Back to Journals » Journal of Experimental Pharmacology » Volume 17
Artemisia annua Extract Ameliorates Atopic Dermatitis: Evidence from 3D Epidermal Model and Complementary in vitro Assays
Authors Tian Y ![]() , Jiao L, Li Y, Tian Y, Chen Y, Jia H, Ma L
, Jiao L, Li Y, Tian Y, Chen Y, Jia H, Ma L
Received 1 July 2025
Accepted for publication 4 October 2025
Published 15 October 2025 Volume 2025:17 Pages 707—720
DOI https://doi.org/10.2147/JEP.S550568
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Abdelwahab Omri
Yuanyuan Tian,1,2,* Lei Jiao,1,2,* Yiran Li,3,* Yi Tian,1,2 Yuanyuan Chen,4 Haidong Jia,4 Lin Ma1,2
1Department of Dermatology, Beijing Children’s Hospital, Capital Medical University, National Center for Children’s Health, Beijing, People’s Republic of China; 2Key Laboratory of Major Diseases in Children, Ministry of Education, Beijing, People’s Republic of China; 3Bioinformatics Facility, National Infrastructure for Translational Medicine, Institute of Clinical Medicine, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, People’s Republic of China; 4Shanghai Jahwa United Co., Ltd., Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lin Ma, Email [email protected] Haidong Jia, Email [email protected]
Background: Atopic dermatitis (AD) is a chronic and recurrent inflammatory skin disease. The disruption of the epidermal barrier and the inflammatory response are the key factors for the occurrence and development of this disease. Artemisia annua extract (AAE), a widely used traditional Chinese medicine, exhibits anti-inflammatory properties that may benefit AD management.
Objective: To evaluate the ability of AAE to inhibit inflammation and promote skin barrier repair in an AD-like three-dimensional (3D) epidermal equivalent model.
Methods: Keratinocytes were treated with three AAE concentrations (0.1%, 0.3%, and 1%) to assess their cytotoxic effects using Cell Counting Kit-8. Methyl-β-cyclodextrin and interleukin (IL)-4, IL-13, and IL-25 were used to induce the AD-like model. The expression of skin structural proteins, inflammatory factors, and histopathological manifestations were compared among AAE-treated AD models, an untreated AD model, and normal control models.
Results: Expression of the skin barrier proteins filaggrin (p < 0.0001, 95% CI: 0.7006 to 0.8265), loricrin (p < 0.0001, 95% CI: 0.2028 to 0.3031), and desmoglein-1 (p < 0.05, 95% CI: 0.0298 to 0.4227) was remarkably restored, whereas that of HAS3 (p < 0.05, 95% CI: 1.169 to 7.207), NELL2 (p < 0.0001, 95% CI: 5.787 to 6.978), TSLP (p < 0.01, 95% CI: 1.657 to 7.513), and IL-1α (p < 0.001, 95% CI: 19.33 to 63.35), IL-6 (p < 0.01, 95% CI: 2.474 to 13.78), and IL-8 (p < 0.0001, 95% CI: 36.55 to 55.63) was reduced significantly in 1% AAE concentration. AAE may exert its effects by inhibiting the over-activation of the MAPK pathway in an AD-like 3D epidermal model.
Conclusion: 1% AAE inhibits inflammation and promotes skin barrier function in an AD-like 3D epidermal equivalent model. AAE which mainly includes Arteannuin B, Chlorogenic Acid, Chrysoplenol D, Scopolin, and Vitexicarpin is identified as the putative targets for AD therapy.
Keywords: atopic dermatitis, epidermal equivalent model, Artemisia annua extract, skin barrier, anti-inflammatory
Introduction
Atopic dermatitis (AD) is a chronic inflammatory skin disease caused by genetic predisposition and environmental variables, associated with skin barrier dysfunction and immune system imbalances.1 AD is characterized by chronic eczema and severe pruritus, often with asthma and allergic rhinitis.2 The statistics of the WHO (World Health Organization) show that AD is the most common non-fatal skin disease.3 Globally, the prevalence of AD ranges from 2.7% and 20% in pediatric populations.4 AD and its related disorders can lead to a greatly reduced quality of life in patients and place increased financial strain on families and society.3,5
The pathogenesis of AD is complex and has not been fully elucidated yet, however, skin barrier defects and down-regulation of the helper T helper type 2 (Th2) immune response are regarded as the key factors causing the disease.6 Keratinocytes play an indispensable role in the human epidermis and serve as a barrier between the internal and external environments. They are immune cells that participate in innate and adaptive skin immune responses and are crucial for maintaining skin integrity.7 Decreased expression of epidermal proteins such as filaggrin (FLG), keratin, loricrin (LOR), and involucrin leads to epidermal barrier dysfunction, which increases sensitivity to allergens and activates keratinocytes. Any change in FLG levels may compromise skin barrier function.8 DSG1 is a protein found in corneodesmosomes that protects against skin dehydration and the entry of external antigens.9 Nevertheless, stimulation of Th2 cytokines increased the expression of genes associated with AD such as neural epidermal growth factor-like 2 (NELL2)10 and hyaluronan synthase 3 (HAS3) which is highly expressed in the middle of the spongiotic epidermis.10,11
Activated keratinocytes increasingly release pro-inflammatory cytokines, such as interleukin (IL)-1, IL-25, IL-33, and thymic stromal lymphopoietin (TSLP), which attract T cells and activate Langerhans and dendritic cells.7 Activated dendritic cells then stimulate Th2 cells, which aggravates the production of inflammatory cytokines, such as IL-4, IL-5, IL-13, and IL-31, and causes dysregulation of the immune response.12 In turn, IL-4 and IL-13 facilitate epidermal protein downregulation. Dendritic cells can also stimulate Th1, Th17, and Th22 cytokines, resulting in the overproduction of inflammatory cytokines, such as IL-1, IL-6, IL-8, IL-18, IL-25, and TSLP.13 Due to its chronic nature, AD requires long-term care and management. In addition to topical glucocorticoids, calcineurin inhibitors, and biologicals, moisturization is an important therapeutic strategy.
Various natural products can effectively reduce inflammation and restore skin barrier function with relatively few side effects. Many physiologically active substances discovered in plants, such as licorice and barley, are widely used in functional medicine and cosmetics.14,15 Artemisia annua is widely used in traditional Chinese medicine as it has diverse chemical constituents and extensive pharmacological activities against infectious and inflammatory diseases including fever, malaria and jaundice.16–18 Our collaborators prepared Artemisia annua extract (AAE), which mainly includes Arteannuin B, Chlorogenic Acid, Chrysoplenol D, Scopolin, and Vitexicarpin (Table S1). They found that AAE exhibited anti-inflammatory properties in AD-like mice.19 Based on this study, we conducted a more comprehensive and in-depth study on the AAE.
