Back to Journals » Infection and Drug Resistance » Volume 19
Applications and Challenges of CRISPR-Cas Technology for the Detection of Antimicrobial Resistance Genes
Received 3 November 2025
Accepted for publication 24 March 2026
Published 9 April 2026 Volume 2026:19 578705
DOI https://doi.org/10.2147/IDR.S578705
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sandip Patil
Wenting Tang,1,* Ninglei Yang,2,* Meijia Shi1
1Department of Laboratory Medicine, The Second Affiliated Hospital of Jiaxing University, Jiaxing, Zhejiang, People’s Republic of China; 2Department of Laboratory Medicine, Shanghai First Rehabilitation Hospital, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Meijia Shi, Department of Laboratory Medicine, The Second Affiliated Hospital of Jiaxing University, Jiaxing, Zhejiang, People’s Republic of China, Email [email protected]
Abstract: The global rise of antimicrobial resistance (AMR) demands rapid and precise diagnostic tools capable of providing actionable results at the point of care. Traditional methods, including bacterial culture and PCR, face critical limitations: culture-based approaches require days to weeks for results, potentially delaying life-saving treatment decisions, while PCR-based methods, though faster, require expensive instrumentation, trained personnel, and are typically confined to centralized laboratory settings. CRISPR-Cas systems offer a promising approach for detecting bacterial drug resistance genes with high specificity, speed, and sensitivity. By utilizing Cas enzymes (eg, Cas12 and Cas13) and guide RNAs, CRISPR enables targeted recognition of resistance markers, typically requiring upstream nucleic acid amplification (eg, recombinase polymerase amplification or loop-mediated isothermal amplification) to achieve clinically relevant sensitivity. Platforms such as SHERLOCK and DETECTR facilitate rapid identification of markers associated with pathogens like MRSA and CRE directly from clinical samples, supporting point-of-care use. Integration with microfluidics and AI further expands its potential for personalized therapy and AMR surveillance. Despite ethical and regulatory challenges, CRISPR-Cas technology holds significant promise for advancing antimicrobial stewardship and public health responses to AMR.
Keywords: CRISPR-Cas systems, antimicrobial resistance, molecular diagnostics, resistance gene markers, point-of-care testing
Introduction
The escalating prevalence and diversity of drug-resistant bacteria on a global scale have rendered drug-resistant bacterial infections a significant threat to public health worldwide.1 The scale of this crisis is underscored by recent epidemiological data: a systematic analysis published in 2024 estimated that bacterial AMR was directly responsible for 1.14 million deaths globally in 2021 and contributed to 4.71 million deaths. Furthermore, the World Health Organization’s Global Antibiotic Resistance Surveillance Report 2025 indicates that one in six laboratory-confirmed bacterial infections worldwide in 2023 was resistant to antibiotics, with resistance rates increasing in over 40% of monitored pathogen–antibiotic combinations between 2018 and 2023.2 Without effective intervention, the annual death toll directly attributable to AMR is projected to rise to 10 million by 2050. By identifying resistance genes, clinicians can optimize antibiotic selection, thereby improving therapeutic outcomes and reducing inappropriate antibiotic use.3–5 Furthermore, rapid identification of these genes plays a crucial role in the management of nosocomial infections, enhancing the implementation of isolation protocols and infection control strategies.6,7
While traditional culture methods are considered the “gold standard” for pathogen detection,8 they are inherently time-consuming, potentially delaying treatment for critically ill patients. For instance, cultures of Mycobacterium tuberculosis can take several weeks to yield results,9 rendering this approach unsuitable for urgent diagnostic needs. Although molecular diagnostics, such as polymerase chain reaction (PCR), offer increased sensitivity, their specificity requires nuanced consideration. Analytical specificity—the ability to correctly identify the target sequence without cross-reactivity—is generally high. However, clinical specificity—the proportion of truly uninfected individuals correctly identified as negative—can be compromised by factors such as sample contamination, suboptimal primer design, and the detection of colonizing rather than infecting organisms, leading to false positive results.10,11 These limitations collectively impair the capacity to meet clinical demands, highlighting the urgent need for diagnostic platforms that are both rapid and highly accurate.
It is important to distinguish between genotypic detection of AMR, which identifies the presence of specific resistance genes or mutations in a pathogen’s genome, and phenotypic susceptibility testing, which directly measures the ability of a bacterium to grow in the presence of an antibiotic to determine the Minimum Inhibitory Concentration (MIC). While genotypic methods offer significant speed advantages, the presence of a resistance gene does not invariably confer phenotypic resistance due to factors such as gene silencing, insufficient expression levels, or epistatic interactions. Conversely, novel or uncharacterized resistance mechanisms may be missed by genotypic approaches. This distinction is clinically significant, as it underscores the complementary roles of both approaches in comprehensive AMR management.
