is significantly lower than previously reported.38 Biofilm formation is also associated with the presence of ica operon and aap genes which is responsible for the production of the PIA and proteinaceous structure of biofilm respectively. Previous studies have shown that the biofilm formation in staphylococci is associated with the presence of both icaA and icaD genes.The expressions of these genes are essential for the full phenotypic expression of biofilm in clinical staphylococcal isolates.39,40
However, in accordance with our findings which demonstrated that the presence of the ica operon was not always associated with biofilm production. Infact four biofilm-producing isolates did not carry the ica operon but indicated the ability of biofilm formation. In conclusion, to our knowledge, recognized risk factors for postpartum septicemia may not always be present, and signs of severe septicemia may be masked or present atypically. In this study, we report a high prevalence of virulence/antimicrobial resistance determinants in S. epidermidis from neonatal septicemia, although the reasons for the increases of invasive S.epidemidis infection are unclear because it has different reasons. Environmental factors that influence S.epidemidis septicemia such as hygiene, maternity setting, health-care workers, social and family contacts,etc, and the host and the pathogen factors affecting the transmission of S.epidemidis. However, processes of infection control including identifying patients at risk of nosocomial infections, observing hand hygiene, include identifying sources of organism, identification of organisms, isolation if required, antibiotic prophylaxis to be used selectively following standard precautions to reduce transmission and strategies to reduce infections. Fortunately, the result of antibiotic resistance in S. epidermidis isolates demonstrated thatlinezolid and vancomycin showgood effects in treatment. Furthermore, a high rate of prevalence of biofilm-forming strains among the tested S. epidermidis isolates was detected, but, S. epidermidis forms a prototypic biofilm, and that biofilm formation in vitro does not necessarily correlate with biofilm formation in vivo.
Acknowledgments
This work is a part of the M.Sc. thesis of Aram Asarehzadegan Dezfuli, which has been approved in the Department of Microbiology of Ahvaz Jundishapur University of Medical Sciences. The authors thank the Department of Microbiology, School of Medicine, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran and Infectious and Tropical Diseases Research Center, Health Research Institute, Ahvaz Jundishapur University of Medical Sciences, Ahvaz, Iran, for financial support (No: IR.AJUMS.REC.1395. 332). The study was sponsored by the authors. The authors received no funding from any other individual or institution.
Author contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. World Health Organization. Sepsis. WHO; 2018. Available from: https://www.who.int/news-room/fact-sheets/detail/sepsis. Accessed April 19, 2018.
2. Sayehmiri K, Nikpay S, Azami M, Pakzad I, Borji M. The prevalence of neonatal septicemia in Iran: a systematic review and meta-analysis study. J Shahrekord Uni Medl Sci. 2017;19:158–169.
3. Thakur S, Thakur K, Sood A, Chaudhary S. Bacteriological profile and antibiotic sensitivity pattern of neonatal septicaemia in a rural tertiary care hospital in North India. Indian J Med Microbiol. 2016;34(1):67. doi:10.4103/0255-0857.174108
4. Cohen-Wolkowiez M, Moran C, Benjamin DK, et al. Early and late onset sepsis in late preterm infants. J Pediatr Infect Dis. 2009;28(12):1–13. doi:10.1097/INF.0b013e3181acf6bd
5. Dong Y, Speer CP. Late-onset neonatal sepsis: recent developments. Arch Dis Child Fetal Neonatal Ed. 2015;100(3):F257–F263. doi:10.1136/archdischild-2014-306213
6. Lee JY, Monk IR, Da Silva AG, et al. Global spread of three multidrug-resistant lineages of Staphylococcus epidermidis. Nat Microbiol. 2018;3(10):1175. doi:10.1038/s41564-018-0230-7
7. Donlan RM. Biofilm formation: a clinically relevant microbiological process. Clin Microbiol Infect Dis. 2001;33(8):1387–1392.
