Back to Journals » Hepatic Medicine: Evidence and Research » Volume 16
Direct Ingestion of Oxidized Red Blood Cells (Efferocytosis) by Hepatocytes
Authors Zheng C ![]() , Li S, Lyu H, Chen C, Mueller J
, Li S, Lyu H, Chen C, Mueller J ![]() , Dropmann A, Hammad S, Dooley S, He S, Mueller S
, Dropmann A, Hammad S, Dooley S, He S, Mueller S ![]()
Received 29 May 2024
Accepted for publication 14 August 2024
Published 3 September 2024 Volume 2024:16 Pages 65—77
DOI https://doi.org/10.2147/HMER.S469990
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Gerry Lake-Bakaar
Supplementary video of “Red blood cell ingestion by hepatocytes” [469990].
Views: 604
Chaowen Zheng,1,2 Siyuan Li,1 Huanran Lyu,1 Cheng Chen,1 Johannes Mueller,1 Anne Dropmann,3 Seddik Hammad,3,4 Steven Dooley,3 Songqing He,2 Sebastian Mueller1,5
1Center for Alcohol Research, University of Heidelberg, Heidelberg, Germany; 2Division of Hepatobiliary Surgery, the First Affiliated Hospital of Guangxi Medical University, Nanning, Guangxi, People’s Republic of China; 3Molecular Hepatology Section, Medical Faculty Mannheim, Heidelberg University, Heidelberg, Germany; 4Department of Forensic Medicine and Veterinary Toxicology, Faculty of Veterinary Medicine, South Valley University, Quena, Egypt; 5Viscera AG Bauchmedizin, Bern, Switzerland
Correspondence: Sebastian Mueller, Center for Alcohol Research, University Hospital Heidelberg Im Neuenheimer Feld 350 69120 Heidelberg, Germany, Tel +49 6221 567121, Email [email protected]
Purpose: Both hepatic iron accumulation and hemolysis have been identified as independent prognostic factor in alcohol-related liver disease (ALD); however, the mechanisms still remain poorly understood. We here demonstrate that hepatocytes are able to directly ingest aged and ethanol-primed red blood cells (RBCs), a process termed efferocytosis.
Methods: Efferocytosis of RBCs was directly studied in vitro and observed by live microscopy for real-time visualization. RBCs pretreated with either CuSO4 or ethanol following co-incubation with Huh7 cells and murine primary hepatocytes. Heme oxygenase-1 (HO-1) and other targets were measured by q-PCR.
Results: As shown by live microscopy, oxidized RBCs, but not intact RBCs, are rapidly ingested by both Huh7 cells and murine primary hepatocytes within 10 minutes. In some cases, more than 10 RBCs were seen within hepatocytes, surrounding the nucleus. RBC efferocytosis also rapidly induces HO1, its upstream regulator Nuclear factor erythroid 2-related factor 2 (Nrf2) and ferritin, indicating efficient heme degradation. Preliminary data further suggest that hepatocyte efferocytosis of oxidized RBCs is, at least in part, mediated by scavenging receptors such as ASGPR1. Of note, pretreatment of RBCs with ethanol but also heme and bilirubin also initiated efferocytosis. In a cohort of heavy human drinkers, a significant correlation of hepatic ASGPR1 with the heme degradation pathway was observed.
Conclusion: We here demonstrate that hepatocytes can directly ingest and degrade oxidized RBCs through efferocytosis, a process that can be also triggered by ethanol, heme and bilirubin. Our findings are highly suggestive for a novel mechanism of hepatic iron overload in ALD patients.
Keywords: alcohol-related liver disease, ASGPR1, efferocytosis, heme oxygenase-1, red blood cells, scavenging receptors, iron metabolism
Introduction
Too much alcohol intake causes ca. 50% of cases of liver cirrhosis worldwide and is considered the leading etiology of advanced liver disease in the coming decades.1,2 Alcohol-related liver disease (ALD) is also the main contributor to a loss of disability-adjusted life years among adults who abuse alcohol among adults who abuse alcohol in Europe. Despite intensive research efforts for over five decades, the underlying molecular mechanisms of the disease are still poorly understood. This also applies to typical laboratory findings in heavy drinkers.2 For instance, besides elevated levels of gamma-glutamyl transferase (GGT) and serum ferritin, the mean corpuscular volume (MCV) of red blood cells is often increased.3 In addition, in liver histology, about 50% of drinkers show histological accumulation of hepatic iron, both in hepatocytes and macrophages.4–6
First preliminary data of a longterm prospective study on mortality in heavy drinkers has recently identified anemia as major long-term predictor of death in this cohort.6,7 Further analysis revealed that the anemia is associated with signs of enhanced hemolysis, ineffective erythropoiesis and an enhanced RBC turnover. Of note, deficiency of vitamin B12 or folic acid as classical causes of ineffective erythropoiesis could not be confirmed. These novel clinical findings have put on stage the enhanced turnover of senescent or impaired RBCs by phagocytic cells, a process termed erythrophagocytosis.
In the last 30 years, a profound progress has been made in order to better understand the molecular mechanisms of iron regulation both at the systemic and cellular level.8,9 Most iron (20–25 mg) is daily recycled by macrophages, which phagocytize senescent RBCs mainly in the spleen and liver through the reticuloendothelial system. For new production of red blood cells, macrophages also directly recycle iron in the bone marrow. Hepatocytes and macrophage ferritin also serve as a storage side for excess iron. The mean survival time of RBCs typically lasts 120 days.10
Besides erythrophagocytosis, free hemoglobin can be either bound to haptoglobin or, after release of toxic heme, hemopexin. It is then internalized through a hemoglobin-haptoglobin complex (CD163) or a heme-hemopexin complex (CD91).11,12 During erythrophagocytosis, heme oxygenases (HO), mainly inducible HO-1, heme is enzymatically degraded into equimolar amounts of carbon monoxide (CO), biliverdin and iron.13–16 In a coupled reaction, biliverdin is further converted into bilirubin via biliverdin reductase.17 While HO-2 is constitutively expressed and mainly found in brain and testis,18 inducible isozyme HO-1 is expressed at low levels in most cells and tissues.15 However, HO-1 can be markedly up-regulated by its substrate heme and a variety of oxidants.19,20 Consequently, HO-1 induction has been considered as an adaptive response to protect against oxidative stress.21–24 Of note, Nrf2, a major inducer of HO-1 coordinates the transcription of various pathways of the hepatic elimination and detoxification phases 0–3.25 Although macrophages, within the context of ALD pathology, inflammation and bacterial endotoxins, have been intensively studied,26 the role of hepatocytes in recycling RBCs has received almost no attention.
In this work, we show that hepatocytes can directly ingest and degrade red blood cells through efferocytosis and that this process can also be initiated by ethanol exposure to RBCs. Our findings are likely to represent a novel mechanism for RBC recycling and could lead to a new explanation for hepatic iron overload in ALD.
Materials and Methods
Cell Culture
Human hepatoma Huh7 cells were cultured under standard conditions in Dulbecco’s modified Eagle medium (DMEM, Sigma-Aldrich, Taufkirchen, Germany) with 25 mM glucose and 10% fetal calf serum (FCS). These cells were obtained from the Japanese Cancer Research Resources Bank (JCRB, Tokyo, Japan). Murine primary hepatocytes, generously provided by Dr. Yujia Li (University of Heidelberg, Germany), were grown under standard conditions using Williams’ medium (Sigma-Aldrich, Taufkirchen, Germany). The isolation of murine primary hepatocytes followed a two-step collagenase (C2-22, Merck Biochrom) perfusion method as described previously.27,28 Freshly prepared hepatocytes were seeded and harvested within 48 h after isolation. Each experiment involving cells was repeated two or three times, with at least triplicate samples for each repetition. THP-1 immortalized human monocytes were bought from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum and 25 mM glucose (Gibco, Thermo Fisher Scientific, Waltham, MA, USA).