Beyond the genotypic–phenotypic distinction, successful clinical implementation of any molecular diagnostic, including CRISPR-based systems, must address several practical considerations. Sample preparation remains a critical bottleneck: clinical specimens (eg, blood, sputum, and urine) contain complex matrices with potential inhibitors—such as hemoglobin, immunoglobulins, and mucopolysaccharides—that can interfere with nucleic acid amplification and Cas enzyme activity, leading to false-negative results. Furthermore, contamination from environmental DNA or cross-contamination between samples poses a risk for false-positive outcomes, particularly in assays with high amplification sensitivity. Standardized protocols for nucleic acid extraction, internal controls for inhibition monitoring, and validated contamination-prevention measures are therefore essential prerequisites for reliable point-of-care deployment. Finally, regulatory frameworks require that novel diagnostics undergo rigorous analytical and clinical validation against established reference methods before routine clinical adoption.
CRISPR technology, a groundbreaking gene-editing platform, has exhibited substantial utility across multiple disciplines, with notable potential in the realms of molecular diagnostics.12–14 By leveraging its precise genome-editing capabilities, the CRISPR/Cas system enables innovative methodologies for the detection of drug-resistant bacteria. This review focuses specifically on the detection of drug resistance genes rather than pathogen identification per se, as resistance gene profiling provides the critical information needed to guide appropriate antibiotic selection. We primarily focus on CRISPR diagnostic platforms based on the collateral-cleavage activity of Cas12 and Cas13 effectors and their derivative systems, as these represent the most actively developed architectures for rapid resistance gene detection.
Biological Foundations of the CRISPR-Cas System
The CRISPR-Cas system represents an adaptive immune defense mechanism that has evolved in bacteria and archaea.15,16 Its primary function is to identify and cleave invading bacteriophage or exogenous plasmid DNA. Upon the invasion of foreign DNA, the system incorporates specific fragments of this DNA as spacer sequences into its CRISPR array, thereby establishing an immunological memory. When the host subsequently encounters the same foreign DNA, the pre-crRNA transcribed from the CRISPR array is processed into mature crRNA.17 This crRNA then associates with Cas proteins to form an effector complex. The complex achieves precise recognition through base pairing between the crRNA and the complementary sequence of the target DNA. Upon recognition, the Cas proteins exhibit endonuclease activity, cleaving the target DNA and thus degrading the exogenous genetic material to protect the host cell.
The CRISPR array is characterized by alternating repeat and spacer sequences, with the latter being derived from previously encountered foreign DNA.17 The Cas protein family includes various members, each with distinct functions.18 Class 1 systems utilize a multi-protein effector complex (eg, the Cascade complex), whereas Class 2 systems rely on a single, multi-domain effector protein (such as Cas9, Cas12, or Cas13) to perform both recognition and cleavage. Notably, the Cas9 protein, directed by guide RNA (gRNA), is capable of specifically recognizing and cleaving double-stranded DNA complementary to the gRNA through its dual nuclease domains.19 This recognition mechanism imparts the CRISPR-Cas system with high specificity.
While CRISPR-Cas systems are noted for their high target specificity, it is important to acknowledge that Cas enzymes can tolerate certain base mismatches between the gRNA and target sequence, particularly at positions distal to the PAM-proximal “seed” region (typically the 10–12 nucleotides immediately upstream of the PAM). This mismatch tolerance could potentially lead to off-target binding and false-positive signals in diagnostic applications. In the context of AMR gene detection, where resistance gene families often share significant sequence homology (eg, different variants of β-lactamase genes), careful gRNA design is essential to ensure variant-level discrimination. Strategies to mitigate off-target effects include the use of high-fidelity Cas variants engineered for improved mismatch discrimination, truncated gRNAs (tru-gRNAs), and rigorous bioinformatic validation of gRNA specificity against comprehensive resistance gene databases.
CRISPR-Cas System Depends On Two Key Components
The core of CRISPR diagnostic technology lies in its programmable target recognition capability, which depends on two key components: a guide RNA (gRNA) (the molecular “GPS”) and a Cas nuclease (the molecular “scissors”).12,20
The gRNA is typically a synthetic single-chain molecule that merges the functions of bio-CRISPR RNA (crRNA) and trans-activating crRNA (tracrRNA).21 The design of gRNAs is crucial, necessitating adequate complementarity to the target gene sequence while minimizing off-target interactions. The use of bioinformatics tools facilitates the identification of optimal gRNA sequences.22
While CRISPR technology was initially famed for gene-editing (Cas9), diagnostic applications have shifted toward enzymes best suited for signal generation (Cas12 and Cas13).