In this study, we first detected Arteannuin B, Chlorogenic Acid, Chrysoplenol D, Scopolin, and Vitexicarpin to identify therapeutic target proteins for AD and conducted bioinformatics analysis of the putative targets of AAE. Furthermore, we mainly investigated the ability of AAE to inhibit inflammation and promote skin barrier repair in AD in human epidermal models. We induced an in vitro AD-like 3D epidermal model to assess the anti-AD activity of AAE by detecting the expression of skin barrier related proteins and inflammatory factors to confirm the effectiveness, and attempting to analyze their possible signaling pathways.
Materials and Methods
The Preparation of AAE
The AAE (Artemisia annua aqueous extract, voucher No. 210706) was prepared by Shanghai Jahwa United Co., Ltd. The dried aerial part of Artemisia annua (1 kg) was extracted twice by refluxing with 15 times of distilled water for 1 h each time. After filtration, the supernatant was concentrated to 1 L under reduced pressure. Subsequently, 95% ethanol was added to the extract to adjust the final concentration to 70% at room temperature. After allowing it to stand overnight, the mixture was filtered to remove impurities (such as proteins and polysaccharides) and retain the small molecules at room temperature. The ethanol in the supernatant was then removed under reduced pressure. The concentrated supernatant was finally freezedried for easy storage. All voucher specimens were kept in the Institute of Chinese Materia Medica, China Academy of Chinese Medical Sciences.
Target Identification, Gene Ontology (GO), and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses of the Putative Targets of AAE
Our collaborators identified AAE, which mainly includes Arteannuin B, Chlorogenic Acid, Chrysoplenol D, Scopolin, and Vitexicarpin. AAE CAS number of effective ingredients from PubChem database (https://pubchem.ncbi.nlm.nih.gov) were retrieved and confirmed their chemical structures. We examined these chemicals to identify therapeutic target proteins for AD and performed GO and KEGG pathway enrichment analyses of the putative targets of AAE. All active ingredients of AAE were imported into the Swiss Target Prediction database (http://swisstargetprediction.ch/) and their corresponding targets were identified. The GeneCards database (https://www.genecards.org/) was used to screen for information on all AD-related genes. The R package clusterProfiler (version 4.12.6) was used for enrichment analysis of genes encoding target proteins, and GO term and KEGG signaling pathways with p values less than 0.05 were selected.
Cell Culture and Cell Cytotoxicity Assay
Keratinocytes obtained from Biocell (Guangdong, China) were maintained in KcGrowth medium (Biocell) supplemented with 35 mg/L cell growth factor (Biocell) and 876 mg/L glutamine. Keratinocytes were serially passaged at 60±5% confluence, and experiments were carried out using subconfluent cells at passage two, at which time they were proliferating actively. Cells were incubated at 37 °C in a humidified atmosphere with 95% air and 5% CO2. The cells were seeded in 96-well plates (1 × 104 cells/well) and incubated overnight to achieve a confluency of 40–60%.
The AAE group was treated with 100 μL AAE (0.03–3%) and incubated for 24 h. The control group (Control) was treated with 100 μL medium, and the wells without cells were used as the blank control group (BC). For the AD-like keratinocyte model, AAE (Shanghai Jahwa United Co., Ltd) was added to adjust the final concentrations to 0.1%, 0.3%, and 1%, and the untreated AD model and normal control model received an equivalent amount of KcGrowth medium. Following another 2 h of incubation, the untreated AD and AAE-treated models were induced using MβCD (3.75 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), and IL-25 (20 ng/mL) (MβCD/IL-mix). After incubation for 24 h, 10 μL Cell Counting Kit-8 (CCK-8; CK04, Dojindo, Japan) solution was added to each well and incubated for 4 h. The absorbance at 450 nm was measured using a TECAN Microplate Reader (Infinite M Nano, Switzerland). Three wells were used for each concentration of the drugs. Cell morphology was observed by the ZEISS AX10 optical microscope.
Construction of AD-Like 3D Epidermal Model
3D epidermal models (EpiKutis, ES210402, Biocell, Guangdong, China) were used to simulate AD-like skin. The 3D epidermal models were initially cultured in EpiGrowth medium (Biocell) at 37 °C and 5% CO2 in six-well plates (2.5×105 cells/well) for 24 h. To induce the AD-like model, MβCD/IL-mix was added into six-well plates and incubated for 24 h. For analysis, the supernatant from each sample was collected for ELISA, epidermis was separated for histopathological examination, and RNA was extracted for quantitative real-time PCR (qPCR) detection.
To evaluate the effects of AAE on the AD-like model, AAE (Shanghai Jahwa United Co., Ltd) was added to adjust the final concentrations to 0.1%, 0.3%, and 1%, and the untreated AD model and normal control model received an equivalent amount of EpiGrowth medium. Following another 2 h of incubation at 37 °C and 5% CO2, the untreated AD and AAE-treated models were induced using MβCD/IL-mix and incubated for 24 h. The analytical method is the same as above.
Histopathology
Slices of 3D epidermal models were subjected to hematoxylin-eosin and immunohistochemical staining for FLG. Paraffin sections of 3D epidermal models were stained with hemalum and erythrosine (HE) for morphological analysis. A light microscope was used to assess histological changes.
Analysis of mRNA Expression by qPCR
To assess the relative mRNA expression of specific genes, we extracted total RNA using the TRIzol reagent following the manufacturer’s protocol. Then, 4 µg of RNA was reverse-transcribed into cDNA using SuperScriptIII reverse transcriptase from Invitrogen. The resulting cDNA aliquots were stored at −20 °C. qPCR was performed using the Applied Biosystems 7500 Fast Real-Time PCR System, and TaqMan site-specific primers and probes were obtained from Applied Biosystems (Foster City, CA, USA). cDNA samples were tested for FLG, LOR, DSG1, Has3, and NELL2 expression, and the results were normalized to GAPDH levels. The primer sequences for these genes are listed in Table S2.