Co-Culture Systems
Direct co-culture system of THP-1, Huh7 and RBCs together was established by seeding THP-1 monocytes for differentiation with PMA at 0.3×105 cells/well in 12-well plates for a ratio hepatocyte to macrophage of 4:1.29,30 After 24 hours of differentiation, Huh7 cells were cultured in 12 well plates using a density of 1.3×105 cells/well and incubated over night for attachment. Then, RBCs were added. Similarly, Huh7 cells and RBCs were also kept in co-culture by seeding Huh7 cells 1.3x105/well first in wells and subsequently RBCs were added to the wells after 24 hours. THP-1 and RBCs were also kept in co-culture by seeding THP-1 for differentiation with PMA at 0.3x105/well first, then RBCs were added to the wells after 24 hours.
Chemicals and Reagents
Phorbol 12-myristate 13-acetate (PMA, 100 ng/mL), CuSO4 (0.2mM), ascorbate (5mM) and staurosporine (1um) were purchased from Sigma-Aldrich (Taufkirchen, Germany).
Live Video Camera System
Real-time videos of the uptake of RBCs by hepatocytes were taken using the IncuCyte®S3 system (Essen BioScience, Royston, UK; Cat. no.: 4647). The live-cell imaging platform facilitates the immediate acquisition of real-time well-wide cell images and videos. Images were taken with a 2-minute time-interval which was the lowest time-resolution for the device.
SiRNA Transfection
ASGPR1 siRNA transfection assay was performed on Huh7 cells. Huh7 cells were seeded as 5×104 in 12-well plates. The density of cells per well was around 80 to 90%. Transfection was performed under a reduced serum medium (Thermo Fisher; 31985062) with 50 nM ASGPR1 siRNA (Thermo Fisher; AM16708) or 50 nM nontargeting siRNA (Thermo Fisher; SIC001) as a negative control by Lipofectamine 2000 (Invitrogen; 13778030). Lipofectamine was applied as 3 μL per well. Transfection reagents were all removed after 6 h, and the transfection efficiency was detected at 48 h. After transfection for 24 h, the general density of cells in 12-well plates was around 50 to 60%. The knockdown efficiency was tested by real-time quantitative PCR (qRT-PCR).
qRT-PCR and ELISA
Total RNA from cells was extracted using Trizol Reagent (Thermo Fisher, 15596018) following the standard protocol provided by the supplier. Reverse transcription was performed as previously reported.31 Primers and specialized probes were synthesized at Eurofins MWG Operon (Ebersbach, Germany). For q-PCR using probes, the probes and primers were designed using the universal Probe Library (LiveScience, Roche Molecular Systems), and the UPL probes were also obtained from Roche. The sequences of each primer are presented in Supplementary. Table 1. A direct ELISA assay was established using a human ASGPR1 Elisa kit (Biorbyt Company, orb562782). ELISA was performed following the manufacturer’s instructions.
Patients
We analyzed frozen sera from patients who were enrolled from 2011 until 2020 at Salem Medical Center Heidelberg during an ongoing study.7 All patients were heavy drinkers (>80 g per day in males and >60 g per day in females) with ALD and a mean alcohol consumption of 187 ± 124 g/day. The study protocol was reviewed and approved by the Ethics Committee of the University of Heidelberg (Ethikkommission der medizinischen Fakultät Heidelberg, S150-15) and conformed to the ethical guidelines of the 1975 Declaration of Helsinki and all patients gave written informed consent prior to inclusion. In n=47 patients with sera before and after 7.0 ± 2.4 days of alcohol detoxification, serum levels of ASGPR1 were determined by ELISA. In n=30 patients with liver biopsy, mRNA expression levels of ASGPR1 and HO-1 were measured by q-PCR in liver tissue. Additionally, all patients had routine laboratory and liver stiffness measurements (Fibroscan, Echosens SA, Paris, France). Patient characteristics are shown in Supplementary Table 2.
Statistical Analysis
All data are shown as mean ± standard deviation. Significant differences (n.s., no significance; *p<0.05; **p<0.01; ***p<0.001) between means of data sets were assessed by one-way ANOVA with Tukey’s test or unpaired two-tailed Student’s t-test using GraphPad Prism 9 software. Correlations were conducted using the Spearman Rho correlation coefficient and comparison of changes in paired data was done using the paired samples t-test.
Results
Direct Indication for RBC Ingestion (Efferocytosis) by Hepatocytes
Since macrophages and hepatocytes closely cooperate in the liver to handle recycled RBCs and iron, we set up in vitro experiments in order to learn more about the modulation of macrophage erythrophagocytosis by co-cultured hepatocytes. To study the effect of co-cultured hepatocytes on macrophages, we differentiated THP1 monocytes with PMA to macrophages (here called THP1 macrophages) and exposed them to intact but oxidized human RBCs for 24 hours (for experimental design see Supplementary Figure 1A). Oxidized or aged RBCs were produced by exposing them to copper sulfate for 120 min. We used THP1 cells alone or in combination with Huh7 cells and Huh7 cells alone. We also used the physiological 4:1 ratio for hepatocytes and THP1 cells.29 HO-1 mRNA was assessed by RT-PCR as a parameter of erythrophagocytosis. As shown in Figure 1A, oxidized RBCs significantly induced HO-1 mRNA by a factor of 7, while non-oxidized RBCs caused only a minor HO-1 mRNA elevation. Seemingly no difference was observed in THP1 cells co-cultured together with Huh7 cells. However, unexpectedly, Huh7 cells alone also showed a strong HO-1 induction when exposed to oxidized RBCs. When performing light microscopy over 24 hours (see Figure 1B), some hepatocytes had ingested multiple RBCs comparable to macrophages during erythrophagocytosis. In some hepatocytes, RBCs were aligned around the cell nucleus which confirmed that these RBCs were indeed ingested and not aligned around the hepatocytes or simply attached to them. We next proofed that RBCs are directly taken up by hepatocytes using a live video camera system to record co-culture of hepatocytes and RBCs over 24 hours. The video is available for download in Video S1 (covering ca. 120 min and starting after 5 hours of a 24 hour recorded experiment). Figure 1C shows representative images for 6 time points within this 120 min sequence. Red arrows indicate RBC efferocytosis. As the term phagocytosis is restricted to professional phagocytes, we will use the term efferocytosis which describes the ingestion of cells by eg epithelial cells. With the limited time resolution of the video system (2 min), our data suggest that RBCs are taken up very rapidly by hepatocytes, approximately within 10–15 minutes. RBCs are ingested once getting into direct contact with hepatocytes. We finally confirmed that RBC ingestion or efferocytosis can be also observed in primary cells. When oxidized murine RBCs were exposed to primary mouse hepatocytes, they also efficiently performed efferocytosis (see Figure 2A and B). In summary, we here show for the first time that hepatocytes are able to rapidly ingest aged or oxidized RBCs, a process generally termed efferocytosis. RBC efferocytosis by hepatocytes is a rapid process and terminated within minutes.