Cas9: The Original Gene Editor. Guided by a gRNA, Cas9 cleaves dsDNA to create a double-strand break. While revolutionary for editing, its diagnostic application typically requires downstream steps—such as amplification or electrophoresis—to detect the cleavage event, as it lacks intrinsic signal amplification mechanisms.23
Cas12 (eg, Cas12a and Cas12b): This enzyme recognizes a specific dsDNA target. Upon binding, it cuts the target and becomes activated to indiscriminately cleave any nearby single-stranded DNA (ssDNA) molecules.14
Cas13 (eg, Cas13a): This enzyme targets ssRNA. Upon binding, it is activated to indiscriminately cleave surrounding ssRNA molecules.24,25 This makes it ideal for detecting RNA viruses or monitoring bacterial gene expression. While the concept of simultaneous DNA/RNA detection using orthogonal Cas12 and Cas13 systems has been demonstrated in proof-of-concept studies, clinical validation of such dual-detection approaches for AMR applications remains limited and represents an active area of development.
CRISPR-Cas Based Bacterial Drug Resistance Gene Detection
The technical approach for CRISPR-Cas-mediated detection encompasses target design, gRNA synthesis, RNP assembly, and detection. The gRNA directs the Cas protein to a precise target sequence. For DNA-targeting enzymes like Cas9 and Cas12, recognition requires a Protospacer Adjacent Motif (PAM).
The requirement for a PAM sequence adjacent to the target site imposes practical constraints on the range of targetable sequences within resistance genes. For the widely used SpCas9, the 5′-NGG-3′ PAM requirement limits approximately one-third of all possible target sites. This constraint can be particularly relevant when attempting to target highly conserved or specific regions of certain resistance genes. To address this limitation, researchers can employ strategics such as utilizing Cas enzymes from different species that recognize alternative PAM sequences (eg, Cas12a recognizes a 5′-TTTN-3′ PAM), using engineered Cas variants with relaxed PAM requirements, or leveraging PAM-independent systems such as Cas13.
By adding reporter molecules (ssDNA for Cas12 and ssRNA for Cas13) labeled with a fluorophore and a quencher, the system is primed for signal amplification. The activation of a single Cas enzyme by a trace amount of target triggers a cascade, acting like “frenzied scissors” to cleave thousands of reporter molecules, releasing a detectable fluorescent signal.
Methodological Framework for Detection of Resistance Genes
The complete CRISPR-Cas diagnostic workflow for AMR gene detection, from sample collection through signal readout, is illustrated in Figure 1. The pipeline integrates isothermal amplification with CRISPR-based recognition and collateral cleavage, enabling rapid and sensitive identification of resistance genes without the need for complex instrumentation.
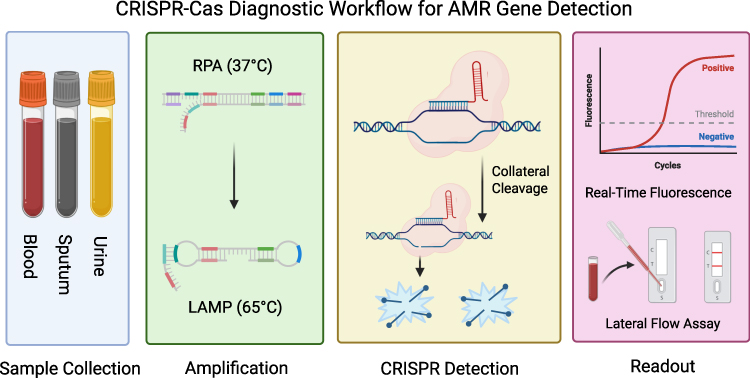 |
Figure 1 CRISPR-Cas Diagnostic Workflow for AMR Gene Detection. This figure provides an overview of the complete CRISPR-Cas diagnostic pipeline. The workflow proceeds through five sequential stages: (1) Sample Collection and nucleic acid extraction from clinical specimens; (2) Isothermal Amplification via RPA (37°C) or LAMP (60–65°C); (3) CRISPR Recognition and Collateral Cleavage, where the Cas-gRNA complex binds the target and activates non-specific reporter cleavage; (4) Signal Generation through fluorescence or lateral flow assay readout. |
Mainstream Diagnostic Platforms
SHERLOCK and DETECTR are two landmark CRISPR diagnostic platforms that standardized the collateral cleavage activity of Cas12 and Cas13.26–29
SHERLOCK (Specific High-sensitivity Enzymatic Reporter unLOCKing): Based on RNA-targeting Cas13, involving isothermal pre-amplification, in vitro transcription (if identifying DNA), and Cas13-mediated collateral cleavage of RNA reporters.27,28,30,31
DETECTR (DNA Endonuclease-Targeted CRISPR Trans Reporter): Based on DNA-targeting Cas12, involving isothermal amplification and direct Cas12-mediated cleavage of ssDNA reporters.26,29,32
Regarding commercialization and regulatory progress, Sherlock Biosciences obtained the first FDA Emergency Use Authorization (EUA) for a CRISPR-based diagnostic in May 2020 for SARS-CoV-2 detection. More recently, the company launched its SHERLOCK Select platform in January 2025 for at-home respiratory infection testing. Mammoth Biosciences, developers of DETECTR, announced a strategic collaboration with Illumina in December 2024 to integrate CRISPR diagnostics with NGS workflows. These developments signal a clear trajectory toward broader commercial availability and regulatory acceptance.