Analysis of Cytokine Arrays by ELISA
Culture supernatants were collected and stored at −80 °C. Cytokines, including TSLP, IL-1α, IL-6, and IL-8 released from 3D epidermal models, were analyzed using a series of Human Cytokine ELISA kits (Abcam, UK), namely TSLP (Abcam, ab192149), IL-1α (Abcam, ab178008), IL-6 (Abcam, ab178013), and IL-8 kits (Abcam, ab214030), respectively. Samples were assayed in three duplicates, and procedures were performed according to the manufacturer’s protocol.
Western Blots
3D models were lysed with RIPA lysis buffer (ThermoFisher, 89901) containing protease inhibitors (ThermoFisher, 78438). Protein concentration was determined by BCA assay, and cell lysates were resolved by SDS-PAGE on 4–12% gradient Bis-Tris gels (GenScript, M000138), transferred to PVDF membranes, and probed with antibodies against filaggrin (ThermoFisher, PA5-115235), IL-1α and TSLP (Cell Signaling Technology, 34819S and 97630S), p38 MAPK and p-p38 MAPK (Proteintech, 66234-1-Ig and 28796-1-AP), GAPDH (Cell Signaling Technology, 14C10), β-actin (Servicebio, GB15001-100), anti-rabbit IgG and anti-mouse IgG HRP-linked Antibody (Cell SignalingTechnology, 7074P2 and 7076P2). GAPDH and β-actin were used to normalize the protein expression using ImageJ software (ImageJ-win64).
Proteome and Transcriptome Sequencing
For the AD-like keratinocyte models, we performed proteomic and transcriptomic sequencing analysis of the control group, AD-like group (treated with MβCD/IL-mix only), and AAE-treated group (concentrations 0.1%, 0.3%, and 1%) (n=2). Omics analysis was conducted at Shanghai Applied Protein Technology Co., Ltd. All samples were prepared by in-solution digestion method. The peptide content was estimated by UV light spectral density at 280 nm. For DIA experiments, iRT (indexed retention time) calibration peptides were spiked into the sample (400 ng/sample). The peptides from each sample were analyzed by OrbitrapTM AstralTM mass spectrometer (Thermo Scientific) connected to an Vanquish Neo system liquid chromatography (Thermo Scientific) in the data-independent acquisition (DIA) mode.
Bioinformatic Analysis
Cluster 3.0 (http://bonsai.hgc.jp/mdehoon/software/cluster/software.htm) and Java Treeview software (http://jtreeview.sourceforge.net) were used to performing hierarchical clustering analysis. CELLO (http://cello.life.nctu.edu.tw/) which is a multi-class SVM classification system, was used to predict protein subcellular localization. Protein sequences are searched using the InterProScan software (interproscan-5.25–64.0) to identify protein domain signatures from the InterPro member database Pfam. The protein sequences of the selected differentially expressed proteins were locally searched using the NCBI BLAST+ client software (ncbi-blast-2.2.28+-win32.exe) and InterProScan, then gene ontology (GO) terms were mapped and sequences were annotated using the software program Blast2GO (BLASTP2.8.0+). Following annotation steps, the studied proteins were blasted against the online Kyoto Encyclopedia of Genes and Genomes (KEGG) database to retrieve their KEGG orthology identifications and were subsequently mapped to pathways in KEGG using KOBAS (KOBAS 3.0) software. Enrichment analysis were applied based on the Fisher’ exact test, considering the whole quantified proteins as background dataset. And only functional categories and pathways with p-values under a threshold of 0.05 were considered as significant. The results were downloaded in the XGMML format and imported into Cytoscape software (http://www.cytoscape.org/, version 3.2.1) to visualize.
Statistical Analysis
The study adopted One-way ANOVA to test the significant differences of continuous variables among different experimental groups. After the ANOVA analysis found significant differences between groups, Tukey’s test method was used to conduct multiple comparisons of the means of each group which had significant mean differences. The significance was indicated by * p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. All experiments were performed with a minimum of three independent biological replicates (n≥3). Results were expressed as mean ± standard deviation (SD). All statistical analyses were completed by SPSS (v26.0), and the visualization results were generated by GraphPad Prism 9 softwear.
Results
GO and KEGG Pathway Enrichment Analyses of the Putative Targets of AAE for AD Therapeutics
As shown in Figure 1, the GO functional enrichment analysis revealed that the biological processes primarily involved regulation of inflammatory responses, regulation of protein processing, response to glucocorticoid, response to stimulus, metabolic process, immune system process, cellular response to tumor necrosis factor, and Th1 and Th2 cell differentiation. KEGG analysis revealed more than 10 overlapping pathways, including MAPK, PI3K-AKT, NF-κB, mTOR, cAMP, calcium, cGMP-PKG, inflammatory mediator regulation of TRP channels, TNF, and JAK-STAT signaling pathways. Bioinformatics analysis indicated that AAE may exert its effects via the MAPK, NF-κB, and PI3K-AKT signaling pathways.
 |
Figure 1 GO and KEGG pathway enrichment analyses of the putative targets of different components of Artemisia annua extract (AAE) for AD treatment (a-e are Arteannuin B, Chlorogenic Acid, Scopolin, Vitexicarpin and Chrysoplenol D, respectively). |
Effect of AAE on Keratinocytes Proliferation in vitro
In order to identify the validity of AAE on AD, keratinocytes were initially used to determine whether AAE affected normal cell growth. Subsequently, experiments were performed using a 3D epidermal model. The AAE stock solution was diluted to final concentrations of 0.03%, 0.1%, 0.3%, 1%, and 3%. None of these concentrations (0.03%, 0.1%, 0.3%, and 1%) exhibited cytotoxicity or significant differences from the control group, as depicted in Figure 2a (* p < 0.05). The AAE dosing concentrations for the subsequent experiments with keratinocytes and 3D models were 0.1%, 0.3%, and 1%. Keratinocytes were treated with MβCD/IL-4/IL-13/IL-25 (MβCD/IL-mix), and the cell viabilities were examined. Compared to the control group, different AAE concentrations showed no obvious effect on cell proliferation activity (Figure 2b). Therefore, no significant cytotoxicity was observed at AAE concentrations ranging from 0.1% to 1%.