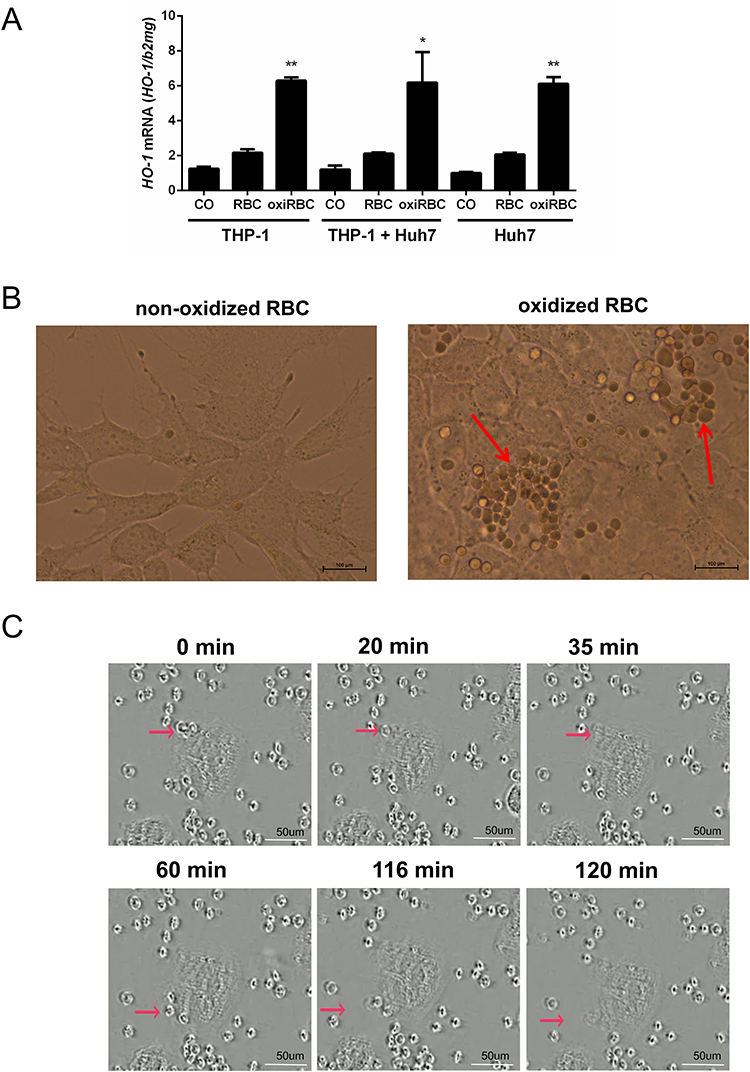 |
Figure 1 Evidence of direct RBC efferocytosis by hepatocytes. (A) Comparable to macrophages (THP-1), hepatocytes also show high induction of HO-1 mRNA expression when exposed to oxidized red blood cells (oxiRBC). THP1 macrophages and Huh7 cells were cultured either alone or co-cultured together and exposed to oxidized RBCs. HO-1 mRNA was measured by RT PCR (n=2 per group). Further conditions: RBCs were used at a concentration of 1% hematocrit. Huh7 to THP-1 cell ratio was 4:1 reflecting the physiological ratio.29 Results are represented as mean of mRNA levels normalized to β2- microglobulin ± SD. Tukey’s multiple comparisons test, *, p<0.05; **, p<0.01. (B) Light microscopy of direct RBCs efferocytosis by hepatocytes. Co-culture of Huh7 cells with oxidized RBC (hematocrit 1%) and light microscopy at 24 hours after co-culture. Note, that multiple RBCs are surrounding the nucleus confirming complete internalization of RBCs (red arrows). Scale bars, 100 µm. (C) Rapid ingestion of RBCs (efferocytosis) by hepatocytes (Video screen shot). Hepatocytes (Huh7 cells) were co-cultured with 0.01% oxidized RBC and then followed up by light microscopy using a video camera system over 24 hours. Representative images are shown after 5 hours (set at time point 0 min for a time period of 120 min). Scale bars, 50 µm. Red arrows indicate examples of RBC efferocytosis by hepatocytes. The complete video with 2 min-intervals is available in Video S1. |
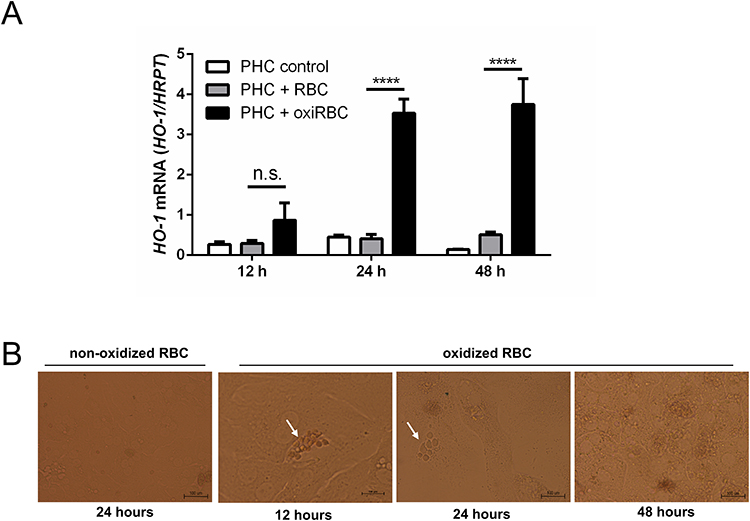 |
Figure 2 RBC efferocytosis was also detected in murine primary hepatocytes (PHC) co-cultured with murine oxidized RBCs (oxiRBC). RBCs (1% hematocrit) from mice were oxidized by CuSO4 for 2 hours and then co-cultured with murine primary hepatocyte for different time points. At least >80% of all hepatocytes showed some efferocytosis of RBCs. (A) After treatment of human primary hepatocytes (PHC) with 1% oxidized RBCs, HO-1 mRNA is significantly increased at all time points. HO-1 mRNA was quantified by quantitative real-time PCR (n=3 per group) and the results are represented as mean of mRNA levels normalized to HRPT± SD. Tukey’s multiple comparisons test, n.s., no significance; ****, p<0.0001. (B) Representative image of RBCs efferocytosis by primary murine hepatocytes at different time points after co-culture. Scale bars, 100 µm. |
Kinetic Studies on RBCs Efferocytosis by Hepatocytes
We next studied in more detail the dose and time dependence of RBCs efferocytosis by hepatocytes. Of note, efferocytosis started at much lower RBC counts as compared to professional phagocytes (Zheng C. et al, unpublished data). A first peak of HO-1 mRNA was already observed at a hematocrit of 0.05% which is 20 times less as compared to macrophages (see Figure 3A). With high reproducibly, a second peak was seen at 1% hematocrit while HO-1 mRNA expression decreased thereafter. Of note, ferritin mRNA, the major iron storage protein, only peaked at the hematocrit of 0.05%. Time-dependent studies confirmed that efferocytosis had started before 2 hours (see Supplementary Figure 2A and 2B). In conclusion, efferocytosis of RBCs by hepatocytes starts at much lower cell counts and is self-limited at higher RBCs counts.