Reaction Kinetics and Signal Saturation
The kinetics of Cas-mediated collateral cleavage and the associated reporter signal generation are subject to saturation effects that influence the quantitative capability of CRISPR diagnostics. At high target concentrations, the rate of reporter cleavage can reach a plateau, resulting in a limited dynamic range for quantitative applications. Additionally, reaction kinetics are influenced by factors such as Cas enzyme concentration, gRNA-target binding efficiency, and temperature, which require careful optimization for each specific diagnostic application to ensure maximal sensitivity without compromising specificity.
Synergy with Nucleic Acid Amplification Technologies
Because pathogen concentrations in clinical samples are often low, most workflows integrate nucleic acid amplification.13,14,29,33
Isothermal Amplification
Recombinase Polymerase Amplification (RPA) and Loop-Mediated Isothermal Amplification (LAMP) operate at constant temperatures, eliminating the need for thermal cyclers and facilitating Point-of-Care Testing (POCT).34
PCR and qPCR
Combining CRISPR with PCR allows leveraging existing infrastructure for precise quantitative analysis, though with higher instrument complexity.
CRISPR-NGS for High-Throughput Surveillance
CRISPR-NGS utilizes Cas9 for target enrichment, depleting background DNA to significantly increase the relative abundance of target antimicrobial resistance genes (ARGs) before sequencing. This approach can increase sequencing read depth by 10-fold and identify low-abundance resistance genes missed by conventional metagenomic NGS.35
Visualizing the Diagnostic Result
Fluorescence Detection
The most common laboratory method, using fluorophore-quencher labeled reporters quantified by fluorescence readers.
Lateral Flow Assays (LFA)
Adapted for POCT, using reporters labeled with tags (eg, FAM/Biotin) captured on strips. A positive result prevents capture at the second line (or vice versa depending on design), providing a visual “yes/no” readout.
However, conventional lateral flow assays are inherently qualitative or semi-quantitative. Key quantitative limitations include the following: (1) a narrow dynamic range compared to fluorescence methods; (2) subjective visual interpretation of band intensity; (3) susceptibility to the “hook effect” at very high analyte concentrations; and (4) the absence of internal calibration standards. To enhance capabilities, researchers are developing smartphone-based image analysis for objective quantification and integrating fluorescent labels.
Clinical Applications in Detection of Drug-Resistant Pathogens
Detection of Methicillin-Resistant Staphylococcus Aureus (MRSA)
Rapid identification of MRSA is crucial for infection control.36 CRISPR technology primarily targets the mecA gene.32
- RAA-Cas12a: Combines RAA with Cas12a, completing detection in 60 minutes with an LoD of 10 copies/µL and high specificity.32
- RPA-Cas12a/Cas13a Single-Tube: Integrates amplification and detection to reduce contamination risk.37
- MCFHCR: Combines magnetic bead enrichment, Cas12a, and HCR signal amplification, achieving five copies/µL sensitivity in 35 minutes.32
Detection of Carbapenem-Resistant Enterobacteriaceae (CRE)
Identification of carbapenemase genes (blaKPC, blaNDM, and blaOXA-48) is critical for CRE management.38–40
- RPA-CRISPR/Cas12a (RCCS): Detects blaKPC and blaNDM in 50 minutes with attomolar sensitivity.40
- LAMP-CRISPR-Cas13a: Detects blaOXA-48 and blaGES in 2 hours without purification, achieving 100% sensitivity/specificity.30
Other Applications in AMR Surveillance
Multiplexing Capability
Detecting multiple resistance genes simultaneously is crucial. Strategies include using orthogonal Cas enzymes (eg, Cas12a/Cas13a) with distinct reporters.31 However, multiplexed CRISPR detection faces significant challenges: potential cross-reactivity between Cas-gRNA complexes, the need for non-interfering reporter systems, complex reaction optimization, and the risk of reduced sensitivity when multiple reactions compete for shared resources.