 |
Figure 2 Cytotoxicity of different AAE concentrations in keratinocytes cells. (a) Cell viabilities of keratinocytes after AAE (0.1–3%) treatment for 24 h; 0 group, treated with DMEM; AAE group, treated with AAE (0.1–3%); DMSO (positive control) group, treated with 10% DMSO (n=5). (b) Cell viabilities of keratinocytes after AAE (0.1–3%) treatment for 24 h in MβCD//IL-4/IL-13/IL-25-induced AD-like models (n=5) (*p < 0.05, significant difference; **p < 0.01, highly significant difference; ***p < 0.001, extremely significant difference; ****p < 0.0001, ultra-significant difference). Abbreviations: AD, Atopic dermatitis; AAE, Artemisia annua extract; MβCD, methyl-b-cyclodextrin; IL, interleukin; DMSO, Dimethyl sulfoxide. |
MβCD/IL-Mix Induced AD-Like 3D Epidermal Model
3D epidermal models were treated with MβCD/IL-mix, and the AD-like characteristics were examined. Histopathology revealed hyperkeratosis, mild intercellular spongiosis, and keratinocyte necrosis in the AD-like groups. Immunohistochemical staining showed that the expression of the structural protein FLG was significantly decreased compared with the control group (Figure 3a). A concentration-dependent decrease in FLG mRNA expression (50: MβCD (7.5 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), IL-25 (20 ng/mL), ****p < 0.0001, 95% Confidence Interval, 95% CI: 0.4008 to 0.7264) and an increase in NELL2 expression (50: MβCD (7.5 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), IL-25 (20 ng/mL), *p < 0.05, 95% CI: 0.05182 to 0.3994) were also observed (Figure 3b). Inflammatory cytokines were detected using ELISA. The results indicated a concentration-dependent increase of TSLP (50: MβCD (7.5 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), IL-25 (20 ng/mL), ****p < 0.0001, 95% CI: 26.39 to 48.33) and IL-1α (50: MβCD (7.5 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), IL-25 (20 ng/mL), ****p < 0.0001, 95% CI: 29.55 to 38.58) in the MβCD/IL-mix (Figure 3c), with statistically significant results. These results suggest that the MβCD/IL-mix can induce an AD-like 3D epidermal model.
 |
Figure 3 MβCD/IL-mix-induced AD-like phenotypes and expression of related genes in 3D epidermal models. (a) Histological sections of 3D models treated with different concentrations of MβCD/IL-4/IL-13/IL-25. 25: MβCD (7.5 mM), IL-4 (25 ng/mL), IL-13 (25 ng/mL), IL-25 (10 ng/mL); 50: MβCD (7.5 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), IL-25 (20 ng/mL); 100: MβCD (7.5 mM), IL-4 (100 ng/mL), IL-13 (100 ng/mL), IL-25 (40 ng/mL). Scale bar 50 µm (n=3). (b) qPCR analysis (n=4) (*p < 0.05, **p < 0.01, ****p < 0.0001, n.s., not significant). (c) Cytokine array TSLP and IL-1α analysis (n=3) (**p < 0.01, ****p < 0.0001). Abbreviations: AD, Atopic dermatitis; AAE, Artemisia annua extract; 3D, three dimensional; MβCD, methyl-b-cyclodextrin; IL, interleukin; FLG, filaggrin; NELL2, neural epidermal growth factor-like 2; TSLP, thymic stromal lymphopoietin; qPCR, quantitative PCR. |
AAE Repairs Abnormal Phenotypes in the MβCD/IL-Mix-Induced AD-Like 3D Epidermal Model
To assess the impact of AAE on 3D epidermal models resembling AD, AAE was administered alongside MβCD/IL-mix during 24 h of air-liquid interface culture. Hematoxylin-eosin staining showed intercellular sponge-like lesions lacking keratin hyaluron granules in the AD-like group (treated with MβCD/IL-mix only), whereas the AAE-treated groups (concentrations 0.1%, 0.3%, and 1%) demonstrated less spongiosis and more keratohyalin granules, especially the 1% AAE-treated group (Figure 4a). Immunohistochemical staining showed that FLG expression was reduced in the AD-like group but increased in the AAE-treated groups (concentrations 0.1%, 0.3%, and 1%) (Figure 4a). In addition, FLG expression was observed in 3D epidermal models using Western blotting (Figure 4b and c). FLG expression was decreased in the untreated AD model group compared to that in the control group. Further, 1% AAE significantly increased the expression of FLG (**p < 0.01). This result is consistent with those of the hematoxylin-eosin and immunohistochemical staining. The isolated 3D epidermis was analyzed for mRNA expression to determine whether AAE influenced AD-related gene expression. The results indicated that AAE treatment led to a dose-dependent restoration of FLG (1% AAE: ****p < 0.0001, CI: 0.7006 to 0.8265), LOR (1% AAE: ****p < 0.0001, 95% CI: 0.2028 to 0.3031) and DSG1 (1% AAE: *p < 0.05, 95% CI: 0.0298 to 0.4227) gene expression, which was decreased in the AD-like group. Similarly, AAE treatment reduced the mRNA levels of HAS3 (1% AAE: *p < 0.05, 95% CI: 1.169 to 7.207) and NELL2 (1% AAE: ****p < 0.0001, 95% CI: 5.787 to 6.978), which were elevated in the AD-like group (Figure 4d).
 |
Figure 4 AAE regulated expression of AD-related genes in MβCD/IL-mix-induced AD-like skin equivalent model. 3D epidermal models were cultured with MβCD/IL-mix (MβCD (3.75 mM), IL-4 (50 ng/mL), IL-13 (50 ng/mL), and IL-25 (20 ng/mL)) and different concentrations of AAE. (a) Histological section of 3D models treated with different concentrations of AAE. Scale bar 50 µm (n=3). (b and c) Protein expressions of FLG determined by Western blot (n=4) (*p < 0.05, **p < 0.01, ***p < 0.001). (d) qPCR analysis (n=3) (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001). Abbreviations: ns, not significant; AD, Atopic dermatitis; AAE, Artemisia annua extract; 3D, three dimensional; MβCD, methyl-b-cyclodextrin; IL, interleukin; FLG, filaggrin; LOR, loricrin; DSG1, desmoglein-1; HAS3, hyaluronan synthase 3; NELL2, neural epidermal growth factor-like 2; GAPDH: glyceraldehyde-3-phosphate dehydrogenase; qPCR, quantitative PCR. |
Additionally, ELISA showed that AAE significantly decreased the production of pro-inflammatory cytokines TSLP (1% AAE, **p < 0.01, 95% CI: 1.657 to 7.513), IL-1α (1% AAE, ***p < 0.001, 95% CI: 19.33 to 63.35), IL-6 (1% AAE, **p < 0.01, 95% CI: 2.474 to 13.78), and IL-8 (1% AAE, ****p < 0.0001, 95% CI: 36.55 to 55.63) which were elevated by MβCD/IL-mix treatment (Figure 5a). Similarly, we also tested the expression of TSLP and IL-1α using Western blotting (Figure 5b). With increasing AAE concentration, the inflammatory response decreased gradually (Figure 5c). These findings suggest that AAE mitigates AD-like symptoms in the 3D epidermal models by regulating AD-related mRNA, proteins, and pro-inflammatory cytokines.