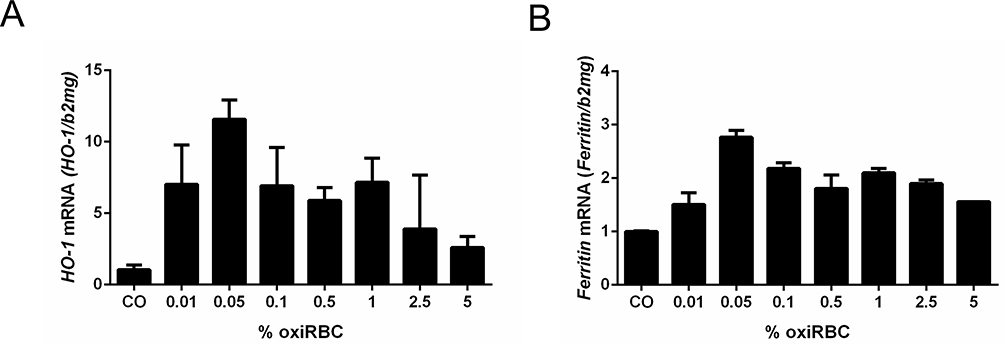 |
Figure 3 Induction of (A) HO-1 mRNA and (B) Ferritin mRNA during RBCs efferocytosis of hepatocytes. RBCs were oxidized by CuSO4 for 2 hours and then co-cultured with Huh7 cells. Note that two peaks can be reproducibly induced at 0.05% and 1% hematocrit. HO-1 and ferritin mRNA was measured by quantitative real-time PCR and the results are represented as mean of mRNA levels normalized to β2-microglobulin ± SD (n=2). |
Potential Mechanisms of RBCs Efferocytosis by Hepatocytes
We first studied the expression of Nrf2 (Nuclear factor erythroid 2-related factor 2), the major upregulator of HO-1 that also orchestrates the transcriptional induction of various enzymes of the hepatic elimination and detoxification phases 0–3.25 Nrf2 was also induced in response to RBCs efferocytosis mirroring the response of HO-1 and peaking at 0.05% hematocrit (see Figure 4A). This confirms that Nrf2 is involved in the efferocytosis response most likely upon the release of heme. Since RBCs are known to have important surface glycoproteins, we next screened for potential hepatocyte receptors that may be involved in RBCs efferocytosis. Figure 4B shows the response to oxidized RBCs (0.05% hematocrit) of Scavenger Receptor Class F Member 1 (SCARF1), ASGPR1 and 2, and TLR4. All scavenger receptors except SCARF1 were induced during efferocytosis. Moreover, as is exemplarily shown in Figure 4C for ASGPR1, it peaked at rather low 0.05% hematocrit and decreased at higher concentrations without having a second peak such as HO-1. Unfortunately, we failed to directly proof whether these receptors are directly involved in efferocytosis. Thus, knock down experiments using eg ASGPR1 siRNA that was able to efficiently knock down ASPGR1 mRNA by 65% had no effect on efferocytosis. However, we believe that this is due to a high sensitivity of phagocytic processes such as efferocytosis to membrane interferences that necessarily occur during treatment with transfection agents. Of note, efferocytosis could be strongly blocked by the unselective kinase inhibitor staurosporine and staurosporine also inhibited significantly the induction of ASGPR1 (see Figure 4D and E). In addition, protein levels of ASGPR1 mirrored mRNA expression levels (see Supplementary Figure 3). Taken together, our first mechanistic studies indicate that RBC efferocytosis by HC involves the important transcription factor Nrf2, is kinase dependent and HO-1 expression is mirrored by important but multiple-scavenging receptors that include ASGPR1.
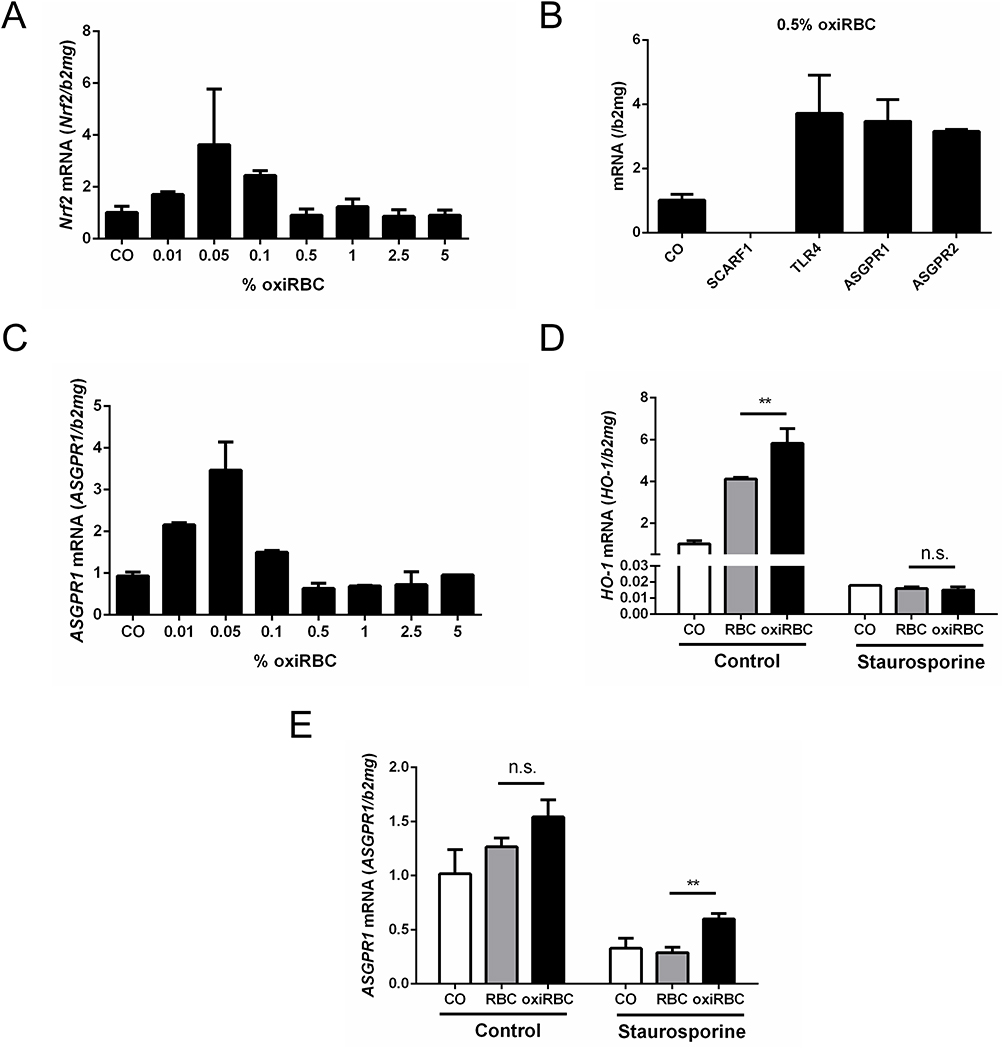 |
Figure 4 Expression of Nrf2 and scavenging receptors during RBCs efferocytosis by hepatocytes. RBCs were oxidized by CuSO4 for 2 hours and then co-cultured with hepatocytes. (A) HO-1 upstream transcription factor Nrf2 is also induced peaking at 0.05% hematocrit (n=2). (B) Screening of various scavenger receptors 24 hour after exposure of Huh7 cells to 0.05% oxidized RBCs. ASGPR1and TLFR4 showed the highest expression (n=2). (C) ASGPR1 mRNA showed a dose–response curve peaking at 0.05% oxiRBC. mRNA was quantified by quantitative real-time PCR and the results are represented as mean of mRNA levels normalized to β2- microglobulin ± SD (n=2). E-G) Inhibition of RBCs efferocytosis by hepatocytes using the kinase inhibitor staurosporine. Pre-treated Huh7 with staurosporine (1uM/well) for 1 hour. After refreshing the medium, Huh7 cells were co-cultured with RBCs/oxiRBC (0.05%) for 24 hours. (D) HO-1 mRNA expression was significantly decreased in the staurosporine treated group by 70%. Tukey’s multiple comparisons test, n.s., no significance; **, p<0.01. (E) Expression of ASGPR1 was significantly decreased by staurosporine. HO-1, ASGPR1 and mRNA was quantified by quantitative real-time PCR and the results are represented as mean of mRNA levels normalized to β2-microglobulin ± SD. Tukey’s multiple comparisons test, n.s., no significance; **, p<0.01 (n=3). |
Ethanol Primes RBCs for Hepatocyte-Mediated Efferocytosis
Similar to our erythrophagocytosis studies (Zheng C. et al Evidence for alcohol-mediated hemolysis and erythrophagocytosis. unpublished 2024), we next set up an in vitro model (see also Supplementary Figure 1) to study in more detail ethanol-mediated aspects of efferocytosis using HO-1 mRNA expression as parameter of RBCs ingestion. Huh7 cells were cocultured with human RBCs that had been exposed of 24 hours to increasing concentrations of ethanol. Oxidized RBCs using copper sulfate served as positive control, while untreated RBCs were used as negative control. As shown in Figure 5A, ethanol concentrations as low as 400 mM started to efferocytosis. These are lower concentrations as compared to priming in macrophages (800 mM) or direct lysis of RBCs by ethanol (2000 mM). At 1400 mM ethanol, HO-1 expression reached the levels of the positive control. Of note, the process is independent on the duration of alcohol exposure and short exposure times of 30 min suffice to cause efferocytosis suggesting most likely a direct interaction of ethanol with the RBCs membrane. In summary, these data demonstrate that ethanol is able to prime RBCs for efferocytosis by hepatocytes, most likely due to direct membrane modification.