Environmental Monitoring
CRISPR-NGS effectively tracks ARG profiles in wastewater for real-time surveillance.35
Comparative Performance Analysis with Traditional Diagnostic Methods
Specificity
CRISPR-Cas minimizes non-specific binding through precise base-pairing, surpassing PCR in specificity for single-target detection.41,42
Sensitivity
Comparable to optimized PCR, capable of detecting <50 copies without complex instrumentation.14
Operational Advantages
Rapid turnaround (hours) compared to days for culture.14
Necessity of Phenotypic Confirmation
It is essential to emphasize that genotypic CRISPR-based detection does not replace the need for phenotypic confirmation. The presence of a resistance gene does not invariably confer phenotypic resistance. Therefore, CRISPR diagnostics should be viewed as a complementary screening tool guiding initial decisions, while phenotypic testing remains necessary for definitive treatment guidance (as shown in Table 1).
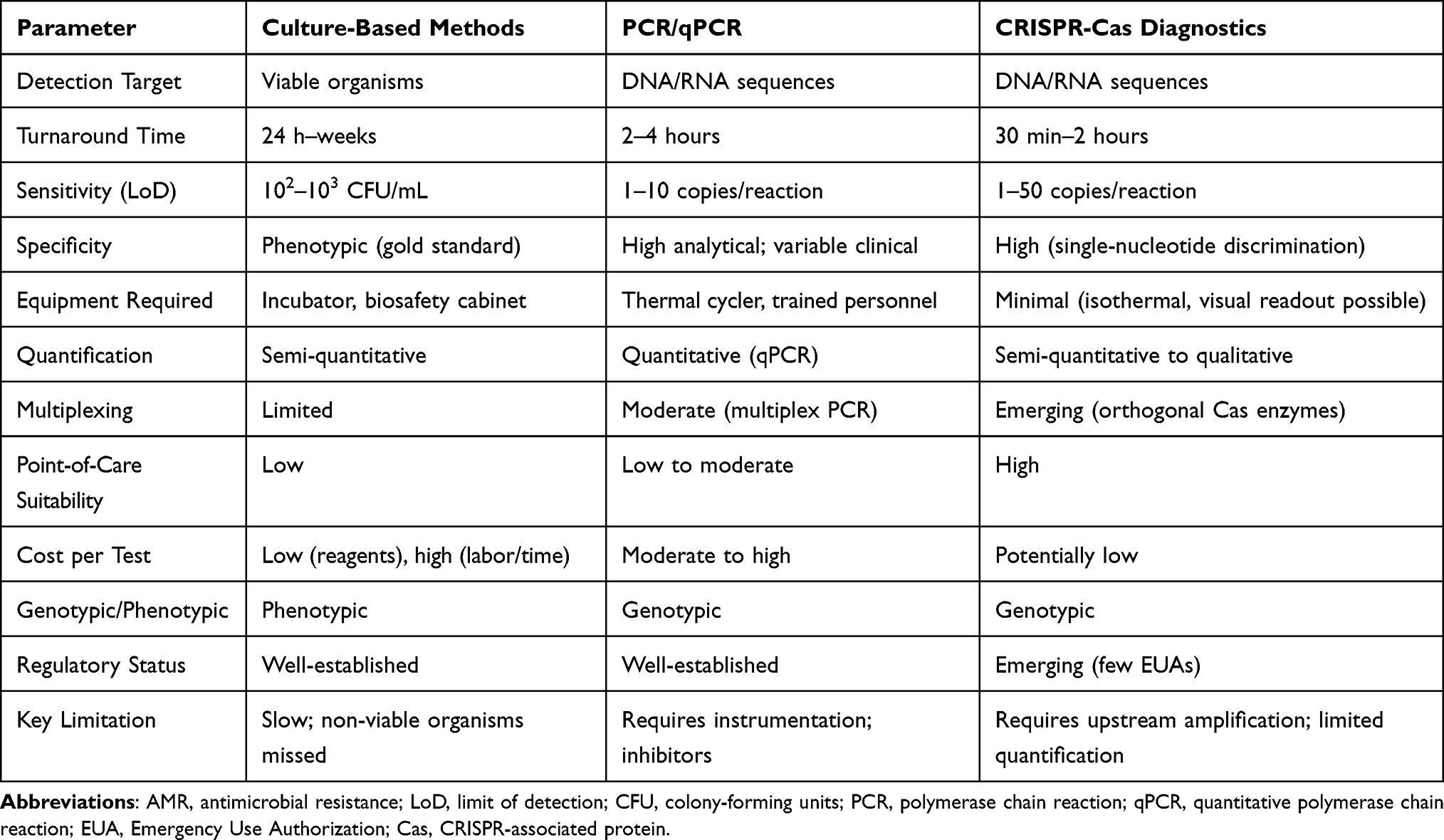 |
Table 1 Comparison of CRISPR-Cas Diagnostics with Traditional Methods for AMR Detection |
Future Directions of CRISPR-Cas-Based Detection
The future of CRISPR diagnostics lies in integration with microfluidics, nanotechnology, and artificial intelligence.