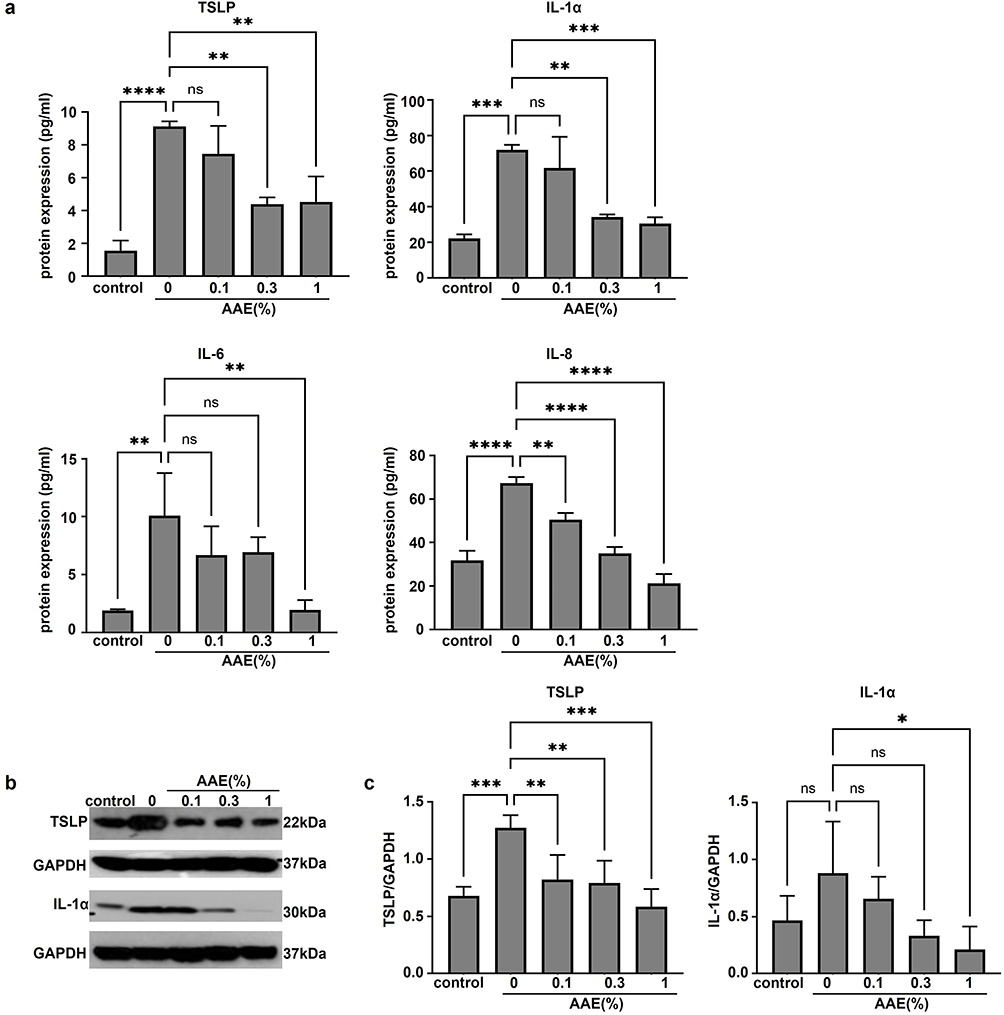 |
Figure 5 AAE partially improved the AD-related cytokine expression in AD-like skin equivalent model. (a) Cytokine array TSLP, IL-1α, IL-6 and IL-8 analysis (n=3) (**p < 0.01, ***p < 0.001, ****p < 0.0001, n.s., not significant). (b and c) Protein expressions of IL-1α and TSLP determined by Western blot (n=4) (*p < 0.05, **p < 0.01, ***p < 0.001). Abbreviations: ns, not significant; AD, Atopic dermatitis; AAE, Artemisia annua extract; IL, interleukin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; TSLP, thymic stromal lymphopoietin. |
AAE Inhibited the Transcriptional Regulation of MAPK Signal Pathway
We performed proteome and transcriptome sequencing of keratinocytes and AD-like models to verify the possible signaling pathway involved in the mitigating effect of AAE on AD. The AAE-treated group consists of three AAE concentrations (0.1%, 0.3%, and 1%), with two biological replicates for each concentration due to constraints in sample availability and resource limitations. When comparing the AD-like group and the control group, the up-regulated proteins were involved in signaling pathways such as MAPK (Figure 6a), whereas signaling pathways enriched in down-regulated proteins in the AAE treated groups and AD-like group also included MAPK (Figure 6b). In addition, the expression of P38 MAPK (mitogen-activated protein kinase) was also detected using Western blot (Figure 6c and d). In the AD-like 3D epidermal model, the phosphorylation of p38 MAPK was enhanced. However, the addition of AAE resulted in decreased phosphorylation levels (*p < 0.05). The results showed that the attenuated symptoms of skin lesions and inflammatory responses were closely related to the transcriptional regulation of the MAPK signaling pathway.
 |
Figure 6 Effects of AAE on MAPK signaling pathway. (a and b) Proteome and transcriptome sequencing for MβCD/IL-mix-induced AD-like keratinocytes models. (a) AD-like group compared with the control group, the up-regulated proteins involved in signaling pathways; (b) signaling pathways involved in down-regulated proteins in AAE treated groups and AD-like group (n=2). (c and d) Western blot of p-P38 and P38 in AD-like 3D epidermal models, n=3/group. Compared with AD-like group (n=3) (*p < 0.05). Abbreviations: ns, not significant; AD, Atopic dermatitis; AAE, Artemisia annua extract; 3D, three dimensional; MβCD, methyl-b-cyclodextrin; IL, interleukin; MAPK, mitogen-activated protein kinase. |
Discussion
AD is characterized by early onset, a persistent nature, and high likelihood of recurrence, with a chronic course that can last for a lifetime.20 Patients with AD experience intense itching, frequent scratching, sleep disturbances, and dietary restrictions, which lead to a significant decline in their quality of life. The complex pathogenesis and clinical recurrence hinder the effective prevention of AD, resulting in a high prevalence rate, an increasing number of affected individuals, and substantial consumption of public health resources. While the emergence of biologics and targeted small molecule drugs has provided hope for AD treatment, some limitations remain.21 Therefore, natural herbal extracts have received increasing attention in research due to their potential efficacy and fewer adverse reactions.