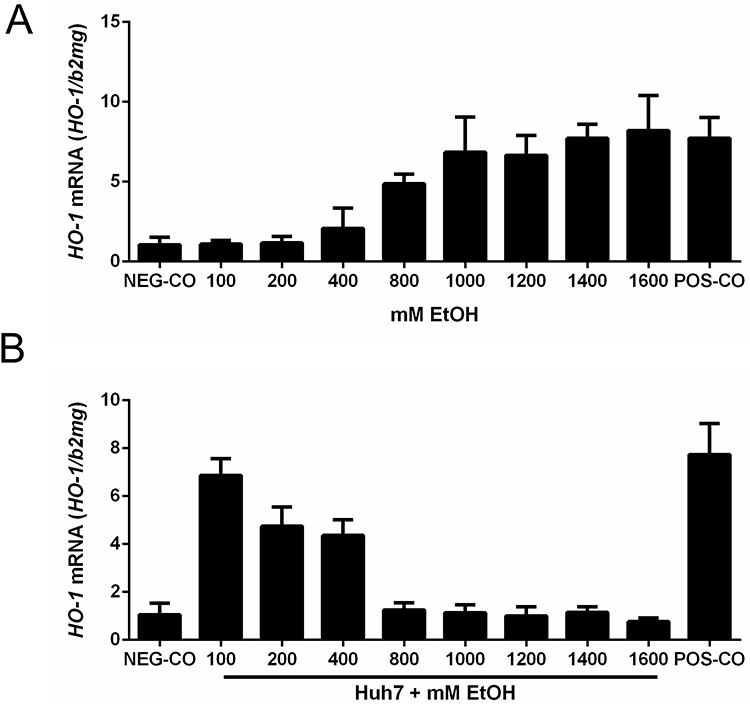 |
Figure 5 Ethanol primes RBCs for hepatocyte-mediated efferocytosis. (A) After treating RBCs with 100–1600 mM ethanol for 24 hours, the expression of HO-1 was measured after co-culture with Huh7 cells at 1% hematocrit for 24 h. At concentrations of ethanol ≥800 mM, efferocytosis was observed as measured by increasing HO-1 mRNA levels. (B) High levels of ethanol efficiently block efferocytosis. RBCs were oxidized by CuSO4 for 2 hours, washed and then co-cultured with Huh7 in the presence of increasing ethanol concentrations (100–1600 mM) for 24 hours. The expression of HO-1 was significantly reduced at ethanol levels ≥800 mM. HO-1 was quantified by quantitative real-time PCR and the results are represented as mean of mRNA levels normalized to β2-microglobulin ± SD (n=2). Oxidized RBCs served as positive control (POS-CO). |
Ethanol Reversibly Blocks RBCs Efferocytosis by Hepatocytes
The in vitro efferocytosis system also allowed us to study potential-inhibiting effects of alcohol on the process of RBCs ingestion. RBCs were oxidized by copper sulfate for two hours and then co-cultured with Huh7 and co-exposed with 100–1600 mM ethanol (0.5–10%) for 24 hours. A significant inhibition of efferocytosis and HO-1 expression was already observed at 200 mM ethanol, a complete inhibition at 800 mM (see Figure 5B). In contrast to macrophages, hepatocytes did not recover at least after 24 hours upon exposure to ethanol (see Supplementary Figure 4). We finally exposed a co-culture model of macrophages (THP1) and hepatocytes (Huh7 cells) to human RBCs pretreated with increasing ethanol concentrations (Supplementary Figure 5). We also studied in this co-culture model the inhibition of efferocytosis (using oxidized RBCs) in the presence of ethanol (Supplementary Figure 6). These data show that both cells together are more resistant to ethanol and RBCs are primed for ingestion at lower ethanol levels.
In conclusion, ethanol efficiently blocks RBCs efferocytosis by hepatocytes. In contrast to macrophages, this inhibition seems to be irreversible for at last 24 hours.
Bilirubin and Hemin Also Prime RBCs for Efferocytosis
As elevated bilirubin and heme are toxic end products of hemolysis and end-stage liver disease and accumulate at higher fibrosis stages, we also set up experiments to learn whether they are able to trigger efferocytosis of control RBCs. As shown in Figure 6, human control RBCs were incubated for 3 hours with different concentrations of bilirubin and hemin and then pretreated RBCs were co-incubated with Huh7 for another 24 hours. As can be seen in Figure 6, higher concentrations caused a significant increase of HO-1 mRNA indicating enhanced efferocytosis. In summary, typical toxic end products of hemolysis such as bilirubin or hemin are able to further enhance efferocytosis potentially allowing to initiate a vicious cycle.