Artificial Intelligence (AI)
AI is becoming a pivotal tool. Intelligent Assay Design: Machine learning algorithms rapidly predict optimal gRNA target sites by analyzing massive genomic datasets, assessing off-target risks more efficiently than manual design.43
AI-Driven Phenotypic Prediction
Recent advances (2024–2025) include deep learning models capable of diagnosing drug-resistant infections directly from microscopy images and analyzing whole-genome sequences to identify resistance mutations with high accuracy.
Data Bias
However, AI-assisted design faces limitations, including potential biases in training datasets that may underrepresent rare resistance genes and the risk of overfitting, necessitating validation across diverse clinical settings.
Microfluidics
“Lab-on-a-Chip” platforms integrate sample prep, amplification, and detection into disposable cartridges, enabling true “sample-in, answer-out” POCT functionality and reducing contamination risks.44
Personalized Therapy
Rapid resistance profiling allows for the customization of antibiotic therapies, moving away from broad-spectrum approaches and supporting antimicrobial stewardship.
Ethical and Technical Challenges
Ethical Considerations in Technological Applications
The implementation of CRISPR-Cas technology for the detection of resistance genes raises substantial concerns regarding genetic privacy. Data pertaining to individual resistance genes is classified as sensitive health information, and its unauthorized disclosure could result in employment discrimination or exclusion from insurance coverage.
Regulatory and Standardization Deficiencies
Present CRISPR-Cas detection systems suffer from a lack of cohesive regulatory oversight and standardization. Variations in detection methodologies, reagents, and protocols across different laboratories undermine the accuracy and comparability of results. For instance, the generation of conflicting outcomes by different laboratories analyzing identical samples leads to diagnostic and therapeutic uncertainties.45
Conclusion
The intensifying global crisis of antimicrobial resistance necessitates innovative diagnostic solutions. CRISPR-Cas systems represent a significant advancement in the detection of antimicrobial resistance genes, offering high specificity, sensitivity, and speed. The integration of CRISPR-Cas with isothermal amplification, lateral flow assays, microfluidics, and AI-driven design holds significant potential for point-of-care application. However, successful translation requires addressing key challenges: standardization of protocols, mitigation of off-target effects, validation of AI models, and regulatory alignment. Furthermore, while CRISPR offers powerful genotypic screening, it must effectively complement phenotypic testing to provide comprehensive guidance for antimicrobial stewardship.
Acknowledgments
Figure 1 was created in BioRender. (https://BioRender.com/dnt3wif) is licensed under CC BY 4.0.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Levy SB, Marshall B. Antibacterial resistance worldwide: causes, challenges and responses. Nat Med. 2004;10:S122–9. doi:10.1038/nm1145
2. World Health Organization. Global Antibiotic Resistance Surveillance Report 2025. Geneva: World Health Organization; 2025.
3. Zhao K, Tu C, Chen W, et al. Rapid identification of drug-resistant tuberculosis genes using direct PCR amplification and Oxford nanopore technology sequencing. Can J Infect Dis Med Microbiol. 2022;2022:1–8. doi:10.1155/2022/7588033
4. Bianco G, Boattini M, Iannaccone M, et al. Detection of antibiotic resistance genes from blood cultures: performance assessment and potential impact on antibiotic therapy management. J Hosp Infect. 2019;102:465–469. doi:10.1016/j.jhin.2019.03.007
5. Ma X, Cui J, Wang J, et al. Multicentre investigation of pathogenic bacteria and antibiotic resistance genes in Chinese patients with acute exacerbation of chronic obstructive pulmonary disease. J Int Med Res. 2015;43:699–710. doi:10.1177/0300060515587577
6. Bloomer E, McKee M. Policy options for reducing antibiotics and antibiotic-resistant genes in the environment. J Public Health Policy. 2018;39:389–406. doi:10.1057/s41271-018-0144-x
7. Voigt AM, Ciorba P, Döhla M, et al. The investigation of antibiotic residues, antibiotic resistance genes and antibiotic-resistant organisms in a drinking water reservoir system in Germany. Int J Hyg Environ Health. 2020;224:113449. doi:10.1016/j.ijheh.2020.113449
8. Leoni E, Legnani PP. Comparison of selective procedures for isolation and enumeration of legionella species from hot water systems. J Appl Microbiol. 2001;90:27–33. doi:10.1046/j.1365-2672.2001.01178.x
9. Wang R, Zhao X, Wan K. Deterioration of cycloserine in drug susceptibility testing of mycobacterium. Infect Drug Resist. 2022;15:135–140. doi:10.2147/IDR.S348428
10. Alanio A, Bretagne S. Difficulties with molecular diagnostic tests for mould and yeast infections: where do we stand? Clin Microbiol Infect. 2014;20:36–41.