8. Li J, Xie S, Ahmed S, et al. Antimicrobial activity and resistance: influencing factors. Front Pharmacol. 2017;13(8):364. doi:10.3389/fphar.2017.00364
9. Klingenberg C, Aarag E, Rønnestad A, et al. Coagulase-negative staphylococcal sepsis in neonates: association between antibiotic resistance, biofilm formation and the host inflammatory response. Pediatr Infect Dis J. 2005;24(9):817–822.
10. Schaeffer CR, Woods KM, Longo GM, et al. Accumulation-associated protein enhances Staphylococcus epidermidis biofilm formation under dynamic conditions and is required for infection in a rat catheter model. Infect Immun. 2015;83(1):214. doi:10.1128/IAI.02177-14
11. Otto M. Molecular basis of Staphylococcus epidermidis infections. Semin Immunopathol. 2012;34(2):201–214. doi:10.1007/s00281-011-0296-2
12. Hiramatsu K, Ito T, Tsubakishita S, et al. Genomic basis for methicillin resistance in Staphylococcus aureus. Infect Chemother. 2013;45(2):117–136. doi:10.3947/ic.2013.45.2.117
13. Saber H, Jasni AS, Jamaluddin TZ, Ibrahim R. A review of Staphylococcal cassette chromosome mec (SCCmec) types in coagulase-negative staphylococci (CoNS) species. Malays J Med Sci. 2017;24(5):7–18.
14. Mahon CR, Lehman DC, Manuselis JG. Textbook of Diagnostic Microbiology. Amsterdam: Elsevier Health Sciences; 2014.
15. Khodaparast L, Khodaparast L, Van Mellaert L, Shahrooei M, Van Ranst M, Van Eldere J. sesC as a genetic marker for easy identification of Staphylococcus epidermidis from other isolates. Infect Genet Evol. 2016;43:222–224. doi:10.1016/j.meegid.2016.05.037
16. Performance standards for antimicrobial susceptibility testing: sixteenth informational supplement. Clinical and Laboratory Standards Institute; 2016
17. Sharma V, Jindal N, Devi P. Prevalence of methicillin resistant coagulase negative staphylococci in a tertiary care hospital. Iran J Microbiol. 2010;2(4):185–188.
18. Moosavian M, Shahin M, Navidifar T, Torabipour M. Typing of staphylococcal cassette chromosome mec encoding methicillin resistance in Staphylococcus aureus isolates in Ahvaz, Iran. New Microbes New Infect. 2018;21:90–94. doi:10.1016/j.nmni.2017.11.006
19. Zhang D, Xia J, Xu Y, et al. Biological features of biofilm-forming ability of Acinetobacter baumannii strains derived from 121 elderly patients with hospital-acquired pneumonia. Clin Exp Med. 2016;16(1):73–80. doi:10.1007/s10238-014-0333-2
20. Kaiser TD, Pereira EM, Dos Santos KR, Maciel EL, Schuenck RP, Nunes AP. Modification of the Congo red agar method to detect biofilm production by Staphylococcus epidermidis. Diagn Microbiol Infect Dis. 2013;75(3):235–239. doi:10.1016/j.diagmicrobio.2012.11.014
21. Gutierrez D, Ruas-Madiedo P, Martínez B, Rodríguez A, García P. Effective removal of staphylococcal biofilms by the endolysin LysH5. PLoS One. 2014;9(9):e107307. doi:10.1371/journal.pone.0107307
22. Calà C, Amodio E, Di Carlo E, Virruso R, Fasciana T, Giammanco A. Biofilm production in Staphylococcus epidermidis strains, isolated from the skin of hospitalized patients: genetic and phenotypic characteristics. New Microbiol. 2015;38(4):521–529.
23. Nguyen TH, Park MD, Otto M. Host response to Staphylococcus epidermidis colonization and infections. Front Cell Infect Microbiol. 2017;21(7):90.