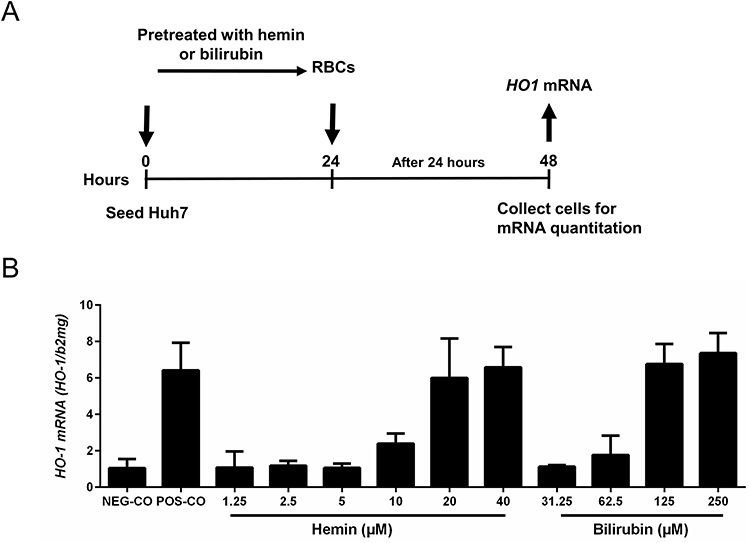 |
Figure 6 Heme-degradation end-products heme and bilirubin also prime RBCs for efferocytosis. (A) Experimental design: RBCs were exposed to increasing concentrations of hemin or bilirubin for 3 hours. RBCs treated with CuSO4 for 2 hours served as positive control, untreated RBCs as negative control. RBCs were then co-cultured with Huh7 for 24 hours and HO-1 mRNA was determined to monitor efferocytosis. (B) Heme and bilirubin prime RBCs for efferocytosis starting at concentrations of ca. 10 and 60 µM, respectively. HO-1 mRNA was quantified by quantitative real-time PCR and the results are represented as mean of mRNA levels normalized to β2- microglobulin ± SD (n=2). Oxidaized RBCs served as positive control (POS-CO). |
Hepatic mRNA Expression of ASGPR1 in Heavy Drinkers
Based on the Heidelberg biobank of heavy drinkers, we were able to measure ASGPR1 mRNA mRNA in liver specimen form n=30 patients and perform a Spearman Rho correlation analysis. Patient characteristics are shown in Supplementary Table 2. Spearman Rho correlations for ASGPR1 are shown in Supplementary Table 3A. ASGPR1 is correlated with mRNA fingerprints from the heme degradation pathways despite the rather small sample size of n=30 frozen liver specimen. Thus, ASGPR1 mRNA showed a good positive correlation with liver HO-1 mRNA (r=0.47, p=0.02) and the HO-1 upstream regulator Nrf2 (r=0.39, p=0.06).
Serum Levels of ASGPR1 Decrease After Alcohol Detoxification and Highly Correlate with Mortality and Hemolysis
Finally, in serum samples from 47 heavy drinkers, we were able to assess serum levels of ASGPR1 prior and after alcohol detoxification (Supplementary Figure 7). ASGPR1 serum levels significantly decrease after a mean detoxification period of 7.0 ± 2.4 days 182.0 ± 8.7 ng/mL to 169.0 ± 6.6 ng/mL, p<0.0001. In addition, serum ASPGR1 levels were highly correlated with serum CD163 levels and other markers of hemolysis and, most importantly, with all-cause death (Supplementary Table 4). In conclusion, first mRNA and serum data from a heavy drinker cohort indicate that ASGPR1 is tightly associated with markers of heme degradation. Moreover, serum ASGPR1 levels are associated with markers of hemolysis and rapidly decrease after alcohol withdrawal. In conclusion, serum ASGPR1 levels are highly associated with markers of hemolysis and all-cause mortality. They decrease after alcohol withdrawal.
Discussion
We here show, to our knowledge for the first time, that hepatocytes are able to directly ingest aged or oxidized RBCs, a process termed efferocytosis. RBC efferocytosis induces the heme degradation pathway including Nrf2 and HO-1 but also iron storage protein ferritin. Second, we further show that ethanol is able to prime RBCs for efferocytosis. Third, in contrast, ethanol can also efficiently inhibit efferocytosis when present during efferocytosis at higher concentrations. We believe that these observations have fundamental implications not only for a better understanding of alcohol-related disease mechanisms such as iron overload or cholestasis but also for physiological RBC recycling in general.
Our observation was made accidentally when learning that hepatocytes respond with HO-1 expression similar to phagocytes in the presence of oxidized RBCs. While, in professional phagocytes, this process is termed erythrophagocytosis,12 ingestion of foreign cells or apoptotic bodies by epithelial cells is termed efferocytosis.32,33 Live video recording confirmed the direct uptake of RBCs by hepatocytes. Moreover, we could recapitulate RBC efferocytosis with primary hepatocytes from mice. The rapid ingestion within 10 minutes indicates a rather fast, membrane and receptor orchestrated process. This short time may be also the major reason why it has not been described so far. In fact, there has been only a descriptive mentioning of intrahepatocellular RBCs without further analysis and these observations have not been published in regular peer-reviewed journals.34 In addition, we were able to identify various reports published more than 70 years ago describing intracellular RBCs in hepatocytes mostly in association with various poisons.35
On the other side, the clearance of apoptotic cells by hepatocytes through efferocytosis has been well documented.32 Although RBCs lack mitochondria, they can undergo a programmed cell death (apoptosis), sometimes termed as eryptosis.36 At least the phenotypical characterization (stinging) of oxidized RBCs matches the definition of spur cells or acanthocytes suggesting that both ascorbate/CuSO4 and ethanol-treated RBCs may eventually undergo eryptosis.36 Apoptotic cells are often recognized through the display of phospholipid phosphatidylserine (PtdSer) on the outer leaf of the plasma membrane. Consequently, receptors such as Stabilin-1, Stabilin-2, Brain-specific angiogenesis inhibitor 1 (BAI1), and RAGE but also scavenging receptors such as SCARF1 are thought to be involved in efferocytosis.32,33 In our setting ASGPR1/2 and TLR4 (but not SCARF1) were induced during hepatocyte efferocytosis of RBCs. Unfortunately, we were not able to further dissect the receptors specifically involved by knock down experiments most likely due to membrane interactions of transfection reagents. In contrast, RBCs ingestion was almost completely blocked by phosphokinase inhibitor staurosporine strongly suggesting the involvement of kinase action as reported earlier in scavenging receptor mediated studies.37,38 However, it will remain an interesting field for further exploration to better understand the role of ASGPR1 in red blood cell elimination. Already early studies showed rapid clearance of desialylated erythrocytes with sequestration by the liver and suggested a reduction in erythrocyte sialic acid as a mechanism of erythrocyte destruction and an involvement in erythrocyte senescence.39,40 By this time it also remained unresolved, how hepatocyte ASGPR would gain access to red blood cells since they are separated by liver sinusoidal endothelial cells.41
What is the potential meaning of RBC efferocytosis for liver pathophysiology? The fact that hepatocytes are able to directly ingest RBCs has several implications. Altogether about 90% of iron is recycled from senescent erythrocytes that typically have a mean survival of 120 days.10 During erythrophagocytosis, heme oxygenase (HO) catalyzes the enzymatic degradation of heme and produces equimolar amounts of carbon monoxide (CO), biliverdin and iron.13–16 Of note, hepatocytes are the almost exclusive side of bilirubin excretion and there is no reversal of bilirubin towards heme such as a reversed synthesis. Thus, RBC efferocytosis by hepatocytes would offer a direct recycling of RBCs material in the cell that is the only location of bilirubin excretion and major cross section for iron regulation, storage and control. In support, we could confirm the activation of the heme degradation and iron storage pathways by demonstrating a robust upregulation of Nrf2, HO-1 and ferritin mRNA during RBCs efferocytosis. Since the redox-sensitive transcription factor Nrf2 also coordinates the transcription of many enzymes of the hepatic elimination and detoxification phases 0–3,25 our findings are likely to have broader implications that also include the metabolism of lipids and bile acids. So, besides HO-1, these target genes also include glucuronidation of bilirubin by UDP glucuronosyltransferase, glutathione S transferase (GST) or multidrug resistance-associated proteins (MRPs). Second, RBCs efferocytosis would also have fundamental implication for iron homeostasis. The hepatocyte is not only a major regulator of systemic iron homeostasis by secreting the peptide hormone hepcidin but also an important storage side. RBCs efferocytosis could represent a highly efficient and direct iron recycling mechanisms that would shortcut eg the release of iron from macrophages to transferrin and secondary uptake by transferrin receptors.