Epidermal equivalent models, which have been developed and used to study skin physiology for more than 30 years and often used to explore new potential treatments for AD and psoriasis, represent a 3D reconstruction of the human epidermis from primary keratinocytes extracted from the skin of healthy individuals. Reconstructing the human epidermis in vitro is a useful tool for overcoming the limitations of AD animal models, which are less deterministic and stable.22 The most common method for studying AD is to treat with Th2 cytokines in 3D epidermal models.10,23 Moreover, IL-4, IL-13, and Th2 cytokines are commonly used to induce AD in 3D epidermal models.24,25 IL-25 was initially identified as a promoter of Th2 immune responses after stimulating Th2 cell cytokines, such as IL-13 and IL-4.26 Furthermore, depleting plasma membrane cholesterol via MβCD administration in 3D epidermal models, followed by exposure to IL-4, IL-13, and IL-25, can effectively replicate the characteristics of AD.23 The 3D epidermal models of EpiKutis used in our research are normal epidermal models commercially produced in China.27 Therefore, to build on previous research, we investigated the responses of 3D epidermal models to MβCD treatment when subsequently exposed to a combination of cytokines, thereby establishing an AD-like model and determining the experimental dose of MβCD and cytokines.
FLG and LOR are proteins involved in the formation and maintenance of the epidermal barrier, and play important roles in the pathophysiology of AD. Any change in FLG levels may compromise the skin barrier function.8 DSG1 is a protein found in corneodesmosomes that protects against skin dehydration and the entry of external antigens entry.9 Therefore, in this study, the expression levels of these genes were used to evaluate skin barrier function. Pro-inflammatory cytokines, including TSLP, IL-1a, IL-6, and IL-8, are increasingly released by keratinocytes in AD. TSLP, which is highly expressed in AD, is responsible for irritation and itching, which are prominent in disease pathology.7 Therefore, the concentrations of these cytokines were measured to evaluate the inflammatory response in AD. Our results revealed that MβCD and a combination of IL-4, IL-13, and IL-25 induced AD-like manifestations in EpiKutis, including skin barrier weakening, typical histological features, and an inflammatory response in keratinocytes.
Artemisia annua is a traditional Chinese medicine used to treat various diseases that contains a variety of chemical components, including sesquiterpenoids, flavonoids, fatty acids, polyphenols, volatile oils, and coumarins.19 Modern pharmacological studies have confirmed the anti-inflammatory, immunomodulatory, anti-malarial, and anti-bacterial properties of this plant.16,28 Since the discovery of artemisinin and its excellent anti-malarial activity, Artemisia annua has received considerable attention. Studies have demonstrated that artemisinin and its derivatives exhibit anti-inflammatory properties in AD-like mice.29 Animal experiments demonstrated that topical application of AAEO (a volatile oil extracted from Artemisia annua) significantly ameliorated skin lesions, reduced dermatitis score, and decreased spleen weight compared to DNCB (2,4-dintrochlorobenzene) treatment.30 Artemisia annua water extract (AWE) attenuates DNCB-induced atopic dermatitis by restraining Th2 cell-mediated inflammatory responses in BALB/c mice.31 Artemisia annua has been widely studied for its medicinal properties, and various extraction methods have been employed to obtain bioactive compounds, including essential oils, ethanol extracts, and aqueous extracts. However, the present study introduces a novel aqueous extract of Artemisia annua (AAE) that differs significantly in both composition and biological activity from previously reported extracts. Essential oils derived from Artemisia annua are rich in volatile terpenes and are primarily investigated for their antimicrobial and antioxidant effects.30 In contrast, ethanol extracts typically contain a broader spectrum of lipophilic compounds, including artemisinin and flavonoids, and have been mainly explored for antimalarial and anti-inflammatory activities.16
AAE, as prepared in this study, is distinguished by its high content of water-soluble polyphenols such as chlorogenic acid, scopolin, chrysoplenol D, and vitexicarpin, which are less abundant in other extract types. Based on existing literature, each of these compounds may contribute to the observed anti-AD effects through distinct mechanisms. Chlorogenic acid, a polyphenolic compound widely reported for its anti-inflammatory and antioxidant properties, has been shown to suppress pro-inflammatory cytokine production and inhibit NF-κB and MAPK signaling pathways in various inflammatory models.32 Scopolin, a coumarin derivative, has demonstrated immunomodulatory effects by reducing Th2-mediated inflammation and inhibiting histamine release, which are key features of AD pathogenesis.33 Chrysoplenol D, a flavonoid, is known for its ability to modulate immune responses and inhibit the activation of inflammatory signaling cascades, including MAPK and STAT pathways.34 Vitexicarpin, another flavonoid, has been reported to exhibit potent anti-inflammatory activity by downregulating the expression of TNF-α, IL-6, and other cytokines involved in AD.35 While our study did not isolate or test each compound individually, the collective presence of these constituents in AAE likely contributes synergistically to its overall anti-AD effects. Future studies should aim to isolate these compounds and evaluate their individual and combined effects in relevant AD models to further elucidate their specific roles.
Importantly, while previous studies have mostly focused on the antimalarial or general anti-inflammatory properties of Artemisia annua extracts, the present work demonstrates the specific efficacy of AAE in improving skin barrier function and suppressing inflammatory responses in an in vitro model of atopic dermatitis. The unique combination of bioactive compounds in AAE appears to contribute to its potent inhibition of the MAPK signaling pathway and reduction of key pro-inflammatory cytokines. AAE treatment resulted in significant inhibition of the MAPK signaling pathway, as evidenced by reduced phosphorylation levels of p38 in the 3D epidermal model. This suggests that AAE may interfere with upstream activators of the MAPK cascade, such as receptor tyrosine kinases or RAS GTPases, thereby attenuating the downstream phosphorylation events that lead to the activation of pro-inflammatory transcription factors like NF-κB.36 Given the critical role of MAPK signaling in the production of pro-inflammatory cytokines and the impairment of skin barrier function, the modulation of this pathway by AAE likely represents a key mechanism underlying its observed anti-AD effects. Future studies should aim to identify the specific molecular targets of AAE within the MAPK cascade and assess whether its effects are mediated through direct interaction with pathway components or indirect modulation via upstream regulators. Sawhney et al has demonstrated the anti-inflammatory effects in murine models and macrophages via down-regulation of MAPK signaling. The Arteannuin-B component plays a role. The conclusions of the two models are consistent.37
While our study provides valuable insights into anti-AD effects of AAE, it is important to acknowledge its limitations. Our experiments were conducted exclusively in in vitro models, which, although useful for mechanistic studies, may not fully mimic the physiological complexity of in vivo systems. Therefore, the translational relevance of our findings remains to be validated in animal models or clinical settings. The sample size in some assays, particularly the omics analyses may limit the statistical power and generalizability of the results, and larger sample sizes will be necessary in future studies to strengthen these findings. While we observed significant changes in key pathways such as MAPK signaling, the precise mechanisms through which the active compounds in AAE exert their effects are not yet fully elucidated. Further fractionation and identification of the specific bioactive components, along with detailed mechanistic studies, are required to clarify their roles and interactions.