11. Erler S, Lommatzsch S, Lattorff HMG. Comparative analysis of detection limits and specificity of molecular diagnostic markers for three pathogens (Microsporidia, Nosema spp.) in the key pollinators Apis mellifera and Bombus terrestris. Parasitol Res. 2012;110:1403–1410. doi:10.1007/s00436-011-2640-9
12. Yang H, Zhang Y, Teng X, et al. CRISPR-based nucleic acid diagnostics for pathogens. TRAC-Trends Anal Chem. 2023;160:116980. doi:10.1016/j.trac.2023.116980
13. Wang T, Bai L, Wang G, et al. SATCAS: a CRISPR/Cas13a-based simultaneous amplification and testing platform for one-pot RNA detection and SNPs distinguish in clinical diagnosis. Biosens Bioelectron. 2024;263:116636. doi:10.1016/j.bios.2024.116636
14. Kostyusheva A, Brezgin S, Babin Y, et al. CRISPR-cas systems for diagnosing infectious diseases. Methods. 2022;203:431–446. doi:10.1016/j.ymeth.2021.04.007
15. Ishino Y, Shinagawa H, Makino K, Amemura M, Nakata A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in escherichia coli, and identification of the gene product. J Bacteriol. 1987;169:5429–5433. doi:10.1128/jb.169.12.5429-5433.1987
16. Zhang F. Development of CRISPR-cas systems for genome editing and beyond. Q Rev Biophys. 2019;52:e6. doi:10.1017/S0033583519000052
17. Zhang X, Huang Z, Zhang Y, et al. Mitigating antibiotic resistance: the utilization of CRISPR technology in detection. Biosensors. 2024;14:633. doi:10.3390/bios14120633
18. Qian Y, Zhou D, Li M, et al. Application of CRISPR-cas system in the diagnosis and therapy of ESKAPE infections. Front Cell Infect Microbiol. 2023;13:1223696. doi:10.3389/fcimb.2023.1223696
19. Tao S, Chen H, Li N, Liang W. The application of the CRISPR-cas system in antibiotic resistance. Infect Drug Resist. 2022;15:4155–4168. doi:10.2147/IDR.S370869
20. Sidik SM, Huet D, Lourido S. CRISPR-Cas9-based genome-wide screening of Toxoplasma gondii. Nat Protoc. 2018;13:307–323. doi:10.1038/nprot.2017.131
21. Mengstie MA, Wondimu BZ. Mechanism and applications of CRISPR/cas-9-mediated genome editing. Biol Targets Ther. 2021;15:353–361. doi:10.2147/BTT.S326422
22. Huang Z, Nair M. A CRISPR/Cas9 guidance RNA screen platform for HIV provirus disruption and HIV/AIDS gene therapy in astrocytes. Sci Rep. 2017;7:5955. doi:10.1038/s41598-017-06269-x
23. Kundar R, Gokarn K. CRISPR-cas system: a tool to eliminate drug-resistant gram-negative bacteria. Pharmaceuticals. 2022;15:1498. doi:10.3390/ph15121498
24. Han Y, Yan T, Zhang C, Zhang J. CRISPR technology in infectious disease management: principles, applications, and future challenges with emphasis on SARS-CoV-2, HIV, and HBV. Highlights Sci Eng Technol. 2023;74:533–540. doi:10.54097/09vggj25
25. Myhrvold C, Freije CA, Gootenberg JS, et al. Field-deployable viral diagnostics using CRISPR-Cas13. Science. 2018;360:444–448. doi:10.1126/science.aas8836
26. Chen JS, Ma E, Harrington LB, et al. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science. 2018;360:436–439. doi:10.1126/science.aar6245
27. Kellner MJ, Koob JG, Gootenberg JS, Abudayyeh OO, Zhang F. SHERLOCK: nucleic acid detection with CRISPR nucleases. Nat Protoc. 2019;14:2986–3012. doi:10.1038/s41596-019-0210-2
28. Mann JG, Pitts RJ. PrimedSherlock: a tool for rapid design of highly specific CRISPR-Cas12 crRNAs. BMC Bioinf. 2022;23:428. doi:10.1186/s12859-022-04968-5
29. Broughton JP, Deng X, Yu G, et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat Biotechnol. 2020;38:870–874. doi:10.1038/s41587-020-0513-4