However, many open questions remain and should be addressed in future studies: Why are internalized RBCs not seen more often in histological sections? How do RBCs get direct access to hepatocytes which are considered to be separated by a fenestrated endothelial layer within the liver sinus? Are there more specific receptor mechanisms for RBCs efferocytosis than the one suggested in this manuscript? Our clinical data suggest that during hemolytic, eryptotic or erythrophagocytotic processes, several mechanisms occur in parallel such as hemoglobin scavenging by haptoglobin, direct heme trapping by hemopexin, erythrophagocytosis and, eventually, efferocytosis. It also remains to be understood why hepatocytes already respond to much smaller numbers of RBCs as compared to macrophages, almost 20 times lower. First data from our human cohort of heavy drinkers confirm that both RBCs efferocytosis and hemolysis/erythrophagocytosis are highly associated with mortality but also serum markers to be elevated in ALD patients such as uric acid or cytokines namely TNF alpha, IL8 and IL6. Finally, it will be of high interest to put RBCs efferocytosis into context with hepatocellular death, energy and oxygen metabolism since the uncontrolled release of heme degradation end-products heme, bilirubin, iron and carbon monoxide are highly reactive and very likely to be engaged in apoptosis and ferroptosis.42
Our novel data on the modulation of efferocytosis by ethanol also deserve discussion.
First, five times lower levels of ethanol are able to prime RBCs for efferocytosis as compared to direct lysis. It is quite conceivable that, locally, these concentrations may occur during alcohol consumptions and especially during consumption of high percentage beverages. Such high concentrations could occur in the upper digestive tract or the portal vein and continuously and steadily prime RBCs for enhanced recycling through erythrophagocytosis or efferocytosis, as described here. On the other side, we have also demonstrated that ethanol is able to block RBCs efferocytosis by hepatocytes. Thus, it is possible that a complex balance of RBCs priming and modulation of efferocytosis by ethanol will result in various individual responses in heavy drinkers. Of note, potentially involved scavenging receptor ASGPR1 was tightly associated with the heme degradation pathways (Nrf2 and HO-1 mRNA) in human heavy drinkers and, ASGPR1, significantly decreased after one week of alcohol detoxification.
Of special clinical interest could be our observations that heme-degradation products such as heme and bilirubin are, in addition to ethanol, also able to further prime RBCs for efferocytosis at rather low levels starting from 10 to 60 µM, respectively. Consequently, this finding is highly suggestive for a potential vicious cycle. If hemolysis exceeds certain thresholds, bilirubin accumulation could perturbate priming of RBCs for efferocytosis. Within this context, it is quite interesting to note that such bilirubin levels are typically measured in ALD patients with manifest liver cirrhosis.6 Since lipids such as cholesterol are an important compartment of RBCs,43,44 RBCs efferocytosis may also contribute to hepatocyte fat loading (steatosis) and, consequently, affect or modulate bile acid synthesis and bile formation.
Conclusion
In conclusion, we here show that hepatocytes are able to directly ingest RBCs that are aged/oxidized or primed by ethanol. We think that, together with erythrophagocytosis, this novel process is likely to contribute to enhanced RBCs turnover and hemolytic anemia, identified in drinkers as most important prognostic factor for long-term mortality. It is expected that a better understanding of RBCs efferocytosis and its control will help to develop novel targeted therapies in diseases such as ALD.
Abbreviations
ALD, alcohol-related liver disease; ASGPR1, asialoglycoprotein receptor 1; b2mg, Beta; 2-microglobulin; GGT, gamma-glutamyltransferase; HO-1, heme oxygenase 1; HTC, hematocrit; MCV, mean corpuscular volume of erythrocytes; Nrf2, Nuclear factor erythroid 2-related factor 2; oxiRBC, oxidized RBC; RBC, red blood cell.
Acknowledgments
This study was funded by the Deutsche Forschungsgemeinschaft (DFG) to S.M. (RA 2677/1-2) and The Federal Ministry of Education and Research-Liver Systems Medicine Program LiSyM-Cancer, Grant PTJ-031L0257A, and by the Stiftung für Biomedizinische Alkoholforschung. C.Z. and S.L. C.C. are also grateful for the financial support of China Scholarship Council (CSC). Further support came in part by “The 111 Project” (D17011). For the publication fee, we acknowledge financial support by Heidelberg University.
Disclosure
Dr Seddik Hammad reports grants from The Stiftung für Biomedizinische Alkoholforschung, the BMBF (German Federal Ministry of Education and Research) Project LiSyM (Grant PTJ-FKZ: 031L0043) and LiSyM-Cancer (Grant PTJ-FKZ: 031L0257A), outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. World Health Organization. Global status report on alcohol and health; 2018. Available from: http://www.who.int/substance_abuse/publications/global_alcohol_report/en/.
2. Mueller S, Heilig M. Alcohol and Alcohol-Related Diseases. Springer International Publishing; 2023.
3. Mueller S, Seitz HK, Rausch V. Non-invasive diagnosis of alcoholic liver disease. World J Gastroenterol. 2014;20(40):14626–14641. doi:10.3748/wjg.v20.i40.14626
4. Mueller S, Rausch V. The role of iron in alcohol-mediated hepatocarcinogenesis. Adv Exp Med Biol. 2015;815:89–112.
5. Silva I, Rausch V, Seitz HK, Mueller S. Does hypoxia cause carcinogenic iron accumulation in alcoholic liver disease (ALD)? Cancers. 2017;9(11):145. doi:10.3390/cancers9110145
6. Mueller S, Chen C, Mueller J, Wang S. Novel insights into alcoholic liver disease: Iron overload, iron sensing and hemolysis. J Transl Int Med. 2022;10(2):92–124. doi:10.2478/jtim-2021-0056
7. Mueller S, Mueller J. Alcohol and mortality: First preliminary lessons from a prospective 15 year follow-up study. In: Mueller S, Heilig M, editors. Alcohol and Alcohol-Related Diseases. Springer International Publishing Cham; 2023:81–102.