Conclusion
In this study, we investigated the potential therapeutic effects of AAE in an AD-like epidermal model. The results showed that AAE improved AD-like characteristics in models stimulated by MβCD/IL-mix. AAE suppressed the expression of the inflammatory cytokines TSLP, IL-1α, IL-6, and IL-8, enhanced the expression of FLG, LOR, and DSG1 mRNA, significantly decreased the mRNA levels of NELL2 and HAS3 genes, and partially repaired AD-like structures. This study provides compelling evidence that AAE significantly enhances skin barrier function and attenuates inflammatory responses through the inhibition of the MAPK signaling pathway. In the present study, we report the concurrent enhancement of skin barrier integrity and inhibition of key pro-inflammatory mediators by AAE in a 3D skin model. Our findings indicate that AAE exerts its effects through the coordinated modulation of barrier repair and immune response pathways, suggesting a dual mechanism of action not previously described for this compound. These results not only advance our understanding of the pathophysiology of AD but also highlight the therapeutic potential of AAE as a multi-target agent for treating inflammatory skin disorders. Further research should focus on elucidating the mechanism underlying the anti-AD effects of AAE to advance its potential application as a treatment for AD. Given the limitations of in vitro models, further studies using animal models are essential to confirm the biological activity and safety profile of AAE. Such studies would help bridge the gap between preclinical findings and potential clinical applications. Upon successful validation in preclinical models, early-phase (Phase I/II) clinical trials should be conducted to evaluate the safety, tolerability, and preliminary efficacy of AAE in humans. These trials would represent a critical step toward translating our findings into clinical practice.
Abbreviations
AD, Atopic dermatitis; Th2, T helper type 2; AAE, Artemisia annua extract; 3D, three dimensional; MβCD, methyl-b-cyclodextrin; IL, interleukin; FLG, filaggrin; LOR, loricrin; DSG1, desmoglein-1; HAS3, hyaluronan synthase 3; NELL2, neural epidermal growth factor-like 2; GAPDH: glyceraldehyde-3-phosphate dehydrogenase; TSLP, thymic stromal lymphopoietin; qPCR, quantitative PCR; MAPK, mitogen-activated protein kinase; DMSO, Dimethyl sulfoxide; CI: Confidence Interval.
Data Sharing Statement
The data that support the findings of this study are available from either corresponding Lin Ma or corresponding Haidong Jia upon reasonable request. Lin Ma is responsible for the basic experimental part, while Haidong Jia is in charge of the extraction of Artemisia annua.
Acknowledgments
Yuanyuan Tian, Lei Jiao and Yiran Li are co-first authors for this study. The authors would like to thank Pro. Kaihu Yao and Dan Yu for support.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
Natural Science Foundation of Beijing Municipality (7244333); National Key R&D Program of China (2023YFC2508101); Beijing Hospitals Authority’s Ascent Plan (DFL20241201).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Yang G, Seok JK, Kang HC, et al. Skin barrier abnormalities and immune dysfunction in atopic dermatitis. Int J Mol Sci. 2020;21(8):2867. doi:10.3390/ijms21082867
2. Weidinger S, Novak N. Atopic dermatitis. Lancet. 2016;387(10023):1109–1122. doi:10.1016/s0140-6736(15)00149-x
3. Vos T, Flaxman AD, Naghavi M, et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380(9859):2163–2196. doi:10.1016/s0140-6736(12)61729-2
4. Barne M, Singh S, Mangal DK, et al. Global Asthma Network Phase I, India: results for allergic rhinitis and eczema in 127,309 children and adults. J Allergy Clin Immunol Global. 2022;1(2):51–60. doi:10.1016/j.jacig.2022.01.004
5. Ständer S. Atopic Dermatitis. New Engl J Med. 2021;384(12):1136–1143. doi:10.1056/NEJMra2023911
6. Humeau M, Boniface K, Bodet C. Cytokine-mediated crosstalk between keratinocytes and T cells in atopic dermatitis. Front Immunol. 2022;13:801579. doi:10.3389/fimmu.2022.801579
7. Das P, Mounika P, Yellurkar ML, et al. Keratinocytes: an enigmatic factor in atopic dermatitis. Cells. 2022;11(10):1683. doi:10.3390/cells11101683
8. Elias MS, Long HA, Newman CF, et al. Proteomic analysis of filaggrin deficiency identifies molecular signatures characteristic of atopic eczema. J Allergy ClinIimmunol. 2017;140(5):1299–1309. doi:10.1016/j.jaci.2017.01.039
9. Park N-J, Bong S-K, Lee S, et al. Compound K improves skin barrier function by increasing SPINK5 expression. J Ginseng Res. 2020;44(6):799–807. doi:10.1016/j.jgr.2019.11.006
10. Lee SH, Bae I-H, Choi H, et al. Ameliorating effect of dipotassium glycyrrhizinate on an IL-4- and IL-13-induced atopic dermatitis-like skin-equivalent model. Arch Dermatol Res. 2019;311(2):131–140. doi:10.1007/s00403-018-1883-z
11. Ohtani T, Memezawa A, Okuyama R, et al. Increased hyaluronan production and decreased E-cadherin expression by cytokine-stimulated keratinocytes lead to spongiosis formation. J Investig Dermatol. 2009;129(6):1412–1420. doi:10.1038/jid.2008.394
12. Kim W-H, An H-J, Kim J-Y, et al. Beneficial effects of melittin on ovalbumin-induced atopic dermatitis in mouse. Sci Rep. 2017;7(1):17679. doi:10.1038/s41598-017-17873-2
13. Purwar R, Werfel T, Wittmann M. IL-13-stimulated human keratinocytes preferentially attract CD4+CCR4+ T cells: possible role in atopic dermatitis. J Investig Dermatol. 2006;126(5):1043–1051. doi:10.1038/sj.jid.5700085