30. Ortiz-Cartagena C, Pablo-Marcos D, Fernández-García L, et al. CRISPR-Cas13a-based assay for accurate detection of OXA-48 and GES carbapenemases. Microbiol Spectr. 2023;11:e01329–23. doi:10.1128/spectrum.01329-23
31. Lv H, Wang J, Zhang J, et al. Definition of CRISPR Cas12a trans-cleavage units to facilitate CRISPR diagnostics. Front Microbiol. 2021;12:766464. doi:10.3389/fmicb.2021.766464
32. Jiang Y, Chen Z, Liu X, et al. A rapid and naked-eye methicillin resistant Staphylococcus aureus screening method based on CRISPR/Cas12a and hybridization chain reaction. Front Microbiol. 2025;16:1592153. doi:10.3389/fmicb.2025.1592153
33. Zhou Z, Cho I-H, Kadam US. CRISPR-Cas-based diagnostics in biomedicine: principles, applications, and future trajectories. Biosensors. 2025;15:660. doi:10.3390/bios15100660
34. Li L, Li S, Wu N, et al. HOLMESv2: a CRISPR-Cas12b-assisted platform for nucleic acid detection and DNA methylation quantitation. ACS Synth Biol. 2019;8:2228–2237. doi:10.1021/acssynbio.9b00209
35. Mao Y, Shisler JL, Nguyen TH. Enhanced detection for antibiotic resistance genes in wastewater samples using a CRISPR-enriched metagenomic method. Water Res. 2025;274:123056. doi:10.1016/j.watres.2024.123056
36. Lakhundi S, Zhang K. Methicillin-resistant staphylococcus aureus: molecular characterization, evolution, and epidemiology. Clin Microbiol Rev. 2018;31:e00020–18. doi:10.1128/CMR.00020-18
37. Chen W, Zhang J, Wei H, et al. Rapid and sensitive detection of methicillin-resistant staphylococcus aureus through the RPA-PfAgo system. Front Microbiol. 2024;15:1422574. doi:10.3389/fmicb.2024.1422574
38. Naghavi M, Vollset SE, Ikuta KS, et al. Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. Lancet. 2024;404:1199–1226. doi:10.1016/S0140-6736(24)01867-1
39. Gao Q, Zhang T, Yuan Y, et al. Detection of KPC-producing carbapenem-resistant Klebsiella pneumoniae based on CRISPR Cas12a. J Microbiol Biotechnol. 2025;35:e2502042. doi:10.4014/jmb.2502.02042
40. Yang JW, Kim H, Hyeon L-S, Yoo JS, Kang S. Development of a recombinase polymerase amplification-coupled CRISPR/Cas12a platform for rapid detection of antimicrobial-resistant genes in carbapenem-resistant enterobacterales. Biosensors. 2024;14:536. doi:10.3390/bios14110536
41. Bhattacharyya RP, Thakku SG, Hung DT. Harnessing CRISPR effectors for infectious disease diagnostics. ACS Infect Dis. 2018;4:1278–1282. doi:10.1021/acsinfecdis.8b00170
42. Uddin F, Rudin CM, Sen T. CRISPR gene therapy: applications, limitations, and implications for the future. Front Oncol. 2020;10:1387. doi:10.3389/fonc.2020.01387
43. Chen H, Zhou X, Wang M, Ren L. Towards point of care CRISPR-based diagnostics: from method to device. J Funct Biomater. 2023;14:97. doi:10.3390/jfb14020097
44. Chen B, Li Y, Xu F, Yang X. Powerful CRISPR-based biosensing techniques and their integration with microfluidic platforms. Front Bioeng Biotechnol. 2022;10:851712. doi:10.3389/fbioe.2022.851712
45. Van Dongen JE, Segerink LI. Building the future of clinical diagnostics: an analysis of potential benefits and current barriers in CRISPR/cas diagnostics. ACS Synth Biol. 2025;14:323–331. doi:10.1021/acssynbio.4c00816
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
CRISPR-Cas Systems in the Fight Against Antimicrobial Resistance: Current Status, Potentials, and Future Directions
Ahmed MM, Kayode HH, Okesanya OJ, Ukoaka BM, Eshun G, Mourid MR, Adigun OA, Ogaya JB, Mohamed ZO, Lucero-Prisno III DE
Infection and Drug Resistance 2024, 17:5229-5245
Published Date: 26 November 2024