8. Ganz T. Systemic iron homeostasis. Physiol Rev. 2013;93(4):1721–1741. doi:10.1152/physrev.00008.2013
9. Wang J, Pantopoulos K. Regulation of cellular iron metabolism. Biochem J. 2011;434(3):365–381. doi:10.1042/BJ20101825
10. Andrews NC. Disorders of iron metabolism. N Engl J Med. 1999;341(26):1986–1995. doi:10.1056/NEJM199912233412607
11. Winn NC, Volk KM, Hasty AH. Regulation of tissue iron homeostasis: The macrophage ”ferrostat”. JCI Insight. 2020;5(2). doi:10.1172/jci.insight.132964
12. Klei TR, Meinderts SM, van den Berg TK, van Bruggen R. From the cradle to the grave: The role of macrophages in erythropoiesis and erythrophagocytosis. Front Immunol. 2017;8:73. doi:10.3389/fimmu.2017.00073
13. Tenhunen R, Marver HS, Schmid R. The enzymatic conversion of heme to bilirubin by microsomal heme oxygenase. Proc Natl Acad Sci U S A. 1968;61(2):748–755. doi:10.1073/pnas.61.2.748
14. Maines MD. The heme oxygenase system: A regulator of second messenger gases. Annu Rev Pharmacol Toxicol. 1997;37:517–554. doi:10.1146/annurev.pharmtox.37.1.517
15. Ryter SW, Alam J, Choi AM. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol Rev. 2006;86(2):583–650. doi:10.1152/physrev.00011.2005
16. Immenschuh S, Baumgart-Vogt E, Mueller S. Heme oxygenase-1 and iron in liver inflammation: A complex alliance. Curr Drug Targets. 2010;11(12):1541–1550. doi:10.2174/1389450111009011541
17. Kapitulnik J, Maines MD. Pleiotropic functions of biliverdin reductase: Cellular signaling and generation of cytoprotective and cytotoxic bilirubin. Trends Pharmacol Sci. 2009;30(3):129–137. doi:10.1016/j.tips.2008.12.003
18. Trakshel GM, Kutty RK, Maines MD. Purification and characterization of the major constitutive form of testicular heme oxygenase. The noninducible isoform. J Biol Chem. 1986;261(24):11131–11137. doi:10.1016/S0021-9258(18)67358-2
19. Choi AMK, Alam J. Heme oxygenase-1: Function, regulation, and implication of a novel stress-inducible protein in oxidant-induced lung injury. Am J Respir Cell Mol Biol. 1996;15:9–19. doi:10.1165/ajrcmb.15.1.8679227
20. Immenschuh S, Ramadori G. Gene regulation of heme oxygenase-1 as a therapeutic target. Biochem Pharmacol. 2000;60:1121–1128. doi:10.1016/S0006-2952(00)00443-3
21. Keyse SM, Applegate LA, Tromvoukis Y, Tyrrell RM. Oxidant stress leads to transcriptional activation of the human heme oxygenase gene in cultured skin fibroblasts. Mol Cell Biol. 1990;10(9):4967–4969. doi:10.1128/mcb.10.9.4967-4969.1990
22. Vile GF, Basu-Modak S, Waltner C, Tyrrell RM. Heme oxygenase 1 mediates an adaptive response to oxidative stress in human skin fibroblasts. Proc Natl Acad Sci U S A. 1994;91(7):2607–2610. doi:10.1073/pnas.91.7.2607
23. Otterbein LE, Choi AM. Heme oxygenase: Colors of defense against cellular stress. Am J Physiol Lung Cell Mol Physiol. 2000;279(6):L1029–1037. doi:10.1152/ajplung.2000.279.6.L1029
24. Gozzelino R, Jeney V, Soares MP. Mechanisms of cell protection by heme oxygenase-1. Annu Rev Pharmacol Toxicol. 2010;50:323–354. doi:10.1146/annurev.pharmtox.010909.105600
25. Lim PJ, Duarte TL, Arezes J, et al. Nrf2 controls iron homeostasis in haemochromatosis and thalassaemia via Bmp6 and hepcidin. Nat Metab. 2019;1(5):519–531. doi:10.1038/s42255-019-0063-6
26. Ju C, Mandrekar P. Macrophages and alcohol-related liver inflammation. Alcohol Res. 2015;37(2):251–262.
27. Hengstler JG, Utesch D, Steinberg P, et al. Cryopreserved primary hepatocytes as a constantly available in vitro model for the evaluation of human and animal drug metabolism and enzyme induction. Drug Metab Rev. 2000;32(1):81–118. doi:10.1081/DMR-100100564
28. Bissell DM. Primary hepatocyte culture: Substratum requirements and production of matrix components. Federation Proc. 1981;40(10):2469–2473.
29. Silva I, Peccerella T, Mueller S, Rausch V. IL-1 beta-mediated macrophage-hepatocyte crosstalk upregulates hepcidin under physiological low oxygen levels. Redox Biol. 2019;24:101209. doi:10.1016/j.redox.2019.101209
30. Silva I, Rausch V, Peccerella T, Millonig G, Mueller S. Independent and additive induction of hepcidin by hypoxia and H2O2: Evidence for NOX4-mediated iron signaling. Z Gastroenterol. 2016;54(12):1343–1404.
31. Millonig G, Ganzleben I, Peccerella T, et al. Sustained submicromolar H2O2 levels induce hepcidin via signal transducer and activator of transcription 3 (STAT3). J Biol Chem. 2012;287(44):37472–37482. doi:10.1074/jbc.M112.358911
32. Davies SP, Reynolds GM, Stamataki Z. Clearance of apoptotic cells by tissue epithelia: A Putative role for hepatocytes in liver efferocytosis. Front Immunol. 2018;9:44. doi:10.3389/fimmu.2018.00044
33. Patten DA, Wilkinson AL, O’Keeffe A, Shetty S. Scavenger receptors: Novel roles in the pathogenesis of liver inflammation and cancer. Semin Liver Disease. 2022;42(1):61–76. doi:10.1055/s-0041-1733876
34. National Institute of Environmental Health Sciences. Intrahepatocyte Erythrocytes and Pigment. The Digitized Atlas of Mouse Liver Lesions. Available from: https://www.niehs.nih.gov/research/resources/visual-guides/liverpath/miscellaneous/intrahepatocyte_erythrocytes/index.cfm.
35. Rosin A, Doljanski L. Erythrocytes in cytoplasms and nuclei of liver cells. The British Journ of Exp Pathol. 1944;25(4):111–114.
36. Pretorius E, du Plooy JN, Bester J. A comprehensive review on eryptosis. Cell Physiol Biochem. 2016;39:1977–2000. doi:10.1159/000447895
37. Fallon RJ, Danaher M. The effect of staurosporine, a protein kinase inhibitor, on asialoglycoprotein receptor endocytosis. Exp Cell Res. 1992;203(2):420–426. doi:10.1016/0014-4827(92)90016-2
38. Tuma DJ, Casey CA, Sorrell MF. Effects of ethanol on hepatic protein trafficking: Impairment of receptor-mediated endocytosis. Alcohol Alcohol. 1990;25(2–3):117–125. doi:10.1093/oxfordjournals.alcalc.a044986
39. Durocher JR, Payne RC, Conrad ME. Role of sialic acid in erythrocyte survival. Blood. 1975;45(1):11–20. doi:10.1182/blood.V45.1.11.11
40. Janicik JM, Schauer R, Andres KH, von During M. Sequestration of neuraminidase-treated erythrocytes. Studies on its topographic, morphologic and immunologic aspects. Cell Tissue Res. 1978;186(2):209–226. doi:10.1007/BF00225532
41. Matsuura S, Nakada H, Sawamura T, Tashiro Y. Distribution of an asialoglycoprotein receptor on rat hepatocyte cell surface. J Cell Biol. 1982;95(3):864–875. doi:10.1083/jcb.95.3.864
42. Bogdan AR, Miyazawa M, Hashimoto K, Tsuji Y. Regulators of iron homeostasis: New players in metabolism, cell death, and disease. Trends Biochem Sci. 2016;41(3):274–286. doi:10.1016/j.tibs.2015.11.012
43. Nikolic M, Stanic D, Antonijevic N, Niketic V. Cholesterol bound to hemoglobin in normal human erythrocytes: A new form of cholesterol in circulation? Clin Biochem. 2004;37(1):22–26. doi:10.1016/j.clinbiochem.2003.10.002
44. Lange Y, Molinaro AL, Chauncey TR, Steck TL. On the mechanism of transfer of cholesterol between human erythrocytes and plasma. J Biol Chem. 1983;258(11):6920–6926. doi:10.1016/S0021-9258(18)32311-1
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.