14. Milam EC, Rieder EA. An approach to cosmeceuticals. J Drugs Dermatol. 2016;15(4):452–456.
15. Min S-Y, Park C-H, Yu H-W, et al. Anti-inflammatory and anti-allergic effects of saponarin and its impact on signaling pathways of RAW 264.7, RBL-2H3, and HaCaT cells. Int J Mol Sci. 2021;22(16):8431. doi:10.3390/ijms22168431
16. Lang SJ, Schmiech M, Hafner S, et al. Antitumor activity of an Artemisia annua herbal preparation and identification of active ingredients. Phytomedicine. 2019;62:152962. doi:10.1016/j.phymed.2019.152962
17. Qiu F, Liu J, Mo X, et al. Immunoregulation by artemisinin and its derivatives: a new role for old antimalarial drugs. Front Immunol. 2021;12:751772. doi:10.3389/fimmu.2021.751772
18. Zhang X-B, Zhao Y-P, Huang X-W, et al. [Review on study of Dao-di herbs Artemisiae Annuae Herba]. Zhongguo Zhong yao za zhi. 2016;41(11):2015–2018. doi:10.4268/cjcmm20161107
19. Zhao Y, Zhu L, Yang L, et al. In vitro and in vivo anti-eczema effect of Artemisia annua aqueous extract and its component profiling. J Ethnopharmacol. 2024;318(Pt B):117065. doi:10.1016/j.jep.2023.117065
20. Guo Y, Zhang H, Liu Q, et al. Phenotypic analysis of atopic dermatitis in children aged 1–12 months: elaboration of novel diagnostic criteria for infants in China and estimation of prevalence. J Eur Acad Dermatol Venereol. 2019;33(8):1569–1576. doi:10.1111/jdv.15618
21. Dębińska A. New treatments for atopic dermatitis targeting skin barrier repair via the regulation of FLG expression. J Clin Med. 2021;10(11):2506. doi:10.3390/jcm10112506
22. Vuyst ED, Salmon M, Evrard C, et al. Atopic dermatitis studies through in vitro models. Front Med. 2017;4:119. doi:10.3389/fmed.2017.00119
23. Pedrosa TDN, Vuyst ED, Mound A, et al. Methyl-β-cyclodextrin treatment combined to incubation with interleukin-4 reproduces major features of atopic dermatitis in a 3D-culture model. Arch Dermatol Res. 2017;309(1):63–69. doi:10.1007/s00403-016-1699-7
24. Danso MO, Drongelen VV, Mulder A, et al. TNF-α and Th2 cytokines induce atopic dermatitis-like features on epidermal differentiation proteins and stratum corneum lipids in human skin equivalents. J Investig Dermatol. 2014;134(7):1941–1950. doi:10.1038/jid.2014.83
25. Rouaud-Tinguely P, Boudier D, Marchand L, et al. From the morphological to the transcriptomic characterization of a compromised three-dimensional in vitro model mimicking atopic dermatitis. Br J Dermatol. 2015;173(4):1006–1014. doi:10.1111/bjd.14012
26. Gregory LG, Jones CP, Walker SA, et al. IL-25 drives remodelling in allergic airways disease induced by house dust mite. Thorax. 2013;68(1):82–90. doi:10.1136/thoraxjnl-2012-202003
27. Qiu J, Zhong L, Zhou M, et al. Establishment and characterization of a reconstructed Chinese human epidermis model. Int J Cosmet Sci. 2016;38(1):60–67. doi:10.1111/ics.12249
28. Feng X, Cao S, Qiu F, et al. Traditional application and modern pharmacological research of Artemisia annua L. Pharmacol Ther. 2020;216:107650. doi:10.1016/j.pharmthera.2020.107650
29. Bai X-Y, Liu P, Chai Y-W, et al. Artesunate attenuates 2, 4-dinitrochlorobenzene-induced atopic dermatitis by down-regulating Th17 cell responses in BALB/c mice. Eur J Pharmacol. 2020;874:173020. doi:10.1016/j.ejphar.2020.173020
30. Huang Z, Fan B, Mao W, et al. Topical application of Artemisia annua L. essential oil ameliorates 2,4-dintrochlorobenzene-induced atopic dermatitis in mice. J Ethnopharmacol. 2024;333:118439. doi:10.1016/j.jep.2024.118439
31. Han X, Chen Z, Yuan J, et al. Artemisia annua water extract attenuates DNCB-induced atopic dermatitis by restraining Th2 cell mediated inflammatory responses in BALB/c mice. J Ethnopharmacol. 2022;291:115160. doi:10.1016/j.jep.2022.115160
32. Huang J, Xie M, He L, et al. Chlorogenic acid: a review on its mechanisms of anti-inflammation, disease treatment, and related delivery systems. Front Pharmacol. 2023;14:1218015. doi:10.3389/fphar.2023.1218015
33. Gao X-Y, Li X-Y, Zhang C-Y, et al. Scopoletin: a review of its pharmacology, pharmacokinetics, and toxicity. Front Pharmacol. 2024;15:1268464. doi:10.3389/fphar.2024.1268464
34. Lang SJ, Schmiech M, Hafner S, et al. Chrysosplenol d, a Flavonol from Artemisia annua, induces ERK1/2-mediated apoptosis in triple negative human breast cancer cells. Int J Mol Sci. 2020;21(11):4090. doi:10.3390/ijms21114090
35. Rakha A, Umar N, Rabail R, et al. Anti-inflammatory and anti-allergic potential of dietary flavonoids: a review. Biomed Pharmacother. 2022;156:113945. doi:10.1016/j.biopha.2022.113945
36. Bahar ME, Kim HJ, Kim DR. Targeting the RAS/RAF/MAPK pathway for cancer therapy: from mechanism to clinical studies. Signal Transduct Target Ther. 2023;8(1):455. doi:10.1038/s41392-023-01705-z
37. Sawhney G, Rasool JU, Saroch D, et al. Arteannuin-B and (3-Chlorophenyl)-2-Spiroisoxazoline derivative exhibit anti-inflammatory effects in LPS-activated RAW 264.7 macrophages and BALB/c mice-induced proinflammatory responses via downregulation of NF-κB/P38 MAPK signaling. Molecules. 2022;27(22):8086. doi:10.3390/molecules27228068
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.