24. Lai Y, Di Nardo A, Nakatsuji T, et al. Commensal bacteria regulate Toll-like receptor 3–dependent inflammation after skin injury. Nat Med. 2009;15(12):1377. doi:10.1038/nm.2062
25. Shobowale EO, Solarin AU, Elikwu CJ, Onyedibe KI, Akinola IJ, Faniran AA. Neonatal sepsis in a Nigerian private tertiary hospital: bacterial isolates, risk factors, and antibiotic susceptibility patterns. Ann Afr Med. 2017;16(2):52. doi:10.4103/aam.aam_34_16
26. Verma P, Berwal PK, Nagaraj N, Swami S, Jivaji P, Narayan S. Neonatal sepsis: epidemiology, clinical spectrum, recent antimicrobial agents and their antibiotic susceptibility pattern. Int J Contemp Pediatrics. 2017;2(3):176–180.
27. Lin CB, Hornik CP, Clark R, et al. Very low birth weight neonates who survive early-onset sepsis do not have an increased risk of developing late-onset sepsis. Early Hum Dev. 2012;88(11):905–909. doi:10.1016/j.earlhumdev.2012.07.009
28. Soroush S, Jabalameli F, Taherikalani M, et al. Investigation of biofilm formation ability, antimicrobial resistance and the staphylococcal cassette chromosome mec patterns of methicillin resistant Staphylococcus epidermidis with different sequence types isolated from children. Microb Pathog. 2016;2016(93):1–24.
29. Ramazanzadeh R, Salimizand H, Shahbazi B, Narenji H. Prevalence of mecA gene of methicillin resistant Staphylococcus spp. Isolated from nosocomial infections and environmental specimens in Sanandaj Hospitals, Kurdistan, Iran. Res Mol Med. 2015;3(3):38–42.
30. Elhassan MM, Ozbak HA, Hemeg HA, Elmekki MA, Ahmed LM. Absence of the mecA gene in methicillin resistant Staphylococcus aureus isolated from different clinical specimens in shendi city, Sudan. Biomed Res Int. 2015;2015:1–5. doi:10.1155/2015/895860
31. Sahal G, Bilkay IS. Multi drug resistance in strong biofilm forming clinical isolates of Staphylococcus epidermidis. Braz J Microbiol. 2014;45(2):539–544.
32. Xu LC, Siedlecki CA. Submicron-textured biomaterial surface reduces staphylococcal bacterial adhesion and biofilm formation. Acta Biomater. 2012;8(1):72–81. doi:10.1016/j.actbio.2011.08.009
33. Baillie GS, Douglas LJ. Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J Antimicrob Chemother. 2000;46(3):397–403.
34. Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002;15(2):167–193.
35. Gurung J, Khyriem AB, Banik A, Lyngdoh WV, Choudhury B, Bhattacharyya P. Association of biofilm production with multidrug resistance among clinical isolates of Acinetobacter baumannii and Pseudomonas aeruginosa from intensive care unit. Indian J Crit Care Med. 2013;17(4):214. doi:10.4103/0972-5229.118416
36. Stevens NT, Tharmabala M, Dillane T, Greene CM, O’Gara JP, Humphreys H. Biofilm and the role of the ica operon and aap in Staphylococcus epidermidis isolates causing neurosurgical meningitis. Clin Microbiol Infect. 2008;14(7):719–722. doi:10.1111/j.1469-0691.2008.02012.x
37. De Silva GD, Kantzanou M, Justice A, et al. The ica operon and biofilm production in coagulase-negative staphylococci associated with carriage and disease in a neonatal intensive care unit. J Clin Microbiol. 2002;40(2):382–388.
38. Tormo MA, Knecht E, Götz F, Lasa I, Penades JR. Bap-dependent biofilm formation by pathogenic species of Staphylococcus: evidence of horizontal gene transfer? Microbiol. 2005;151(7):2465–2475. doi:10.1099/mic.0.27865-0
39. Zhou S, Chao X, Fei M, Dai Y, Liu B. Analysis of S. Epidermidis icaA and icaD genes by polymerase chain reaction and slime production: a case control study. BMC Infect Dis. 2013;13(1):242. doi:10.1186/1471-2334-13-242
40. Gad GF, El-Feky MA, El-Rehewy MS, Hassan MA, Abolella H, El-Baky RM. Detection of icaA, icaD genes and biofilm production by Staphylococcus aureus and Staphylococcus epidermidis isolated from urinary tract catheterized patients. J Infect Dev Ctries. 2009;3(05):342–351.
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.